Реймерс Н.Ф. Экология - теории, законы, правила принципы и гипотезы
Подождите немного. Документ загружается.

Существуют два основных способа изображения ареалов на картах:
путем вычерчивания их границ в виде сплошной или пунктирной (в слу-
чае сомнений в верности проведения) линии и методом нанесения точек в
географические места обнаружения особей вида. В первом случае обычно
производится довольно смелая экстраполяция, как правило, путем соеди-
нения крайних точек обнаружения вида, т. е. оба способа отображения
границ ареала фактически совпадают.
Если в какой-то географической точке никто никогда не вел фаунисти-
ческие или флористические исследования, точек находок, естественно,
не возникает. Кроме того, при многолетнем изучении широких регионов,
как это приходилось делать автору в Средней Сибири, нетрудно убедиться,
что некоторые птицы, например, коростель, порой фактически исчезают
на краю своего ареала на ряд лет, а затем вновь появляются. Для упомя-
нутого вида территориальная изменчивость границ ареала составляет сот-
ни километров, так что выразима в самом мелком масштабе. Для менее
подвижных животных и растений такие колебания границ ареала невоз-
можны, но в годы депрессий численности экологическая роль видов, нахо-
дящихся в минимуме, практически равна нулю или стремится к нему.
Исследователь может вид не обнаружить по чисто техническим при-
чинам.
Возможности сопряжения и прогнозирования границ ареалов связаны
с конгруэнтностью видов в составе биоценозов и сообществ (разд. 3.8.).
Как показывает опыт, имеется соответствие видов и ценозов (см. ниже),
и эти физиономически ясные черты позволяют отображать такие законо-
мерности на картах. Однако довольно трудно (хотя и возможно) найти
два вида, которые бы имели абсолютно сходное распределение в простран-
стве. Эту особенность Л. Г. Раменский (1924) и Г. А. Глизон (1926) сфор-
мулировали в виде индивидуалистической гипотезы, правила или принци-
па экологической индивидуальности видов: каждый вид пространственно
распределен в соответствии со своими генетическими, физиологическими
и другими биологическими особенностями и глубоко специфично относит-
ся к факторам среды, в том числе к другим видам. Как нет абсолютно
идентичных особей (кроме однояйцевых близнецов, тем не менее нередко
фенотипически отличающихся друг от друга), так нет и биоэкологически
идентичных видов. Следовательно, их распространение сугубо индивиду-
ально, а потому подвержено столь же индивидуальным нормам реакции
на возникающие изменения среды жизни. Как сказано выше, этот прин-
цип не абсолютен.
В виде регистрации факта можно сформулировать правило колебания
границ ареала: многие виды не имеют четко отграниченной постоянной
области распространения и временами либо исчезают на границе ареала,
либо достигают исчезающе малой численности. В. Тишлер в 1955 г. сформу-
лировал принцип воздействия факторов, согласно которому границы,
размер и характер ареала вида или местообитания популяции обусловле-
ны их биологическими особенностями, и наоборот, биологические особен-
ности вида или популяции могут указывать (быть индикаторами) места,
где их можно найти
1
. Точнее говоря, каждый вид или популяция входят
в какие-то биоценозы и соответствующие экосистемы (биогеоценозы) и
там, где есть вид или его специфическая популяция, очевидно, сущест-
вует тот ценоз, куда они входят и где вырабатывают свою экологическую
нишу. Если не существует такого соответствия, то, очевидно, либо вид,
' Очень примитивно говоря, рыбы живут в воде — одна в реке, другая в озере, третья
в море; одна на глубине, другая на поверхности и т. д.
91
представленный нипулициси, «не
iui»
^шнулиция
nivicci
unynj
OIVJJIUIM-I
ческую нишу), либо ценоз при физиономическом сходстве все же другой!
(иначе сформирован исторически и функционально).
Как показывают широкомасштабные исследования в Сибири и на Даль-1
нем Востоке, виды очень четко соответствуют ценозам, а если наблюдается
разница в постоянно регистрируемом видовом составе всего в один вид,!
