Смирнов В.М. Физиология человека: Учебник
Подождите немного. Документ загружается.


44 %, а в 90 лет — более 50 % общей емкости
легких.
В покое парциальное давление кислорода
в альвеолах остается неизменным, чему спо-
собствует учащение дыхания. Вместе с тем
показатель максимальной вентиляции легких
уменьшается более чем в 2 раза, соответст-
венно снижаются и резервные возможности
дыхания.
Увеличение сопротивления дыхания
вследствие снижения бронхиальной проходи-
мости ведет при форсированном дыхании к
ограничению вентиляции легких. В этих ус-
ловиях объемная скорость выдоха и вдоха
после 80 лет уменьшается соответственно в
2,2-2,7 и 1,5—2 раза.
Развиваются существенные изменения аль-
веолокапиллярного газообмена. В то время
как парциальное давление кислорода в легких
к старческому возрасту почти не изменяется,
его напряжение и содержание в артериальной
крови заметно снижаются: насыщение арте-
риальной крови кислородом после 60 лет
на 5—7 % меньше, чем в зрелом возрасте. На-
пряжение кислорода в артериальной крови
60-летнего человека соответствует такого рода
параметру молодого человека, находящегося
на высоте
1,2—2,0
км над уровнем моря. Эту
величину (/>) можно рассчитать по формуле:
Р~
103,5 - 0,42 х возраст в годах (мм рт.ст.).
Это связано в основном с ухудшением ус-
ловий диффузии кислорода из легких в
кровь, что может быть обусловлено несколь-
кими факторами: 1) уменьшением количест-
ва альвеол (к 70 годам — примерно на 40 %);
2) увеличением неравномерности вентиля-
ции легких; 3) ухудшением кровоснабжения
альвеол и соотношения между вентиляцией и
13.1.
ОТКРЫТИЕ КРОВООБРАЩЕНИЯ.
ОБЩИЙ ПЛАН СТРОЕНИЯ СЕРДЕЧНО-
СОСУДИСТОЙ СИСТЕМЫ
А. Открытие. Эразистрат (III в. до н.э.) счи-
тал,
что артерии несут тканям воздух. Отсюда
название «артерия» (греч.: аег — воздух, te-
гео
—
содержу, храню). Это положение развил
основоположник экспериментальной меди-
цины Гален (II в. н.э.): он считал, что кровь
образуется в печени из пищи, которая после
обработки в желудке и кишечнике переходит
кровотоком; 4) уменьшением за счет сниже-
ния числа альвеол площади диффузии и уве-
личением расстояния диффузии, т.е. пути,
проходимого молекулами кислорода от аль-
веол до стенок кровеносных капилляров.
В результате диффузионная способность лег-
ких к 60 годам снижается примерно на 30 %.
Таким образом, для поглощения кровью оп-
ределенного количества кислорода из легких
необходимо увеличение их вентиляции по
сравнению с уровнем зрелого возраста.
Содержание в эритроцитах крови 2,3-ди-
фосфоглицерата у пожилых и старых людей
увеличивается, что облегчает отдачу кислоро-
да в крови, т.е. кривая диссоциации оксиге-
моглобина сдвигается вправо.
Перестраивается и регуляция дыхания:
увеличивается чувствительность хеморецеп-
торов каротидных телец к С0
2
и снижается
чувствительность рецепторов растяжения
легких, что в целом способствует поддержа-
нию адекватной вентиляции легких.
Таким образом, возрастная активация хе-
морецепторов, учащение дыхания и улучше-
ние диссоциации оксигемоглобина в опреде-
ленной степени компенсируют структурные и
функциональные изменения системы дыха-
ния. Эти механизмы включаются в состоянии
покоя. При предъявлении же к организму по-
вышенных требований, например при мы-
шечной работе или при снижении содержания
кислорода во вдыхаемом воздухе, ограниче-
ние функционального резерва дыхания суще-
ственно снижает адаптационные возможнос-
ти организма. Известно, в частности, что в
возрасте 20—40 лет максимальная физическая
нагрузка может вызвать девятикратное по-
вышение потребления кислорода. Однако с 26
до 63 лет максимальное потребление кислоро-
да снижается почти в 2 раза.
в печень по протокам. Далее кровь из печени
разносится по венам ко всем частям тела, где
и потребляется. Согласно Галену, часть кро-
ви поступает в правый желудочек, затем пе-
реходит через отверстия перегородки в левый
желудочек — наличие крови в нем Гален до-
казал с помощью пункции. В левом желудоч-
ке кровь смешивается с воздухом, поступа-
ющим из легких, а затем разносится по арте-
риям во все органы тела, в мозг. В мозге
кровь превращается в «животный дух», не-
обходимый для движения каждой части тела.
Глава 13 СЕРДЕЧНО-СОСУДИСТАЯ СИСТЕМА
п\

Ибн-аль-Нафиз (XIII в.) впервые пришел к
заключению, что вся кровь из правого желу-
дочка проходит через сосуды легких и воз-
вращается в левое сердце. М.Сервет (1509—
1553) описал малый круг кровообращения.
Он,
в частности, установил, что кровь к лег-
кому идет по сосуду (легочной артерии), диа-
метр которого равен диаметру аорты, причем
по артериям течет венозная кровь, которая в
легких освобождается от «сажи».
У.Гарвей (1628) открыл кровообращение в
организме. В своей работе «Анатомическое
исследование о движении сердца и крови у
животных» он с безупречной логикой опро-
верг господствовавшую в течение более 1500
лет доктрину Галена. Измерив у овцы вели-
чину систолического объема крови, частоту
сокращений сердца в минуту и общее коли-
чество крови, Гарвей утверждал: «Во всем
теле крови не больше 4 фунтов, как я убедил-
ся в этом на овце». Он подсчитал, что за 1,5—
2 мин вся кровь должна пройти через сердце,
а в течение 30 мин через сердце должно
пройти количество крови, равное массе тела
животного. Столь быстрое и непрерывное
производство крови в организме невозмож-
но.
Гарвей допускал возврат одной и той же
крови к сердцу через посредство замкнутого
цикла. Замкнутость круга кровообращения
Гарвей объяснил прямым соединением арте-
рий и вен через посредство мельчайших тру-
бочек (капилляров), которые были открыты
М.Мальпиги спустя 4 года после смерти Гар-
вея.
Замкнутая система кровообращения, по
Гарвею, имеет два круга — большой и малый
(легочный), которые соединены между собой
через сердце. Малый круг кровообращения
осуществляет прямой контакт с внешней сре-
дой, а большой — с органами и тканями ор-
ганизма.
Б.
Общий план строения. Центральным
звеном сердечно-сосудистой системы являет-
ся четырехкамерное сердце. Оно состоит из
левой и правой половин, каждая из которых
включает предсердие и желудочек. Сердце —
полый орган, стенки которого представляют
собой поперечнополосатую мускулатуру. Од-
нако сердце иннервируется не соматической,
а вегетативной нервной системой. Оно окру-
жено конусовидным мешком — околосердеч-
ной сумкой (перикардом). Наружный слой
перикарда состоит из нерастяжимой белой
фиброзной ткани, внутренний слой — из
двух листков: висцерального и париетально-
го.
