Фёршт Э. Структура и механизм действия ферментов
Подождите немного. Документ загружается.


ОСНОВНЫЕ УРАВНЕНИЯ ФЕРМЕНТАТИВНОЙ КИНЕТИКИ Ц1
Концентрация
субстрата, при которой v =-^
V
max
,
обозна-
чается /См и называется константой
Михаэлиса.
Обратите вни-
мание,
что при низких значениях [S], когда [S] -С Км,
S].
(3.3)
2. Интерпретация кинетических
параметров
для
односубстратных
реакций:
механизм
Михаэлиса — Ментен
В 1913 г. Михаэлис и Ментен, развив идеи
других
исследо-
вателей, предложили
следующую
схему:
*S *cat
E + S
=Р=*
ES -—»• ЕР. (3.4)
Каталитическая реакция состоит из
двух
стадий. Сначала
фермент и
субстрат
соединяются и образуют фермент-субстрат-
ный
комплекс ES. Предполагается, что эта стадия протекает
быстро и обратимо и не сопровождается какими-либо химиче-
скими
изменениями; фермент и
субстрат
удерживаются вместе
за счет сил физической природы. На второй стадии протекают
химические процессы, характеризующиеся константой скорости
первого порядка
&
ca
t
(число оборотов).
Уравнение скорости выводится следующим образом. Из
уравнения (3.4) получаем
(3.5)
(3.6)
Далее, поскольку суммарная концентрация фермента [Е]
о
и
концентрация
свободного фермента [Е] связаны соотношением
[E] =
[E]
0
-[ES],
(3.7)
имеем
1
Е8
1-Т??Уп!г
(3-8)
(3.9)
Это уравнение идентично уравнению (3.1), где Кгл равна
константе диссоциации фермент-субстратного комплекса.

112
ГЛАВА
3
Концепция
фермент-субстратного комплекса является краеу-
гольным камнем ферментативной кинетики и лежит в основе
наших представлений о механизмах ферментативного катализа.
В честь ученого, который ввел это понятие, комплекс субстрата с
ферментом,
образованный без участия ковалентных связей, ча-
сто называют
комплексом
Михаэлиса.
3.
Последующие
модификации
механизма
Михаэлиса
— Ментен
Не
следует
отождествлять уравнение и механизм, предло-
женные Михаэлисом и Ментен. Уравнение выполняется для
многих механизмов, механизм же не всегда согласуется с полу-
ченными
данными.
Механизм Михаэлиса — Ментен предполагает, что фермент-
субстратный комплекс находится в термодинамическом равно-
весии
со свободными ферментом и субстратом. Это справедливо
только в том случае, если в уравнении
(3.10)
ki «С k-\\
E+S
=f=fc
ES >- EP.
(3.10)
Случай, когда k
2
сравнима с k-\, впервые был рассмотрен
Бриггсом и Холдейном в 1925 г.
а.
Кинетика
Бриггса
—
Холдейна
[2]: Км > Ks
Вывести уравнение скорости для схемы
(3.10)
немного
труд-
нее,
чем для механизма Михаэлиса — Ментен. При расчете кон-
центрации
ES используется принцип стационарности, т. е. счи-
тается, что если измеряемая скорость реакции приблизительно
постоянна
в рассматриваемом промежутке времени, то [ES]
также постоянна:
1
0 = fti [E] [S] - *, [ES] _ *_, [ES] (3.11)
Из
уравнений (3.7) и
(3.11)
получаем
и,
поскольку v = k
2
[ES],
[E]
0
[S]*,
(3.13)
Это уравнение идентично уравнению Михаэлиса — Ментен (3.1);
здесь
(*
2
+
*_,)/*,
= *«•
(3.14)

ОСНОВНЫЕ
УРАВНЕНИЯ
ФЕРМЕНТАТИВНОЙ
КИНЕТИКИ
ИЗ
Поскольку
константа диссоциации комплекса ES равна /Cs =
=
k-ilk
h
ТО
(3.15)
Естественно, что при k-i^ k
2
уравнение
(3.14)
сводится к виду
Км = Ks, т. е. константа Михаэлиса представляет собой в этом
случае
константу диссоциации фермент-субстратного комплек-
са.