то ценоз явно резко отличается от географически или топографически
соседних.
Как правило, смена видового состава происходит не на один, а на!
группу видов, и действительно, вид служит индикатором того сообщества,!:
куда он входит.
Там, где природа сильно нарушена человеческой деятельностью,!
описанное выше правило соответствия вида и ценоза заметно не столь!
четко. Лоскутные природные образования складываются не в рамках быв-В
ших границ элементарных биогеоценозов, а составляют объединенные!?
экосистемы топографически (но не экологически) более высокого иерархи!
ческого уровня. Вид как бы вписывается не в «свой» изначальный ценоз,!
а в мозаику биогеоценотических парцелл разных экосистем, его экологи!
ческая ниша оказывается соотнесенной с этой мозаикой. Отсюда следуещ
вывод, что правило соответствия вида и ценоза в этом случае распростра!
няется на более географически широкие образования, а колебания граж
92
ниц ареалов могут в этом случае быть менее выраженными и не столь
заметными.
Необходимо обратить внимание на то, что в ходе упомянутых процес-
сов нарушается принцип территориальной общности физико-географи-
ческих единиц (разд. 3.7.3), они укрупняются из-за мозаичности возни-
кающих «обрывков» экосистем и их возрастающей отдаленности друг от
друга. Иногда «промежуточные» экосистемы (по иерархии соподчинения)
вообще исчезают, как, впрочем, и элементарные, в своем первозданном
виде. Иерархия экосистем резко упрощается по числу уровней — сохра-
няется как целое лишь 2-й снизу уровень биома или ландшафтной полосы
(см. главу 2) и только им соответствуют по распространению виды, зани-
мая микроучастки, едва ли составляющие полноценные экосистемы (ско-
рее, их парацеллы).
Там, где сохранилась первичная структура природных образований,
соответствие вида и ценоза дает очень четкую пространственную картину
кружева ареала, закономерно меняющегося при движении в географичес-
ком пространстве. Эта закономерность в разд. 3.6 была названа правилом
топографического, или популяционного, кружева ареала. Это правило бы-
ло сформулировано для топографического пространства одной геогра-
фической точки. Теперь следует обратить внимание на изменения этого
кружева в более широком пространстве — правило географической из-
менчивости кружева ареала. Обычно оно состоит в том, что на севере вид
приурочен к приречным, низменным участкам, затем к югу заселяет более
широкие пространства, в горах обитает лишь в локальных участках, а
на южном пределе распространения вновь приурочен к приречным более
влажным и прохладным местообитаниям. У каждого широко распростра-
ненного вида характер изменения кружева ареала специфичен, но смены
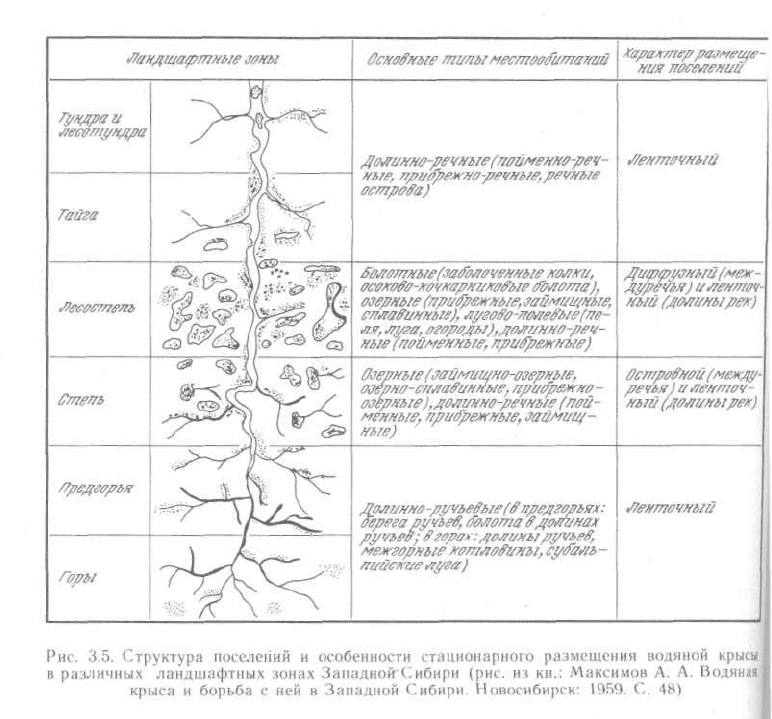
происходят закономерно. Такие смены, пожалуй, лучше всего известны
для водяной крысы в Сибири
1
(рис. 3.5). Пользуясь выявленными законо-
мерностями, можно получить более надежные критерии для экстрапо-
ляции границ ареалов видов.
