Шендрик А.Н. Химия белка. Структура, свойства, методы исследования
Подождите немного. Документ загружается.

92
зина. Если из реакционной смеси удалить галоген расщепление пептидной свя-
зи не идет. Предполагается, что механизм процесса аналогичен другим реакци-
ям с использованием галогенирующих агентов (в том числе N-
бромсукцинимида), а окислительное галогенирование - есть свойство системы:
о-иодбензойная к-та : 4 М солянокислый гуанидин. Уменьшить вероятность
расщепления по тирозину удается добавками п-крезола. В этом случае метод
применим для избирательного расщепления по триптофану.
5.
Известны другие методы расщепления по триптофану. Среди них - рас-
щепление активным иодом. Последний генерируют в системе: пероксид водо-
рода - соли HJ - лактопероксидаза (или пероксидаза хрена). Выход 30-40%. С
аналогичным выходом идет расщепление под действием иодирующих агентов
J
2
, J
3
-
, JCl, смеси: (хлорамин Т)/KJ.
Расщепление по карбоксильной группе триптофана в кислой среде можно
осуществить под действием N-хлорсукцинимида. Остальные связи не затраги-
ваются, в том числе и по тирозину или гистидину. Выходы и скорости реакций
меньше, чем с N-бромсукцинимидом.
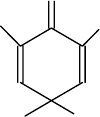
Избирательно, пептидные связи триптофана в белках и пептидах, можно
расщепить с помощью 2,4,6-трибромо-4-метилциклогексадиенона (трибромкре-
зола):
Пептидные связи тирозина и гистидина не затрагиваются. Однако, тиро-
зин превращается в 3,5-дибромпроизводное, цистеин окисляется до цистеино-
вой кислоты, метионин - в метионин-S-оксид и идет модификация гистидина.
O
BrBr
BrCH
3
93
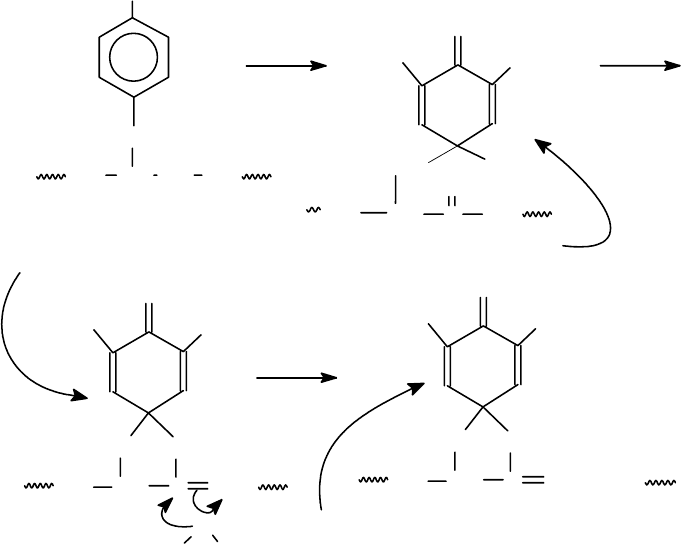
Расщепление по тирозину. При отсутствии в молекуле белка остатков
триптофана избирательное расщепление по тирозину можно осуществить с по-
мощью N-бромсукцинимида.
CH
2
CHNH CO NH
OH
O
BrBr
H
2
CO
CH CHN O
H
2
O
+
O
BrBr
H
2
CO
CH CHN
N
+
H
O
Br
BrBr
CH
2
CH
NH
C
O
NH
+3
NBS
O
H
H
H
2
N
+
Трибромдиенон
Спироиминолактон
Дибромдиеноспиролактон
Диеноспиролактон - сильный хромофор с λ
мах
= 260 нм (ε = 10000-11000).
Метод часто применяется для исследования гистонов, которые не содер-
жат остатков триптофана. Выход, обычно неколичественный, но увеличивается
при увеличении избытка реагента.
Условия проведения реакции: 20
0
С, 6 часов, 50% СН
3
СООН, глубина 30-
65%. В присутствии триптофана расщепление по тирозину идет только при
большом избутке NBS.