Висцеральный листок сращен с сердцем,
а париетальный — с фиброзной тканью.
В щель между этими листками выделяется
перикардиальная жидкость, которая умень-
шает трение между стенками сердца и пери-
кардом. Плотность ткани перикарда препят-
ствует излишнему растяжению сердца или
переполнению его кровью. В зависимости от
морфологических и функциональных осо-
бенностей в сердце различают два типа мы-
шечных волокон: 1) волокна рабочего миокар-
да предсердий и желудочков, составляющие
основную массу сердца и обеспечивающие
его нагнетательную функцию; 2) волокна про-
водящей системы сердца, отвечающие за гене-
рацию и проведение возбуждения к клеткам
рабочего миокарда. Левая и правая половины
сердца непосредственно не сообщаются
между собой, отверстия имеются только
между предсердиями и желудочками каждой
половины сердца. Левое предсердие и левый
желудочек разделены между собой двуствор-
чатым клапаном; правое предсердие и пра-
вый желудочек — трехстворчатым клапаном.
Вход в аорту и легочную артерию перекрыва-
ют полулунные клапаны. В области впадения
вен в предсердия имеется кольцевая мускула-
тура (сфинктеры, выполняющие функцию
клапанов — обеспечение одностороннего
тока крови по сердцу и направление ее в
большой и малый круг кровообращения).
В.
Главная роль сердечно-сосудистой сис-
темы — обеспечение выполнения транспорт-
ной функции крови; только при движении
кровь может выполнять свою главную функ-
цию — транспорт различных веществ в орга-
низме, причем обмен веществ между кровью
и тканями происходит только в капиллярах,
их суммарная площадь огромна — достигает
1000 м
2
. В стенках сердца и сосудов выраба-
тываются многие биологически активные ве-
щества, например атриопептид — натрийуре-
тический гормон (см. раздел 17.4), гепарин,
гистамин, серотонин, факторы свертывания
крови, эндотелии — сосудосуживающее ве-
щество, антитела.
Деление на большой и малый круги кро-
вообращения условно: они сообщены между
собой, один является продолжением другого,
т.е.
два круга включены последовательно, это
замкнутая система (рис. 13.1).
Две части сердечно-сосудистой системы
названы так потому, что каждая из них начи-
нается в сердце и возвращается в сердце, но
по отдельности замкнутых кругов они не об-
разуют. Фактически имеется один общий
замкнутый круг кровообращения. Из левого
желудочка кровь поступает в аорту, далее по
артериям она следует в капилляры всех орга-
нов и тканей организма, по венам возвраща-
ется в правое предсердие, правый желудочек
и по легочной артерии поступает в легкие. Из
272
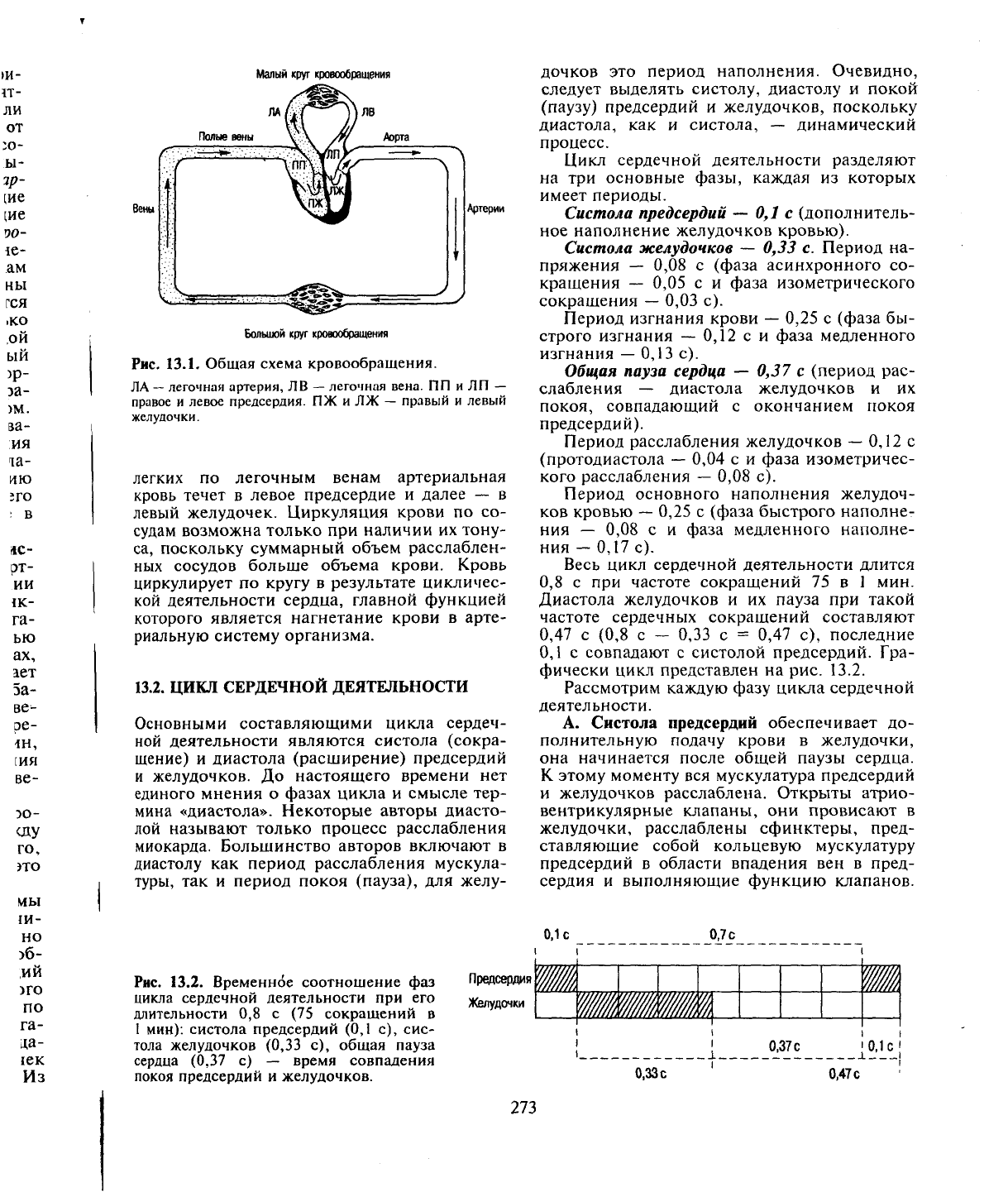
Малый круг кровообращения
Вены
Артерии
Большой круг кровообращения
Рис. 13.1. Общая схема кровообращения.
ЛА
—
легочная артерия,
ЛВ
—
легочная вена. ПП
и ЛП —
правое
и
левое предсердия. ПЖ
и
ЛЖ
—
правый
и
левый
желудочки.
легких по легочным венам артериальная
кровь течет в левое предсердие и далее — в
левый желудочек. Циркуляция крови по со-
судам возможна только при наличии их тону-
са, поскольку суммарный объем расслаблен-
ных сосудов больше объема крови. Кровь
циркулирует по кругу в результате цикличес-
кой деятельности сердца, главной функцией
которого является нагнетание крови в арте-
риальную систему организма.