Проверка приведенных теоретических положений в свое время была
сделана автором на Дальнем Востоке методом артиллерийской «вилки»,
т. е. были намечены точки, где вид уже не наблюдался и где наверняка
существовал, а в промежуточном регионе по характеру растительности
части экосистемы должен был бы присутствовать. В этой срединной точке
проводились исследования. Все ожидавшиеся виды (в том числе не склон-
ные к флуктуации границ ареала) были обнаружены вне зависимости от
того, на каком расстоянии от точки наблюдения были проведены на картах
ранее известные предельные границы их' распространения. Часто находка
была в сотнях километров к северу от «северного предела распростра-
нения» вида.
Правило соответствия вида и ценоза позволило автору предложить
новый тип зоогеографических и шире — биогеографических — карт
2
,
основанных на экосистемном подходе. При этом правило колебаний гра-
ниц ареала в мелком масштабе удалось отразить лишь частично. Зато в
крупном масштабе оно вполне может быть реализовано — ценозы, где
1
Очень показательная схема была опубликована (с. 48) в сборнике «Водяная крыса
и борьба с ней в Западной Сибири». Новосибирское кн. изд-во, 1959 (Максимов А. А. Рас-
пределение водяной крысы по зонам и ландшафтам Западной Сибири. С. 19—48). См. так-
же Пантелеев П. А. Популяционная экология водяной полевки и меры борьбы. М.: Наука,
1968. 255 с.
2
РеймерсН. Ф. Птицы и млекопитающие южной тайги Средней Сибири. М,-Л.: Нау-
ка, 1966. 420 с; Атлас Сахалинской области. М.: ГУГК, 1967.
93
наблюдается пульсация ареалов, графически на картах отличаются о
тех, где вид постоянно присутствует.
Наиболее общим объяснением причин формирования границ ареал;
вида служит правило ограничивающих факторов: факторы среды, наибо
лее удаляющиеся от оптимума экологических потребностей вида, лимита
руют возможности его существования в данных условиях. Поскольку i
лимитирующим факторам относятся любые условия существования ви
да — как абиотические, так и биотические, включая антропогенные,-
правило ограничивающих факторов, ведущее свое начало от группы за
конов минимума (см. разд. 3.5.2), включая закон толерантности Шел
форда (см. разд. 3.5.1), практически дополнительно ничего не объясняет
а лишь резюмирует перечисленные закономерности.
Для объяснения механизма действия принципа воздействия факторо!
В. Тишлера (см. выше) было предложено несколько теоретических осно
ваний. Одно из них — правило стимулирующего действия температур
В. Шелфорда — Т. Парка, в умеренных широтах определяющее стиму-
лирующее воздействие, которое оказывают изменения температуры среди
обитания (правило сформулировано независимо друг от друга В. Э. Шел-
фордом в 1929 г. и Т. Парком в 1930 г.). Это правило в 1972 г. А. Н. Голи-
ков и О. А. Скарлато дополнили правилом лимитирующего значения край-
них летних и зимних температур, или «биогеографическим методом»
указанных авторов: в умеренных широтах Северного полушария распро-
странение видов ограничено необходимыми для размножения температу-
рами. Эта потребность является генотипически закрепленным видовым
признаком и прямо не связана с диапазоном температур выживаемости
особей вида. У видов, переносящих широкие колебания температур, тем-
пература размножения часто более контрастна, чем у видов с относительно
низким порогом выносливости к переменам количества тепла.