Расщепление по остатку цистеина. В основе метода избирательного
расщепления пептидной связи по аминогруппе цистеина лежит превращение
SH-групп в тиоциановые группировки. Впервые расщепление по цистеину под
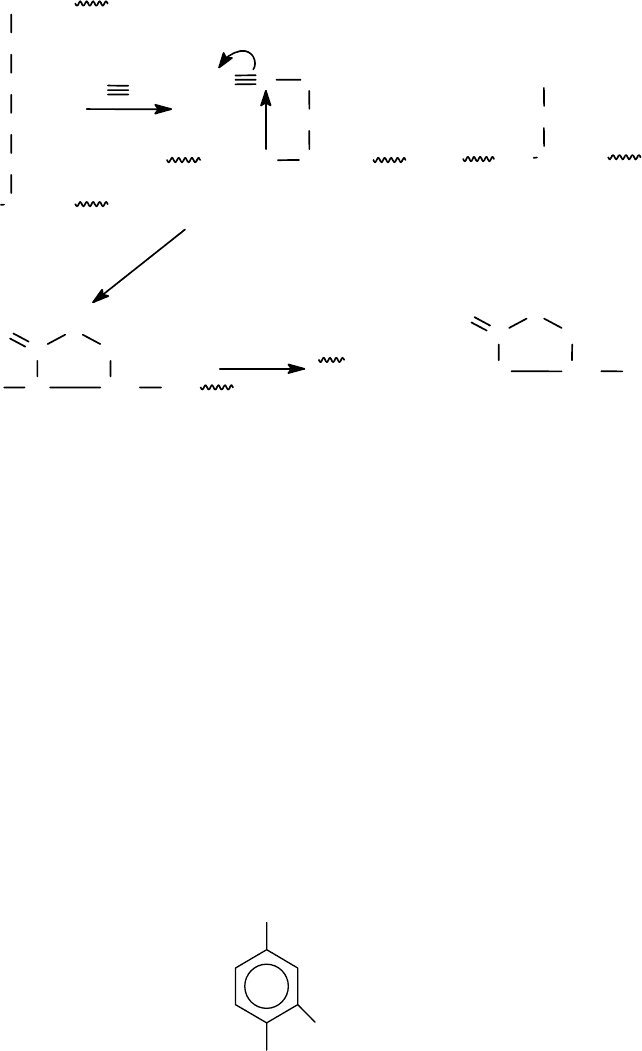
94
действием цианидов было выполнено в 1964-65гг. Механизм реакции представ-
лен ниже на схеме.
C
N
-
NN
CHCO
CH
2
S
S
CH
2
NHCHCO
CH
2
SCN
CHCOOCNH
NN
CHCO
CH
2
S
-
+
C
NCH
CH
2
S
NH
OC CO
H
2
O
COOH
C
HN CH
CH
2
S
NH
CO
Остатки
тиоцианоаланина
Циклизация
Ацилиминотиазолидин
В реакции образуется вначале остатки тиоцианоаланина, которые цикли-
зуются в ацилиминотиазолидин. Затем идет быстрый гидролиз пептидной свя-
зи. Расщепление дисульфидных связей цианидами приводит к частичному и
случайному образованию тиоцианогрупп
Предложен метод прямого и количественного превращения SH-групп в
соответствующие тиоцианопроизводные с помощью 2-нитро-5-
тиоцианобензойной кислоты:
SCN
NO
2
COOH
Показано также, что активным цианирующим агентом SH-групп в белках
является соли 1-циано-4-диметиламино-пиридиния:
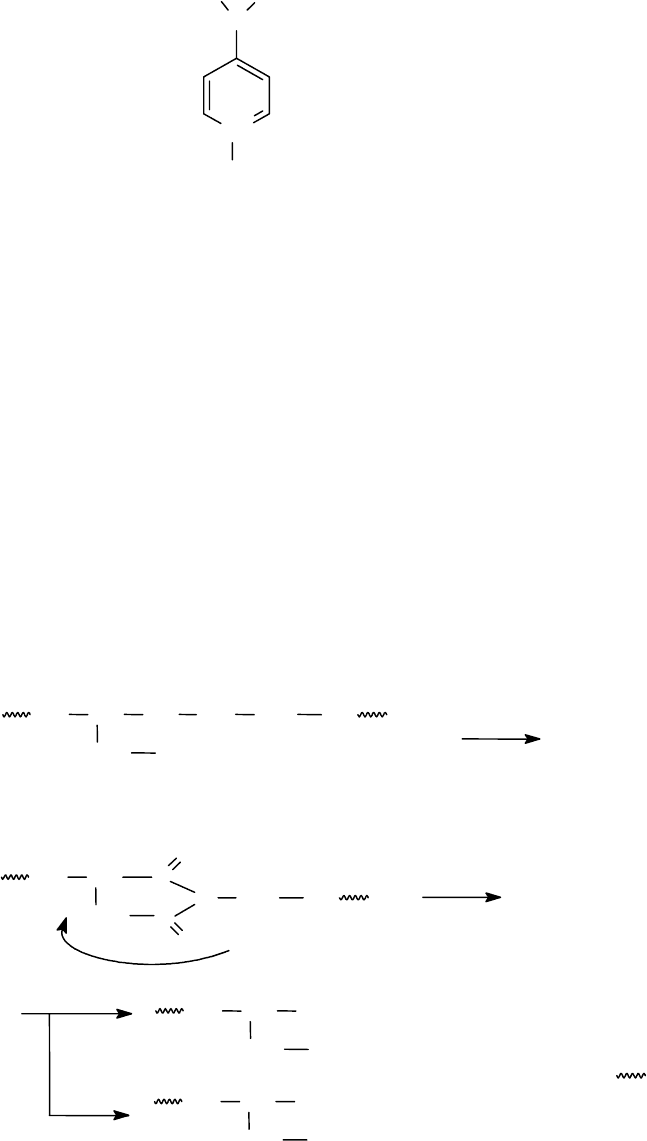
95
N
N
+
CH
3
CH
3
CN
X
-
X = Br
BF
4
ClO
4
Другие химические методы расщепления пептидных связей
Расщепление по связи Асн-Гли гидроксиламином основано на способно-
сти боковой цепи остатка аспарагина циклизоваться с образованием замещен-
ного сукцинимида. Последний подвергается нуклеофильной атаке со стороны
гидроксиламина с расщеплением пептидной связи. Повышенная лабильность
связи Асн-Гли в этой реакции обусловлена, видимо, отсутствием стерических
препятствий циклизации.
Краткая схема механизма реакции выглядит следующим образом:
NH CH CO NH CH
2
CO
CH
2
CONH
2
-
NH
3
NH CH C
CH
2
C
N
O
O
CH
2
CO
NH CH COOH
CH
2
CONHOH
NH CH CONHOH
CH
2
COOH
+
NH
2
CH
2
CO
NH
2
OH
Сукцинимид
Связь Гли-Гли не расщепляется. Условия 20
0
С, 4 часа, рН=9, 6М гуанидин•НСl
+ 2% NH
2
OH•HCl
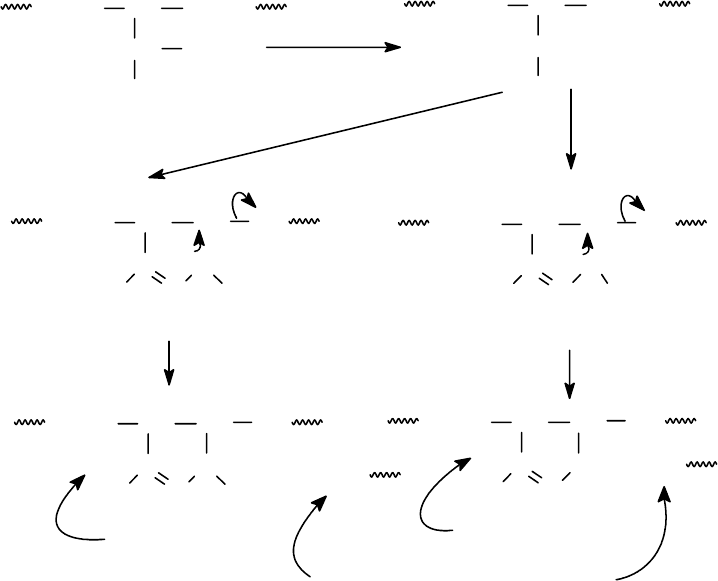
96
Расщепление по связи Асп-Про. Эта связь наиболее лабильна в условиях
кислотного гидролиза и легко расщепляется уже в мягких условиях: 10%
CH
3
COOH + пиридин, рН = 2.5, 7 М солянокислый гуанидин, 40
0
С, 4 суток.
