Zelditch M.L. (и др.) Geometric Morphometrics for Biologists: a primer
Подождите немного. Документ загружается.


chap-06 4/6/2004 17: 23 page 148
148 GEOMETRIC MORPHOMETRICS FOR BIOLOGISTS
at the ith landmark is denoted (X
i
, Y
i
):
V =
X
1
Y
1
X
2
Y
2
.
.
.
.
.
.
X
K
Y
K
00
00
00
=
f
X
(
X
1
, Y
1
)
f
Y
(
X
1
, Y
1
)
f
X
(
X
2
, Y
2
)
f
Y
(
X
2
, Y
2
)
.
.
.
.
.
.
f
X
(
X
K
, Y
K
)
f
Y
(
X
K
, Y
K
)
00
00
00
= LW (6.46)
where LW is the product of two matrices L and W. L is the (K +3) ×(K +3) matrix:
L =
U
(
0
)
U
R
1, 2
U
R
1, 3
··· U
R
1, K
1 X
1
Y
1
U
R
2, 1
U
(
0
)
U
R
2, 3
··· U
R
2, K
1 X
2
Y
2
U
R
3, 1
U
R
3, 2
U
(
0
)
··· U
R
3, K
1 X
3
Y
3
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
U
R
K,1
U
R
K,2
U
R
K,3
··· U
R
K, K
1 X
K
Y
K
111··· 1000
X
1
X
2
X
3
··· X
K
00 0
Y
1
Y
2
Y
3
··· Y
K
00 0
(6.47)
in which U(R) is the function appearing in Equations 6.44 and 6.45 evaluated at each
landmark location (X
i
, Y
i
). W is the (K +3) ×2 matrix of weights and uniform terms
appearing in Equations 6.44 and 6.45:
W =
W
X1
W
Y1
W
X2
W
Y2
.
.
.
.
.
.
W
XK
W
YK
A
X1
A
Y1
A
XX
A
YX
A
XY
A
YY
(6.48)
So we have the equation:
V = LW (6.49)
in which L and W are the matrices just described. We wish to solve for W, the matrix of
coefficients in our spline model, which gives us:
W = L
−1
V (6.50)
We can use the weights in the matrix W in conjunction with the spline functions in Equa-
tions 6.44 and 6.45 to interpolate the observed deformation at the landmarks over the
entire specimen. However, it turns out that we can make some further use of the matrix
L
−1
. This matrix is (K +3) by (K +3); if we take the first K rows and the first K columns
of L
−1
, we can form L
−1
K
, which is called the bending energy matrix.

chap-06 4/6/2004 17: 23 page 149
THE THIN-PLATE SPLINE 149
The bending energy matrix can be rearranged into a series of eigenvectors E
i
, and
eigenvalues, λ
i
, such that:
L
−1
K
E
i
= λ
i
E
i
(6.51)
The eigenvectors E
i
have the usual properties of eigenvectors, and consequently they are a
basis (or a set of coordinate axes) of a space. In this case, the eigenvectors are the basis of
the Euclidean space tangent to shape space at the reference shape. This means that we can
express our matrix of observed deformations V as a linear combination of the eigenvectors
of the bending energy matrix. The eigenvalues are the bending energies required to effect
a change (of a given amount of shape difference, i.e. a unit of Procrustes distance) at that
spatial scale.
Three of the eigenvalues of the bending energy matrix are zero, corresponding to the
components with no bending (with X- and Y-coefficients, these eigenvectors account for
the six uniform components of the deformation). The remaining K −3 eigenvectors are
the explicitly localized components of a deformation. These eigenvectors are called the
partial warps; the vector multipliers of the partial warps are called the partial warp scores
(following Slice et al., 1996). They are “partial” because they describe part of a deforma-
tion. We should note that Bookstein (1991) called the eigenvectors of the bending energy
matrix principal warps, analogous to principal components. By “partial warp,” he meant
the vector multiple of a principal warp. Slice and colleagues use the term principal warp
to refer to a partial warp interpreted as a bent surface of the thin-plate spline, and because
the latter terminology has become standard, we use it here.
