Щербак Г.Й., Царичкова Д.Б., Вервес Ю.Г. Зоологія безхребетних. Книга 1
Подождите немного. Документ загружается.

воді або в гастральній порожнині материнського організму.
Плаваючі личинки -— планули — розносяться морськими
течіями на значні відстані.
Актинії трапляються в усіх морях і океанах — від Арк-
тики до антарктичних вод, але найрізноманітніші вони в
тропіках. У Чорному морі мешкає лише два види: Actino-
thoe clavata та Actinia equina, останній вид знайдено й у
• Азовському морі.
Більшість видів мешкає в прибережному мілководді,
але є й глибоководні форми, деякі з них живуть на макси-
мальних глибинах Світового океану (до 11 км).
Окремі види актиній вступають у мутуалістичні відно-
сини з іншими тваринами. Класичний приклад — це спів-
існування раків-самітників і актиній Adamsia. Актинія
оселяється на черепашці молюска, в якій живе рак-саміт-
ник. Така асоціація є корисною для обох видів. Актинія
^одержує залишки їжі рака-самітника, набуває рухливо-
сті, а рак-самітник перебуває під надійним захистом щу-
палець актинії.
Один із середземноморських крабів постійно тримає в
клешнях двох актиній для захисту від ворогів і захоплен-
ня їжі, яку актинії ловлять своїми щупальцями, а краб
відбирає.
Має місце також симбіоз велетенських актиній Stoi-
chactis, що живуть на західному узбережжі Австралії, з
рибками-ам-фіприонами, які перебувають серед щупалець
актинії, ховаючись від ворогів. У щупальцях виробляється
особлива речовина, що гальмує вистрілювання жалких
капсул. Ця речовина поступово накопичується в слизі, що
вкриває тіло рибок, тому вони після перших, дуже обереж-
них контактів із актинією поступово стають несприйнятли-,
вими до її отрути. Актинії одержують залишки їжі ри-
бок, крім того, риби постійним биттям плавців сприяють
поліпшенню газообміну актинії.
Ряд Склерактинії, або Мадрепорові корали
(Scleractinia, або Madreporaria)
Склерактинії — найчисленніша група коралів, яка на-
лічує, за сучасними відомостями, близько 550 видів. Це пе-
реважно колоніальні, рідше поодинокі корали, що мають
вапняковий скелет ектодермального походження. Скелет у
склерактиній суцільний, масивний.
У поодиноких поліпів скелет утворюється спочатку на
підошві, епідерма якої складається з особливих клітин —
калікобластів (склеробластів). На поверхні цих клітин
160
виділяються найтонші хітинові ниточки, які, переплітаю-
чись між собою, утворюють дуже ніжну щільну сіточку.
На ній і відкладаються кристалики вапна (СаСО
3
), які
зливаються потім у суцільний скелет, що постійно нарос-
тає. Таким чином утворюється підошовна пластинка, з якої
розвиваються скелетні перетинки — склеросепти, що врос-
тають у м'які тканини поліпа. У периферійній області по-
ліпа від підошви відходить невисока чашечка-ге/со, яка
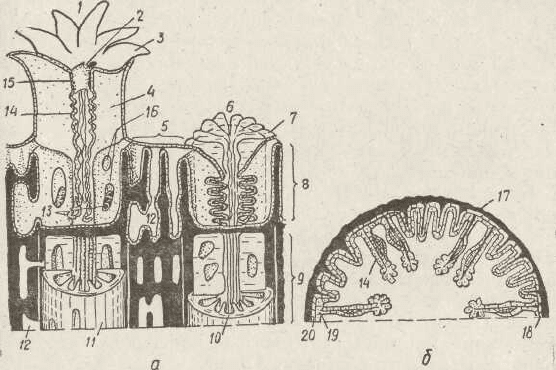
Рис. 118. Схема будови склерактиній — ділянка колонії (а), попереч-
ний зріз'окремого поліпа (б):
/ — розправлений поліп; 2 — рот; 3 —щупальця; 4 — м'яка септа; 5 — ценосарк?
6 — втягнутий поліп; 7 — склеросепта; 8 — м'яке тіло; 9 — відмерлі поліп»
(голий скелет)- 10 — денце; // —кораліт; 12 — порожнина ценосарка; ІЗ —
аконції; 14 — мезентеріальні нитки; 15 — глотка; 16 — отвори у склеросептах;;
17 — склеротика; 18 — мезоглея; 19 — гастродерма; 20 — епідерма
оточує його нижню частину. Так виникає кораліт — скелет
окремого поліпа. М'які тканини опиняються всередині ко-
раліта, вони щільно прилягають до нього та повторюють
його рельєф (рис. 118).
У колонії склерактиній розрізняють окремі поліпи (зор-
їди) та ценосарк. Тіло зооїда складається з нижньої, за-
нуреної в скелетну чашечку кораліта частини, та верх-
ньої, що піднімається над коралітом. Під час подразнення
корала верхня частина поліпа вгортається всередину
нижньої. Верхня частина тіла поліпа розправляється за
рахунок нагнітання води через рот. Сифоногліфів у склер-
актиній немає. Рот і глотка в них виконують функцію кла-
пана, що запобігає відтоку води з гастральної порожнини,
181
й результаті чого тіло поліпа опирається на замкнену все-
редині нього воду — своєрідний гідроскелет.
На підошві в зооїдів колонії утворюються скелетні
денця, що відокремлюють скелети живих поліпів від ске-
летів попередніх поколінь, які лежать під ними.
* М'яке тіло колонії розташоване на поверхні загально-
го скелета - - поліпняка. Набуваючи його форми, воно
вкриває скелет у вигляді сплющеного мішка, стінки якого
складаються з епідерми, тонкої мезоглеї та гастродерми.