б. Промежуточное соединение, образующееся вслед за
образованием ES: Км < Ks
Схему
Михаэлиса — Ментен можно использовать для анали-
за самых разных процессов, отличительной особенностью кото-
рых является образование в
ходе
реакции дополнительных ко-
валентно или нековалентно связанных промежуточных соедине-
ний.
Установлено, что во
всех
этих
случаях
применимо уравнение
Михаэлиса — Ментен, хотя теперь Км и k
ca
t представляют собой
комбинации
различных констант скорости и констант равнове-
сия.
При этом
всегда
Км ^ Ks. Допустим, например, что реак-
ция
протекает с образованием ряда промежуточных соединений
[схема
(3.16)]
и конечная каталитическая стадия является мед-
ленной:
KS
* К'
k
* /г, ,^\
E+S
^=? ES ч=* ES' <=* ES" -> ЕР, (3.16)
Медленно
где [ES']
=tf[ES],
a [ES"] = /C[ES].
Следовательно,
K
K
=
K
S
/(1
+ K + KK')
(3.17)
*
c
,t
=
*
4
ffff
7(1+ * +
**').
(3.18)
Катализируемый химотрипсином гидролиз эфиров и амидов
протекает согласно
следующему
механизму:
Е+
S
=<=^=
ES ~^ ЕА -^*
ЕР
2>
(3.19)
Pi
где ЕА — «ацилфермент».
Считая,
что концентрация комплекса ЕА стационарна, мы
можем записать
{№}
(3
'
20)

114
ГЛАВА
3
г. е. получаем уравнение Михаэлиса — Ментен, где
*м =
*зУ(*2
+ *з)
(3.21)
*cat = W(*2 +
*
3
).
(3.22)
или
l/*
cat
= l/k
2
+ l/k
3
.
(3.23)
В более сложных случаях неравенство /См < /Cs может не вы-
полняться.
4. Все три механизма встречаются на практике
В
случае
механизма Бриггса — Холдейна, для которого
k%
S> k-\, отношение
kcat/Км
равно k\ — константе скорости свя-
зывания
фермента и субстрата. В гл. 4
будет
показано, что
константы скорости связывания должны быть порядка
10
8
М^-С"
1
. Это позволяет сделать вывод, что для механизма
Бриггса — Холдейна отношение
kat/Км
равно ~10
7
—
10
8
М-
1
-с"
1
. Каталаза, ацетилхолинэстераза, карбоангидраза,
кротоназа, фумараза и триозофосфатизомераза — все эти фер-
менты по указанному критерию подчиняются кинетике Бриг-
гса— Холдейна, о чем свидетельствуют данные табл. 4.4. Еще
одним ферментом такого типа является пероксидаза, выделен-
ная
из хрена,— один из первых ферментов, к которому были
применены методы исследования быстрых реакций [3]. Сначала
пероксидаза образует с перекисью водорода компекс Михаэли-
са, который затем взаимодействует с донором водорода (реак-
ция
второго порядка). При достаточно высоких концентрациях
донора скорость второй реакции значительно превышает ско-
рость диссоциации комплекса Михаэлиса.
Образование вслед за фермент-субстратным комплексом
промежуточных соединений [как в
схеме
(3.19)
] —чрезвычайно
широко распространенное явление. Однако в
случае
физиологи-
ческих субстратов эти промежуточные соединения часто не на-
капливаются и медленной стадией в последовательности реак-
ций
(3.19)
является стадия с константой скорости k
2
. (Обуслов-
лено это причинами принципиального характера, обсуждающи-
мися
в гл. 10, где приведены соответствующие примеры.) При
этих условиях /См равна /Cs (константе диссоциации) и первона-
чально предложенный механизм Михаэлиса — Ментен оказыва-
ется всегда выполнимым. Для многих экспериментальных систем
наблюдается обратная картина. Исследователи часто использу-
ют в опытах синтетические высокореакционные субстраты; в этих
условиях часто происходит накопление содержащих ковалент-
ные связи промежуточных соединений.

ОСНОВНЫЕ
УРАВНЕНИЯ ФЕРМЕНТАТИВНОЙ КИНЕТИКИ 116
Б.