13.2.
ЦИКЛ СЕРДЕЧНОЙ ДЕЯТЕЛЬНОСТИ
Основными составляющими цикла сердеч-
ной деятельности являются систола (сокра-
щение) и диастола (расширение) предсердий
и желудочков. До настоящего времени нет
единого мнения о фазах цикла и смысле тер-
мина «диастола». Некоторые авторы диасто-
лой называют только процесс расслабления
миокарда. Большинство авторов включают в
диастолу как период расслабления мускула-
туры, так и период покоя (пауза), для желу-
дочков это период наполнения. Очевидно,
следует выделять систолу, диастолу и покой
(паузу) предсердий и желудочков, поскольку
диастола, как и систола, — динамический
процесс.
Цикл сердечной деятельности разделяют
на три основные фазы, каждая из которых
имеет периоды.
Систола
предсердий — 0,1 с (дополнитель-
ное наполнение желудочков кровью).
Систола желудочков — 0,33 с. Период на-
пряжения — 0,08 с (фаза асинхронного со-
кращения — 0,05 с и фаза изометрического
сокращения
—
0,03 с).
Период изгнания крови — 0,25 с (фаза бы-
строго изгнания — 0,12 с и фаза медленного
изгнания
—
0,13 с).
Общая пауза сердца — 0,37 с (период рас-
слабления — диастола желудочков и их
покоя, совпадающий с окончанием покоя
предсердий).
Период расслабления желудочков
—
0,12 с
(протодиастола — 0,04 с и фаза изометричес-
кого расслабления
—
0,08 с).
Период основного наполнения желудоч-
ков кровью
—
0,25 с (фаза быстрого наполне-
ния — 0,08 с и фаза медленного наполне-
ния
—
0,17 с).
Весь цикл сердечной деятельности длится
0,8 с при частоте сокращений 75 в 1 мин.
Диастола желудочков и их пауза при такой
частоте сердечных сокращений составляют
0,47 с (0,8 с — 0,33 с = 0,47 с), последние
0,1 с совпадают с систолой предсердий. Гра-
фически цикл представлен на рис. 13.2.
Рассмотрим каждую фазу цикла сердечной
деятельности.
А. Систола предсердий обеспечивает до-
полнительную подачу крови в желудочки,
она начинается после общей паузы сердца.
К этому моменту вся мускулатура предсердий
и желудочков расслаблена. Открыты атрио-
вентрикулярные клапаны, они провисают в
желудочки, расслаблены сфинктеры, пред-
ставляющие собой кольцевую мускулатуру
предсердий в области впадения вен в пред-
сердия и выполняющие функцию клапанов.
Рис. 13.2. Временное соотношение фаз
цикла сердечной деятельности при его
длительности 0,8 с (75 сокращений в
1 мин): систола предсердий (0,1 с), сис-
тола желудочков (0,33 с), общая пауза
сердца (0,37 с) — время совпадения
покоя предсердий и желудочков.
0,1с
Предсердия
Желудочки
W//M
W/////
0,7 с
ы
'////////
0,37 с
•
0,1с
L
0,33 с
0,47с
273

Поскольку весь рабочий миокард расслаблен,
давление в полостях сердца равно нулю. Из-
за градиента давления в полостях сердца и
артериальной системе полулунные клапаны
закрыты.
Возбуждение и, следовательно, волна со-
кращения предсердий начинаются в области
впадения полых вен, поэтому одновременно
с сокращением рабочего миокарда предсер-
дий сокращается и мускулатура сфинктеров,
выполняющих функцию клапанов, — они за-
крываются, давление в предсердиях начинает
расти, и дополнительная порция крови (при-
мерно 1/5 от конечно-диастолического объе-
ма) поступает в желудочки.
Во время систолы предсердий кровь из них
обратно в полые и легочные вены не возвра-
щается, так как сфинктеры закрыты. К концу
систолы давление в левом предсердии возрас-
тает до 10—12 мм рт.ст., в правом — до 4—
8 мм рт.ст. Такое же давление к концу систо-
лы предсердий создается и в желудочках. Та-
ким образом, во время систолы предсердий
сфинктеры предсердий закрыты, атриовент-
рикулярные клапаны открыты. Поскольку в
аорте и легочной артерии давление крови в
этот период больше, то полулунные клапаны,
естественно, все еще закрыты. После оконча-
ния систолы предсердий, через 0,007 с (интер-
систолический интервал), начинаются систо-
ла желудочков, диастола предсердий и их
покой. Последние длятся 0,7 с, предсердия
при этом наполняются кровью (резервуарная
функция предсердий). Значение систолы
предсердий заключается еще и в том, что воз-
никающее при этом давление обеспечивает
дополнительное растяжение миокарда желу-
дочков и последующее усиление их сокраще-
ний во время систолы желудочков.
Б.
Систола желудочков состоит из двух пе-
риодов — напряжения и изгнания, каждый
из которых делится на две фазы. В фазе асин-
хронного (неодновременного) сокращения воз-
буждение мышечных волокон распространя-
ется по обоим желудочкам. Сокращение на-
чинается с ближайших к проводящей системе
сердца участков рабочего миокарда (сосочко-
вых мышц, перегородки, верхушки желудоч-
ков).
К концу этой фазы в сокращение вовле-
чены все мышечные волокна, поэтому давле-
ние в желудочках начинает быстро повы-
шаться, вследствие чего закрываются атрио-
вентрикулярные клапаны и начинается фаза
изометрического сокращения. Сокращающие-
ся вместе с желудочками сосочковые мышцы
натягивают сухожильные нити и препятству-
ют выворачиванию клапанов в предсердия.
Кроме того, эластичность и растяжимость су-
хожильных нитей смягчают удар крови об ат-
риовентрикулярные клапаны, что обеспечи-
вает долговечность их работы. Общая поверх-
ность атриовентрикулярных клапанов боль-
ше площади атриовентрикулярного отверс-
тия,
поэтому их створки плотно прижимают-
ся друг к другу. Благодаря этому клапаны на-
дежно смыкаются даже при изменениях
объема желудочков и кровь не возвращается
во время систолы желудочков обратно в
предсердия. Во время фазы изометрического
сокращения давление в желудочках быстро
нарастает. В левом желудочке оно увеличива-
ется до 70—80 мм рт.ст., в правом — до 15—
20 мм рт.ст. Как только давление в левом же-
лудочке окажется больше диастолического
давления в аорте (70—80 мм рт.ст.), а в пра-
вом желудочке — больше диастолического
давления в легочной артерии (15—20 мм
рт.ст.),
открываются полулунные клапаны и
начинается период изгнания.
Оба желудочка сокращаются одновремен-
но,
причем волна их сокращения начинается
в верхушке сердца и распространяется вверх,
выталкивая кровь из желудочков в аорту и
легочный ствол. В период изгнания длина
мышечных волокон и объем желудочков
уменьшаются, атриовентрикулярные клапа-
ны закрыты, так как давление в желудочках
высокое, а в предсердиях оно равно нулю.