Нижній бік мішка щільно обгортає поверхню скелета, за-
ходячи вглиб його порожнин. Підошви окремих поліпів
з'єднані між собою в суцільний покрив, епідерма (каліко-
бласти) якого на нижньому боці виділяє вапно, що запов-
нює простір між -поліпами й утворює скелет ценосарка.
Порожнини ценосарка сполучаються одна з одною та з
гастральними порожнинами отворами або трубочками; всі
порожнини вистелені гастродермою (див. рис. 118).
Отже, живі поліпи містяться лише в поверхневому шарі
колонії, а вся товща поліпняка утворена скелетами мину-
лих поколінь. Під час росту колонії в процесі поновлення
поліпів вони створюють над старими днищами нові, при-
чому м'які тканини залишаються лише в поверхневій час-
тині кораліта, а' стовпчик із днищ тягнеться вглиб скеле-
та. За об'ємом і масою скелет багаторазово переважає жи-
ві тканини, які ніби розтікаються тоненьким шаром по
поверхні масивного поліпняка.
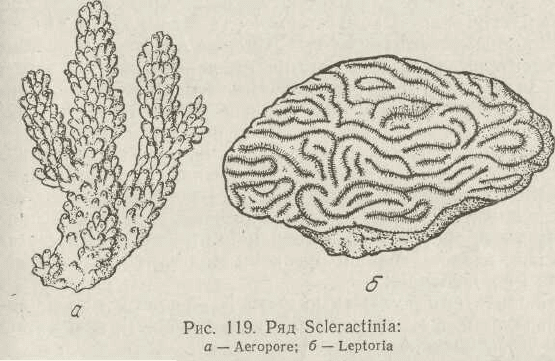
За формою колонії склерактиній надзвичайно різнома-
нітні, причому будова колонії залежить переважно від ха-
рактеру руху водних мас. Один і той же вид коралів у
більш спокійній воді будує кулястий поліпняк, в менш
спокійній — гіллястий, в бурхливій — плескатий, що сте-
леться по дну (рис. 119).
Тривалий час особливості живлення, дихання та про-
цеси, -що зумовлюють утворення могутнього скелета, не
вивчалися. За аналогією з іншими кишковопорожнинними
склерактиній вважали хижаками. Нині з'ясовано, що жив-
лення склерактиній відбувається трьома способами.
1. Склерактинії використовують ту їжу, яку приносять
їм морські течії: дрібні планктонні організми, яйця, цисти,
органічні
1
часточки тваринного та рослинного походження
(псевдапланктон). Важливу роль в уловлюванні та погли-
нанні їжі відіграють війки, що вкривають оральний диск,
а також слиз. Поліпи випускають з рота слизисті тяжі,
які витягуються між щупальцями. Під дією сильних токів
води, що виникають внаслідок биття війок, слиз розпушу-
ється, утворюючи сітку клейких ниток, до якої прили-
182
пають харчові часточки. Періодично слиз разом із осіли-
ми на ньому часточками їжі втягується до рота. Цей ме-
ханізм поліпи використовують не тільки для фільтрації, а
й для хижого живлення. Рухлива здобич вбивається жал-
кими клітинами щупалець, потрапляє в слизові сітки, при-
клеюється до них і заковтується разом із слизом.
Нещодавно відкрито ще один спосіб живлення склер-
актиній— перетравлення осілих на*поверхні тіла харчо-
вих часточок ферментами, що секретуються всією поверх-
нею епідерми, її клітини здатні також захоплювати бак-
терій, що осідають на їх поверхні. Проте тропічні води
бідні на планктон, тому склерактинії в такий спосіб свої
потреби не задовольняють.
2. Результати дослідів із радіоізотопами свідчать, що
склерактинії здатні засвоювати з води через покриви ор-
ганічні речовини (амінокислоти, пептиди тощо), вилуча-
ючи їх із навколишнього середовища. Епідерма поліпів
має надзвичайно складну будову. Вона вкрита мікровор-
синками, впинаннями мембрани, що збільшує контакт по-
верхні епідермальних клітин із зовнішнім середовищем, по-
легшуючи засвоєння з води молекул розчинених у ній ор-
ганічних речовин. Великі колоїдні частки їжі засвоюють-
ся шляхом піноцитозу, а дрібні молекули — шляхом актив-
ного ферментного транспорту. Це було доведено в дорлі-
.дах з коралами роду Pocillopora.
3. Склерактинії одержують поживні речовини також за
допомогою симбіонтів - - зооксантел. Це вид динофлаге-
лят Symbiodinium microadriaticum.
183
Симбіодініум не трапляється у вільному стані, проте
він здатний вступати в симбіотичні відносини з багатьма
безхребетними — кораловими поліпами, губками, меду-
зами, війчастими червами, молюсками і навіть найпрості-
шими (інфузоріями, радіоляріями, форамініферами). Сим-
біодініуми містяться в клітинах гастродерми або в між-
клітинниках. Вирости клітин гастродерми хазяїна утворю-
ють своєрідну капсулу навколо зооксантели. Від клітини
хазяїна її відокремлює складної будови тришарова обо-
лонка, що містить клітковину. Ця оболонка надійно ізо-
лює симбіонта від атаки лізосом хазяїна, запобігаючи
його перетравленню. Разом із тим вона проникна для ме-
таболітів у процесі обміну симбіонта з його клітинами.
За допомогою мічених за С
14
молекул С0
2
виявлено, що
зооксантели синтезують органічні речовини, які засвоюють
корали. У процесі фотосинтезу в зооксантелах утворю-
ються переважно жири, але вони не накопичуються в зоо-
ксантелах, а переходять у клітини поліпа. Жир — це ос-
новний енергетичний матеріал поліпів, який є також ос-
новною запасною речовиною. Зворотний рух метаболітів
із клітин хазяїна в клітини симбіонта полягає перш за все
у споживанні зооксантелами біогенних речовин — солей
азоту та фосфору. Зооксантели можуть також засвоювати
з клітин хазяїна складні органічні молекули типу нуклео-
тидів і поліпептидів.