Очевидно, нельзя считать, что температура — абсолютно определяю-
щий фактор в формировании границ ареала. Существенную роль играют
влажность, субстраты, межвидовые отношения, преследование человеком,
а в водной среде также скорость движения ее потоков и соленость вод.
Саму температуру регулируют складывающийся мезо-, микро- и био-|
климат. Играют роль и генетические закономерности взаимоотношения]
видов.
Среди них выделяется правило викариата Д. Джордана: ареалы близко-]
родственных форм организмов (правило установлено для животных) обыч-
но занимают смежные территории и существенно не перекрываются;]
родственные формы, как правило, викарируют, т. е. географически заме-
щают друг друга. Согласно правилу представительства рода одним видон\
А. Монара (1919 г.), в однородных условиях и на ограниченной террито-
рии систематический (таксономический) род, как правило, представлен
только одним видом. Видимо, это связано с близостью экологических ниш
видов одного рода.
В связи с этим необходимо вспомнить группу закономерностей, свя-1
занных с принципом конкурентного исключения, или законом (теоремой]
Г. Ф. Гаузе (1934 г.): два вида с близкими экологическими требованиями!
длительное время не могут занимать одну экологическую нишу и, как
правило, входить в одну экосистему. Следовательно, одновременно про-
исходит и географическое замещение видов, что констатирует принцип
видо-родового представительства Й. Иллиеса: поскольку два близкород-,
ственных вида не могут занимать одинаковые экологические ниши в одно»!
биотопе, и, соответственно, в биоценозе, богатые видами роды обычно
представлены в экосистеме единственным своим представителем. Матема-|
94
тически подобные отношения были доказаны уравнениями А. Лотки —
В. Вольтерры (независимо этими авторами в 1925 и 1926 гг.). Однако
поскольку принцип конкурентного исключения, видимо, не абсолютно уни-
версален, был сформулирован принцип сосуществования, или парадокс
Дж. Хатчинсона: два вида в порядке исключения могут сосуществовать
в одной экологической нише, и если близкие виды вынуждены использо-
вать одни и те же ресурсы, то естественный отбор может благоприятство-
вать их сосуществованию, и они способны эволюционировать в близком
направлении. Парадокс Дж. Хатчинсона наблюдается главным образом в
водных экосистемах
1
.
Эмпирическим обобщением всех биогеографических закономерностей,
касающихся ареалов и условий существования видов внутри них, служит
правило географического оптимума: условия обитания вида наиболее опти-
мальны для него в центре ареала. Очевидно, предполагается, что абиоти-
ческая и биотическая среда тут наиболее благоприятна, а экологическая
ниша выработана с наибольшим совершенством. Однако часто имеется в
виду не пространственно-геометрический центр ареала, а его экологи-
ческий «центр». Например, для морских котиков, многих других морских
млекопитающих, колониальных птиц и любых наземных видов с дизъюнк-
тивным или другим типом прерывистого ареала едва ли геометрический
центр их ареала будет оптимален для жизни. Вид может вообще тут отсут-
ствовать. Скорее, понятийно наоборот, экологический оптимум формирует
часть ареала с наибольшей плотностью видового населения — оно имеет
наивысшую плотность и общую численность в наиболее благоприятных
местах существования популяций вида. Эти места могут располагаться
как в глубине очерченного ареала, так и у его географических границ.
В «чистом» виде правило географического оптимума справедливо лишь
для видов с обширным сплошным ареалом, как правило занимающим
всю либо большую часть биома или географической зоны. Однако и в
этом случае нередко возможны варианты, когда наиболее благоприят-
ными оказываются какие-то края области распространения вида. Напри-
мер, многие таежные животные (лось, волк и др.) явно приурочены не к
глухой срединной тайге, а к так называемой подтаежной южной ее полосе.