Известен метод расщепления пептидных связей серина и треонина, осно-
ванный на предварительном окислении гидроксильной группы дициклогексил-
карбодиимидом (ДЦК) в присутствии фосфорной кислоты в среде ДМСО и по-
следующей реакции с фенилгидразином. Промежуточный продукт реакции фе-
нилгидразон циклизуется в пиразол с одновременным отщеплением фрагмента
со свободной NH-концевой аминогруппой.
Возможен и другой вариант, когда под действием гидроксиламина обра-
зуется промежуточный гидразон. Он циклизуется в изооксазолин с образовани-
ем фрагмента пептидной цепи со свободной аминогруппой. Схема этих реак-
ций:
OCNH CH CONH
CH OH
R
ДМСО/ДЦК
H
3
PO
4
OCNH CH CONH
CO
R
C
6
H
5
NHNH
2
OCNH CH CO NH
C
RN
NH
C
6
H
5
NH
2
OH
OCNH CH CO NH
C
RN
O
H
Фенилгидразон
Гидразон
OCNH CH CO NH
C
RN
N
C
6
H
5
OCNH CH CO NH
C
RN
O
H
2
N
H
2
N
+
+
Пирозалон
Изооксазолин
Фрагменты со свободной
аминогруппой
R = H, CH
3

97
2.1.5.2 Установление аминокислотной последовательности
фрагментов белковых молекул
Первым белком, для которого была установлена его аминокислотная по-
следовательность был инсулин. На разработку стратегии этого эксперимента и
его реализацию выдающемуся биохимику Ф. Сенгеру (Sanger) потребовалось
несколько лет и в 1951 г. он расшифровал первичную структуру инсулина.
Фермент состоит из двух цепей (21 и 30 аминокислотных остатка), связанных
между собой дисульфидными связями.
В 1958 г Сенгер был удостоен Нобелевской премии
за исследование первичной структуры инсулина. В
1980 г ему была присуждена вторая Нобелевская
премия за установление последовательности нук-
леиновых оснований в полинуклеотидах.
СЕНГЕР (Sanger), Фредерик
род. 13 августа 1918 г.
Нобелевская премия по химии, 1958 г.
Нобелевская премия по химии, 1980 г., совместно с Полом Бергом и Уолтером Гилбертом
Английский биохимик Фредерик Сенгер (Сангер) родился в Рендкомбе (графство Глостершир). Его
мать была дочерью преуспевающего текстильного магната. Отец работал врачом. С 1932 по 1936 г. будущий
ученый обучался в Брайанстонской школе в Блэндфорде (графство Дорсетшир), а в 1936 г. поступил в колледж
св. Иоанна Кембриджского университета. Первоначально С. планировал заняться медициной, но его заинтере-
совала биохимия. «Мне казалось, – писал он много лет спустя, – что это был путь к действительному понима-
нию живой материи и к разработке более научных основ для решения многих проблем, стоящих перед медици-
ной».
В 1939 г. в Кембриджском университете С. получил степень бакалавра естественных наук, а в 1943 г.
докторскую степень. Он вошел в исследовательскую группу, возглавляемую Э.Ч. Чибналлом, который как раз
перед этим сменил Фредерика Хопкинса в должности профессора биохимии Кембриджского университета. В то
время Чибналл занимался исследованиями в области химии белков.
В 1902 г. Эмиль Фишер предположил, что белки состоят из аминокислот, связанных между собой пеп-
тидными связями. К началу 40-х гг. гипотеза Фишера была широко, хотя и не повсеместно признана. Чибналл
предложил С. установить конечную последовательность пептидной цепи химическим путем.
В 1945 г. С. сообщил, что в мягких щелочных условиях динитрофенол может присоединяться к атому
азота аминокислоты благодаря связи более сильной, чем пептидная. Следовательно, белок может быть расщеп-
лен на составляющие его аминокислоты с разрушением пептидных связей, а структуру аминокислот можно ус-
тановить с помощью хроматографии.
