Смирнов В.М. Физиология человека: Учебник
Подождите немного. Документ загружается.


участок легкого плохо вентилируется, то
кровеносные сосуды в этой области сужают-
ся и даже полностью закрываются. Это осу-
ществляется с помощью механизмов мест-
ной саморегуляции — посредством реакций
гладкой мускулатуры: при снижении в аль-
веолах Р0
2
возникает вазоконстрикция.
В эксперименте она наблюдается уже при
небольшом снижении содержания кислоро-
да (до 15—16 %) в газовой смеси, которой
вентилируют легкое. Вентиляция легкого
азотом также ведет к закрытию капилляров,
вентиляция кислородом ведет к раскрытию
капилляров легких. Эта реакция сохраняется
даже на изолированном легком. При по-
вышении С0
2
также возникает вазокон-
стрикция, а при снижении РС0
2
сужаются
бронхи. Последнее ведет к сохранению
угольной кислоты при гипокапнии. Реакция
усиливается при повышении рН крови. Сле-
дует заметить, что, хотя и существует меха-
низм, обеспечивающий корреляцию крово-
обращения и вентиляции соответствующих
участков легких, он нарушается в результате
механического сдавления сосудов: при вы-
дохе, когда давление в альвеолах повышает-
ся,
кровоток может сильно уменьшиться.
Нередко отмечают, что при выдохе кровоток
в сосудах легких может прекратиться вслед-
ствие повышения давления воздуха в альве-
олах на 1—2 мм рт.ст. Однако это мнение
необоснованно, так как давление в капилля-
рах легких составляет 6—7 мм рт.ст., т.е. в
несколько раз больше давления воздуха в
альвеолах на выдохе.
Из-за того что верхушки легких перерас-
тянуты и по этой причине хуже вентили-
руются, они чаще поражаются туберкулезом.
В нормальных условиях у здорового челове-
ка активно функционирует примерно '/7 аль-
веол, эти активно функционирующие участ-
ки легких непрерывно меняются. Тот факт,
что одновременно функционирует лишь
часть альвеол, весьма важен. В случае пора-
жения части легкого или даже всего легкого
и невозможности излечения терапевтичес-
кими средствами можно одно легкое удалить
полностью. Оставшееся легкое обеспечит га-
зообмен, достаточный для удовлетворитель-
ной жизнедеятельности организма.
В.
Газообмен в легком, естественно, ве-
дет к изменению газового состава в легком
по сравнению с составом атмосферного воз-
духа. В покое человек потребляет около
250 мл 0
2
и выделяет около 230 мл С0
2
.
Поэтому в альвеолярном воздухе уменьшает-
ся количество 0
2
и увеличивается — С0
2
(табл. 12.2).
Таблица 12.2. Состав атмосферного воздуха и
газовой смеси легких (%)
Компонент
0
2
С0
2
Азот и
инертные
газы
Пары воды
Атмосферный
воздух
20,93
0,04
78,5
0,5
Выдыхаемая
смесь газов
16,0
4,0
74,9
5,5
Альвеолярная
смесь газов
14,0
5,5
74,5
5,6
Изменения содержания 0
2
и С0
2
в альвео-
лярной смеси газов являются следствием по-
требления организмом 0
2
и выделения С0
2
.
В выдыхаемом воздухе количество 0
2
не-
сколько возрастает, а С0
2
уменьшается по
сравнению с альвеолярной газовой смесью
вследствие того, что к ней добавляется воздух
воздухоносного пути, не участвующий в газо-
обмене и, естественно, содержащий С0
2
и 0
2
в таких же количествах, как и атмосферный
воздух. Азот в газообмене не участвует, неко-
торое увеличение содержания его в альвео-
лярном воздухе является относительным:
объем выдыхаемого воздуха несколько мень-
ше объема вдыхаемого. Это объясняется тем,
что С0
2
выделяется из организма несколько
меньше, нежели потребляется 0
2
, из-за раз-
личного содержания углерода и кислорода в
различных окисляемых веществах организма.
На долю инертных газов в атмосферном воз-
духе приходится около 1 %. Кровь, обога-
щенная 0
2
и отдавшая С0
2
, из легких посту-
пает в сердце и с помощью артерий и капил-
ляров распределяется по всему организму, в
различных органах и тканях отдает 0
2
и полу-
чает С0
2
.
12.3.
ТРАНСПОРТ ГАЗОВ КРОВЬЮ
Газы в крови находятся в виде физического
растворения и химической связи. Количество
газа, физически растворенного в крови (g),
можно рассчитать по формуле:
a-VP
8--д—•
где а
—
коэффициент абсорбции
—
количест-
во газа, растворенного в 1 мл жидкости при
температуре 0 "С и атмосферном давлении
760 мм рт.ст. (для плазмы крови а0
2
= 0,024;
аС0
2
= 0,49); V
—
объем жидкости; Р
—
пар-
циальное давление газа над жидкостью; А
—
атмосферное давление (760 мм рт.ст.). Если
251

состав жидкости и ее температура постоян-
ны,
что характерно для плазмы крови, то ко-
личество растворенного газа зависит только
от парциального давления. Количество физи-
чески растворенного в крови 0
2
= 0,3 об%;
С0
2
= 4,0 об%; N
2
= 1 об%. Общее содержа-
ние 0
2
и С0
2
в крови во много раз больше,
нежели их физически растворенных фаз
(табл. 12.3).
Таблица 12.3. Общее количество газов в кро-
ви,
об%
(мл в 100 мл крови)
Газ
0
2
С0
2
N
2
Артериальная кровь
19-20
58
1
Венозная кровь
14,5-15,5
54
1
Сравнивая количество растворенных газов
в крови с общим их содержанием, видим, что
0
2
и С0
2
в крови находятся главным образом
в виде химических соединений, с помощью
которых и переносятся.
12.3.1.
ТРАНСПОРТ КИСЛОРОДА
Практически весь 0
2
(около 20 об% — 20 мл
0
2
на 100 мл крови) переносится кровью в
виде химического соединения с гемоглоби-
ном. В виде физического растворения транс-
портируется только 0,3 об%. Однако эта фаза
весьма важна, так как 0
2
из капилляров к
тканям и 0
2
из альвеол в кровь и в эритроцит
проходит через плазму крови в виде физичес-
ки растворенного газа. Если учесть, что чело-
век хорошо переносит чистый 0
2
при давле-
нии до 3 атм, то можно добиться увеличения
физически растворенного 0
2
до 6,84 об%, что
и используется в клинике при оперативных
вмешательствах на сердце или при сердечно-
легочной недостаточности. Но даже дыхание
чистым кислородом с помощью газообмен-
ной маски при обычном для человека давле-
нии в
1
атм увеличивает количество физичес-
ки растворенного 0
2
до 2,28 об% и несколько
улучшает снабжение организма кислородом в
условиях патологии или при подъеме на вы-
соту. При продолжительном же дыхании чис-
тым кислородом у здоровых лиц появляются
боли, особенно при глубоких вдохах, умень-
шается жизненная емкость легких (по-види-
мому, из-за ограничения глубины вдоха);
возможны перевозбуждение ЦНС и возник-
новение судорог, что, очевидно, объясняется
инактивацией некоторых ферментов, в част-
ности дегидрогеназ.