Физический
смысл
параметров
уравнения
Михаэлиса — Ментен
1.
Смысл константы
k
cat
Когда имеется только один фермент-субстратный комплекс и
все стадии связывания достаточно быстрые, параметр k
ca
t в
уравнении Михаэлиса — Ментен представляет собой просто
константу скорости первого порядка для химического превра-
щения
комплекса ES в комплексе ЕР. В более сложных ситуа-
циях
fecat является функцией
всех
констант скорости первого
порядка
и не может быть отнесена ни к какому конкретному
процессу, за исключением крайних случаев. Например, для ме-
ханизма Бриггса — Холдейна, когда комплекс ЕР диссоциирует
достаточно быстро, k
ca
t = k
2
[уравнение (3.10)]. Однако если
диссоциация
комплекса ЕР протекает медленно, то константа
скорости этого процесса вносит вклад в параметр k
ca
t и в
предельном случае, когда скорость диссоциации ЕР значительно
меньше скоростей химических стадий, параметр k
ca
t
будет
ра-
вен
константе скорости диссоциации. Из рассмотрения схемы
(3.19)
видно, что в этом
случае
k
ca
t является функцией констант
k
2
и k
3
. Однако если одна из этих констант много меньше другой,
k
cat
становится равной меньшей константе. Например, если
къ < h, то, согласно (3.22), A>
ca
t = k
3
. Из этого
следует,
что k
ca
t
не
может превышать ни одной из констант скорости первого
порядка
для реакционного пути в прямом направлении и пред-
ставляет собой, таким образом,
нижний
предел для констант
скорости химических реакций.
Часто параметр k
ca
t называют
числом
оборотов
фермента,
поскольку он определяет максимальное число молекул субстра-
та, превращающихся в продукт одним активным центром в
единицу времени.
2. Смысл /С
м
: истинные и кажущиеся константы
равновесия
Несмотря
на то что Км равна /Cs — истинной константе дис-
социации
фермент-субстратного комплекса — только для про-
стого механизма Михаэлиса — Ментен или в аналогичных слу-
чаях, при решении определенных задач /См можно рассматри-
вать как
кажущуюся
константу
диссоциации.
Например, кон-
центрацию свободного фермента можно рассчитать из соотно-
шения
fE][S] ,„
ол
ч
=
^'
(
'
24)

116
ГЛАВА
3
где X [ES] — сумма концентраций
всех
фермент-содержащих
форм
'.
Концепция
кажущихся
величин очень плодотворна и исполь-
зуется во многих
случаях,
например при определении р/С
а
. До-
вольно часто значения р/(а не отражают микроскопическую
ионизацию
данной группы, а представляют собой комбинацию
соответствующей константы с константами равновесия
между
разными
конформационными состояниями данной молекулы.
Эту
кажущуюся
константу можно определить титрованием как
простую константу рЛ*
а
. При анализе влияния температуры, рН
и
других
факторов на кажущуюся Д*м не
следует
слишком пола-
гаться на подобный упрощенный подход; необходимо помнить,
что константы скорости, функцией которых является кажущаяся
константа /См, также зависят от указанных факторов. То же
самое относится к значениям k
ca
i- В литературе имеются при-
меры, когда наличие изломов на температурной зависимости
&cat объяснялось тем, что фермент претерпевает конформацион-
ные
изменения,
тогда
как в действительности имела место раз-
ная
температурная зависимость индивидуальных констант ско-
рости, входящих в параметр
fe
C
at,
т. е. констант k
2
и k
3
в урав-
нении
(3.22).
То,
что /См является мерой количества фермента, связанного
в
той или иной форме с субстратом, иллюстрируется следую-
щей
схемой (сравните с механизмом действия химотрипсина):
E+S
=?=*
ES
>•
ES' > Е + Р.
(3.25)
Применяя
условие стационарности к комплексу ES', получаем
[ES'] = [ES] kt/k
3
.