Значительная часть исследований, проводимых в лаборатории Чибналла, была связана с инсулином,
одним из немногих белков, доступных в то время в чистом виде и в больших количествах. Первоначальное изу-
чение С. инсулина показало, что он содержит две различные N-концевые аминокислоты, т.е. каждая молекула
инсулина состоит их двух видов полипептидных цепей. Две молекулы цистеина могут соединяться с образова-
нием цистина посредством дисульфидного мостика между двумя полипептидными цепями, либо между раз-
личными участками одной цепи. В 1949 г. С. сообщил, что он открыл способ разрушения этих дисульфидных
мостиков и, следовательно, метод разделения двух цепей.
Сенгер и приехавший из Вены Ганс Туппи разработали план установления последовательности чередо-
вания аминокислот в каждой полипептидной цепи инсулина, используя для этих целей различные ферменты
(протеазы). В 1950 г. Туппи, после установления структуры более длинной из двух инсулиновых цепей, уехал
98
из Кембриджа. Более короткая инсулиновая цепь не так легко поддавалась химическому анализу, и поэтому по-
следовательность чередования в ней аминокислот была полностью установлена только в 1953 г. С. продолжил
работу по установлению местоположения дисульфидных мостиков между двумя цепями, и в 1955 г. представил
законченную структуру молекулы инсулина. Это была первая белковая молекула, так подробно изученная.
Работа С. имела важные последствия для биохимии и зарождающейся науки – молекулярной биологии. Резуль-
таты проведенных им исследований окончательно доказали, что белки состоят из аминокислот, соединенных в
цепи пептидными связями. С. также доказал, что некоторые ферменты могут разрывать пептидные цепи в зара-
нее установленных местах. Применение этого метода помогло биохимикам определить впоследствии структуру
многих других белков.
В 1958 г. С. была присуждена Нобелевская премия по химии «за установление структур белков, осо-
бенно инсулина». В своей Нобелевской лекции С. подчеркнул большое практическое значение проведенной им
работы. «Установление структуры инсулина, безусловно, открывает путь к исследованию других белков, – ска-
зал он. – Можно также надеяться, что изучение белков поможет выявить изменения, которые происходят в ор-
ганизме во время болезни, и что наши усилия могут принести человечеству большую практическую пользу».
Еще до получения Нобелевской премии С. занялся изучением генетики. Отчасти это произошло под
влиянием дружбы ученого с Фрэнсисом Криком. Для С. одним из наиболее поразительных фактов, касающихся
последовательности чередования отдельных групп в инсулине, было явное отсутствие какого бы то ни было
принципа уникального расположения аминокислот. А ведь от этого, казалось бы, случайного порядка зависела
важная физиологическая деятельность. С. не понимал, каким образом белок может соединяться именно в такой
последовательности, однако было очевидно, что у этого порядка должны быть определенные истоки. В середи-
не 50-х гг. Крик (который вместе с Джеймсом Д. Уотсоном первый описал структуру генетического вещества
дезоксирибонуклеиновой кислоты, или ДНК) объяснил сделанные С. открытия, прибегнув к «гипотезе после-
довательности», которая заключалась в том, что информацию, определяющую последовательность аминокис-
лот в белке, несут гены. Позднее было установлено, что сами гены представляют собой последовательность
звеньев, отдельные группы которых соответствуют определенной аминокислоте.
Нуклеиновые кислоты – ДНК и рибонуклеиновая кислота (РНК) – это цепи связанных нуклеотидов.