А. Свойства гемоглобина и его содержание
в крови. Этот красный кровяной пигмент, со-
держащийся в эритроцитах как переносчик
0
2
, обладает замечательным свойством при-
соединять 0
2
, когда кровь находится в лег-
ком, и отдавать 0
2
, когда кровь проходит по
капиллярам всех органов и тканей организ-
ма. Гемоглобин является хромопротеидом;
его мол. м. составляет 64 500, он состоит из
четырех одинаковых групп — гемов. Гем
представляет собой протопорфирин, в центре
которого расположен ион двухвалентного
железа, играющего ключевую роль в перено-
се 0
2
. Кислород образует обратимую связь с
гемом, причем валентность железа не изме-
няется. При этом восстановленный гемогло-
бин (НЬ) становится окисленным НЬ0
2
, точ-
нее НЬ(0
2
)
4
. Каждый гем присоединяет по
одной молекуле кислорода, поэтому одна мо-
лекула гемоглобина связывает четыре моле-
кулы 0
2
. Один грамм гемоглобина может
присоединить 1,39 мл 0
2
. Однако при анали-
зе газового состава крови получают несколь-
ко меньшую величину (1,34—1,36 мл 0
2
на
1
г
НЬ).
Это объясняется тем, что небольшая
часть гемоглобина (2—3 %) в физиологичес-
ких условиях не связана с 0
2
, а минимальная
часть оксигемоглобина (около 1 %) не диссо-
циирует (не отдает 0
2
). Содержание гемогло-
бина в крови у мужчин 130—160 г/л, у жен-
щин 120—140 г/л. Количество 0
2
, которое
может быть связано в 100 мл крови, у муж-
чин составляет около 20 мл (20 об%) (кисло-
родная емкость крови), у женщин она на 1—
2 об% меньше. После разрушения старых
эритроцитов в норме и в результате патоло-
гических процессов прекращается и дыха-
тельная функция гемоглобина, поскольку он
частично «теряется» через почки, частично
фагоцитируется клетками мононуклеарной
фагоцитирующей системы.
Гем может подвергаться не только оксиге-
нации, но и истинному окислению. При этом
железо из двухвалентного превращается в
трехвалентное. Окисленный гем носит назва-
ние гематина (метгема), а вся полипептидная
молекула в целом — метгемоглобина. В кро-
ви человека в норме метгемоглобин содер-
жится в незначительных количествах, но при
некоторых заболеваниях, отравлениях неко-
торыми ядами, при действии некоторых ле-
карств, например кодеина, фенацетина, его
содержание увеличивается. Опасность таких
состояний заключается в том, что окислен-
ный гемоглобин очень слабо диссоциирует
(не отдает 0
2
тканям) и, естественно, не мо-
252
жет присоединять дополнительно молекулы
0
2
, т.е. он теряет свойства переносчика кис-
лорода. Так же опасно соединение гемогло-
бина с угарным газом (СО) — карбоксигемо-
глобином, поскольку сродство гемоглобина к
СО в 300 раз больше, чем к кислороду, и
НЬСО диссоциирует в 10 000 раз медленнее,
чем НЬ0
2
. Даже при крайне низких парци-
альных давлениях угарного газа гемоглобин
превращается в карбоксигемоглобин: НЬ +
СО = НЬСО. В норме на долю НЬСО прихо-
дится лишь 1 % общего количества гемогло-
бина крови, у курильщиков — значительно
больше: к вечеру оно достигает 20 %. Если в
воздухе содержится 0,1 % СО, то около 80 %
гемоглобина переходит в карбоксигемогло-
бин и выключается из транспорта 0
2
. Опас-
ность образования большого количества
НЬСО подстерегает пассажиров на автомо-
бильных дорогах. Известно много случаев со
смертельным исходом при включении двига-
теля автомобиля в гараже в холодное время
года с целью обогрева. Первая помощь по-
страдавшему заключается в немедленном
прекращении его контакта с угарным газом.
Специфическое лечение — дыхание чистым
кислородом под повышенным давлением;
поступление кислорода в кровь ускоряет дис-
социацию НЬСО. Таким образом, свойством
гемоглобина как переносчика кислорода яв-
ляется, во-первых, то, что он обладает боль-
шим сродством к 0
2
и быстро соединяется с
ним при увеличении Р0
2
, не изменяя валент-
ности железа; во-вторых, он легко отдает
кислород при уменьшении Р0
2
. Скорость
процесса связывания кислорода гемоглоби-
ном в легких и отдачи его тканям хорошо ил-
люстрирует кривая образования и
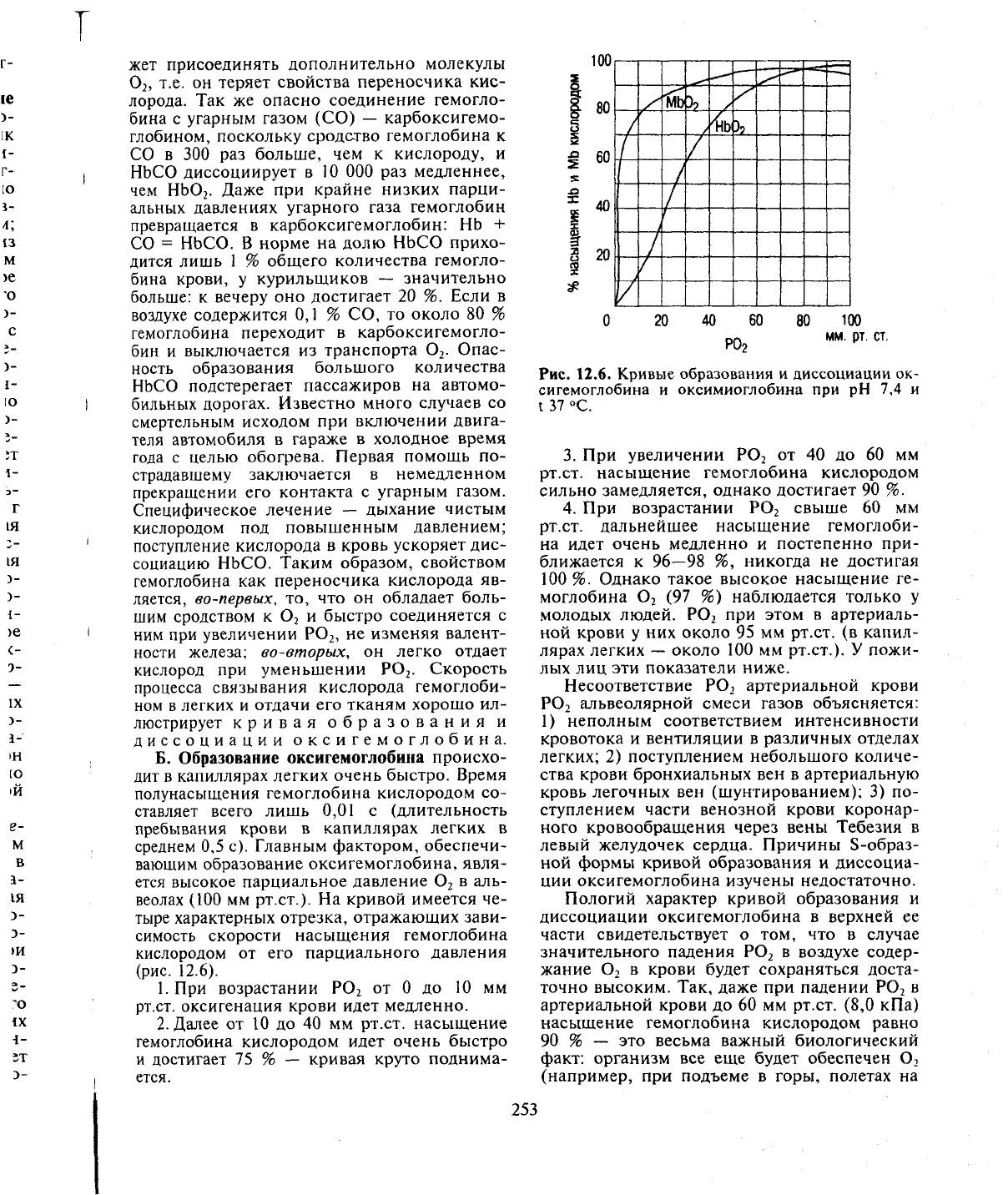
диссоциации оксигемоглобина.
