Карнаухов В.Н. Люминесцентный анализ клеток
Подождите немного. Документ загружается.

По прошествий недели после введения флуорексона изготавливали препараты костей и их
люминесценцию исследовали микроскопически.
В результате такой обработки вокруг гаверсовых полостей кости образовывались
концентрические зоны разного цвета люминесценции. Наиболее удаленная от гаверсова канала
область обладала красной люминесценцией ализарина, далее следовала область золотисто-желтой
люминесценции окситетрациклина и непосредственно к гаверсовой полости примыкало
кольцо
зелёной люминесценции флуорексона. Эта работа представляет собой пример изящного применения
трехцветного люминесцентного метода, позволяющего легко проследить направление и скорость
минерализации кости.
Одним из важнейших и наиболее трудных вопросов, возникающих при попытке использовать
люминесцентные, красители-метки для изучения функциональных механизмов клетки является
вопрос о природе субстрата, связывающего люминесцентную метку. Большая
сложность и
многокомпонентность клетки, наличие в ней взаимодействующих структурных органоидов,
имеющих зачастую сходные механизмы действия, не позволяют использовать непосредственно
представления о взаимодействии метки с какими-либо чистыми веществами in vitro
для
интерпретации данных, полученных при исследовании клетки.
Необходимым, обычно, является проверка этих представлений на все более усложняющихся
модельных объектах, таких, как суспензия митохондрий и их наружных и внутренних мембран,
саркоплазматический ретикулум, сократительный аппарат и т.д. Не менее полезным является
использование в качестве моделей узкоспециализированных клеток, в которых изучаемый механизм
представлен
наиболее ярко. Некоторые подходы подобного типа были описаны выше.
Определенную пользу может принести также прием, заключающийся в применении к одному
и тому же препарату последовательно нескольких люминесцентных меток одного и того же
субстрата. Наблюдаемые при этом зачастую конкурентные отношения меток, обладающих разной по
цвету люминесценцией, могут доставить ценную информацию.
Например,
изложенные выше данные, полученные при исследовании суспензий митохондрий,
указывают на возможность взаимодействия аурамина с местами специфического связывания кальция
на белках. Поэтому представлялось интересным сравнить локализацию аурамина с локализацией
одиночных молекул ализаринового красного S на белках одного и того же препарата.
Оказалось, что если препарат рецептора растяжения рака, окрашенный ализариновым
красным, дополнительно обработать
аурамином, то последний вытесняет с белков ализариновый
красный. Это видно не только по замене его красной люминесценции на зеленую люминесценцию
аурамина, но и по ослаблению плотности ализарина в проходящем свете в местах его
непосредственной связи с белками (мышца). Таким образом, аурамин (катион) выступает здесь как
своеобразный конкурент молекул ализаринового красного S (
анион) за места связывания на белках.
Места, за которые может происходить конкуренция, представляют собой, по-видимому,
отрицательно заряженные группы белков, блокированные Са
2+
и приобретающие таким образом
положительный заряд. Анион ализаринового красного электростатически связывается с ионом Са
2+
, в
свою очередь связанным с отрицательно заряженной группой белка. Появившийся в среде катион
аурамина, взаимодействуя с отрицательно заряженными группами белков, вытесняет с них Са
2+
,
через который осуществлялась связь аниона ализаринового красного с этими группами.
Точно также можно наблюдать взаимоотношения ализаринового красного S и
хлортетрациклина, о котором известно, что интенсивность его люминесценции на несколько
порядков выше, если он связан с кальцием и находится в неполярном окружении — в мембранах
[308, 309, 313]. При окрашивании препарата рецептора растяжения рака хлортетрациклином можно
наблюдать яркую зеленую люминесценцию мышц, приядерных и примембранных областей обоих
нейронов. Дополнительная окраска препарата ализарином приводит почти к полному замещению
зеленой люминесценции хлортетрациклина красной люминесценцией ализарина. Докраска этого же
препарата хлортетрациклином только частично возвращает зеленую люминесценцию, наиболее
заметную в медленно адаптирующейся мышце. Следует при этом отметить, что в отличие от
аурамина хлортетрациклин не вытесняет из препарата красный продукт взаимодействия ализарина с
белками мышц, наблюдаемый в проходящем свете. По-видимому, места связывания этих
люминесцентных меток различны. Результаты подробных исследований взаимодействия
хлортетрациклина с митохондриальной суспензией (рис. 86) указывают на то, что основной вклад в
люминесценцию вносит комплекс хлортетрациклина с Са
2+
, находящийся в гидрофобной области
митохондриальной мембраны [313].
Приведенные выше сравнительные наблюдения показывают, что хотя все три красителя
имеют отношение к кальцию, они характеризуют, однако, разные аспекты его обмена в клетке.
Свободный кальций определяется ализариновым красным S по местам локализации зеленой
люминесценции его одиночных комплексов с красителем (520 нм), по узкополосной (рис. 84, 4)
красной
люминесценции (685 нм) комплексов кальция с двумя молекулами красителя или по местам
локализации красного нелюминесцирующего лака, наблюдаемого в проходящем свете.
Молекулы ализаринового красного S, являющегося анионным красителем, способны к
электростатическому взаимодействию с Са
2+
, связанным с отрицательно заряженными группами
белков. В этом случае ализариновый красный S обладает характерной широкополосной
люминесценцией кирпично-красного цвета (рис. 84, 2; рис. 85, 1, 3) и может вытесняться вместе с
Са
2+
с групп связывания аурамином. Возможно, что в число мест взаимодействия обоих этих
красителей входят и специфические места связывания Са
2+
.