В период быстрого изгнания давление в
левом желудочке достигает 120—140 мм рт.ст.
(систолическое давление в аорте и крупных
артериях большого круга), а в правом желу-
дочке — 30—40 мм рт.ст. В период медленно-
го изгнания давление в желудочках начинает
падать. Состояние клапанов сердца пока не
изменяется — закрыты только атриовентри-
кулярные клапаны, полулунные клапаны от-
крыты, предсердные сфинктеры также от-
крыты, потому что весь миокард предсердий
расслаблен, кровь заполняет предсердия.
Во время периода изгнания крови из же-
лудочков реализуется процесс засасывания
крови из крупных вен в предсердия. Это обу-
словлено тем, что плоскость атриовентрику-
лярной «перегородки», которую формируют
соответствующие клапаны, смещается по на-
правлению к верхушке сердца, при этом
предсердия, находящиеся в расслабленном
состоянии, растягиваются, что способствует
заполнению их кровью.
Вслед за фазой изгнания начинаются диа-
стола желудочков и их пауза (покой), с кото-
рой частично совпадает и пауза предсердий,
поэтому данный период сердечной деятель-
ности предлагается называть общей паузой
сердца.
274

В.
Общая пауза сердца начинается с про-
тодиастолы —
это период от начала расслаб-
ления мышц желудочков до закрытия полу-
лунных клапанов. Давление в желудочках
становится несколько ниже, чем в аорте и ле-
гочной артерии, поэтому полулунные клапа-
ны закрываются. В фазе изометрического рас-
слабления
полулунные клапаны уже закрыты,
а атриовентрикулярные еще не открыты. По-
скольку расслабление желудочков продолжа-
ется, давление в них падает, что приводит к
открытию атриовентрикулярных клапанов
массой крови, накопившейся во время диа-
столы в предсердиях. Начинается период на-
полнения желудочков, расширение которых
обеспечено несколькими факторами.
1.
Расслабление желудочков и расширение
их камер происходит в основном за счет
части энергии, которая расходуется во время
систолы на преодоление сил упругости серд-
ца (потенциальная энергия). Во время систо-
лы сердца сжимаются его соединительно-
тканный упругий каркас и мышечные волок-
на, которые имеют различное направление в
разных слоях. Желудочек в этом отношении
можно сравнить с резиновой грушей, которая
принимает прежнюю форму после того, как
на нее надавили, расширение желудочков
оказывает некоторое присасывающее дейст-
вие.
2.
Левый желудочек (правый — в мень-
шей степени) во время фазы изометрическо-
го сокращения мгновенно становится круг-
лым, поэтому в результате действия сил гра-
витации обоих желудочков и находящейся в
них крови быстро растягиваются крупные
сосуды, на которых «висит» сердце. При
этом атриовентрикулярная «перегородка»
несколько смещается вниз. При расслабле-
нии мускулатуры желудочков атриовентри-
кулярная «перегородка» вновь поднимается
вверх, что также способствует расширению
камер желудочков, ускоряет наполнение их
кровью.
3.
В фазе быстрого наполнения кровь,
скопившаяся в предсердиях, сразу провали-
вается в расслабленные желудочки и способ-
ствует их расправлению.
4.
Расслаблению миокарда желудочков
способствует давление крови в коронарных
артериях, которая в это время начинает уси-
ленно поступать из аорты в толщу миокарда
(«гидравлический каркас сердца»).
5.
Дополнительное растяжение мускулату-
ры желудочков осуществляется за счет энер-
гии систолы предсердий (повышение давле-
ния в желудочках во время систолы предсер-
дий).
6. Остаточная энергия венозной крови,
сообщенная ей сердцем во время систолы
(этот фактор действует в фазе медленного на-
полнения).
Таким образом, во время общей паузы
предсердий и желудочков сердце отдыхает,
его камеры наполняются кровью, миокард
интенсивно снабжается кровью, получает
кислород и питательные вещества. Это весь-
ма важно, так как во время систолы коронар-
ные сосуды сжимаются сокращающимися
мышцами, при этом кровоток в коронарных
сосудах практически отсутствует.
13.3.
ОСОБЕННОСТИ СВОЙСТВ СЕРДЕЧНОЙ
МЫШЦЫ
Свойства сердечной мышцы определяются
структурными особенностями кардиомиоци-
тов и их функциональными взаимоотноше-
ниями. В сердце имеются мышечные волокна
двух типов — клетки рабочего миокарда, со-
ставляющие основную массу сердца и обес-
печивающие его механическую деятельность,
а также атипические мышечные волокна, об-
разующие проводящую систему сердца и
обеспечивающие его автоматию. В этих клет-
ках зарождается возбуждение, которое потом
распространяется по всему сердцу. Мышеч-
ные волокна рабочего миокарда предсердий
и желудочков разделены соединительноткан-
ной предсердно-желудочковой перегородкой,
связь между ними осуществляется только в
одном участке, посредством проводящей сис-
темы сердца (см. раздел 13.5).
В желудочках имеется три мышечных
слоя, идущих в различных направлениях: на-
ружный косой; средний, самый мощный
(кольцевой) и внутренний — продольный
слой, формирующий сосочковые мышцы.
Последние предотвращают выворачивание
атриовентрикулярных клапанов в сторону
предсердий во время систолы желудочков.
Предсердия имеют два мышечных слоя: на-
ружный — циркулярный (единый для обоих
предсердий) и внутренний — продольный
(раздельный для каждого предсердия). Тол-
щина стенок отделов сердца сильно различа-
ется: самую толстую стенку имеет левый же-
лудочек, тоньше стенка правого желудочка,
еще тоньше — предсердия, что определяется
различной нагрузкой на мышцу. Постепенно
увеличивающаяся в процессе эволюции на-
грузка на желудочек сердца и последующее
его разделение на две изолированные камеры
сформировали мощный рабочий миокард ле-
вого желудочка.
275

Сердечной мышце в целом присущи те же
свойства, что и скелетной мышце, — возбу-
димость, проводимость, сократимость, растя-
жимость, эластичность. Вместе с тем есть су-
щественные отличия.
13.3.1.
ВОЗБУДИМОСТЬ И ПРОВОДИМОСТЬ
А. Возбудимость — это способность клеток
генерировать потенциал действия (ПД).
1.
Параметры потенциала покоя и ПД.
Потенциал покоя клеток рабочего миокарда
формируется в основном градиентом К
+
, его
величина в среднем составляет 85—90 мВ.
Величина ПД составляет 120 мВ. Длитель-
ность ПД кардиомиоцитов желудочков (300—
400 мс) почти соответствует длительности со-
кращения мышцы сердца. У кардиомиоцитов
предсердия продолжительность ПД составля-
ет 100 мс, почти столько же длится систола
предсердий. Длительность ПД кардиомиоци-
та значительно уменьшается, если очередной
импульс приходит раньше — сразу после
окончания рефрактерной фазы. Этот фено-
мен можно наблюдать в эксперименте при
укорочении интервалов между отдельными
раздражениями сердечной мышцы. Меха-
низм уменьшения продолжительности ПД в
описанном опыте (рис. 13.3) связан с ускоре-
нием процесса реполяризации в цикле воз-
буждения, что объясняется еще сохраняю-
щейся повышенной проницаемостью мем-
браны для калия.