Б.
Образование оксигемоглобина происхо-
дит в капиллярах легких очень быстро. Время
полунасыщения гемоглобина кислородом со-
ставляет всего лишь 0,01 с (длительность
пребывания крови в капиллярах легких в
среднем 0,5 с). Главным фактором, обеспечи-
вающим образование оксигемоглобина, явля-
ется высокое парциальное давление 0
2
в аль-
веолах (100 мм рт.ст.). На кривой имеется че-
тыре характерных отрезка, отражающих зави-
симость скорости насыщения гемоглобина
кислородом от его парциального давления
(рис.
12.6).
1.
При возрастании Р0
2
от 0 до 10 мм
рт.ст. оксигенация крови идет медленно.
2.
Далее от 10 до 40 мм рт.ст. насыщение
гемоглобина кислородом идет очень быстро
и достигает 75 % — кривая круто поднима-
ется.
/
к
/
/
МЬЗг_
'
/
/
г
4
/
/
У
ъъъ^
О
20 40 60 80 100
Р0
2
мм
' Р
т
'
ст
'
Рис. 12.6. Кривые образования и диссоциации ок-
сигемоглобина и оксимиоглобина при рН 7,4 и
t 37 °С.
3.
При увеличении Р0
2
от 40 до 60 мм
рт.ст. насыщение гемоглобина кислородом
сильно замедляется, однако достигает 90 %.
4.
При возрастании Р0
2
свыше 60 мм
рт.ст. дальнейшее насыщение гемоглоби-
на идет очень медленно и постепенно при-
ближается к 96—98 %, никогда не достигая
100 %. Однако такое высокое насыщение ге-
моглобина 0
2
(97 %) наблюдается только у
молодых людей. Р0
2
при этом в артериаль-
ной крови у них около 95 мм рт.ст. (в капил-
лярах легких
—
около 100 мм рт.ст.). У пожи-
лых лиц эти показатели ниже.
Несоответствие Р0
2
артериальной крови
Р0
2
альвеолярной смеси газов объясняется:
1) неполным соответствием интенсивности
кровотока и вентиляции в различных отделах
легких; 2) поступлением небольшого количе-
ства крови бронхиальных вен в артериальную
кровь легочных вен (шунтированием); 3) по-
ступлением части венозной крови коронар-
ного кровообращения через вены Тебезия в
левый желудочек сердца. Причины S-образ-
ной формы кривой образования и диссоциа-
ции оксигемоглобина изучены недостаточно.
Пологий характер кривой образования и
диссоциации оксигемоглобина в верхней ее
части свидетельствует о том, что в случае
значительного падения Р0
2
в воздухе содер-
жание 0
2
в крови будет сохраняться доста-
точно высоким. Так, даже при падении Р0
2
в
артериальной крови до 60 мм рт.ст. (8,0 кПа)
насыщение гемоглобина кислородом равно
90 % — это весьма важный биологический
факт: организм все еще будет обеспечен 0
2
(например, при подъеме в горы, полетах на
253

низких высотах — до 3 км), т.е. имеется вы-
сокая надежность механизмов обеспечения
организма кислородом.
Процесс насыщения гемоглобина кисло-
родом в легких отражает верхняя часть кри-
вой (от 75 до 96—98 %). В венозной крови,
поступающей в капилляры легких, Р0
2
равно
40 мм рт.ст., и достигает оно в артериальной
крови 100 мм рт.ст., как Р0
2
в альвеолах.
Имеется ряд вспомогательных факторов, спо-
собствующих оксигенации крови: 1) отщепле-
ние от карбгемоглобина С0
2
и удаление его
(эффект Вериго); 2) понижение температуры
в легких; 3) увеличение рН крови (эффект
Бора).
Следует также отметить, что с возрас-
том связывание 0
2
гемоглобином ухудша-
ется.
В.
Диссоциация оксигемоглобина происхо-
дит в тканевых капиллярах большого круга
кровообращения (рис. 12.6). При этом гемо-
глобин не только отдает 0
2
тканям, но и при-
соединяет образовавшийся в тканях С0
2
.
Главным фактором, обеспечивающим диссо-
циацию оксигемоглобина, является падение
Р0
2
,
который быстро потребляется тканями.
Обычно отмечают, что этот процесс отражает
нижняя часть кривой.
Однако если учесть, что напряжение Р0
2
в
венозной крови, как правило, не падает ниже
40 мм рт.ст. и что при этом гемоглобин насы-
щен кислородом на 75 % (в этом мнение всех
исследователей едино), то станет очевидным,
что и образование оксигемоглобина в легких,
и диссоциация его в тканях проходят в пре-
делах одного и того же верхнего участка кри-
вой (75—96 % насыщения гемоглобина кис-
лородом). Ведь эритроциты, содержащие ге-
моглобин, находятся в капиллярах, напряже-
ние кислорода в венозной части которых
равно 40 мм рт.ст. По этой же причине, со-
гласно расчетам, и артериовенозная разница
по кислороду равна не более 4,5 об% (4,5 мл
О
2
/100 мл крови), поскольку падение напря-
жения Р0
2
до 40 мм рт.ст. в венозной крови
ведет к уменьшению насыщения гемоглобина
кислородом с 97 до 75 %, т.е. снижение при-
мерно на 22 %.