As evident in the definition of L
−1
K
, only one matrix of landmarks enters into the cal-
culation of bending energy; the coordinates of the form usually called the reference or
starting form. Thus, the eigenvectors that give us a coordinate system for shape analy-
ses are a function of one single form. This may be highly counterintuitive, because more
familiar eigenvectors, such as principal components, are functions of an observed variance–
covariance matrix. They are functions of variation (or differences) among observed forms.
That is not the case for the eigenvectors of the bending energy matrix. The eigenvalues
of the bending energy are the bending energies that would be required to modify a given
shape by a single unit of shape difference at each spatial scale. Thus the partial warps are
not themselves features of shape change, they are simply a coordinate system or basis for
the space in which we analyze shape change.
The “A” coefficients in Equation 6.48 describe the uniform deformation of the shape.
There are six of these coefficients, which is enough to describe the six components of the
uniform deformation of shape. However, we know that the reference and the target do
not differ by rotation, rescaling or translation, because those differences were removed by
the superimposition process. Consequently, we do not need six parameters to describe the
uniform component of the deformation, only the two components derived earlier in this
chapter.
By convention, partial (or principal) warps are numbered from the lowest to highest
bending energy; the one with the highest number corresponds to the one with greatest bend-
ing energy. The two uniform components are sometimes called the zero
th
principal warp.
Thinking of the uniform components in those terms is useful because it emphasizes that the
uniform components cannot be viewed separately from the non-uniform ones. Including

chap-06 4/6/2004 17: 23 page 150
150 GEOMETRIC MORPHOMETRICS FOR BIOLOGISTS
the uniform terms also completes the tally of shape variables. The K −3 partial warps
contribute 2K −6 scores; adding the two uniform scores brings the count up to 2K −4.
Using the thin-plate spline to visualize shape change
The combination of the uniform and non-uniform components completely describes any
shape change. The set of partial warp scores (including scores on the uniform component)
can be used in any conventional statistical analysis and, like the coordinates obtained by
GLS, the sum of their squares equals the squared Procrustes distance from the reference.
Moreover, like Bookstein’s shape coordinates, they have the correct degrees of freedom.
Thus we can use partial warps in any statistical procedure, such as regression, and diagram
the results as a deformation.
Interpreting changes depicted by the thin-plate spline
Interpretations should be presented in terms of the total deformation, not by detailing the