Здесь больше кормов по естественным причинам и из-за нарушения лесов
человеком, к тому же теплее, и как правило, менее глубок снежный по-
кров.
Ясно, что перечисленные биогеографические закономерности неравно-
значны по глубине обобщения и вкладываемому смыслу.
Совершенно очевидно, что все закономерности системы «организм —
среда» (разд. 3.5) отражаются на формировании границ ареалов видов.
Наибольшее индикаторное значение имеет правило соответствия вида и
ценоза, которое хотя и не объясняет причин такого соответствия, но отра-
жает реально существующий факт, возникающий на почве сложного пере-
плетения многих закономерностей генетического, аутоэкологического и
синэкологического характера. Они формируют кружево видового ареала.
Его географические пределы зависят от толерантности особей вида к
физическим условиям среды и способности выработать экологическую
нишу в одном или в нескольких ценозах, само существование которых
определяется свойствами не столько отдельных видов, сколько их сооб-
ществ (разд. 3.8). Однако и тут важны характеристики видов-средообра-
зователей — эдификаторов среды. Лишь во взаимодействии свойств попу-
' Кроме того, особи морфологически сходных видов, обитающих совместно, стати-
стически различаются по размерам.
95

ляций и их системных объединений складывается картина ареала. Поэтому
все эти закономерности важны для анализа его границ и особенно-
стей.
Нельзя сказать, чтобы внутренние механизмы формирования ареалов
были до конца ясны, а главное, складывались в совокупность знаний,
вполне пригодную для практической деятельности. Лучше обстоит дело с
описанием внешних свойств географически сменяющих друг друга попу-
ляций, особенно животных. Эти закономерности сведены в следующем
разделе.
3.7.2. Изменение особей (популяций) в пределах
видового ареала
Группа закономерностей, характерных для пространственных измене-
ний особей в популяциях, сменяющих друг друга при движении с севера
на юг (или наоборот) и живущих в различных условиях островной изо-
ляции, наиболее известна. Кажется, еще Фридрих I Барбаросса (1125—
1190) знал правило измельчания размеров тела теплокровных животных
при движении с севера на юг, названное затем правилом К- Бергмана
(1847): у теплокровных животных, подверженных географической измен-
чивости, размеры тела особей статистически (в среднем) больше у попу-
ляций, живущих в более холодных частях ареала вида. Правилу Бергма-
на подчиняются около 50% видов теплокровных животных, подавляющая
часть которых (до 90%) птицы. Механизм действия правила Бергмана
очевиден — укрупнение размеров особей термодинамически позитивно ме-
няет отношение объема и площади тела, снижает уровень основного обме-
на. В то же самое время правило не может действовать в тех случаях,
когда наблюдается недостаток кормов или колеблется их доступность:
при увеличении размеров тела общая масса потребляемого корма воз-
растает
1
. Очевидна связь правила Бергмана с правилом поверхностей
(разд. 3.4.2).
Как бы дополнением к правилу Бергмана служит правило Р. Гессе:
особи популяций в северных районах обладают относительно большей
массой сердца (при ее сравнении с массой тела), чем особи популяций,
живущих в южных районах или в более теплых местообитаниях. Большая
интенсивность обмена и необходимость поддерживать относительно по-
1
Но если вид способен к временному снижению метаболизма, а соответственно и тем-
пературы тела, то особи могут не укрупняться при распространении с юга на север. К тому же
результату ведет способность использовать терморегулирующие укрытия, что характерно
для млекопитающих.
96
стоянную температуру тела при более низких температурах требует
лучшего кровоснабжения. А для него нужна более мощная сердечная
мышца.