В межклеточной жидкости Р0
2
уменьша-
ется до
5—20
мм рт.ст., а в клетках падает до
1 мм рт.ст. и меньше (когда Р0
2
в клетке ста-
новится равным 0,1 мм рт.ст., клетка погиба-
ет).
Поскольку возникает большой градиент
Р0
2
(в пришедшей артериальной крови оно
около 95 мм рт.ст.), диссоциация оксигемо-
глобина идет быстро и 0
2
переходит из ка-
пилляров в ткань. Длительность полудиссо-
циации равна 0,02 с (время прохождения
каждого эритроцита через капилляры боль-
шого круга — около 2,5 с), что достаточно для
отщепления 0
2
(огромный запас времени).
Весьма важно, что средняя часть кривой
имеет крутой наклон. В случае увеличения
потребности в кислороде отдельных органов
или организма в целом и увеличения расхода
0
2
даже небольшой сдвиг напряжения 0
2
ниже 40 мм рт.ст. ведет к значительному ус-
корению диссоциации оксигемоглобина и
обеспечению кислородом интенсивно рабо-
тающих органов. Если напряжение кислоро-
да в венозной крови падает лишь на 5 мм
рт.ст. (0,7 кПа), то насыщение гемоглобина
кислородом снижается на 7 %, высвобождаю-
щийся при этом 0
2
может быть сразу же ис-
пользован для процессов метаболизма.
Кроме главного фактора (градиента Р0
2
),
имеется и ряд вспомогательных факторов,
способствующих диссоциации оксигемогло-
бина в тканях. К ним относятся: 1) накопле-
ние С0
2
в тканях; 2) закисление среды; 3) по-
вышение температуры. Ускорение диссоциа-
ции оксигемоглобина при накоплении С0
2
и
Н
+
-ионов в крови объясняется конформаци-
онными изменениями белковой части моле-
кулы гемоглобина. Влияние РС0
2
на связь ге-
моглобина с кислородом открыл отечествен-
ный физиолог Б.Ф.Вериго в 1898 г. (эффект
Вериго); влияние действия рН открыл дат-
ский физиолог Ч.Бор в 1904 г. (эффект Бора).
В зарубежной печати оба эффекта называют
эффектами Бора. Степень выраженности вли-
яния всех факторов (падение Р0
2
и рН, на-
копление С0
2
и повышение температуры ор-
гана) возрастает при увеличении интенсив-
ности обмена веществ в активно работающем
органе. Чем активнее функционирует рабочий
орган, тем, естественно, быстрее потребляется
0
2
и падает его напряжение, быстрее накапли-
вается С0
2
, закисляется среда и повышается
температура ткани вследствие интенсифика-
ции метаболизма — все это ускоряет диссо-
циацию оксигемоглобина и улучшает достав-
ку 0
2
к активно функционирующим органам.
При этом каждые 100 мл крови, содержащие
19—20 мл 0
2
, отдают тканям около 4,5 мл
0
2
— артериовенозная разница по кислороду
составляет 22—23 %. Эта часть 0
2
, поглощае-
мая тканями, называется
коэффициентом
ути-
лизации. Около 15 мл 0
2
остаются в каждых
100 мл венозной крови, возвращающейся
вновь за кислородом в легкие (это резерв ор-
ганизма). Коэффициент утилизации кислоро-
да в разных тканях неодинаков: он колеблется
в пределах 20—30 %; в миокарде, сером веще-
стве мозга, печени достигает 50—60 %.
Таким образом, усиление метаболизма
любой ткани ведет к улучшению диссоциа-
254

ции оксигемоглобина. Кроме того, диссоциа-
ции оксигемоглобина способствует 2,3-ди-
фосфоглицерат — промежуточный продукт,
образующийся в эритроцитах при расщепле-
нии глюкозы. При гипоксии его образуется
больше, что улучшает диссоциацию оксиге-
моглобина и обеспечение тканей организма
кислородом. Ускоряет диссоциацию оксиге-
моглобина также и АТФ, но в значительно
меньшей степени, так как 2,3-дифосфоглице-
рата в эритроцитах содержится в 4—5 раз
больше, чем АТФ.
Г.
Миоглобин также присоединяет 0
2
. По
последовательности аминокислот и третич-
ной структуре молекула миоглобина очень
сходна с отдельной субъединицей молекулы
гемоглобина. Однако молекулы миоглобина
не соединяются между собой с образованием
тетрамера, что, по-видимому, объясняет
функциональные особенности связывания
0
2
. Сродство миоглобина к 0
2
больше, чем у
гемоглобина: уже при напряжении Р0
2
3—
4 мм рт.ст. 50 % миоглобина насыщено кис-
лородом, а при 40 мм рт.ст. насыщение до-
стигает 95 %. Однако миоглобин труднее от-
дает кислород. Это своего рода запас 0
2
, ко-
торый составляет 14 % от общего количества
0
2
, содержащегося в организме. Оксимиогло-
бин начинает отдавать кислород только пос-
ле того, как парциальное давление 0
2
падает
ниже 15 мм рт.ст. Благодаря этому он играет
в покоящейся мышце роль кислородного
депо и отдает 0
2
только тогда, когда исчер-
пываются запасы оксигемоглобина: в част-
ности, во время сокращения мышцы крово-
ток в капиллярах может прекращаться в ре-
зультате их сдавливания, мышцы в этот пе-
риод используют запасенный во время рас-
слабления кислород. Это особенно важно для
сердечной мышцы, источником энергии ко-
торой является в основном аэробное окисле-
ние.
В условиях гипоксии содержание мио-
глобина возрастает. Сродство миоглобина к
СО меньше, чем гемоглобина к СО.
12.3.2.
ТРАНСПОРТ УГЛЕКИСЛОГО ГАЗА
Транспорт С0
2
осуществляется кровью в
виде физического растворения и химической
связи, причем С0
2
, как и 0
2
, переносится и
плазмой, и эритроцитами [Сеченов И.М.,
1859].
Однако соотношения фракций С0
2
,
переносимых плазмой и эритроцитами, су-
щественно отличаются от таковых для 0
2
.
Ниже приведены усредненные показатели
содержания С0
2
в крови.