Записанный в них генетический код для аминокислот определяется последовательностью трех оснований. Про-
цесс строительства белка начинается с того, что соответствующий участок молекулы ДНК, который включает
полные указания для сбора соединения, «расстегивает молнию» для связи, соединяющей основания друг с дру-
гом. Свободные нуклеотиды (как попало плавающие в клетке) оказываются привязанными вдоль открытой для
этого последовательности молекулы ДНК, образуя зеркально отображенную цепь, называемую матричной РНК
(мРНК). Законченная цепь мРНК покидает ДНК (которая тогда снова «закрывает молнию») и продвигается к
клеточным структурам, которые называются биросомами, где и будет собираться белок. Участки более корот-
кой цепи формируются мРНК и затем движутся в сторону, с тем чтобы вобрать в себя соответствующие сво-
бодные нуклеотиды, которые они затем приносят обратно мРНК для включения в белковую структуру. Эти ко-
роткие цепи называются транспортными РНК (тРНК). К тому времени, когда С. приступил к изучению нуклеи-
новых кислот, об этих процессах мало что было известно, а о нуклеотидных последовательностях не было из-
вестно вообще ничего.
Последовательности ДНК и РНК представляют большие трудности для анализа, чем белковые после-
довательности, поскольку они длиннее. Типичная белковая цепь может содержать до пятидесяти аминокислот,
а типичная мРНК содержит сотни нуклеотидов. ДНК даже крошечного вируса состоит из тысяч нуклеотидов. И
тем не менее последовательности нуклеиновых кислот легче поддаются раскодированию, чем белковые после-
довательности, из-за их фундаментального различия: в то время как каждое место в белковой цепи может быть
занято любой из 20 различных аминокислот, существует только 4 «претендента» на каждое место в последова-
тельности ДНК – нуклеотиды, сокращенно называемых А, Т, Ц и Г.
В 1958 г. Роберт У. Холли предпринял попытку установить последовательность цепи тРНК. Несмотря
на то что длина этих коротких цепей не превышает 100 нуклеотидов, эта работа из-за сложности установления
последовательности затянулась до 1965 г. На С. произвела глубокое впечатление работа Холли, но он искал бо-
лее действенный метод установления последовательности, доступный для применения к цепям мРНК, длина
которых нередко достигает нескольких сотен нуклеотидов. В начале 60-х гг. он и его коллеги разработали та-
кую технологию. Применив ферменты, они избирательно расчленили цепи мРНК на более мелкие фрагменты и
проследили последовательность в каждой из них. Затем на основании заключений о взаимоотношении между
фрагментами была определена последовательность во всей цепи.
Такой подход, однако, требовал массы времени и терпения, и С. решил разработать аналитический ме-
тод установления последовательности в ДНК. Он добился этого в 1973 г. Предложенная им процедура заклю-
чалась в том, что двойная цепь молекулы ДНК разбивалась на одинарные цепи (называемые стренгами), а затем
полученный материал группировался в четыре образца. Каждый образец начинают восстанавливать до перво-
начальной последовательности двойной цепи, исходя из шаблона одинарной цепи. Однако исследователи оста-
навливают процесс восстановления на разных нуклеотидах для каждого образца либо путем ограничения кон-
центрации того или иного свободного нуклеотида, либо помещая в цепь определенный нуклеотид с таким хи
99
микатом, который предотвращает дальнейший синтез. В результате этого реконструированные цепи представ-
ляют собой образцы различной длины, но каждая заканчивается одинаковым нуклеотидом. Затем эти четыре
образца одновременно пропускают через фильтрующий материал - сверхтонкий акриламидным гелем, который
разделяет эти цепи в соответствии с их длиной, поскольку более короткие цепи проходят через гель быстрее. И
тогда нуклеотидная последовательность первоначальной цепи ДНК может быть прочитана прямо с геля путем
сравнения следов, оставленных образцами.
В то время как С. м его коллеги работали над этим методом (названным дидекоксидным методом по
типу используемого при этом ограничивающего химиката), американские ученые Уолтер Гилберт и Аллан
Мэксам разрабатывали другую процедуру установления нуклеотидных последовательностей. В соответствии с
их методом фрагменты цепи ДНК различной длины получают, разрывая цепь на специфических основаниях.
Этот подход напоминает метод, который применил С. для установления последовательностей в белковых цепях
и цепях РНК. Как технология С., так и технология Гилберта стали важнейшим инструментом генной инжене-
рии, хотя метод С. несколько более эффективен при работе с очень длинными последовательностями. Еще в
1978 г. С. и его коллеги продемонстрировали действенность дидезоксидного метода, установив последователь-
ность 5375 оснований в цепи ДНК бактериального вируса. Это был первый случай такой подробной расшиф-
ровки цепи ДНК.