Склерактинії дихають киснем, розчиненим у воді, але
види поліпів, що мають симбіотичні водорості, одержують
його безпосередньо в товщі тканин.
Симбіодініум знаходить у коралах притулок і захист
від ворогів. Він використовує для фотосинтезу вуглекис-
лий газ, який виділяють поліпи в результаті дихання, а
також утилізує азот і фосфор, що містяться в продуктах
метаболізму коралів.
Ті види коралів, які мають симбіотичні водорості, нази-
ваються герматипними, а ті, що не мають, — агерматип-
ними. Герматипні корали можуть довго жити в чистій воді
без усякої їжі, лише за рахунок симбіонтів. Якщо їх
штучно позбавити водоростей (утримуючи деякий час у
темряві), вони припиняють ріст і розмноження, а потім,
незважаючи на достатню кількість їжі, гинуть.
Крім зазначених переваг у живленні та диханні, гер-
матипні корали значно швидше (майже в десять разів),
ніж агерматипні, можуть нарощувати свій скелет. Тому
серед рифоутворювачів переважають саме герматип-
ні види. Механізм нарощування вапнякового скелета та-
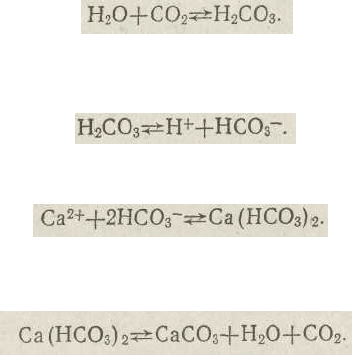
кий. У воді завжди є невеличка кількість СОг та іонів
184
Са
24
-. Під час розчинення СО
2
у воді незначна його части-
на (близько 1 %) вступає в реакцію з водою, утворюючи-
вугільну кислоту:
Вугільна кислота — нестійка сполука, вона може існу-
вати лише у водному розчині, причому розпадається на
іони:
Під час взаємодії іонів Са
2+
і НСО
3
~ утворюється вуг-
лекальцієва сіль (гідрокарбонат кальцію):
У разі надлишку СО
2
реакція зсувається ліворуч, тоб-
то в бік утворення розчинного гідрокарбонату, а внаслідок
нестачі СО
2
— праворуч, що супроводжується випадінням
осаду вапна. Протягом світлого часу доби симбіодініуми
в процесі фотосинтезу безперервно поглинають із тканин
коралів ССЬ, завдяки цьому вміст його у внутрішньому се-
редовищі організму коралів знижується, що сприяє зсу-
ванню хімічної реакції в бік осадження вапна.
Фотосинтез у симбіодініума відбувається найбільш-
ефективно в умовах інтенсивного освітлення. Тому герма-
типні корали оселяються на незначній глибині, що не пе-
ревищує 50 м, утворюючи рясні зарості — рифи. Корало-
вими рифами називаються масові поселення морських ор-
ганізмів, що мають твердий вапняковий скелет. Основу
такого рифу становлять мадрепорові корали (склеракти-
нії), але існує багато організмів, які теж мають вапняко-
вий скелет і беруть участь в утворенні рифу — вапнякові
водорості, гідрокорали, сидячі поліхети, що живуть у вап-
някових трубках, молюски, вусоногі раки. Проте чільну^
роль в утворенні рифу відіграють все ж таки склерактинії.
Корали-рифоутворювачі потребують досить високої й
постійної температури води (не нижче 20°С), нормальної
океанічної солоності (35 °/оо) і освітлення. Тому коралові"
рифи поширені винятково в тропіках.
Розрізняють берегові рифи, що оточують береги остро-
вів або материків, бар'єрні рифи, які тягнуться вздовж
185-
Ця речовина розчинна у воді, але нестійка й легко пе-
ретворюється на нерозчинну сіль — карбонат кальцію-
(вапно):
берега на деякій відстані від нього, атоли — кільцеподібні
коралові острови. € також коралові банки, що виникають
на мілинах, і густі коралові поселення в лагунах і бухтах.
Походження коралових споруд здавна цікавило зооло-
гів і геологів. У різні часи було висунуто багато гіпотез,
але найбільш переконливою залишається теорія, створена
ще Ч. Дарвіном у 1842 р. На його думку, першою стадією
утворення коралових островів є облямовуючий береговий
риф. Корали використовують у цьому випадку береги ост-
ровів як субстрат для будівництва рифу. Якщо в резуль-
таті тектонічних явищ у земній корі дно моря починає
опускатися, й острів поступово занурюється у воду, бере-
говий риф, який облямовує, з тією ж швидкістю росте
вгору та вздовж зовнішнього краю, де більш чиста й ба-
гата на кисень вода. Корали внутрішнього краю., оберне-
ного в бік острова, поступово відмирають, і береговий риф
'перетворюється на бар'єрний, який все далі відступає від
берега. Коли острів повністю заглиблюється під воду, на
його місці утворюється лагуна, а бар'єрний риф перетво-
рюється на атол.
У Індійському та Тихому океанах розкидані сотні та-
ких атолів діаметром від 1 до 160 км, але більшість з них
має
діаметр
3—30
км.. Загальна площа сучасних рифів
становить близько 600 тис. км
2
.
Кораловий риф — це своєрідна екосистема, до складу
якої входять не тільки організми-рифоутворювачі, а й ве-
личезна кількість інших тварин і рослин. Автотрофні ор-
ганізми (в основному симбіодініум) синтезують органіч-
ну речовину, тварини її споживають, поїдаючи м'які тка-
нини поліпів. Серед них переважають молюски, поліхети,
морські зірки, краби, риби. Багато тварин, такі, як сидячі
в трубках поліхети, вусоногі раки, голотурії, знаходять
тут притулок, а живляться завислими у воді органічними
рештками; інші, такі як нематоди, морські їжаки, офіури,
живляться детритом. Є й хижаки, які вживають в їжу ін-
ших мешканців рифів.