2.
Ионный механизм возникновения ПД
кардиомиоцитов. Фазы деполяризации и ин-
версии (вся восходящая часть ПД) осущест-
вляется в основном за счет входа Na
+
в клет-
ку, как у миоцитов скелетной мышцы. В дан-
ный период увеличена проницаемость бы-
стрых Na-каналов, когда снижение мембран-
ного потенциала достигает —60 мВ, Na
+
лави-
ной поступает в клетку. При дальнейшей де-
поляризации до —40 мВ активируются мед-
ленные электрочувствительные Na/Ca-кана-
лы,
по которым дополнительно Na
+
и в боль-
шей степени Са
2+
начинают входить в клетку.
В фазе деполяризации Na
+
и Са
2+
по бы-
стрым и медленным каналам идут в клетку
согласно концентрационному и электричес-
кому градиентам (клетка в эту фазу внутри
еще имеет отрицательный заряд). В фазе ин-
версии оба иона входят в клетку только со-
гласно концентрационному градиенту и во-
преки электрическому — в этот период клет-
ка перезаряжается: внутри нее возникает по-
ложительный заряд, снаружи — отрицатель-
ный.
+30-,
к
+20-
^-^ кЧ
О- \^ ОХ
-20-
\ \ \
-40-
\ \ \
-60-
\ \ V
-80-
_] \|^ V \
-100-1
Рис. 13.3. Укорочение потенциала действия сокра-
тительных миоцитов желудочков с увеличением
частоты раздражения.
Далее нарастание ПД прекращается вслед-
ствие инактивации быстрых Na-каналов, не-
смотря на то, что ток Na
+
и Са
2+
внутрь клет-
ки по медленным каналам продолжается. На-
чинается спад ПД в результате активации
К-каналов и выхода К
+
из клетки. Вначале
снижение ПД происходит быстро, очевидно,
вследствие входа СГ в клетку согласно кон-
центрационному и электрическому градиен-
там (отрицательно заряженных ионов хлора,
как известно, больше вне клетки, которая во
время инверсии внутри заряжена положи-
тельно относительно наружной поверхнос-
ти).
В кардиомиоцитах обнаружены хлорные
потенциалчувствительные каналы. Затем, в
фазе инверсии, медленный суммарный вход
Na
+
и Са
2+
в клетку примерно равен медлен-
ному току К
+
из клетки, что обеспечивает
возникновение плато ПД. Медленный спад
(плато) свидетельствует о том, что выход К
+
из клетки несколько превышает вход Na
+
и
Са
2+
в клетку вследствие начинающейся
инактивации медленных Na/Ca-каналов.
Полная инактивация этих каналов заканчи-
вается при мембранном потенциале 0 —
(+10) мВ, т.е. сразу после фазы инверсии или
в начале фазы реполяризации. К этому мо-
менту активация К-каналов быстро нараста-
ет, и К
+
начинает лавиной выходить из клет-
ки,
что обеспечивает фазу реполяризации
кардиомиоцита. Мембранный потенциал
возвращается к исходной величине 85—
90 мВ. В фазе инверсии К
+
выходит из клет-
ки согласно концентрационному и электри-
ческому градиентам, в фазе реполяризации
—
согласно концентрационному, но вопреки
электрическому: в этой фазе клетка снаружи
уже снова заряжена положительно (рис.
13.4).
3.
Длительность
фазы абсолютной реф-
рактерности кардиомиоцитов в 100—300 раз
больше
таковой миоцитов скелетной мышцы.
У последней, как известно, пик ПД составля-
276
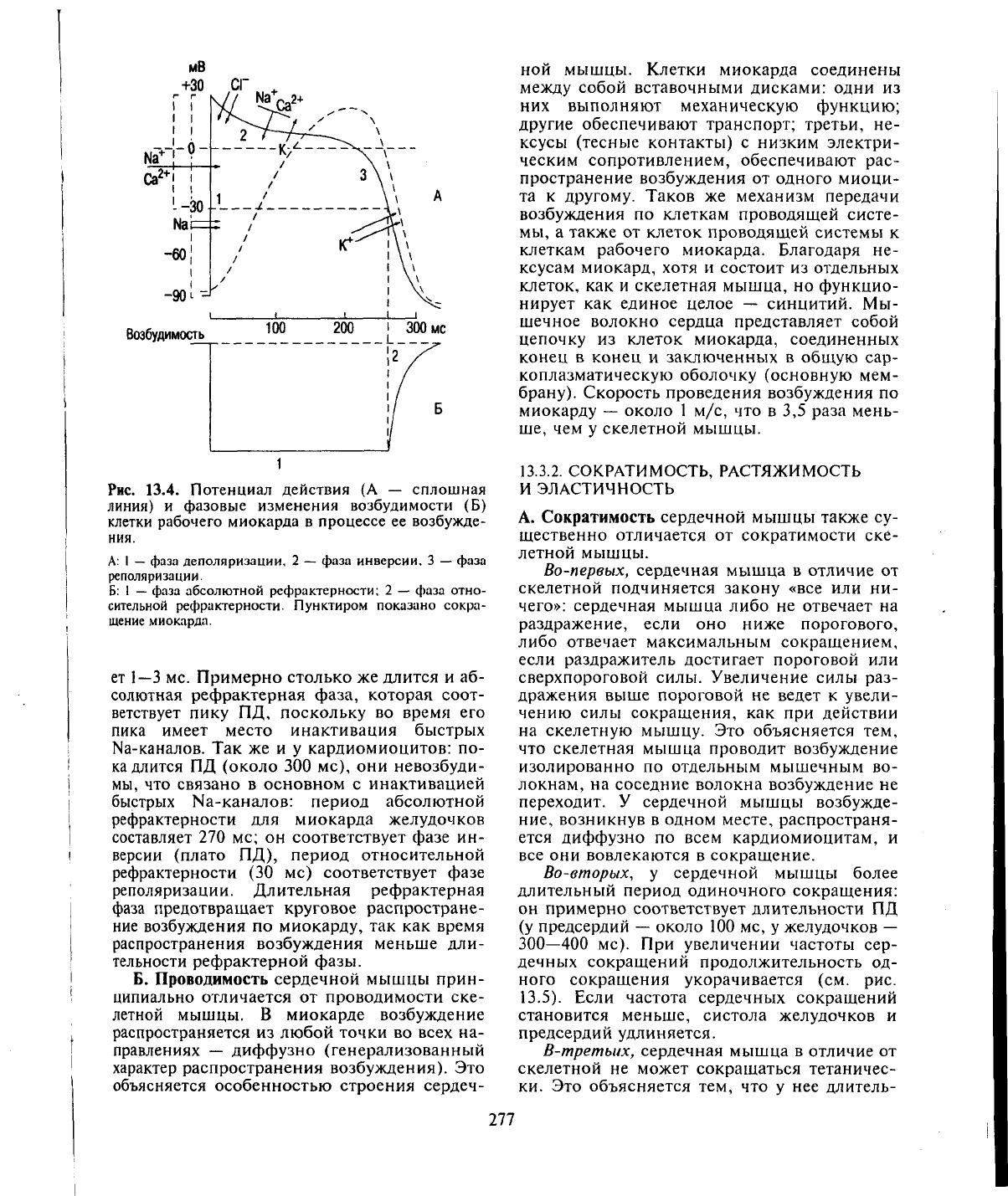
Рис. 13.4. Потенциал действия (А — сплошная
линия) и фазовые изменения возбудимости (Б)
клетки рабочего миокарда в процессе ее возбужде-
ния.