separate uniform and non-uniform components (or the more finely subdivided components
of them). Just as we cannot talk about individual landmarks as if they were separately
moved, we cannot talk about components of the total deformation as if they were separate
parts of the whole. It is important to remember that the changes depicted are based on
an interpolation function – we do not actually know what occurs between landmarks. If
we have sparsely sampled some regions of the body, we cannot assume that the spline
provides a realistic picture of their changes; there might be many highly localized changes
that cannot be detected in the absence of closely spaced landmarks. All we can say is that
our data do not require any more localized changes.
We cannot show an example of a biological transformation depicted by the thin-plate
spline until we have results to show, so we will borrow examples from a later chapter
(Chapter 10) to discuss the description of shape change using the thin-plate spline. In
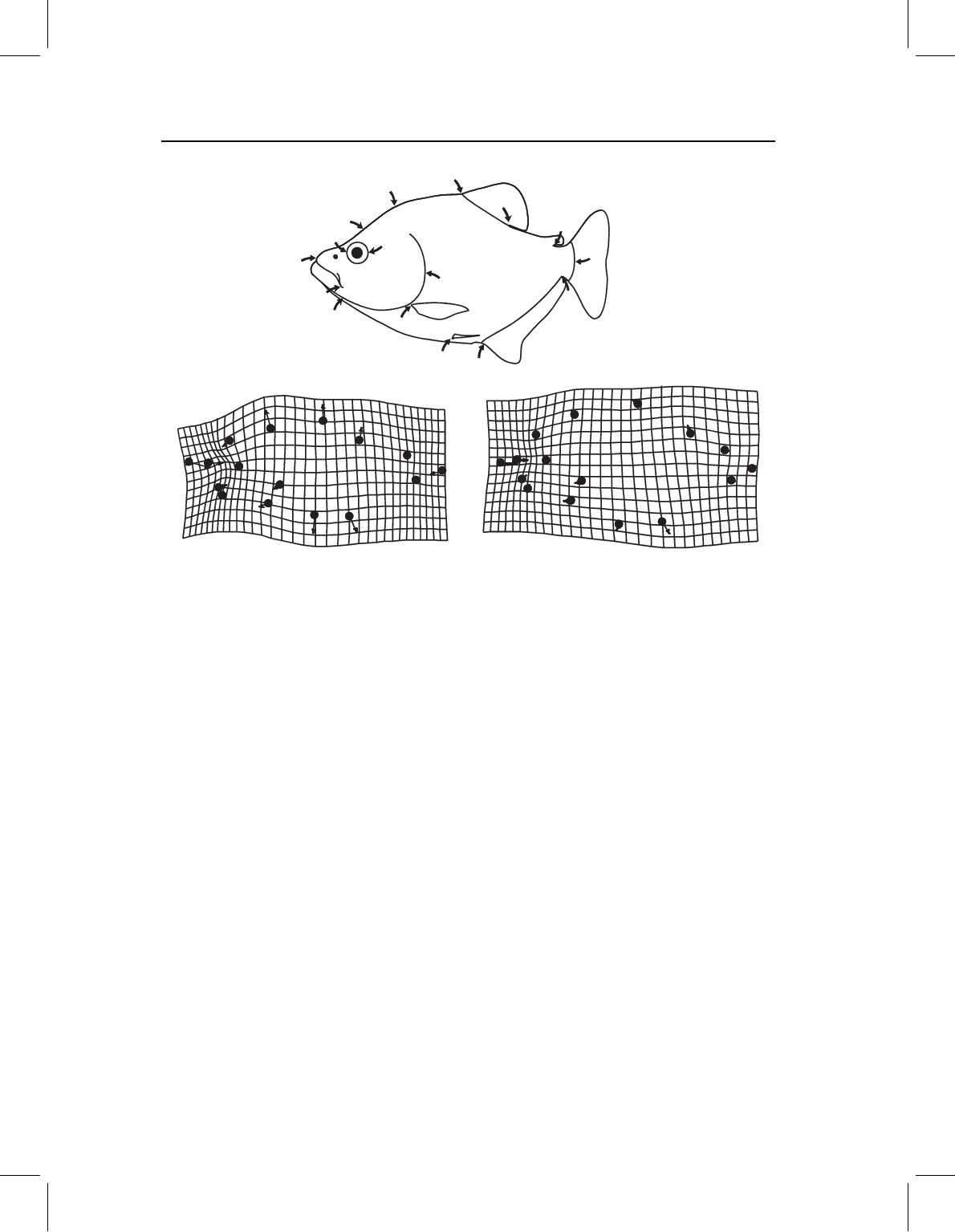
Figure 6.7 we depict the ontogenetic changes in body shape of two species of piranhas:
S. gouldingi (Figure 6.7A), which we used earlier in this chapter, and Pygopristis denticulata
(Figure 6.7B). In both species the head (as a whole) grows less rapidly than the middle of
the body, and the eye grows far more slowly than the head. In neither species does the
shortening of the eye result solely from the generally lower cranial growth rates; rather,
there is an abrupt (and localized) deceleration of growth rates in the orbital region. How-
ever, that does not, by itself, fully account for the apparent contraction of the grid in the
head, especially in S. gouldingi. Part of the relative shortening of the head, supraorbitally,
results from the displacement of the landmark at the epiphyseal bar (landmark 2) towards
the anterior landmark of the eye (landmark 14). Suborbitally, the apparent shortening
of the head results from the displacement of the posterior jaw landmark (landmark 13)
towards the posterior eye landmark (landmark 15), as well as from the more general short-
ening of the snout and eye. These two species also differ in the ontogeny of posterior body
shape. In S. gouldingi, the caudal peduncle (the region bounded by landmarks 6, 7, and
8) appears to contract, but no change appears to be localized there – the posterior body
generally shortens (as does the head). Growth rates appear to decrease, moving posteriorly
from the midbody to the tail. Because the caudal peduncle is the most posterior part of
the body, the growth rates are lowest there. In P. denticulata, growth rates decrease more
chap-06 4/6/2004 17: 23 page 151
THE THIN-PLATE SPLINE 151
4
9
1
2
3
5
6
7
8
11
12
13
14
15
16
10
(A) (B)
Figure 6.7 Ontogenetic shape change for two species of piranhas: (A) Serrasalmus gouldingi;
(B) Pygopristis denticulata.