В 1980 г. С. и Гилберту была присуждена половина Нобелевской премии по химии «за вклад в уста-
новлении основных последовательностей в нуклеиновых кислотах». Другая половина премии была присуждена
Полу Бергу. Эти трое ученых, сказал в своей вступительной речи от имени Шведской королевской академии
наук Б.Г. Мальстрем, «сделали возможным проникновение в еще большие глубины в нашем понимании взаи-
мосвязи между химической структурой и биохимической функцией генетического материала».
В 1983 г. С. вышел в отставку с занимаемого им поста в Медицинском научно-исследовательском со-
вете. Скромный, склонный к уединению человек, он живет в Кембридже со своей женой Маргарет Джоан Хоув.
Брак с ней был зарегистрирован в 1940 г. У супругов два сына и дочь.
С. удостоен многочисленных наград. Среди них: медаль Кордей-Моргана и премия, присужденные ему
Британским химическим обществом (1951), премия Альфреда Бензонса Фонда Альфреда Бензонса (1966), Ко-
ролевская медаль Лондонского королевского общества (1969), ежегодная награда Гарднеровского фонда (1971
и 1979), памятная медаль Хэнбери Фармацевтического общества Великобритании (1976), медаль Копли Лон-
донского королевского общества (1977) и премия Альберта Ласкера за фундаментальные медицинские иссле-
дования (1979). С. – почетный член Американского общества биохимиков и американской Национальной ака-
демии наук, обладатель почетных степеней университетов Лестера и Страсбурга, а также Кембриджа и Окс-
форда.
Источник информации: Лауреаты Нобелевской премии: Энциклопедия: Пер. с англ.– М.: Прогресс, 1992.
К настоящему времени количество белков, для которых установлена ами-
нокислотная последовательность, исчисляется тысячами и их число очень бы-
стро растет. Одним из выдающихся достижений в этой области было определе-
ние аминокислотной последовательности молекулы антитела гамма-
глобулиновой фракции крови, которая включает 1320 аминокислот. В настоя-
щее время все полученные и получаемые данные о пространственной структуре
белков заносятся в банк данных (см. в сети "Internet": http://nist.rcsb.org/pdb/).
По мере накопления информации этот банк предполагается использовать в бу-
дущем для предсказания форм белковых молекул и их функций исходя из ами-
нокислотной последовательности.
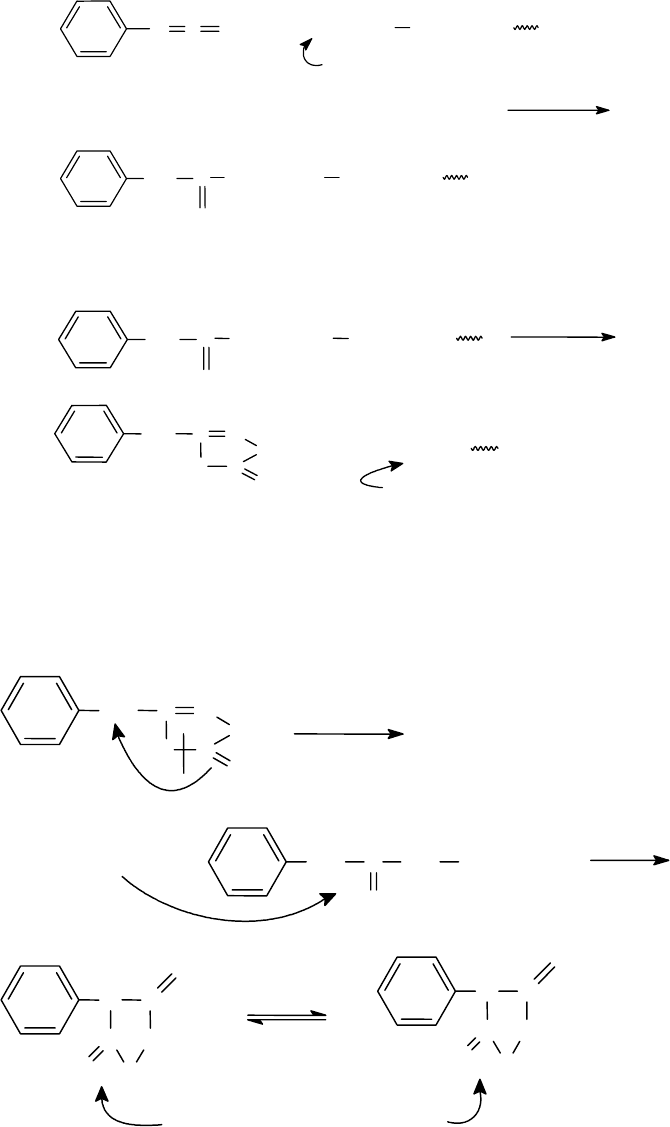
100
В основе методов установления аминокислотной последовательности
пептидов лежит реакция Эдмана (1950-1956г) - это деградация полипептидной
цепи со стороны N-концевой аминокислоты с помощью ФИТЦ.