А. Распределение С0
2
в плазме и эритроци-
тах. Большая часть С0
2
транспортируется
плазмой крови, причем около 60 % всего С0
2
находится в виде бикарбоната натрия
(NaHC0
3
, 33 об%), т.е. в виде химической
связи, 4 об% — в виде физически растворен-
ного С0
2
и около 2 об% С0
2
находится в
виде Н
2
С0
3
. Всего в венозной крови содер-
жится 58 об% С0
2
. Несмотря на то что РС0
2
ниже, чем Р0
2
, количество физически рас-
творенного С0
2
в 13 раз больше. Это объяс-
няется гораздо более высоким коэффициен-
том растворимости С0
2
. Содержание С0
2
в
крови, как и 0
2
, в физически растворенном
состоянии относительно невелико, но это со-
стояние играет огромную роль в жизнедея-
тельности организма. Для того чтобы свя-
заться с теми или иными веществами, дыха-
тельные газы сначала должны быть доставле-
ны к ним в физически растворенном виде.
Газообмен между кровью организма и окру-
жающей средой также осуществляется с по-
мощью физически растворенных С0
2
и 0
2
в
плазме крови.
В эритроците С0
2
находится в форме хи-
мических соединений карбгемоглобина
(ННЬС0
2
, около 5 об%) и бикарбоната калия
(КНС0
3
, 14 об%). Несмотря на то что в виде
карбаминовой связи (ННЬС0
2
) находится
только 5,0 об% С0
2
, это соединение обеспе-
чивает выделение из организма до '/3 объема
всего выделяемого через легкие С0
2
. Это
объясняется особенностями соединения
HHbC0
2
(Hb-NH-COOH): оно легко обра-
зуется в эритроцитах крови при прохождении
ее в капиллярах тканей организма и легко
диссоциирует, когда кровь проходит по ка-
пиллярам легких. Практически весь С0
2
,
связанный с гемоглобином, покидает орга-
низм с выдыхаемым воздухом в результате га-
зообмена в легких. Из других химических со-
единений С0
2
освобождается в легких лишь
частично. Углекислый газ, образуемый в ор-
ганизме, выделяется в основном через легкие
(около 98 %) и только 0,5 % — через почки,
около 2 % — через кожу в виде НСО
э
-бикар-
бонатов.
Следует отметить, что некоторое увеличе-
ние содержания С0
2
в крови оказывает бла-
гоприятное влияние на организм: увеличива-
ет кровоснабжение мозга и миокарда, стиму-
лирует процессы биосинтеза и регенерацию
поврежденных тканей. Увеличение содержа-
ния С0
2
в крови стимулирует также сосудо-
двигательный и дыхательный центры.
Б.
Образование соединений углекислого
газа. В результате окислительных процессов
и образования С0
2
его напряжение в клетках
и, естественно, в межклеточных пространст-
вах значительно больше (достигает 60—80 мм
255

рт.ст.),
чем в поступающей к тканям артери-
альной крови (40 мм рт.ст.). Поэтому С0
2
,
согласно градиенту напряжения, из интерс-
тиция переходит через стенку капилляров в
кровь. Небольшая его часть остается в плазме
в виде физического растворения. В плазме
образуется также небольшое количество
Н
2
С0
3
(Н
2
0+С0
2
-> Н
2
С0
3
), но этот процесс
идет очень медленно, так как в плазме крови
нет фермента карбоангидразы, катализирую-
щего образование Н
2
С0
3
.
Карбоангидраза имеется также в лейкоци-
тах и тромбоцитах. С0
2
поступает и в эти
клетки, где также образуются угольная кис-
лота и ионы НСОз- Однако роль этих клеток
в транспорте С0
2
невелика, так как они не
содержат гемоглобина, их число значительно
меньше, нежели эритроцитов, их размеры
очень маленькие (тромбоциты имеют диа-
метр 2—3 мкм, эритроциты — 8 мкм).
Углекислый газ, проникая в эритроцит,
соединяется с водой и образует угольную
кислоту очень быстро (процесс катализирует-
ся содержащейся в эритроците карбоангидра-
зой примерно в 15 тыс. раз). Оксигемогло-
бин, соединенный с ионом калия (КНЬ0
2
), в
тканях с высокой концентрацией С0
2
легко
отдает кислород. В этом состоянии он теряет
ионы калия и легко принимает от угольной
кислоты ионы водорода, превращаясь в ге-
моглобиновую кислоту (ННЬ). В свою оче-
редь ионы калия связываются с ионами гид-
рокарбоната, образуя бикарбонат калия:
КНЬ0
2
+ Н
2
С0
3
(ННЬ + КНСОз + 0
2
). При-
соединяя ион водорода, гемоглобин действует
как буферная система, поэтому большое ко-
личество угольной кислоты может перено-
ситься к легким без значительного измене-
ния рН крови.
Мембрана эритроцитов слабо проницаема
для Na
+
и К
+
, а проникающие ионы транс-
портируются Na/K-помпой обратно: Na
+
—
в
плазму, К
+
— в эритроцит. Основная масса
НС0
3
,
образующихся в эритроцитах, выходит
в плазму, соединяется там с Na
+
, образуя
NaHC0
3
. Выход НСОз из эритроцитов ком-
пенсируется поступлением туда СГ, диффун-
дирующего из плазмы. Ион НСОз
из
эритро-
цита выходит согласно концентрационному
градиенту, а СГ входит в эритроцит согласно
электрическому градиенту, так как в плазме
крови накапливается избыток отрицатель-
ных ионов НСО]. На стенках мембранных
каналов, пропускающих эти ионы в клетку и
из клетки, могут располагаться фиксирован-
ные заряды, способствующие прохождению
ионов с определенным зарядом. Часть С1-
ионов, проникающих в эритроциты, соеди-
няется с освобождающимися ионами калия,
при диссоциации КНС0
3
, образуя KC1. С0
2
связывается с гемоглобином путем непосред-
ственного присоединения к аминогруппам
белкового компонента гемоглобина. При
этом образуется так называемая карбамино-
вая связь:
ННЬ + С0
2
= HHbC0
2
(Hb-NH-COOH-
карбогемоглобин, точнее карбаминогемогло-
бин).
Небольшое количество С0
2
(1—2 %) пере-
носится белками плазмы крови также в виде
карбаминовых соединений.
В.
Диссоциация соединений углекислого
газа. В легких происходят обратные процес-
сы — выделение из организма С0
2
(за сутки
выделяется около 850 г С0
2
). В первую оче-
редь начинается выход в альвеолы физически
растворенного С0
2
из плазмы крови, по-
скольку парциальное давление РС0
2
в альве-
олах (40 мм рт.ст.) ниже, чем в венозной
крови (46 мм рт.ст.). Это ведет к уменьше-
нию напряжения РС0
2
в крови, причем, при-
соединение кислорода к гемоглобину ведет к
уменьшению сродства углекислого газа к ге-
моглобину и расщеплению карбогемоглобина
(эффект Холдена). Общая схема процессов
образования и диссоциации всех соединений
кислорода и углекислого газа, происходящих
во время прохождения крови в капиллярах
тканей и легких, представлена на схеме 12.1.
12.4.