(3.26)
Когда
k
2
~^>k3,
[ES'] намного превышает [ES], так что ES' вно-
сит в /См больший вклад, чем ES, и является преобладающей
1
Любопытно, что кинетика Михаэлиса — Ментен может наблюдаться и
в
отсутствие насыщения фермента субстратом. Рассмотрим, например, сле-
дующую
схему:
Е + S —
1
~> Е' + Р,
I»
Здесь взаимодействие активной формы фермента с субстратом представляет
собой реакцию второго порядка; в
результате
образуются продукты и неак-
тивная
форма Е', которая медленно переходит в активную. Этот случай слу-
жит примером кинетики с кажущимся насыщением; при этом fccat = k
2
, a
Км = Да/*1. Если рассматривать Е' как
«связанную»
форму фермента, то
можно применять уравнение (3.24).

ОСНОВНЫЕ
УРАВНЕНИЯ ФЕРМЕНТАТИВНОЙ
КИНЕТИКИ
117
ферментсодержащей формой. Даже не решая написанных урав-
нений,
мы интуитивно
чувствуем,
что /См должна быть меньше
Ks приблизительно в ka/k
2
раз, т. е.
(3.27)
K
3. Смысл параметра
k
cai
/K
Как
было показано ранее [уравнение (3.3)], скорость реак-
ции
при низких концентрациях субстрата определяется уравне-
нием
v =(k
cll
t/KfA) [E]o[S], т. е. отношение
kcat/Км
представ-
ляет собой
кажущуюся
константу скорости второго порядка.
Этот параметр не является истинной микроскопической кон-
стантой, за исключением крайнего случая, когда реакция лими-
тируется столкновением фермента с субстратом.
Ценность
параметра
kcat/Кгл
состоит в том, что он связывает
скорость реакции с концентрацией свободного фермента, а не с
его общей концентрацией. В этом нетрудно убедиться из рас-
смотрения
приведенного выше уравнения (3.3), поскольку при
низкой
концентрации субстрата фермент находится преимуще-
ственно в свободном состоянии ([Е]«[Е]о), и, следовательно,
скорость реакции есть
Ниже
будет
показано Сравнение (3.41)], что этот вывод спра-
ведлив для любой концентрации субстрата. Мы покажем также
[уравнение (3.44)], что параметр
k
ca
t/Kn
определяет специ-
фичность
действия фермента при наличии конкурирующих суб-
стратов.
Отношение fe
ca
t//(M не может превышать ни одну из констант
скорости второго порядка [4] для последовательности реакций,
идущих в прямом направлении, и представляет собой, таким
образом,
нижний
предел для константы скорости при связыва-
нии
фермента с субстратом.
В. Графическое представление данных
Для графического анализа данных и обнаружения отклоне-