Столь же очевидны причины действия закона покровов (покрытия)
тела: плотность (густота и терморегулирующая способность) покровов
тела млекопитающих и птиц достигает максимума в холодных и засуш-
ливых областях. Это помогает животным противостоять экстремальным
температуре и влажности. Кстати, этому закону следуют и люди, одевая
наиболее плотную одежду в тех же местах (меховая одежда на севере,
стеганые халаты и длинная одежда в пустынях). Уточнение к закону покро-
вов (покрытия) тела служит правило густоты мехового покрова: у млеко-
питающих холодных местообитаний мех как правило гуще, чем у живущих
в теплых географических зонах и поясах. Часто это правило справедливо
и для млекопитающих сухого климата, где густой мех предохраняет от
излишнего испарения воды с поверхности тела, а также от перегрева-
ния.
Как бы частным случаем правила Бергмана, а скорее, более общего
правила поверхностей (разд. 3.4.2) служит правило Д. Аллена (1877):
увеличение выступающих частей тела одного вида теплокровных живот-
ных (конечностей, хвоста, ушей и т. п.) при продвижении с севера на юг.
Достаточно ясно, что удлинение выступающих частей тела увеличивает
поверхность тела и его теплоотдачу.
Для рыб существует особое правило числа позвонков, или правило
А. Жордана, согласно которому (у сельдей, трески и др.) в водоемах с
повышенной соленостью и более низкими температурами в хвостовой
части тела возрастает число позвонков. Правило Жордана не имеет отно-
шения к правилу Аллена как таковому, а связано с приспособлением к
движению в более плотной среде. Подобного рода закономерностей мно-
го у почвенных и глубоководных организмов — их удлиненность, часто
червеобразность и т. п.
Климатические особенности воздействуют и на цвет особей популя-
ций, живущих на севере и на юге, в сухих и влажных местообитаниях.
Эту специфику отражает правило, или закон К. Глогера (1833): виды
животных северных и влажных географических зон имеют более интен-
сивную окраску, чем обитатели южных сухих регионов. Темная окраска,
отражая меньше тепла, улучшает его аккумуляцию телом животного
1
.
В то же самое время многие виды млекопитающих и птиц Севера имеют
светлую окраску, что объясняется не столько самим цветом мехового по-
крова, сколько его структурой, строением волос, одновременно отражаю-
щих свет и работающих как кумулирующие линзы. Кроме того, на льду
белый цвет служит и защитной окраской (например, у бельков тюле-
ней).
Многие признаки островных популяций видов отличаются от имеющих-
ся у материковых. Объясняет это явление принцип основателя: единичные
особи-основатели новой (изолированной) колонии или популяции несут в
себе лишь часть общей генетической информации, присущей виду. Не-
хватка потенциала наследственной изменчивости может лежать в основе
правила островного измельчания: популяции видов животных, обитающие
на островах, как правило, образуются более мелкими особями, чем матери-
1
Так называемый индустриальный меланизм — потемнение некоторых животных, на-
пример бабочек, в задымленных регионах — видимо, имеет подосновой необходимость более
темной защитной окраски.
4 Н. Ф. Реймерс 97

ковые, живущие в аналогичных условиях. Правило островного измельча-
ния имеет много исключений, особенно на крупных островах (например,
кабарга Сахалина крупнее чем материковая), но, видимо, статистически
верно.
Закономерности географического распространения сообществ также
привлекли внимание экологов и биогеографов. Перейдем к краткому их
перечислению.
3.7.3. Закономерности распространения сообществ
Наиболее общей закономерностью, видимо, следует признать сформу-
лированное А. Уоллесом в 1859 г. правило увеличения видового разнообра-
зия по мере продвижения с севера на юг, или правило Уоллеса. Оно каса-
ется как видов, так и составляемых ими сообществ: в тропиках значительно
больше абсолютное число видов, чем на Севере, и в составе южных сооб-
ществ их также намного больше. Причины возникновения такой разницы
многообразны. Их можно разделить на две группы — эволюционно-гео-
исторические и географо-экологические. Северные ценозы исторически мо-
ложе и находятся в условиях меньшего поступления энергии от Солнца.