РОЛЬ СИСТЕМЫ ДЫХАНИЯ
В РЕГУЛЯЦИИ рН
Наибольшее количество ионов Н
+
удаляется
из организма в ходе выведения С0
2
. В раз-
личных клетках организма, особенно в эрит-
роцитах, содержится карбоангидраза, под
действием которой из воды и углекислого га-
за образуется угольная кислота. Важную роль
в выведении Н
+
-ионов легкими играет карбо-
ангидраза эритроцитов. Образовавшийся в
тканях С0
2
поступает в кровь, из крови — в
эритроцит, где образуется Н
2
С0
3
. Угольная
кислота транспортируется кровью в виде раз-
личных химических соединений, в легких
вновь превращается в Н
2
С0
3
и распадается
на С0
2
и Н
2
0. Вода выводится из клеток ор-
ганизма в основном почками, а С0
2
— в ос-
новном легкими. В течение 1 мин в покое в
организме образуется около 230 мл С0
2
, что
составляет 15 тыс. ммоль в сутки, а из крови
выводится примерно эквивалентное количе-
ство Н
+
-ионов.
Во время физической работы образуется
значительно больше угольной кислоты. Од-
256

Схема 12.1. Основные процессы газообмена между кровью и тканями (А);
между кровью и альвеолярным воздухом (Б)
Клетки тканей Альвеолярная смесь газов
НСОз
Плазма
нако существенных сдвигов рН внутренней
среды организма не происходит, что достига-
ется за счет буферных систем крови, возник-
новения гиперпноэ и усиленного удаления
угольной кислоты. Следует заметить, что бу-
ферные системы не обеспечивают выделения
кислых или щелочных ионов из организма.
Они только уменьшают (сглаживают) колеба-
ния рН внутренней среды в различных усло-
виях. Человек этого обычно не замечает.
У здорового человека ацидоз (сдвиг рН в
кислую сторону) и алкалоз (сдвиг рН в сторо-
ну оснований) могут иметь как дыхательную,
так не дыхательную природу. Дыхательный
ацидоз может развиваться при длительном
вдыхании воздуха с повышенным содержани-
ем углекислого газа, например, в замкнутых
помещениях небольшого объема. Дыхатель-
ный алкалоз может развиться в результате
длительного гиперпноэ, когда возникает гор-
ная болезнь и из организма вымывается
большое количество угольной кислоты. Не
дыхательные
ацидоз или алкалоз у здорового
человека развиваются редко — при избыточ-
ном потреблении с пищей кислых или ще-
лочных пищевых продуктов. Алкалоз может
возникнуть при длительном приеме мине-
ральной щелочной воды. Сдвиги рН, связан-
ные с диетой, во многом корригируются по-
чками.
Ацидоз со сдвигом рН внутренней среды
до 7,0 может привести к тяжелым патологи-
ческим состояниям в виде нарушения работы
сердца (фибрилляция желудочков), ЦНС
(кома),
угнетения дыхания. Чаще это возни-
кает при метаболическом ацидозе как следст-
вие нарушения кровообращения в тканях,
что ведет к развитию избыточного анаэроб-
ного гликолиза и накоплению недоокислен-
ных продуктов — молочной, адениловой кис-
лот. Схема функциональной системы, регу-
лирующей рН крови, представлена на схеме
12.2.
Чем больше нагрузка на организм, тем
больше образуется угольной кислоты и тем
больше ее выводится из организма. Увеличе-
ние выделения С0
2
из организма через лег-
кие обеспечивается увеличением интенсив-
ности дыхания, что осуществляется с помо-
щью регуляторных механизмов.
12.5.
РЕГУЛЯЦИЯ ДЫХАНИЯ
Для дыхания клеток организма необходимо
не только постоянное поступление к ним
кислорода, но и удаление углекислоты, так
как ее накопление привело бы к нарушению
равновесия участвующих в дыхании реакций
и изменениям рН, которые могли бы нару-
шить ферментативные процессы. Организм
осуществляет тонкое регулирование напря-
жения 0
2
и С0
2
в крови: их содержание оста-
ется относительно постоянным, несмотря на
колебания количества доступного кислорода
и потребности в нем, которая во время ин-
тенсивной мышечной работы может увеличи-
ваться в 20 раз. Увеличение потребления 0
2
257
9-5484

Схема 12.2. Функциональная система, поддерживающая оптимальную
для метаболизма активную реакцию крови (по К.В.Судакову, с изменениями)
Поведение
в
экстремальных
условиях
Почки
Легкие
Кожа
ЖКТ
—
/
\
Метаболиты
(рН
крови
]
1
о
Обратная связь
Буферные
системы
обеспечивает организм освобождающейся
при этом энергией. Во всех случаях регуля-
ция интенсивности дыхания направлена на
конечный приспособительный результат —
оптимизацию газового состава внутренней
среды организма.
Частота и глубина дыхания регулируются
дыхательным центром, нейроны которого
расположены в различных отделах ЦНС;
главными из них являются продолговатый
мозг и мост. Дыхательный центр по соответ-
ствующим нервам ритмично посылает к диа-
фрагме и межреберным мышцам импульсы,
которые вызывают дыхательные движения.
В основе своей ритм дыхания является не-
произвольным, но может изменяться в неко-
торых пределах и произвольно корой боль-
шого мозга.
12.5.1.
ХАРАКТЕРИСТИКА ДЫХАТЕЛЬНОГО
ЦЕНТРА
А. Локализация отдельных структурных эле-
ментов дыхательного центра установлена с
помощью перерезок ствола мозга на различ-
ных уровнях, разрушения и раздражения раз-
личных структур мозга, регистрации его
электрической активности. Дыхательный
центр, как и любой другой, представляет
собой совокупность нейронов, расположен-
ных на различных уровнях ЦНС.
В продолговатом мозге нахо-
дится главная часть дыхательного центра. Об
этом свидетельствуют исследования М.Флу-
ранса (1794—1867). Он обнаружил, что разру-
шение медиальной части продолговатого
мозга в нижнем углу ромбовидной ямки
ведет к полной остановке дыхания. Позже
Н.А.Миславский (1885) установил наличие
двух структур, ответственных за вдох и
выдох.
Мост играет важную роль в регуляции
продолжительности фаз вдоха, выдоха и
паузы между ними. Нейроны моста при взаи-
модействии с нейронами продолговатого
мозга обеспечивают нормальный цикл дыха-
ния.
Мотонейроны спинного м о з-
г а получают импульсы от нейронов продол-
говатого мозга и посылают их к дыхательным
мышцам по диафрагмальному и межребер-
ным нервам. Центр диафрагмальных нервов
находится в основном в
3—4-м
шейных сег-
ментах спинного мозга. Центры межребер-
ных нервов, иннервирующих мускулатуру
грудной клетки, локализуются в грудном от-
деле спинного мозга (4—10-й сегменты), ин-
нервация мышц живота осуществляется
Th|
V
—Lm-сегментами.