ний
от идеальности весьма полезно преобразовать уравнение
Михаэлиса — Ментен в линейную форму. Чаще всего для этого
используют метод Лайнуивера—Бэрка. Взяв величины, обрат-
ные
правой и левой частям уравнения (3.1), и подставив выра-
жение (3.2), мы получим уравнение Лайнуивера—Бэрка [5]:
(3.28)
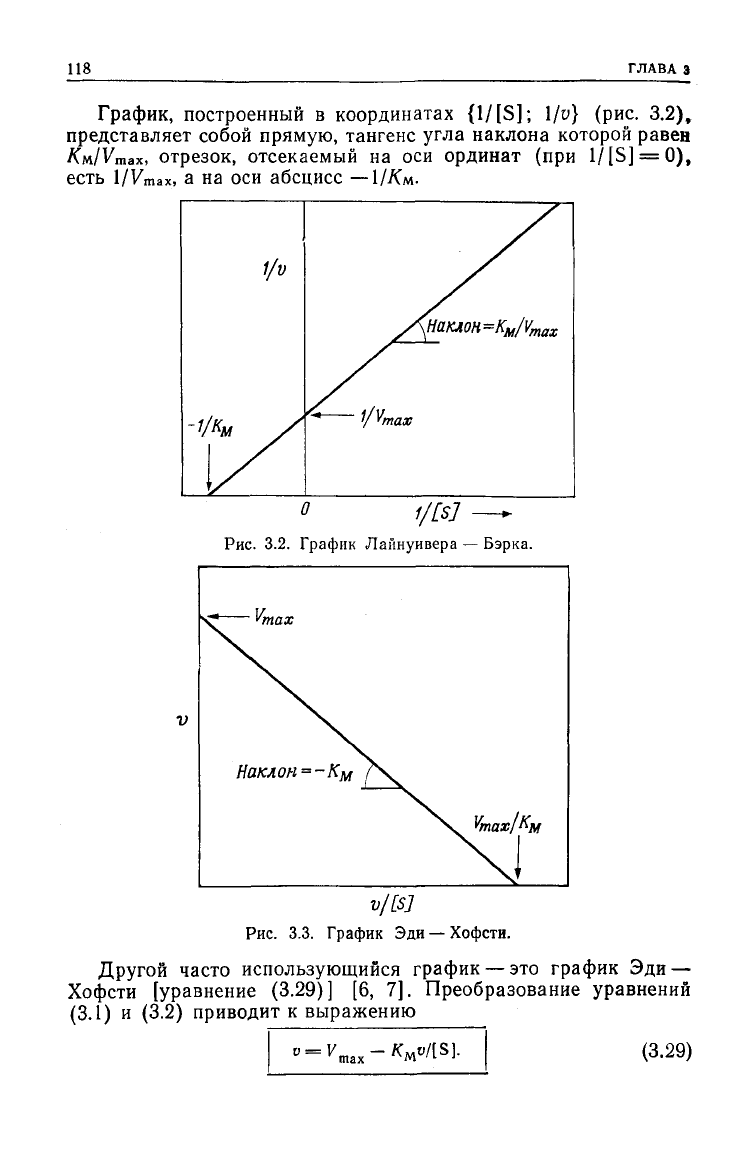
118
ГЛАВА
Э
График,
построенный в координатах
{1/[S];
l/v} (рис. 3.2),
представляет собой прямую, тангенс
угла
наклона которой равен
/См/Vmax,
отрезок, отсекаемый на оси ординат (при 1/[S] = O),
есть
1/Vmax,
а на оси абсцисс —
1//См-
О
f/£
S
J
—+
Рис.
3.2. График Лайнуивера —
Бэрка.
v/[sj
Рис.
3.3. График Эди — Хофсти.
Другой часто использующийся график — это график Эди —
Хофсти [уравнение
(3.29)]
[6, 7]. Преобразование уравнений
(3.1) и (3.2) приводит к выражению
(3.29)

ОСНОВНЫЕ
УРАВНЕНИЯ ФЕРМЕНТАТИВНОЙ
КИНЕТИКИ
Ц9
График,
построенный в координатах
{y/[S];
v) (рис. 3.3),
представляет собой прямую, отсекающую на оси ординат (при
y
/[S] = 0) отрезок, равный Fmax. Тангенс
угла
наклона этой
прямой
равен —К/л, а отрезок, отсекаемый ею на оси абсцисс,
График
Лайнуивера —
Бэрка
имеет тот недостаток, что при
высоких концентрациях
субтрата
экспериментальные точки по-
падают на небольшой отрезок, в
результате
чего точкам, со-
ответствующим низким концентрациям, придается слишком
большой вес. Достоинство же этого графика состоит в том, что
он
позволяет просто оценить значение v при данной концентра-
ции
субстрата [S].
График
Эди — Хофсти лишен указанного выше недостатка,
однако
определить с его помощью значение скорости труднее.
Считается, что этот график
дает
более точные результаты, по-
этому для определения параметров Кгл и
V
ma
x
исследователи
предпочитают пользоваться именно им [8, 9].
Г. Ингибирование
Помимо
необратимой инактивации ферментов при нагрева-
нии
или действии химических реагентов, может происходить
обратимое ингибирование в
ходе
нековалентного связывания
ингибиторов.
Различают четыре основных типа ингибирования.
1. Конкурентное ингибирование
Если
ингибитор I связывается с активным центром фермента
и
препятствует связыванию субстрата S и наоборот, т. е. I и S
конкурируют за связывание с активным центром, то говорят, что
I
является
конкурентным
ингибитором. Для простого механизма
Михаэлиса — Ментен [уравнение (3.4),
KM.