Характерна особливість коралових рифів полягає в їх
надзвичайно високій біологічній продуктивності. Авто-
трофна фотосинтетична продукція в екосистемі- рифу ста-
новить від 50 до 300 г сирої біомаси на 1 м
2
за добу.
Отже, екосистеми коралових рифів --одні з найбільш
зрілих і ефективно функціонуючих біогеоценозів нашої
лланети. Завдяки надзвичайно високій продуктивності й
здатності до біофільтрації сучасні рифи відіграють важ-
ливу роль у глобальних біогеохімічних процесах, а також
у динаміці Світового океану.
186
Протягом тисячоліть коралові рифи становили основу
життя цілих народів, мешканців островів Океанії. Ці ост-
рови складаються з коралового вапняка. Завдяки багато-
віковому досвіду полінезійці, меланезійці та мікронезійці
користувалися всіма багатствами рифу, не завдаючи шко-
ди його існуванню.
Для коралових рифів згубними є опріснення (наприк-
лад, тропічні зливи під час відливу), осушення, урагани —
після таких катастроф риф відновлюється протягом десят-
ків років. Коралами живляться деякі риби, краби, мор-
ські зірки та інші організми, але їх кількість регулюється
іншими хижими тваринами. У разі порушення біологічної
рівноваги внаслідок забруднення води або посиленого ви-
лову окремих видів тварин можуть надмірно розмножи-
тися певні види хижаків, які живляться коралами, що
призводить до загибелі цілих масивів коралів. Така ката-
строфа загрожувала в останні роки Великому Бар'єрному
Рифу, що розташований біля берегів Австралії, через над-
мірне розмноження морської зірки «терновий вінець»
(Acanthaster planci), якби не своєчасні заходи уряду. От-
жк, екосистема коралового рифу дуже своєрідна й уразли-
ва, вона здатна існувати лише в дуже обмеженому діапа-
зоні зовнішніх умов, тому потребує захисту.
ВИКОПНІ КИШКОВОПОРОЖНИННІ
Найстародавніші кишковопорожнинні, що відомі нау-
ці,— це безскелетні медузоподібні організми, залишки яких
знайшли у відкладах кінця протерозойської ери (венд.).
Палеонтологи віднесли їх до класу Cyclozoa (рис. 120).
Ці організми досягали кількох сантиметрів у діамет-
рі, вели переважно прикріплений спосіб життя, мали доб-
ре розвинений шар мезоглеї, що виконувала опорну функ-
цію. Цикломедузи прикріплювалися аборальним полюсом
до субстрату, їх гастральна порожнина не мала поділу на
шлунок і гастроваскулярні канали, як у сучасних медуз, а
була суцільною, досить великою й простягалася до пери-
ферії дзвона. Більшість цикломедуз мала досить вираже-
ні циклічні структури" на аборальному боці дзвона. У дея-
ких з них концентрична покресленість комбінувалася з ра-
діальними елементами (є припущення, що це були,раді-
альні канали).
Серед вендських медуз траплялися також радіально-
симетричні форми, але їх радіальна симетрія мала неви-
значений порядок, який змінювався навіть у онтогенезі -
у процесі розвитку організму виростали нові гілки раді-
187

альних каналів, щупальця тощо. Ці організми палеонто-
логи об'єднали в клас Inordozoa.
Рідше траплялися медузи з трипроменевою симетрією,
що охоплювала кишені шлунка, радіальні канали гастро-
Рис. 120. Реконструкція будови вендських кишковопорож-
нинних:
а, б — Cyclomedusa plana (Cyclozoa); в — Armillifera parva; е — Во-
nata septata (Inordozoa); д — Albumares brunsae (Trilobozoa)
васкулярної системи, ротові лопаті, гонади, їх віднесли
до класу Trilobozoa.
Були також і медузи з чотирипроменевою симетрією,
яка властива сучасним сцифоїдним медузам, ці організми
умовно віднесли до класу Scyphozoa.
У вендській фауні знайдено також перших гідроїдних.
Це були поодинокі безскелетні організми, близькі до су-
часних хондрофор (клас Hydrozoa, ряд Chondrophora).
Вони мали форму диска, на верхньому боці якого містив-
ся кіль, подібний до паруса сучасних Velella. На відміну
від більшості сидячих цикломедуз вони плавали на поверх-
ні океану.
Вважають, що вендська фауна існувала порівняно не-
довго й майже вся вимерла ще до початку кембрійського
періоду.
Початок кембрію ознаменувався появою тварин, що
утворювали скелет, але серед кишковопорожнинних таких
ще не існувало. Перші скелетні корали з'явилися всереди-
ні ордовика. Найстародавніші корали відрізнялися від су-
часних тим, що мали вигляд невеличких келихів, на внут-
188 '
рішніх стінках яких ще не було справжніх септ, а лише
невеличкі реберця. Вони не утворювали великих колоній,
а стелилися по субстрату. Ці організми віднесли до класу
Anthozoa, підкласу Tabulata (рис. 121). Деякі з них уже
були рифоутворювачами.
Пізніше виникла ще одна група коралів, які мали
справжні септи в гастральній порожнині, причому в них
закладалися шість первинних склеросепт, а наступні сеп-
ти утворювалися не в шести, а в чотирьох секторах, тому
їх вважають чотирипроменевими коралами (підклас Ru-
gosa).
Ще одна група викопних коралів — геліолітоідеї (під-
клас Heliolithoidea) утворювала масивні колонії з добре
І
Рис. 121. Викопні корали:
а — кзлонія коралів Tabulata; б — поодинокий чотирипроменевий
корал Omphiuma; s — колонія Heliolithoidea
розвиненим ценосарком, їх кораліти характеризувалися на-
явністю дванадцяти склеросепт.