Каждый цикл деградации включает три стадии.
1.
Образование фенилтиокарбамоилпептида (ФТК-пептида)
NCS
+
NH
2
CHRCO NHCHR'CO
N
-
конец пептидной цепи
HO
-
pH 9-9.5
NH
C
S
NHCHR'CONHCHRCO
2.
Отщепление N-концевой аминоксилоты в форме анилинотиазолинона:
C
S
NHCHR'CONHCHRCO
NH
CF
3
COOH
NH C
S
N
C
CHR
O
+
NH
2
CHR'CO
n-1
Пептидная цепь
3.
Изомеризация тиазолинона в 3-фенил-2-тиогидантоин (ФТГ) и его иден-
тификация. Эта стадия идет через образование фенилтиокарбамоиламинокис-
лоты (ФТК - аминокислота):
NH C
S
N
C
CHR
O
H
2
O,
H
+
NH C
S
NH CHRCOOH
NC
O
CCHR
NH
S
NC
S
CNH
O
CHR
3
-
ф
енил-2-тиоги
д
антоин
ФТК аминокислота
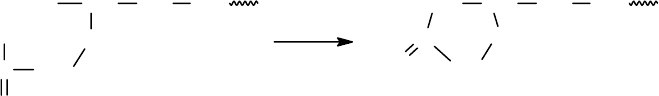
101
Первая стадия - присоединение ФИТЦ к непротонированной α-
аминогруппе концевой аминокислоты. (рН=9-9.5, легко летучие буферные сис-
темы). В качестве оснований можно использовать третичные или гетероцикли-
ческие амины (триэтиламин, диметилаллиламин, пиридин).
Побочные реакции.
¾ Окислительное десульфирование ФТК-группы под действием кислорода
воздуха (все стадии проводят, поэтому, в среде инертного газа).
¾ Гидролиз ФИТЦ с образованием дифенилтиомочевины и анилина и обра-
зование побочных продуктов, которые затруднят идентификацию фенилтиоги-
дантоинов аминокислот. Поэтому по завершению первой стадии экстрагируют
бензолом остаток ФИТЦ и его побочных продуктов.
Вторая стадия - отщепление тиазолинона. Эта реакция идет легко и, как
правило, без образования побочных продуктов. Исключение - концевые остатки
глутамина могут превращаться в остатки пироглутаминовой кислоты, блокируя
тем самым дальнейшую деградацию пептидной цепи:
NH
2
CH
CH
2
CH
2
C
O
NH
2
CO NH
-
NH
3
NH CH
C
CH
2
CH
2
O
CO NH
Вторую стадию проводят в присутствии безводной трифторуксусной кислоты.
Третья стадия - изомеризация. Протекает в два этапа. Вначале проходит
быстрый гидролиз с превращением тиазолинона в ФТК-аминокислоту и даль-
нейшая циклизация в фенилтиогидантоин (ФТГ). Эта реакция протекает в усло-
виях: 1н НСl, 80
0
С, 10 мин. Некоторые ФТГ в этих условиях частично разру-
шаются. Так например, ФТГ серина и треонина подвергаются дегидратации.
На втором этапе добавляют бутилацетат и разделяют фазы. Органическая
фаза содержит ФТГ, водная остаточный пептид с длинной цепи меньшей на
единицу. Водную фазу отделяют и направляют на следующий цикл деградации.