В регуляции дыхания принимают участие
также средний мозг, гипотала-
мус,
лимбик о-р етикулярный
комплекс, кора большого моз-
га. В частности, средний мозг играет важ-
ную роль в регуляции тонуса всей мускула-
туры организма, в том числе и дыхательной.
Гипоталамус выполняет интегрирующую
роль в вегетативном обеспечении сомати-
ческой деятельности, в том числе участвует
в регуляции частоты и глубины дыхания при
физической деятельности, повышении тем-
пературы внешней и внутренней среды (теп-
ловая одышка).
Об участии коры большого мозга в регуля-
ции дыхания свидетельствует тот факт, что
частоту и глубину дыхания можно изменять
произвольно в широком диапазоне. Но про-
258

извольная задержка дыхания не может быть
длительной — наступает непреодолимая по-
требность возобновить дыхание. О роли коры
мозга свидетельствует также усиление дыха-
ния перед стартом или по любому условно-
рефлекторному сигналу. Минимальная физи-
ческая нагрузка (несколько шагов в течение
1—2 мин) бескоркового животного в экспе-
рименте вызывает у него длительную одыш-
ку. Благодаря коре большого мозга при вы-
полнении физических упражнений интен-
сивность дыхания становится адекватной по-
требностям организма (более экономное ды-
хание). Это связано также и с тем, что сами
движения становятся более экономными.
Б.
Автоматия дыхательного центра. В ос-
нове автоматии дыхательного центра лежит
циркуляция возбуждения в его нейронах,
обеспечивающая саморегуляцию вдоха и вы-
доха. Проявления автоматии дыхательного
центра впервые наблюдал с помощью гальва-
нометра на изолированном продолговатом
мозге лягушки И.М.Сеченов (1882). Эдриан
(1931) впервые зарегистрировал ритмическую
активность изолированного продолговатого
мозга золотой рыбки. С помощью микро-
электродной техники подтверждено, что про-
долговатый мозг способен самопроизвольно
генерировать электрические импульсы. Ос-
новная часть нейронов дыхательного центра
в продолговатом мозге относится к ретику-
лярной формации, а эти нейроны обладают
спонтанной активностью. Кроме спонтанной
активности, автоматии дыхательного центра
способствуют гуморальные влияния (глав-
ным образом С0
2
в крови), оказываемые не-
посредственно на клетки центра, афферент-
ная импульсация от хемо- и механорецепто-
ров рефлексогенных зон, реципрокные отно-
шения нейронов дыхательного центра, воз-
буждающие влияние вышележащих отделов
ЦНС.
В.
Нейронная организация дыхательного
центра (продолговатый мозг и мост). Дыха-
тельные нейроны (клетки, которые возбужда-
ются в различных фазах дыхательного цикла)
обнаружены почти на всем протяжении про-
долговатого мозга. Однако в обеих половинах
продолговатого мозга есть участки ретику-
лярной формации, где имеются скопления
дыхательных нейронов. В правой и левой по-
ловинах продолговатого мозга имеется по два
таких скопления
—
дорсальное и вентральное
[Баумгартен Р., 1956]. Они локализуются
вблизи задвижки (obex), которая расположе-
на у нижнего угла ромбовидной ямки.
Дорсальная группа дыхательных нейронов
примыкает к ядру одиночного пучка и состо-
ит примерно на 90 % из инспираторных ней-
ронов (нейронов, возбуждающихся в фазе
вдоха, условно — центра вдоха). Аксоны этих
нейронов идут к другим нейронам дыхатель-
ного центра и к мотонейронам диафрагмаль-
ного нерва в передних рогах шейного отдела
(главным образом сегменты 2—4). Нейроны
диафрагмального ядра спинного мозга воз-
буждаются непрерывно, но с учащением в
фазе вдоха или залпами, как и связанные с
ними нейроны продолговатого мозга. Колла-
терали от аксонов нейронов дорсального ды-
хательного ядра идут также в вентральное
дыхательное ядро продолговатого мозга, об-
разуя возбуждающие синапсы на его инспи-
раторных нейронах и тормозные — на экспи-
раторных. Экспираторные нейроны в дор-
сальном ядре встречаются редко (их несколь-
ко процентов).
Вентральная группа дыхательных
нейронов
расположена вблизи обоюдного, ретроамби-
гуального ядер и простирается до 2-го шей-
ного сегмента спинного мозга включительно.
В вентральной группе содержатся инспира-
торные и экспираторные нейроны (послед-
них большинство). Экспираторные нейроны
расположены между обеими зонами инспи-
раторных клеток, а также ростральнее, в об-
ласти заднего ядра лицевого нерва. Часть
нейронов вентральной группы посылает свои
аксоны в спинной мозг к мотонейронам
межреберных мышц и мышц живота, часть
—
к ядру диафрагмального нерва, часть — к
другим нейронам дыхательного центра. Ин-
спираторные нейроны в спинном мозге рас-
положены в основном в Th„_vi, а экспира-
торные — в Thvin—x- В вентральной группе
находятся эфферентные нейроны центров
блуждающего нерва, регулирующие просвет
воздухоносного пути в ритме дыхательного
цикла. Максимум активности этих нейронов
регистрируется в конце выдоха, что ведет к
сужению просвета воздухоносного пути в ре-
зультате повышения тонуса гладких мышц и
способствует выдоху; минимум активности
нейронов наблюдается в конце вдоха, что со-
провождается уменьшением тонуса гладких
мышц воздухоносного пути, расширяет его и
облегчает вдох.
12.5.2.