Але жодна згадана група коралів не збереглася до на-
ших днів. Протягом геологічної історії кілька разів масо-
во вимирали одні групи й виникали інші. Коралові ком-
плекси вимирали тричі — наприкінці середнього девону
(палеозой), на межі пермського та тріасового періодів, на
початку крейдяного періоду (кінець мезозойської ери).
Перші справжні восьмипроменеві корали з'явилися в си-
лурійському періоді палеозойської ери, а склерактинії —
в середньому тріасі (початок мезозойської ери). Близькі
до сучасних гідроїдні (гідрокорали) відомі з ордовика.
Вони були представлені колоніями з хітиноїдним і вапня-
ковим скелетами.
189
ТИП РЕБРОПЛАВИ (CTENOPHORA)
До цього типу належать винятково морські тварини.
Відомо близько 120 видів реброплавів, більшість яких
трапляється в тропічних морях; в полярних регіонах окре-
мі види утворюють великі скупчення. У Чорному та Азов-
ському морях живе лише один вид Pleurobrachi chodopis,
що з'явився тут нещодавно. Реброплави — переважно віль-
ноплаваючі хижаки, однак ще відомі повзаючі, а також
сидячі детритоїдні види. Розміри реброплавів коливають-
ся від 2—3 мм (наприклад, блакитний тинерфе — Tinerfe
cyanea) до 2,5 м (венерин пояс — Cestus veneris).
Реброплави — радіально-симетричні двошарові тварини
(з зачатковою мезодермою)., в яких на відміну від Cnida-
гіа, немає жалких клітин, Іюліпоїдної фази розвитку та
метагенезу. План будови реброплавів характеризується
поєднанням восьми- та двопроменевої симетрії. На .відмі-
ну від Cnidaria реброплави рухаються за допомогою видо-
змінених війок -- гребних пластинок. Вони є гермафроди-
тами — запліднення зовнішнє, розвиток прямий. До цього
типу належить один клас Реброплави (Ctenophora).
КЛАС РЕБРОПЛАВИ (CTENOPHORA)
Тіло реброплавів має мішкоподібну овальну, округлу
або грушоподібну форму. На одному його полюсі —
оральному міститься ротовий отвір, на протилежному —
аборальному — аборальний орган. Головна вісь тіла про-
ходить через обидва полюси (рис. 122).
На поверхні тіла реброплавів у меридіональному на-
прямку (від орального до аборального полюса) розміще-
ні вісім валків, або ребер (звідси українська назва типу
та класу), на яких розташовані поперечні гребні пластин-
ки, утворені з'єднаними війками. Це найдовші в тварин-
ному царстві війки (довжина — кілька міліметрів), які
до того ж в центрі мають не дві, а три мікротрубочки.
Гребні пластинки під час биття розкладають світло, тому
при русі вздовж них пробігають райдужні переливи. Греб-
ні пластинки •— основні органи руху, завдяки їх роботі
тварина рухається оральним полюсом уперед.
Більшість реброплавів має два щупальця, інколи знач-
но довші за тіло, що втягуються в спеціальні щупальцеві
кишені. На одній з поверхонь щупальця розгалужені та
вкриті специфічними клейкими клітинами, які є лише в
реброплавів (рис. 123).
190
>
Кожна клітина має напівсферичну форму та приєдну-
ється до щупальця еластичним прямим тяжем (видозмі-
нене ядро) й тяжем, спіральне закрученим навколо пря-
мого (видозмінений джгутик). Клітина продукує клейку
речовину, за допомогою якої на неї налипають планктонні
організми, й щупальце підтягує здобич до рота. Якщо здо-
бич намагається вирватися, тяжі пом'якшують струси, що
відчувають при цьому реброплави.
Тіло реброплавів, як і кишковопорожнинних, склада-
ється з епідерми (ектодерми) та гастродерми (ентодерми),
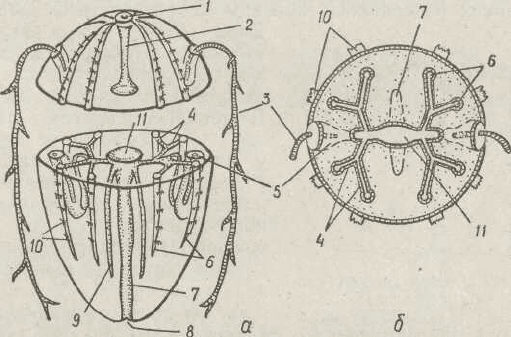
Рис. 122. Схема будови реброплава — загальна схема (а), поперечний
зріз на рівні шлунка (б):
/ — аборальний орган; 2 — аборальний канал (акрогастер); 3 — щупальце) 4 —
екваторіальні канали; 5 — піхва щупальця; 6 — меридіональні канали; 1 —
-лотка- 8 — рот- 9 — глотковий канал; 10 — гребні пластинки; 11 — шлунок
розділених добре розвиненою мезоглеєю. Часто вона криш-
талево прозора, тому тварину не помітно у воді. Такі ви-
ди важко добувати, бо ніжне тіло тварин руйнується. Про-
те є деякі глибоководні реброплави з щільною мезоглеєю.
Більшість повзаючих форм яскраво забарвлена.
М'язова система реброплавів добре розвинена, особли-
во у форм, що повзають. Мускулатура реброплавів склада-
ється з самостійних м'язових клітин, як і в коралових по-
ліпів. Вона розташована в товщі мезоглеї й складається
з поздовжніх і кільцевих волокон під епідермою, подібних
до них волокон навколо глотки та шлунка, а також раді-
альних волокон, що йдуть від шлунка до покривів. Най-
більше розвинена мускулатура щупалець. Мускулатура
191

забезпечує захоплення їжі ротом або щупальцями та вико-
нує захисну функцію —• м'язи, скорочуючись, прикривають
аборальний орган. У деяких реброплавів мускулатура ви-
конує ще й локомоторну функцію, особливо у форм, що
-ведуть повзаючий спосіб життя або плавають завдяки зміє-
лодібним рухам стрічковидного тіла чи за допомогою ро-
тових лопатей.