Однако едва ли можно считать, что со временем, скажем, биоценозы тун-
дры станут богаче видами. Если обратить внимание на степень замкну-
тости круговорота веществ в тропиках и в северных районах, то нетрудно
заметить, что и там, и там практически не образуется накоплений орга-
нического вещества — гумуса почв. Видимо, энергетическое совершен-
ство в соответствии с законом максимализации энергии и информации
(разд. 3.2.3) выработано экосистемами всех ландшафтно-географических
зон. В них существует столько видов, сколько необходимо для максималь-
ной утилизации приходящей энергии и обеспечения круговорота веществ
в рамках энергетического потока. В связи с этим следует к правилу Уоллеса
добавить принцип эколого-географического максимума (стабильности
числа) видов: число видов в составе географических зон и их экосистем
относительно постоянно и регулируется вещественно-энергетическими
процессами. Это число всегда естественно стремится к необходимому и
достаточному максимуму.
Этот принцип явно связан с общим правилом числа видов в биосфере
Земли (разд. 3.11).
Человеческая деятельность снижает видовое разнообразие. Биоти-
ческие системы начинают работать в необычном режиме нехватки видов.
Компенсация происходит за счет увеличения числа особей согласно прин-
98
ципу экологического дублирования (разд. 3.8.1). Однако значительное
и быстрое снижение числа видов и составляющих их особей может резко
нарушить действие принципа эколого-географического максимума (ста-
бильного числа) видов. Это ведет к тому, что не удовлетворяется и требо-
вание принципа Ле Шателье — Брауна (разд. 3.2.3). Экосистема высо-
кого ранга начинает саморазрушаться. Если этот процесс идет с большой
скоростью, происходит заметное опустынивание территории (песчаные
пустыни известны, например, даже в тайге Забайкалья и Сахалина).
В других случаях складывается природная системная совокупность с иным
максимумом видов и новым балансом круговорота веществ. Для этого
наиболее благоприятны условия умеренных географических поясов, где
круговорот веществ явно незамкнут, вернее, его долговременный баланс
опосредован накоплением органики в почвах, в торфах болот, а в тайге
также и в столетиями гниющих погибших деревьях. Буферность эко-
систем умеренных поясов Земли выше, чем тропических и северных, поэто-
му уничтожение видов в этих двух последних и отход от удовлетворения
условий принципа эколого-географического максимума (стабильности
числа) видов тут предоставляет большую опасность для экологического
благополучия этих регионов, а, возможно, и биосферы в целом.
Принцип конкурентного исключения Г. Ф. Гаузе и правило викариата
Д. Джордана (разд. 3.7.1) характерны не только для видов, занимающих
одну экологическую нишу, но и для функционально аналогичных сооб-
ществ, селящихся в очень близких условиях среды. И. И. Дедю в неодно-
кратно упоминавшемся словаре приводит пример фаун ракообразных кас-
пийского и древнепресноводного происхождения, которые в пределах
Каспийско-Азово-Черноморского бассейна не смешиваются вопреки их
сходству по экологическим требованиям. Популяции взаимоисключающих
видов становятся членами лишь одной из викарирующих соседних эко-
систем (это показатель значительной взаимозависимости членов сформи-
рованного ценоза). Указанный автор формулирует принцип взаимоисклю-
чаемости биотических комплексов: две генетически сходные группы орга-
низмов разного происхождения, обладающие близкими (одинаковыми)
требованиями к окружающей их среде, взаимоисключают друг друга.
Совершенно очевидно, что этот принцип следует учитывать при попытках
акклиматизировать виды, относящиеся к различным, но близким биоти-
ческим комплексам. Такие попытки либо окончатся неудачей, либо будут
разрушительны. Это обычно и наблюдается.
Правило А. Уоллеса, с которого начался обзор в этом разделе, спра-
ведливо для географической зональности в целом и для аналогичных био-
тических сообществ, но именно лишь для аналогичных, так как отсутствие
или присутствие одного или (как правило) группы видов свидетельствует
о том, что мы имеем дело не с той же, а с другой экосистемой (согласно
правилу соответствия вида и ценоза —см. разд. 3.7.1). При этом анало-
гичные экосистемы могут оказываться в рамках различной вертикаль-
ной зональности — чем южнее, тем в более высоких поясах гор (правило
смены вертикальных поясов), или на склонах иной экспозиции; например,
на северных склонах образуются экосистемы более северных ландшафт-
ных разностей. Последнее явление было формально установлено в 1951 г.