slightly, and most of the change in the posterior body seems to result from the posterior
displacement (and relative shortening) of the anal fin. That increases the distance between
the pelvic and anal fins (which expands the grid between them), but because that is not a
part of the general expansion of the midbody (it is limited to the ventral region between
the fins) the change is ventrally localized. Due to the sparse sampling of landmarks in the
middle of the body, there is no abrupt contraction or expansion of the grid such as we see
in the head. Sparse sampling of that region makes it difficult to detect localized changes
because we cannot show what happens between landmarks when we have not sampled
them (quoting Gertrude Stein, “there is no there there”).
Software
Until we have results to depict, the spline serves the purpose of providing variables, with
the correct degrees of freedom, for statistical analysis. A file of partial warps, along with
the uniform components, can be computed by several programs in the IMP software series,
all of which output the data in a form that can be input into statistical packages (i.e. they
are in the X1,Y1,…CS format). They are perhaps most easily obtained from the Principal
Components Analysis program (PCAGen, discussed in Chapter 7), which calculates partial
warps relative to the mean shape (that is, the mean serves as the reference form). Within
that software, as in all the others, the explicit uniform terms are always calculated using the
partial Procrustes superimposition (meaning that centroid size is fixed to one). To draw the

chap-06 4/6/2004 17: 23 page 152
152 GEOMETRIC MORPHOMETRICS FOR BIOLOGISTS
deformations in different registrations, the software simply calculates the implicit uniform
deformations corresponding to the desired method of depicting the shape change. We will
return to this when we have results to depict.
References
Bookstein, F. L. (1989). Principal warps: thin-plate splines and the decomposition of deformations.
IEEE Transactions on Pattern Analysis and Machine Intelligence, 11, 567–585.
Bookstein, F. L. (1991). Morphometric Tools for Landmark Data: Geometry and Biology.
Cambridge University Press.
Bookstein, F. L. (1996). Standard formula for the uniform shape component in landmark data. In
Advances in Morphometrics (L. F. Marcus, M. Corti, A. Loy et al., eds) pp. 153–168. Plenum
Press.
Dryden, I. L. and Mardia, K. V. (1998). Statistical Shape Analysis. John Wiley & Sons.
Myers, P., Lundrigan, B. L., Gillespie, B. W. and Zelditch, M. L. (1996). Phenotypic plasticity in
skull and dental morphology in the prairie deer mouse (Peromyscus maniculatus bairdii). Journal
of Morphology, 229, 229–237.
Slice, D. E., Bookstein, F. L., Marcus, L. F. and Rohlf, F. J. (1996). Appendix I: A glossary for
geometric morphometrics. In Advances in Morphometrics (L. F. Marcus, M. Corti, A. Loy et al.,
eds) pp. 531–551. Plenum Press.
Thompson, D’Arcy W. (1942). On Growth and Form: A New Edition. Cambridge University Press.
(Reprinted in 1992 as On Growth and Form: The Complete Revised Edition, Dover Publications.)

chap-07 4/6/2004 17: 24 page 153
PART
II
Analyzing Shape Variables

chap-07 4/6/2004 17: 24 page 154

chap-07 4/6/2004 17: 24 page 155
7
Ordination methods
In this chapter, we discuss two methods for describing the diversity of shapes in a sam-
ple: principal components analysis (PCA) and canonical variates analysis (CVA). Our
discussion of these methods draws heavily on expositions presented by Morrison (1967),
Chatfield and Collins (1980), and Campbell and Atchley (1981). Both methods are used to
simplify descriptions rather than to test hypotheses. PCA is a tool for simplifying descrip-
tions of variation among individuals, whereas CVA is used for simplifying descriptions
of differences between groups. Both analyses produce new sets of variables that are lin-
ear combinations of the original variables. They also produce scores for individuals on
those variables, and these can be plotted and used to inspect patterns visually. Because
the scores order specimens along the new variables, the methods are called “ordination
methods.” It is hoped that the ordering provides insight into patterns in the data, per-
haps revealing patterns that are convenient for addressing biological questions. The most
important difference between PCA and CVA is that PCA constructs variables that can
be used to examine variation among individuals within a sample, whereas CVA con-
structs variables to describe the relative positions of groups (or subsets of individuals) in
the sample.