Травна система починається рухливим ротовим отво-
ром, що веде в об'ємну ектодермальну глотку, де на від-
міну від інших тварин за рахунок ферментів, що виділя-
ються її стінками, в основному перетравлюється їжа.
У деяких реброплавів глотка може вивертатися назовні.
Вона відкривається в ентодер-
мальний шлунок, від якого
відходять три яруси каналів
гастроваскулярної системи.
Перший (акрогастер) підхо-
дить до аборального полюса
у вигляді одного каналу, що
розгалужується на чотири ко-
ротенькі гілки. Дві з них за-
кінчуються сліпо, а дві від-
криваються порами назовні.
Другий ярус складається з
двох каналів, які розташовані
біля екватора перпендикуляр-
но до осі шлунка. Вони двічі
розгалужуються, утворюючи
вісім екваторіальних каналів,
що впадають у вісім меридіональних. Меридіональні ка-
нали розташовані під гребними пластинками. Третій ярус
утворений двома глотковими каналами, які відходять від
шлунка й тягнуться вздовж глотки до орального полюса.
Залежно від форми тіла та розвитку додаткових каналів,
їх відгалужень і переплетень будова гастроваскулярної
системи може значно відрізнятися від описаної.
Глотка в реброплавів має вигляд сплющеної трубки,
ллощина, в якій вона сплющена, називається глотковою.
Шлунок тварин теж сплющений, але в площині, перпенди-
кулярній до глоткової, у цій же площині розташовані два
щупальця (якщо вони є), тому вона називається щупаль-
цевою.
Через тіло реброплава можна провести дві взаємно
перпендикулярні площини — глоткову та щупальцеву,
тобто реброплави побудовані за двопроменевим типом си-
метрії. Проте більшість органів (ряди гребних пластинок,
192
Рис. 123. Будова клейких клі-
тин реброплава:
1 — півкулі з краплинами липкого
секрету; 2 — спіральна нитка; З —
мускульний тяж щупальця
радіальні канали, гонади) розташована навколо головної
осі. Реброплави мають по вісім таких органів. Отже, за-
гальний план будови реброплавів характеризується поєд-
нанням двох типів симетрії — двопроменевого та восьми-
променевого.
Живляться реброплави різними дрібними планктонни-
ми організмами, особливо рачками, полюють на сальп,
дрібних риб, інших реброплавів. Деякі види — коменсали
(від фр. commensal — співтрапезник), тобто тварини, що
живуть за рахунок інших, не завдаючи їм шкоди. Вони
мешкають на поверхні морських зірок, коралів, губок.
Нервова система має вигляд плетива, з невеликими
скупченнями нервових клітин навколо рота, уздовж греб-
них пластинок. Найбільше нер-
вових клітин міститься на або-
ральному полюсі, де є своє-
рідний орган чуття — абораль-
ний орган (рис. 124).
Він складається з статолі-
та—-кульки з вапнякових зе- І,'
рен, що розташований на чо-
тирьох вигнутих еластичних
дужках, утворених злитими
джгутиками. Клітини, до яких .
вони належать, сприймають
зміну тиску статоліта
у
разі
Рис 124
Будова
аборального
зміни положення тіла ребро-
плава. Над статолітом є своє-
рідний ковпачок із війок, що
склеєні разом. Від дужок від-
ходять чотири війчасті боро-
зенки, які розгалужуються й підходять до кінців гребних
пластинок. Під аборальним органом міститься згущення
нервового плетива. Аборальний орган виконує функції ор-
гана рівноваги та регулює рух гребних пластинок. Ребро-
плави реагують на світло. Наприклад, забарвлення мор-
ських огірків (ряд Beroida) на світлі змінюється від мо-
лочно-білого до рожево-фіолетового завдяки дії особ-
ливих клітин-хроматофорів у епідермі.
Багато реброплавів мають здатність до флюоресцен-
ції, яка пов'язана з наявністю особливих клітин-фотоцитів,
локалізованих у гастродермі меридіональних каналів. Се-
ред них найвідомішими є венерин пояс (Cestus veneris),
мнеміопсис (Mnemiopsis), берое (Beroe) та плевробрахія
(Pleurobrachia). Найсильніше світиться берое — світла
однієї особини достатньо для читання. Світіння ребропла-
органа реброплава:
/ — статоліт; 2 — пружні дужки;
З — війчасті борозенки; 4 — епідер-
мальна подушечка; 5 — ковпачок з
війок
13 — 5-856
193
вів, як і інших тварин, зумовлене взаємодією субстрату
люциферину з ферментом люциферазою, в результаті яко-
го випромінюється світло. Світіння в реброплавів коорди-
новане з рухово-захисною реакцією. У разі сильного ме-
ханічного подразнення спостерігаються зупинка биття ві-
йок, різке скорочення тіла й світловий сполох.