В. В. Алехиным и Й. Вальтером в виде правила предварения, или правила
постоянства местообитания (плакорный вид или плакорный биоценоз пред-
варяется на юге или на севере в соответствующих условиях местообитания,
что связано с меньшей или большей инсоляцией склонов различной экспо-
зиции). В ряде случаев образуются участки незональных ценозов, соот-
4» 99
ветствующие более северным или южным зонам (например, степные участ-
ки и тайга). Эти участки нельзя считать адекватными зональными (соот-
ветственно степные участки в тайге называют «степоидами»). Обычно в
этих маргинальных образованиях ценозы беднее видами.
Видимо, именно обедненность ценозов в зонах контакта климатических
и экологических зон формирует закономерность, известную как закон
минимума видов, или эффект А. Ремане (этот автор открыл и сформули-
ровал правило в 1934 г.): минимум морских и пресноводных видов живот-
ных наблюдается в солоноватых водах (соленость 5—8%
0
). В данном
случае возникает не географическая, а экологическая, функциональная
маргинальность.
В пограничных полосах между географическими зонами и вертикаль-
ными поясами часто число видов больше, чем в центральных частях этих
зон и поясов за счет мозаичности экосистем, принадлежащих к соседним
зонам, и так называемого эффекта экотона (разд. 3.9). При этом физионо-
мические «гибриды» ландшафтно-географических зон (лесотундра, лесо-
степь и тому подобное) часто оказываются самостоятельными образова-
ниями со своим видовым составом биоты, особыми биогеоценотическими
закономерностями и другими характеристиками.
Островные экосистемы, возникшие в результате дробления изначаль-
ных при их нарушении человеком, составляют целостные природные обра-
зования (надсистемы) более высокого иерархического уровня, и, соответ-
ственно, функционально целостные физико-географические территориаль-
ные единицы образуются лишь в том случае, если эти экосистемы распо-
ложены не как изолированные, удаленные друг от друга участки, а как
относительно компактная, хотя и мозаичная совокупность. Этот очевидный
факт носит название принципа территориальной общности физико-геогра-
фических единиц (районов, провинций и т. п.).
Менее очевиден сформулированный Г. Ф. Хильми и оставшийся почти
незамеченным научной общественностью закон обеднения разнородного
живого вещества (биоты) в островных его сгущениях. В авторской трак-
товке: «индивидуальная система, работающая в ... среде с уровнем орга-
низации более низким, чем уровень самой системы обречена: постепенно
теряя структуру, система через некоторое время растворится в окру-
жающей... среде»
1
. Другие названия этого обобщения — принцип орга-1
низационной деградации и закон растворения системы в чуждой среде!
(разд. 3.5.2). Фактически это общесистемный закон. Он тесно связан с[
законом оптимальности и в значительной мере отражает термодинамику!
малой системы, находящейся в чуждой среде. Здесь мы вновь возвраща-1
емся к нему, акцентируя внимание на биоте, поскольку искусственное!
сохранение экосистем лишь малого размера (на ограниченной террито-1
рии, например, при заповедании) ведет к их постепенной деструкции и!
не обеспечивает целей сохранения видов и их сообществ. Чем выше разни-1
ца между уровнем организации островной биосистемы и ее окружения, тем!
скорее происходит деградация биоты. Одновременно меняются и все!
остальные компоненты экосистемы, так что сохранить островную биоту
изолировано на малых территориях при любых условиях в длительном
интервале времени практически невозможно.
Законом обеднения разнородного живого вещества Хильми объясня!
ется бедность ценозов видами при действии правила предварения (см,|
выше). Одновременно он тесно связан с известным биогеографическиш!
1
Хильми Г. Ф.' Основы физики биосферы. Л.: Гидрометеоиздат, 1966. С. 272.
100