САМОРЕГУЛЯЦИЯ ВДОХА И ВЫДОХА
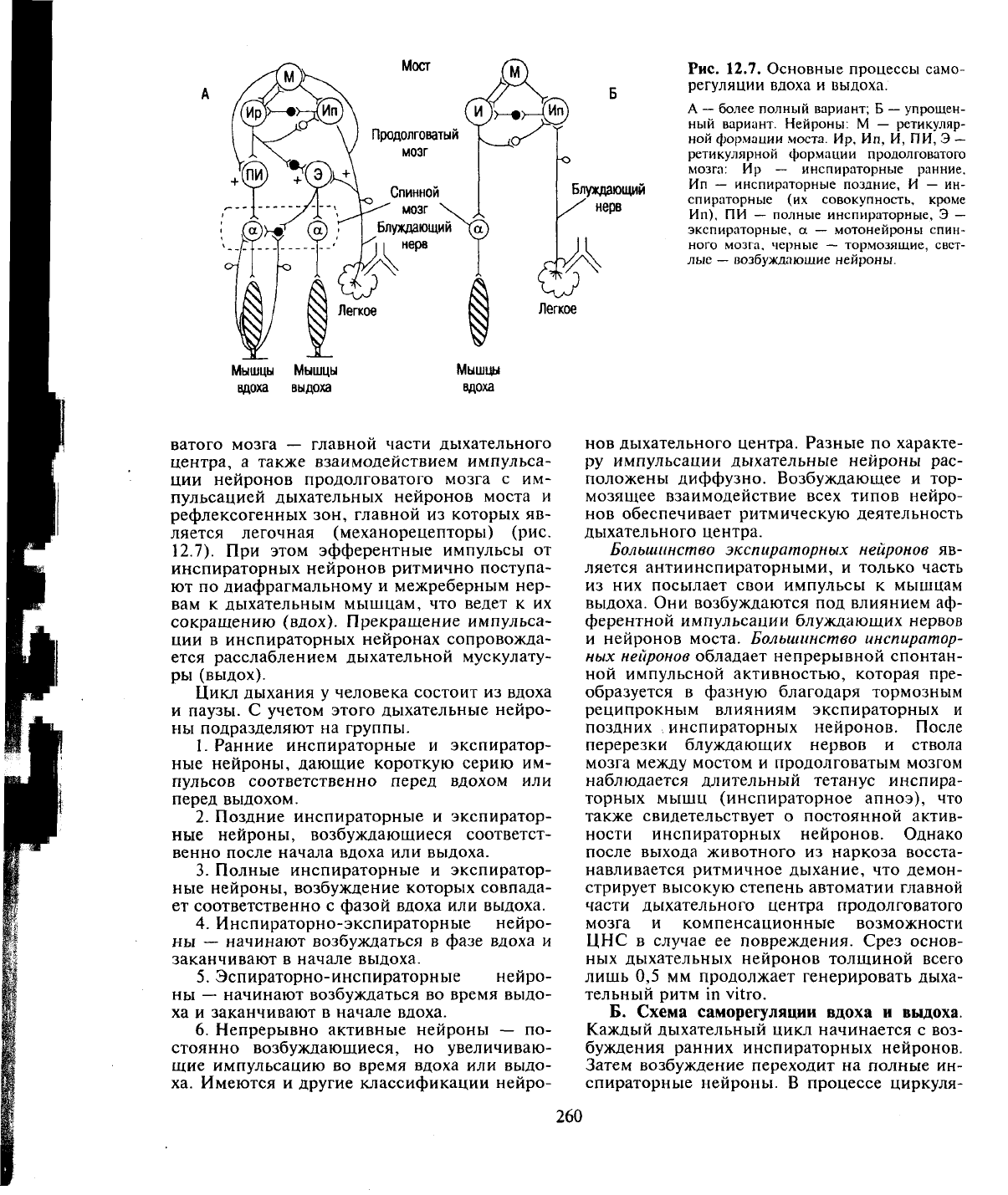
А. Взаимодействие нейронов дыхательного
центра заключается в следующем. Ритмичес-
кая смена вдоха и выдоха (постоянное их че-
редование) обеспечивается циркуляцией воз-
буждения в дыхательных нейронах продолго-
9"
мост
Продолговатый
мозг
Спинной
мозг
Блуждающий
^(а
нерв
Блуждающий
нерв
Легкое
Рис.
12.7. Основные процессы само-
регуляции вдоха и выдоха.
А
—
более полный вариант; Б
—
упрощен-
ный вариант. Нейроны: М — ретикуляр-
ной формации моста. Ир, Ип, И, ПИ, Э
—
ретикулярной формации продолговатого
мозга: Ир — инспираторные ранние.
Ип — инспираторные поздние, И — ин-
спираторные (их совокупность, кроме
Ип),
ПИ — полные инспираторные, Э
—
экспираторные, а — мотонейроны спин-
ного мозга, черные — тормозящие, свет-
лые
—
возбуждающие нейроны.
Мышцы Мышцы
вдоха выдоха
Мышцы
вдоха
ватого мозга — главной части дыхательного
центра, а также взаимодействием импульса-
ции нейронов продолговатого мозга с им-
пульсацией дыхательных нейронов моста и
рефлексогенных зон, главной из которых яв-
ляется легочная (механорецепторы) (рис.
12.7).
При этом эфферентные импульсы от
инспираторных нейронов ритмично поступа-
ют по диафрагмальному и межреберным нер-
вам к дыхательным мышцам, что ведет к их
сокращению (вдох). Прекращение импульса-
ции в инспираторных нейронах сопровожда-
ется расслаблением дыхательной мускулату-
ры (выдох).
Цикл дыхания у человека состоит из вдоха
и паузы. С учетом этого дыхательные нейро-
ны подразделяют на группы.
1.
Ранние инспираторные и экспиратор-
ные нейроны, дающие короткую серию им-
пульсов соответственно перед вдохом или
перед выдохом.
2.
Поздние инспираторные и экспиратор-
ные нейроны, возбуждающиеся соответст-
венно после начала вдоха или выдоха.
3.
Полные инспираторные и экспиратор-
ные нейроны, возбуждение которых совпада-
ет соответственно с фазой вдоха или выдоха.
4.
Инспираторно-экспираторные нейро-
ны — начинают возбуждаться в фазе вдоха и
заканчивают в начале выдоха.
5.
Эспираторно-инспираторные нейро-
ны — начинают возбуждаться во время выдо-
ха и заканчивают в начале вдоха.
6. Непрерывно активные нейроны — по-
стоянно возбуждающиеся, но увеличиваю-
щие импульсацию во время вдоха или выдо-
ха. Имеются и другие классификации нейро-
нов дыхательного центра. Разные по характе-
ру импульсации дыхательные нейроны рас-
положены диффузно. Возбуждающее и тор-
мозящее взаимодействие всех типов нейро-
нов обеспечивает ритмическую деятельность
дыхательного центра.
Большинство экспираторных нейронов яв-
ляется антиинспираторными, и только часть
из них посылает свои импульсы к мышцам
выдоха. Они возбуждаются под влиянием аф-
ферентной импульсации блуждающих нервов
и нейронов моста. Большинство инспиратор-
ных нейронов обладает непрерывной спонтан-
ной импульсной активностью, которая пре-
образуется в фазную благодаря тормозным
реципрокным влияниям экспираторных и
поздних инспираторных нейронов. После
перерезки блуждающих нервов и ствола
мозга между мостом и продолговатым мозгом
наблюдается длительный тетанус инспира-
торных мышц (инспираторное апноэ), что
также свидетельствует о постоянной актив-
ности инспираторных нейронов. Однако
после выхода животного из наркоза восста-
навливается ритмичное дыхание, что демон-
стрирует высокую степень автоматии главной
части дыхательного центра продолговатого
мозга и компенсационные возможности
ЦНС в случае ее повреждения. Срез основ-
ных дыхательных нейронов толщиной всего
лишь 0,5 мм продолжает генерировать дыха-
тельный ритм in vitro.
Б.
Схема саморегуляции вдоха и выдоха.
Каждый дыхательный цикл начинается с воз-
буждения ранних инспираторных нейронов.
Затем возбуждение переходит на полные ин-
спираторные нейроны. В процессе циркуля-
260