= KS]
следует
рас-
сматривать еще одну стадию:
Е =г=^ ES —^-»» Е + Р.
I.
(3.30)
EI
Используя при выводе уравнения скорости соотношение
[E]
0
= [ES] + [EI] + [E],
(3.31)
получаем
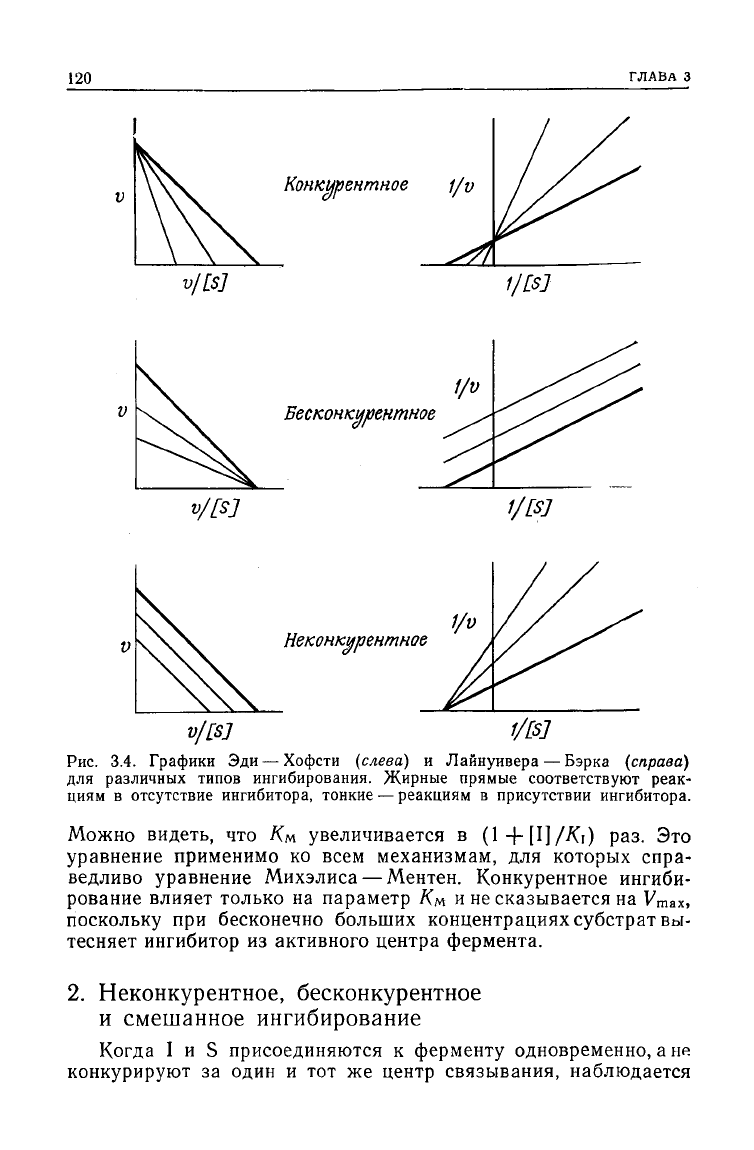
120
ГЛАВА
3
Конкурентное
1/v
v/[S]
f/[Sj
Бесконкурентное
v/[s]
Неконкурентное
v/[sj
Рис.
3.4. Графики Эди — Хофсти
(слева)
и Лайнуивера — Бэрка
(справа)
для различных типов ингибирования. Жирные прямые
соответствуют
реак-
циям
в
отсутствие
ингибитора, тонкие — реакциям в присутствии ингибитора.
Можно видеть, что Км увеличивается в (1 -f-
[Ц/Ki)
раз. Это
уравнение применимо ко всем механизмам, для которых спра-
ведливо уравнение Михэлиса—Ментен. Конкурентное ингиби-
рование влияет только на параметр Км и не сказывается на
У
т
ах,
поскольку при бесконечно больших концентрациях
субстрат
вы-
тесняет ингибитор из активного центра фермента.
2. Неконкурентное, бесконкурентное
и
смешанное ингибирование
Когда I и S присоединяются к ферменту одновременно, а не
конкурируют за один и тот же центр связывания, наблюдается