А: I
—
фаза деполяризации, 2
—
фаза инверсии. 3
—
фаза
реполяризации.
Б:
1
—
фаза абсолютной рефрактерности; 2 — фаза отно-
сительной рефрактерности. Пунктиром показано сокра-
щение миокарда.
ет 1—3 мс. Примерно столько же длится и аб-
солютная рефрактерная фаза, которая соот-
ветствует пику ПД, поскольку во время его
пика имеет место инактивация быстрых
Na-каналов. Так же и у кардиомиоцитов: по-
ка длится ПД (около 300 мс), они невозбуди-
мы,
что связано в основном с инактивацией
быстрых Na-каналов: период абсолютной
рефрактерности для миокарда желудочков
составляет 270 мс; он соответствует фазе ин-
версии (плато ПД), период относительной
рефрактерности (30 мс) соответствует фазе
реполяризации. Длительная рефрактерная
фаза предотвращает круговое распростране-
ние возбуждения по миокарду, так как время
распространения возбуждения меньше дли-
тельности рефрактерной фазы.
Б.
Проводимость сердечной мышцы прин-
ципиально отличается от проводимости ске-
летной мышцы. В миокарде возбуждение
распространяется из любой точки во всех на-
правлениях — диффузно (генерализованный
характер распространения возбуждения). Это
объясняется особенностью строения сердеч-
ной мышцы. Клетки миокарда соединены
между собой вставочными дисками: одни из
них выполняют механическую функцию;
другие обеспечивают транспорт; третьи, не-
ксусы (тесные контакты) с низким электри-
ческим сопротивлением, обеспечивают рас-
пространение возбуждения от одного миоци-
та к другому. Таков же механизм передачи
возбуждения по клеткам проводящей систе-
мы,
а также от клеток проводящей системы к
клеткам рабочего миокарда. Благодаря не-
ксусам миокард, хотя и состоит из отдельных
клеток, как и скелетная мышца, но функцио-
нирует как единое целое — синцитий. Мы-
шечное волокно сердца представляет собой
цепочку из клеток миокарда, соединенных
конец в конец и заключенных в общую сар-
коплазматическую оболочку (основную мем-
брану). Скорость проведения возбуждения по
миокарду — около 1 м/с, что в 3,5 раза мень-
ше,
чем у скелетной мышцы.
13.3.2.
СОКРАТИМОСТЬ, РАСТЯЖИМОСТЬ
И ЭЛАСТИЧНОСТЬ
А. Сократимость сердечной мышцы также су-
щественно отличается от сократимости ске-
летной мышцы.
Во-первых, сердечная мышца в отличие от
скелетной подчиняется закону «все или ни-
чего»: сердечная мышца либо не отвечает на
раздражение, если оно ниже порогового,
либо отвечает максимальным сокращением,
если раздражитель достигает пороговой или
сверхпороговой силы. Увеличение силы раз-
дражения выше пороговой не ведет к увели-
чению силы сокращения, как при действии
на скелетную мышцу. Это объясняется тем,
что скелетная мышца проводит возбуждение
изолированно по отдельным мышечным во-
локнам, на соседние волокна возбуждение не
переходит. У сердечной мышцы возбужде-
ние,
возникнув в одном месте, распространя-
ется диффузно по всем кардиомиоцитам, и
все они вовлекаются в сокращение.
Во-вторых, у сердечной мышцы более
длительный период одиночного сокращения:
он примерно соответствует длительности ПД
(у предсердий
—
около 100 мс, у желудочков
—
300—400 мс). При увеличении частоты сер-
дечных сокращений продолжительность од-
ного сокращения укорачивается (см. рис.
13.5).
Если частота сердечных сокращений
становится меньше, систола желудочков и
предсердий удлиняется.
В-третьих, сердечная мышца в отличие от
скелетной не может сокращаться тетаничес-
ки.
Это объясняется тем, что у нее длитель-
277

ный рефрактерный период: он равен продол-
жительности ПД и одиночного сокращения
мышцы. Это означает, что весь период со-
кращения мышца сердца невозбудима. Воз-
никновение тетанического сокращения
могло бы нарушить нагнетательную функцию
сердца.
В-четвертых, существенно отличается
роль внеклеточного Са
2+
в электромеханичес-
ком сопряжении мышцы сердца. Система
продольных трубочек, служащих внутрикле-
точным резервуаром Са
2+
, выражена в мень-
шей степени, чем в скелетных мышцах. Поэ-
тому Са
2+
, входящий в клетку при возбужде-
нии, не только играет важную роль в электро-
механическом сопряжении, но участвует в де-
поляризации клетки и увеличивает длитель-
ность ПД кардиомиоцитов. Входящий Са
2+
за-
пускает процесс освобождения внутриклеточ-
ного Са
2+
из саркоплазматического ретикулу-
ма, далее он обеспечивает до 30 % общего
внутриклеточного количества Са
2+
, необходи-
мого для взаимодействия нитей актина и мио-
зина. Остальные 70 % Са
2+
выбрасываются
саркоплазматическим ретикулумом.
Естественно, что все факторы, способст-
вующие входу Са
2+
в клетку, увеличивают
силу сердечных сокращений. Например, пар-
ные стимулы увеличивают длительность
плато ПД и вход Са
2+
в клетку, что ведет к
увеличению силы сокращений миокарда.
Сердечные гликозиды способствуют накоп-
лению Са
2+
в кардиомиоцитах и увеличению
их сокращений. Увеличение содержания Са
2+
в крови также ведет к усилению сердечных
сокращений. В лабораторных условиях чрез-
мерно высокая концентрация Са
2+
в перфу-
зирующем растворе вызывает остановку
сердца в систоле. Уменьшение содержания
Са
2+
в крови, естественно, приводит к ослаб-
лению сокращений сердца. При отсутствии
Са
2+
в окружающей кардиомиоцит среде со-
кращение невозможно, хотя ПД при этом
практически не изменяется, так как по
Na/Ca-каналам в отсутствие ионов Са
2+
в
клетку входит больше ионов Na
+
. В обычных
(нормальных) условиях расслабление мио-
карда после его сокращения обеспечивается
вследствие выброса Са
2+
из клетки и погло-
щения его саркоплазматическим ретикулу-
мом в результате работы Са-насоса.
Б.
Растяжимость и эластичность сердеч-
ной мышцы также играют важную роль в вы-
полнении нагнетательной функции сердца.
Во-первых, оба этих свойства смягчают
гидравлический удар, возникающий в резуль-
тате несжимаемости жидкости и быстро со-
кращающихся стенок сердца.