Реброплави — гермафродити. Гонади утворюються за
рахунок ентодерми по боках меридіональних каналів га-
строваскулярної системи, причому з одного боку каналу
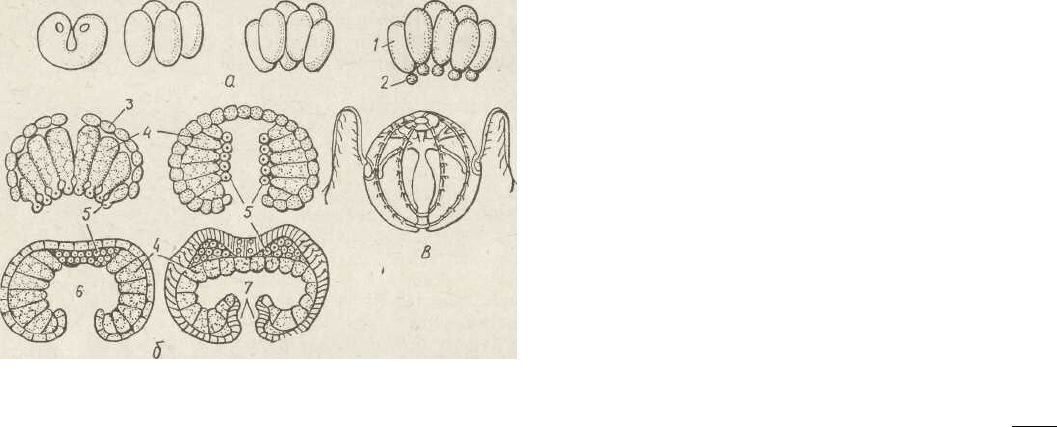
Рис, 125. Ембріональний розвиток реброплавів — дробіння яйцекліти-
ни (а), гаструляція і початок органогенезу (б), личинка (s):
1 — макромери; 2 — мікромери; 3 — ектодерма; 4 — ентодерма; 5 — мезодер-
мальний зачаток; 6 — гастральна порожнина; 7 — глотка
залягає ковбасоподібний яєчник, з другого — такий же
сім'яник. Статеві клітини виводяться у воду через рот або
розриви та тілі, рідше — через спеціальні протоки. Мож-
ливе самозапліднення, тоді перші стадії розвитку відбу-
ваються в гастральній порожнині.
Життєвий цикл у реброплавів^ простий, без метаморфо-
зу. Запліднена • яйцеклітина зазнає повного, але нерівно-
мірного дробіння, внаслідок чого на одному полюсі зарод-
ка (анімальному) утворюються дрібні клітини (мікроме-
. ри), а на другому (вегетативному) —великі (макромери).
З мікромерів розвивається ектодерма, з макромерів —
ентодерма. Гаструляція відбувається шляхом обростання
макромерів мікромерами (епіболія). Частин? клітин
ентодерми заходить у проміжок між ектодермою та
ентодермою. Ці клітини вчені розцінюють як третій зарод-
194
ковий листок — мезодерму, але у реброплавів з нього фор-
муються не основні тканини, а м'язи щупалець і клітини
мезоглеї, які рідко розсіяні в ній. З яйця виходить молода
особина, що за будовою нагадує реброплава найпрості-
шої будови з ряду Cydippida, тому вона називається ціди-
підною личинкою. Далі будова ускладнюється до рівня
дорослих тварин (рис. 125).
У деяких плаваючих видів, описане явище дисогонїї —
своєрідного розмноження на стадії личинки (неотенія),
коли після виходу з яйця вона починає продукувати дрібні
яйця, з яких виходять подібні личинки, що ростуть і пе-
ретворюються на тварин нормальних розмірів. Деякі пов-
заючі форми турбуються про нащадків. У материнському
організмі утворюються виводкові камери, де розвивают'іь-
ся яйця. І/
Нестатевого розмноження в більшості реброплавів йе-
має. Виняток становлять деякі повзаючі форми (Соеіоріа-
na, Planoctena, Vallicula), які розмножуються шляхом ла-
церації подібно до кишковопорожнинних.
В основу сучасної системи реброплавів покладено бу-
дову гастральних каналів у оральній частині тіла. Ця сис-
тема має такий вигляд:
Тип Клас Підклас
Typhlocoela - - -
Ctenophora — Ctenophora —
— Cyclocoela
Ряд
Cydippida
_ Platyctenida
Beroida
— Cestida
— Lobiferida
—Thalassocolycida
Підклас СліпоканальнІ (Typhlocoela)
Підклас об'єднує реброплавів, що не мають анастомо-
зів (з'єднань) між каналами гастроваскулярної системи
біля орального кінця тіла.
Ряд Цідипіди (Cydippida)
Вважається, що цідипіди - - це найдавніший ряд ''реб-
роплавів. За рівнем організації цідипіди схожі- на початко-
ву стадію розвитку всіх інших реброплавів. Типовий пред-
ставник ряду Pleurobrachia pileus трапляється в усіх мо-
рях Світового океану, часто масово (рис. 126, а).
13*
195

Ряд Повзаючі реброплави (Platyctenida)
Деякі види зберегли здатність плавати, наприклад реб-
роплави роду Ctenoplana, проте більшість втратила в до-
рослому стані гребні пластинки й перейшла до повзаючо-
го (рід Coeloplana) або навіть сидячого (рід Tjalfiella)
способу життя. Представники
ряду, що повзають, перетворю-
ються на пласкі істоти, сплю-
щені в орально-аборальному
напрямку. Вони можуть силь-
но вивертати глотку, яка стає
І/1/ /71 повзаючою поверхнею тіла.
/І r/w Часто повзаючі реброплави
\Ч ^&t~-^ трапляються на коралах, де
поїдають їх м'які тканини
(рис. 126,6, в).
196
Підклас Петлеканальш (Cyclocoela)
Для представників цього підкласу характерна наяв-
ність системи перемичок — анастомозів — між гастраль-
ними каналами поблизу орального полюса.
Ряд Морські огірки (Beroida)
У представників ряду на відміну від інших ребропла-
вів немає щупалець на всіх стадіях життєвого циклу. Біль-
шу частину тіла займає велика дзвоноподібна глотка, до
неї веде величезний ротовий отвір, на внутрішньому краї
якого розташовані війкові утвори - - макроцилії, які ут-
римують здобич, зокрема великих сальп, медуз, інших
реброплавів (рис. 126, г).
Найбільші розміри мають арктичні та антарктичні ви-
ди. Морські огірки є кормом для промислових риб, на-
приклад пікші та тріски.