We discuss PCA and CVA in the same chapter because both serve a similar purpose,
and because the mathematical transformations performed in the two analyses are similar.
We describe PCA first because it is somewhat simpler, and because it provides a founda-
tion for understanding the transformations performed in CVA. We begin the description
of PCA with some simple graphical examples, and then present a more formal exposi-
tion of the mathematical mechanics of PCA. This is followed by a presentation of an
analysis of a real biological data set. The description of CVA follows a similar outline;
the only difference is that we begin with a discussion of groups and grouping variables.
CVA requires that the individuals be grouped, because the method analyzes the relative
positions of groups in the sample. Consequently, the sample must be divided into groups
before the analysis begins. The analysis of groups requires a few more computational steps
than PCA, but none of the steps in CVA introduce new mathematical concepts. CVA
will be just a new application of ideas you have already encountered in the discussion
of PCA.
Geometric Morphometrics for Biologists Copyright © 2004 Elsevier Ltd
ISBN 0–12–77846–08 All rights of reproduction in any form reserved

chap-07 4/6/2004 17: 24 page 156
156 GEOMETRIC MORPHOMETRICS FOR BIOLOGISTS
Principal components analysis
Geometric shape variables are neither biologically nor statistically independent. For exam-
ple, the shape variables produced by the thin-plate spline describe variation in overlapping
regions of an organism or structure. Because the regions overlap, they are under the influ-
ence of the same processes that produce variation; and therefore we expect them to be
correlated. Even when they do not describe overlapping regions, morphometric variables
(both geometric and traditional) are expected to be correlated because they describe fea-
tures of the organism that are functionally, developmentally or genetically linked. Their
patterns of variation and covariation are often complex and difficult to interpret. The pur-
pose of PCA is to simplify those patterns and make them easier to interpret by replacing the
original variables with new ones (principal components, PCs) that are linear combinations
of the original variables and independent of each other.
One might wonder why it would be a worthwhile exercise to take simple variables that
covary with each other and replace them with complex variables that do not covary. Part
of the value of this exercise arises from the fact that the new complex variable is a function
of the covariances among the original variables. It thus provides some insight into the
covariances among variables that can direct future research into the identity of the causal
factors underlying those covariances. Another useful purpose served by PCA is that most
of the variation in the sample usually can be described with only a few PCs. Again this is
useful, because it simplifies and clarifies what needs to be explained. Another important
benefit of PCA is that the presentation of results is simplified. It is much easier to produce
and explain plots of the three PCs that explain 90% of the variation than it is to separately
plot and explain the variation on each of 30 original variables.
An indirect benefit of PCA that is useful (but often misused) is that it simplifies the
description of differences among individuals. Clusters of individuals are often more appar-
ent in plots of PCs than in plots of the original variables. Finding such clusters can be quite
valuable, but those clusters do not represent evidence of statistically distinct entities. Legit-
imate methods for testing the hypothesis that a priori groups are statistically significantly
different will be presented in Chapters 8 and 9 (computer-based statistical methods and
multivariate analysis of variance, respectively).