Во-вторых, эластические силы, возникаю-
щие вследствие растяжения стенок сердца
при его наполнении кровью, в том числе и
при сокращении предсердий, увеличивают
силу сокращений миокарда в начале систолы.
В-третьих, эластичность структурных эле-
ментов сердца обеспечивает возникновение в
конце систолы упругих сил, способствующих
расслаблению сократившегося (сжавшегося)
миокарда после прекращения его возбужде-
ния. Стенки (особенно желудочков) расслаб-
ляются — расправляются, как предваритель-
но сдавленная резиновая груша, так как раз-
личное направление отдельных мышечных
слоев друг относительно друга увеличивает
упругость стенок сердца, которая превышает
упругость скелетной мышцы.
13.4.
ОСОБЕННОСТИ ЭНЕРГЕТИЧЕСКОГО
ОБЕСПЕЧЕНИЯ СЕРДЕЧНОЙ МЫШЦЫ
Главным источником энергии для сердца явля-
ется процесс аэробного окисления. Анаэроб-
ное окисление (анаэробный гликолиз) для
сердца в отличие от скелетной мышцы играет
незначительную роль. Потенциальными но-
сителями энергии являются главным образом
неуглеводные субстраты. Это свободные
жирные кислоты и молочная кислота (около
60 %), пировиноградная кислота, кетоновые
тела и аминокислоты (менее 10 %). Только
около 30 % расходуемой сердцем энергии по-
крывается за счет глюкозы; при физической
нагрузке увеличивается энергетическая доля
жирных и молочной кислот при одновремен-
ном снижении энергетической доли глюко-
зы.
Таким образом, сердце утилизирует недо-
окисленные продукты, накапливающиеся в
результате интенсивной мышечной работы, и
тем самым препятствует закислению внут-
ренней среды организма.
Большая зависимость деятельности сердеч-
ной мышцы от аэробного окисления делает
сердце весьма зависимым от поступления кис-
лорода к кардиомиоцитам. Сердце массой 300 г
потребляет около 30 мл Ог в минуту, что со-
ставляет 10—12 % от общего количества по-
требляемого организмом кислорода в покое
(масса сердца составляет 0,5 % от массы тела).
На
1
кг массы весь организм потребляет около
4 мл Ог в
1
мин, а сердце
—
около 100 мл, т.е. в
25 раз больше. Скелетная мышца может неко-
торое время работать вообще без кислорода (в
долг) за счет гликолиза. Коэффициент полез-
ного действия сердца в среднем составляет
около 30 %, т.е. несколько больше, нежели
скелетной мышцы,
—
20—25 %.
278

При ухудшении коронарного кровотока и
недостаточном поступлении кислорода к сер-
дечной мышце в ней могут развиваться пато-
логические процессы, вплоть до инфаркта.
Однако это случается относительно редко
благодаря миоглобину, имеющемуся в сердеч-
ной мышце в количестве около 4 мг/г ткани.
Он обладает большим сродством к 0
2
, запа-
сает его во время диастолы и покоя сердца и
отдает во время систолы, когда кровоток в
коронарных артериях желудочков прекраща-
ется: 1 г миоглобина связывает 1,34 мл 0
2
,
что составляет 0,005 мл на 1 г ткани. Этого
количества кислорода хватает для работы
сердца в течение 3—4 с (в норме систола же-
лудочков длится 0,3 с). Даже в случае кратко-
временного спазма коронарных сосудов, на-
пример, при сильных отрицательных эмоци-
ях кислород, связанный с миоглобином,
смягчает трудную для сердца ситуацию.
13.5.
АВТОМАТИЯ СЕРДЦА И АРИТМИЯ
В ЕГО
ДЕЯТЕЛЬНОСТИ
Автоматия сердца — это способность сердца
сокращаться под действием импульсов, воз-
никающих в нем самом. Свойством автома-
там обладают только атипические мышечные
волокна сердца, формирующие его проводя-
щую систему. Клетки рабочего миокарда
автоматией не обладают. Доказательством
автоматии являются ритмические сокраще-
ния изолированного сердца лягушки, поме-
щенного в раствор Рингера. Сердце млекопи-
тающих, помещенное в теплый, снабжаемый
кислородом раствор Рингера, также продол-
жает ритмически сокращаться.
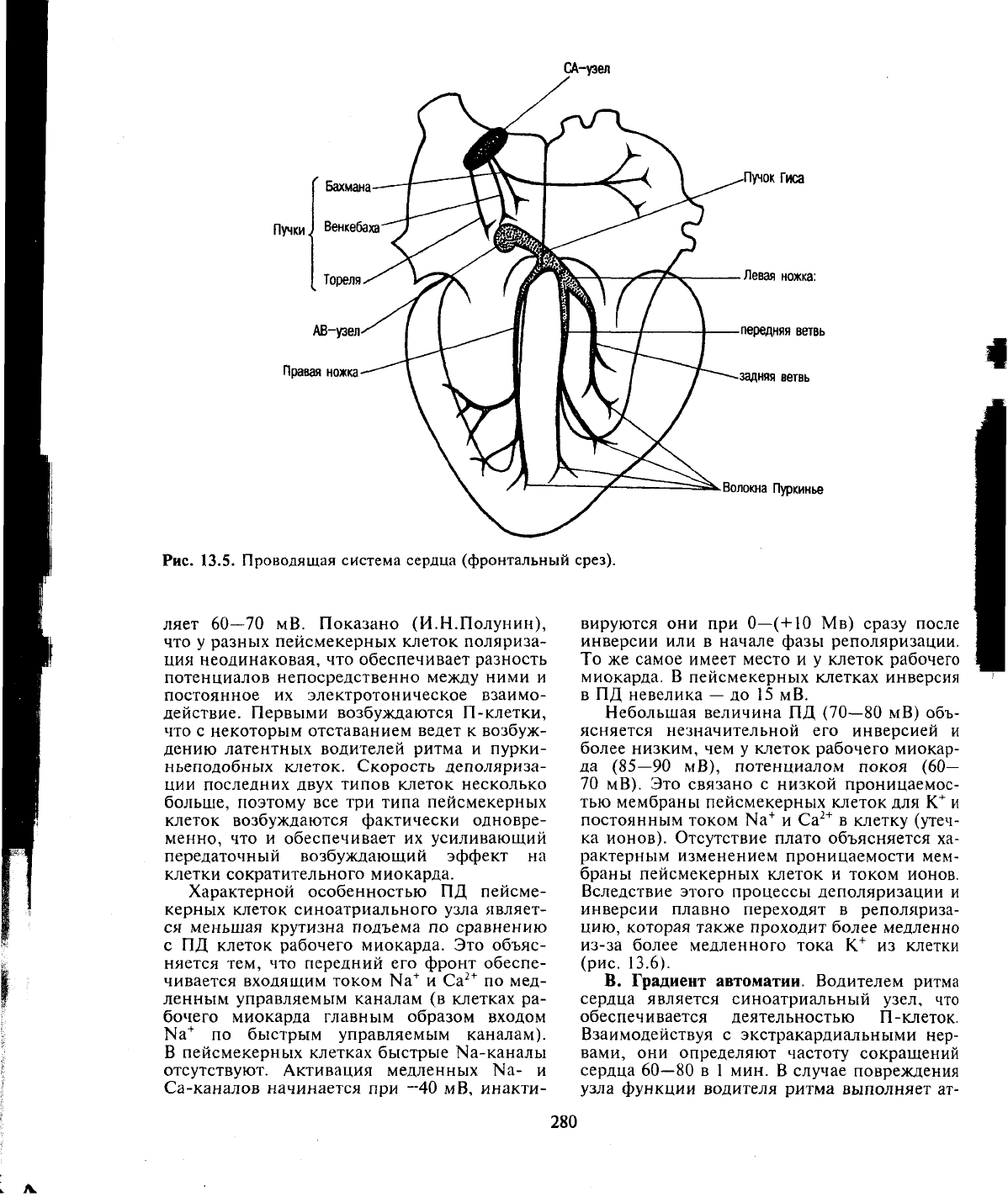
А. Проводящая система сердца имеет в
своем составе узлы, образованные скоплени-
ем атипических мышечных клеток, пучки и
волокна, с помощью которых возбуждение
передается на клетки рабочего миокарда
(рис.