Ряд Стрічкоподібні реброплави (Cestida)
Щупальця в представників ряду є, проте в дорослому
стані вони містяться всередині щупальцевих кишень. Тіло
їх сплющене, але не по орально-аборальній осі, як у пов-
заючих реброплавів, а з боків. Плавають ці реброплави
завдяки синусоїдальним рухам стрічкоподібного тіла, що
зумовлені почерговим скороченням мускульних стрічок,
які тягнуться вздовж боків тіла. Робота гребних пласти-
нок спрямована лише' на підтримку горизонтального поло-
ження головної осі тіла. Найбільший за розмірами пред-
ставник ряду — венерин пояс (Cestus veneris). Живлять-
ся стрічкоподібні реброплави планктонними рачками (рис.
126,
д).
Ряд Лопатеносні реброплави (Lobiferida)
Назва ряду пов'язана з наявністю навколо ротового
отвору або по всьому тілу лопатей різної форми та розмі-
ру, які беруть участь у вловлюванні здобичі — планктонних
мешканців. Вони мають ніжну консистенцію, проте є не-
нажерливими хижаками. Наприклад, мускулястий Осуго'р-
sis crystallina швидко плаває в товщі води, рухаючи ро-
товими лопатями як крилами; він здатний наздоганяти та
заковтувати навіть невеликих риб.
197
Ряд Морські ковпаки (Thalassocolycida)
Це дуже ніжні істоти, що нещодавно були знайдені в
Саргаеовому морі. Своїм куполоподібним тілом вони на-
гадують медуз; живляться пасивно, збираючи ротом
планктонні організми, що прилипають до внутрішньої по-
верхні купола. Рот міститься на особливому вирості, й до
нього підтягуються різні ділянки купола.
Реброплави за рівнем організації близькі до кишково-
порожнинних і раніше об'єднувалися з ними в один тип.
Але вони істотно відрізняються від кишковопорожнинних
•тим, що мають аборальний орган, рухаються переважно
за допомогою війчастих утворень, не мають прикріпленої
* стадії в життєвому циклі, замість жалких мають клейкі
клітини. Реброплави — єдина група багатоклітинних, яка
зберегла первиннопланктонний спосіб життя й походить
від вільноплаваючих предків. Кишковопорожнинні, навпа-
ки, перейшли до прикріпленого способу життя, втратили
аборальний орган і здатність до війчастого руху, що за-
лишилася тільки в личинок. Отже, реброплави та кишко-
вопорожнинні хоча й мають спільних предків, розвивалися
різними еволюційними шляхами.
Реброплави заслуговують на особливу увагу дослідни-
ків у зв'язку з тим, що принцип їх будови часто викори-
стовується для вирішення важливих еволюційних питань.
Наприкінці минулого століття швейцарський зоолог
А. Ланг обгрунтував, ктенофорну гіпотезу походження
плоских червів, яка базувалася на подібності будови тур-
белярій і повзаючих реброплавів. На його думку, сучас-
ні війчасті черви, походять від найдавніших ктенофоропо-
дібних тварин. У наш час ця гіпотеза має лише історичне
значення. Вітчизняний дослідник І. І. Мечников звернув
увагу на риси подібності реброплавів, личинки напівхор-
дових — торнарії та личинки голкошкірих — диплеврули й
висловив думку про походження вторинноротих (голко-
шкірих, напівхордових, хордових) від найдавніших ктено-
фороподібних предків.
ТИП ПЛОСКІ ЧЕРВИ (PLATHELMINTHES)
Переважна більшість плоских червів веде паразитич-
ний спосіб життя. Вільноживучі черви трапляються в мор-
ських і прісних водоймах і значно рідше на суходолі, у
вологому грунті. Розміри плоских червів значно варіюють.
Дрібні види досягають 0,5 мм, але відомі й тридцятимет-
198
рові паразити кишечника китів. Відомо близько 12 тис.
видів плоских червів.
Плоскі черви — білатерально-симетричні тварини з чіт-
ко визначеним головним кінцем. У них розрізняють черев-
ну (вентральну) та спинну (дорзальну) сторони, їх тіло,
як правило, сплющене в дорзовентральному напрямку,
що відображає назва типу.
За рівнем організації плоскі черви стоять дещо вище
кишковопорожнинних і реброплавів. У процесі ембріо-
нального розвитку в них закладаються не два, а три за-
родкових листки — екто-, енто- та мезодерма. За рахунок
5
987
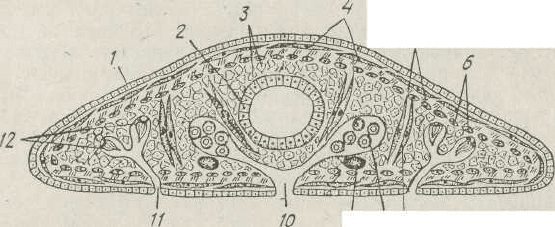
Рис.. 127. Схема поперечного розрізу плоского черва:
/ — ектодерма; 2 — ентодерма (середня кишка); 3 —паренхіма; 4 — кільцеві
м'язи; 5— діагональні м'язи; 6 — поздовжні м'язи; 7 — дорзо-вентральні м'язи;
S — гонада; 9 — нервовий стовбур; 10 — статевий отвір; // —видільна пора;
12 — протонефридій
мезодерми формуються мускулатура, статева система
г
а
також недиференційована сполучна тканина — паренхіма
(мезенхіма), що заповнює всі проміжки між внутрішніми
органами.
Характерною ознакою плоских червів є наявність у них
шкірно-м'язового мішка. Він складається з одношарового
шкірного епітелію, що має різну будову в представників,
різних класів, і кількох шарів м'язів — кільцевих, косих
або діагональних й поздовжніх. CKOp04eHHHM
v
різних груп
м'язів шкірно-м'язового мішка зумовлений червоподібний
рух плоских червів. Паренхіму пронизують пучки дор-
зовентральних м'язів. Крім них, є ще м'язи, що пов'я-,
зані з глоткою, органами прикріплення, статевою систе-
мою тощо.
Безпосередньо під шкірно-м'язовим мішком залягає
паренхіма, що заповнює проміжки між внутрішніми орга-
нами (рис. 127). Плоскі черви належать до пар.енхімних
199