Geometric description of PCA
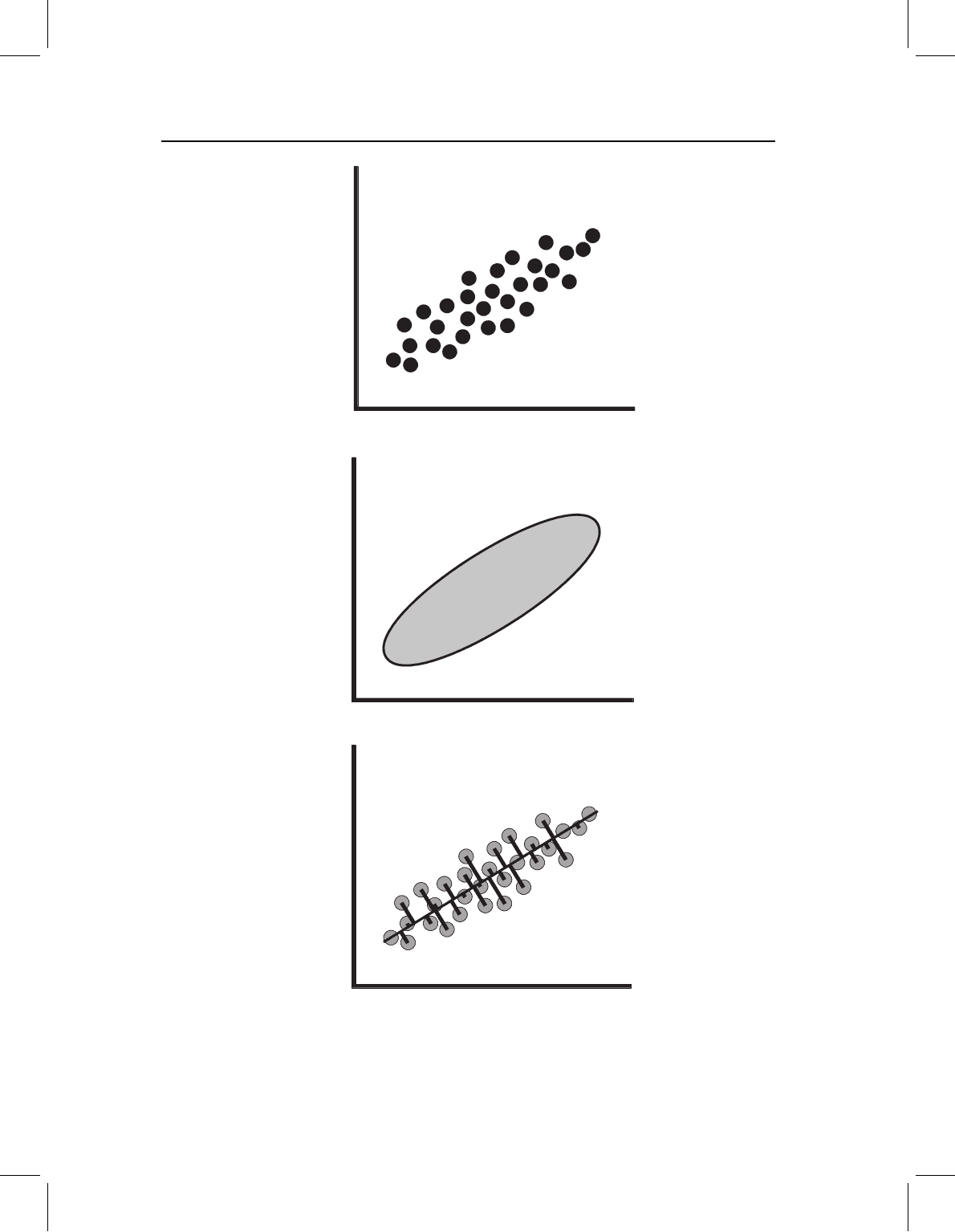
Figure 7.1A shows the simple case in which there are two observed traits, X
1
and X
2
.
These traits might be two distance measurements or the coordinates of a single landmark
in a two-dimensional shape analysis. Each point in the scatter plot represents the paired
values observed for a single specimen. We expect that the values of each trait are normally
distributed, and we expect that one trait is more variable than the other because one
variable, (in this case, X
1
) has a larger range of observed values and a higher variance. In
addition, the values of X
1
and X
2
are not independent; higher values of one are associated
with higher values of the other. This distribution of values can be summarized by an ellipse
that is tilted in the X
1
, X
2
coordinate plane (Figure 7.1B). PCA solves for the axes of this
ellipse, and uses those axes to describe the positions of individuals within that ellipse.
The first step of PCA is to find the direction through the scatter that describes the
largest proportion of the total variance. This direction, the long axis of the ellipse, is the
chap-07 4/6/2004 17: 24 page 157
ORDINATION METHODS 157
X
1
X
2
X
1
X
2
X
1
X
2
(A)
(B)
(C)
Figure 7.1 Graphical representation of the problem to be solved by PCA. (A) Scatter plot of indi-
viduals scored on two traits, X
1
and X
2
; (B) an ellipse enclosing the scatter of points shown in part
(A); (C) a line through the scatter and the perpendicular distances of the individuals from that axis.
The goal of PCA is to find the line that minimizes the sum of those squared distances.