13.5).
Водителем ритма (пейсмекером) сердца яв-
ляется синоатриальный узел, расположенный
в стенке правого предсердия между впадением
в него верхней полой вены и ушком правого
предсердия. В предсердиях имеются также
пучки проводящей системы сердца (Бахмана,
Венкебаха, Тореля и др.), идущие в различных
направлениях. В межпредсердной перегород-
ке у границы с желудочком расположен атри-
овентрикулярный узел, образующий пучок
Гиса — единственный путь, связывающий
предсердия с желудочками. Пучок Гиса делит-
ся на две ножки (левую и правую) с их конеч-
ными разветвлениями — волокнами Пурки-
нье.
В центре синоатриального узла имеется
«плотная зона», содержащая три вида клеток:
П-клетки (истинные водители ритма), латент-
ные водители ритма и пуркиньеподобные
клетки. П-клетки имеются также в атриовент-
рикулярном узле, но в меньшем количестве.
П-клетки содержат мало миофибрилл, имеют
слабо развитую эндоплазматическую сеть и
напоминают малодифференцированные эмб-
риональные кардиомиоциты. Связь П-клеток
с клетками сократительного миокарда осу-
ществляется с помощью промежуточных
Т-клеток. Они близки по структуре клеткам
сократительного миокарда. ПД клеток латент-
ных водителей ритма напоминает таковой
П-клеток. ПД пуркиньеподобной клетки по-
добен таковому клеток сократительного мио-
карда, но в 1,5—2 раза менее продолжителен.
Каждый вид клеток: П-клеток, латентных во-
дителей ритма и пуркиньеподобных — объ-
единен в собственные группы — кластеры.
Клетки Пуркинье обнаруживаются в основ-
ном в ножках пучка Гиса и волокнах Пурки-
нье.
По сравнению с клетками рабочего мио-
карда они содержат меньше митохондрий и
миофибрилл. Эти клетки имеют мало Т-сис-
тем и много вставочных дисков, что обеспечи-
вает малое сопротивление и большую ско-
рость проведения возбуждения.
Б.
Механизм автоматии. Ритмичное воз-
буждение пейсмекерных клеток объясняется
ритмичным спонтанным изменением прони-
цаемости их мембраны для некоторых ионов.
Непосредственные причины следующие.
1.
Медленно увеличивается проницае-
мость мембраны для Na
+
и Са
2+
, вследствие
чего они в большем количестве поступают в
клетку, что ведет к ее деполяризации.
2.
Уменьшение проницаемости мембраны
для К
+
также снижает поляризацию клеток.
3.
Увеличение выхода из клеток СГ со-
гласно электрическому градиенту. Все это ве-
дет к развитию медленной диастолической
деполяризации этих клеток (см. рис. 13.7) и
достижению критического уровня деполяри-
зации (—40—50 мВ), обеспечивающего воз-
никновение ПД и распространение возбуж-
дения — сначала по предсердиям, а затем и
по желудочкам.
Вся восходящая часть ПД клеток-пейсме-
керов обеспечивается входом Na
+
и Са
2+
по
одним и тем же медленным каналам, бы-
стрые Na-каналы в этих клетках отсутствуют.
Достижению критического уровня деполяри-
зации пейсмекерных клеток способствует
также сравнительно небольшая величина
мембранного потенциала в начале медленной
диастолической деполяризации: она состав-
279
СА-узел
Пучки.
Пучок Гиса
Левая ножка:
передняя ветвь
задняя ветвь
Волокна Пуркинье
Рис.
13.5. Проводящая система сердца (фронтальный срез).
ляет 60—70 мВ. Показано (И.Н.Полунин),
что у разных пейсмекерных клеток поляриза-
ция неодинаковая, что обеспечивает разность
потенциалов непосредственно между ними и
постоянное их электротоническое взаимо-
действие. Первыми возбуждаются П-клетки,
что с некоторым отставанием ведет к возбуж-
дению латентных водителей ритма и пурки-
ньеподобных клеток. Скорость деполяриза-
ции последних двух типов клеток несколько
больше, поэтому все три типа пейсмекерных
клеток возбуждаются фактически одновре-
менно, что и обеспечивает их усиливающий
передаточный возбуждающий эффект на
клетки сократительного миокарда.
Характерной особенностью ПД пейсме-
керных клеток синоатриального узла являет-
ся меньшая крутизна подъема по сравнению
с ПД клеток рабочего миокарда. Это объяс-
няется тем, что передний его фронт обеспе-
чивается входящим током Na
+
и Са
2+
по мед-
ленным управляемым каналам (в клетках ра-
бочего миокарда главным образом входом
Na
+
по быстрым управляемым каналам).
В пейсмекерных клетках быстрые Na-каналы
отсутствуют. Активация медленных Na- и
Са-каналов начинается при —40 мВ, инакти-
вируются они при 0—(+10 Мв) сразу после
инверсии или в начале фазы реполяризации.
То же самое имеет место и у клеток рабочего
миокарда. В пейсмекерных клетках инверсия
в ПД невелика
—
до 15 мВ.
Небольшая величина ПД (70—80 мВ) объ-
ясняется незначительной его инверсией и
более низким, чем у клеток рабочего миокар-
да (85—90 мВ), потенциалом покоя (60—
70 мВ). Это связано с низкой проницаемос-
тью мембраны пейсмекерных клеток для К
+
и
постоянным током Na
+
и Са
2+
в клетку (утеч-
ка ионов). Отсутствие плато объясняется ха-
рактерным изменением проницаемости мем-
браны пейсмекерных клеток и током ионов.
Вследствие этого процессы деполяризации и
инверсии плавно переходят в реполяриза-
цию,
которая также проходит более медленно
из-за более медленного тока К
+
из клетки
(рис.
13.6).
В.
Градиент автоматии. Водителем ритма
сердца является синоатриальный узел, что
обеспечивается деятельностью П-клеток.
Взаимодействуя с экстракардиальными нер-
вами, они определяют частоту сокращений
сердца 60—80 в 1 мин. В случае повреждения
узла функции водителя ритма выполняет ат-
280