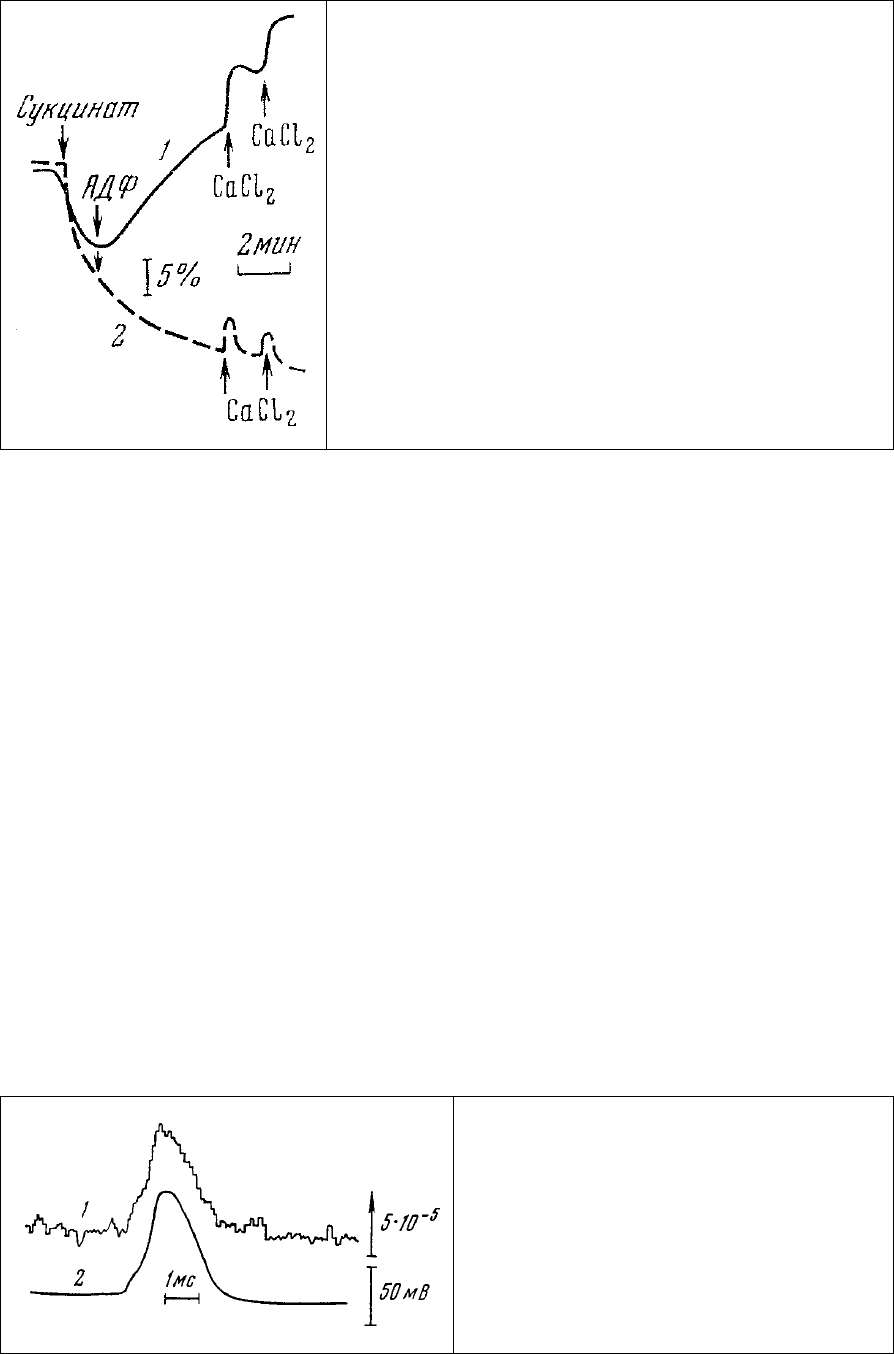
Рис. 86. Изменение люминесценции хлортетрациклина
(длина волны 540 нм), связанного интактными
митохондриями (1) и митохондриями, ингибированными
олигомицином (2) [313]. Длина волны возбуждения 436
нм.
Что касается хлортетрациклина, то его люминесценция выявляет, по-видимому, присутствие
кальция в гидрофобных участках препарата. Это уже упоминавшееся качество хлортетрациклина
резко (в 200-400 раз) увеличивать квантовый выход при связывании его с кальцием в неполярных
(мембраны) структурах клетки нашло применение и при изучении динамики кальция в мембране
аксона при генерации потенциала действия
[314].
4.3. Исследование структуры мембраны при генерации
потенциала действия
Для полноты изложения представляется полезным кратко суммировать на основе
литературных данных попытки применения люминесцентных красителей-меток в изучении
структуры мембраны нерва во время генерации потенциала действия.
Наиболее подходящими для этих целей являются именно красители или комбинации
красителей с переменным квантовым выходом. Естественно, что одной из первых меток,
примененных для изучения
структурных изменений мембраны, оказался АНС. Было установлено, что
изменение интенсивности люминесценции нерва, окрашенного АНС, коррелирует с суммарным
потенциалом действия тонких волокон [57]. Исследования на гигантских аксонах кальмара [315-322]
показали, что изменения интенсивности люминесценции АНС имеют разный знак в зависимости от
того, внутри аксона или снаружи он находится (рис. 87). Аналогичным образом изменяется
люминесценция АНС
в искусственных бислойных лецитиновых мембранах под действием импульсов
напряжения, когда краситель находится в растворе с одной стороны мембраны [323].
Рис. 87. Изменение интенсивности
люминесценции (1) аксона кальмара,
флуорохромированного АНС во время
потенциала действия (2) [322]. АНС на
внешней поверхности аксона. Использовано
синхронное накопление сигнала.
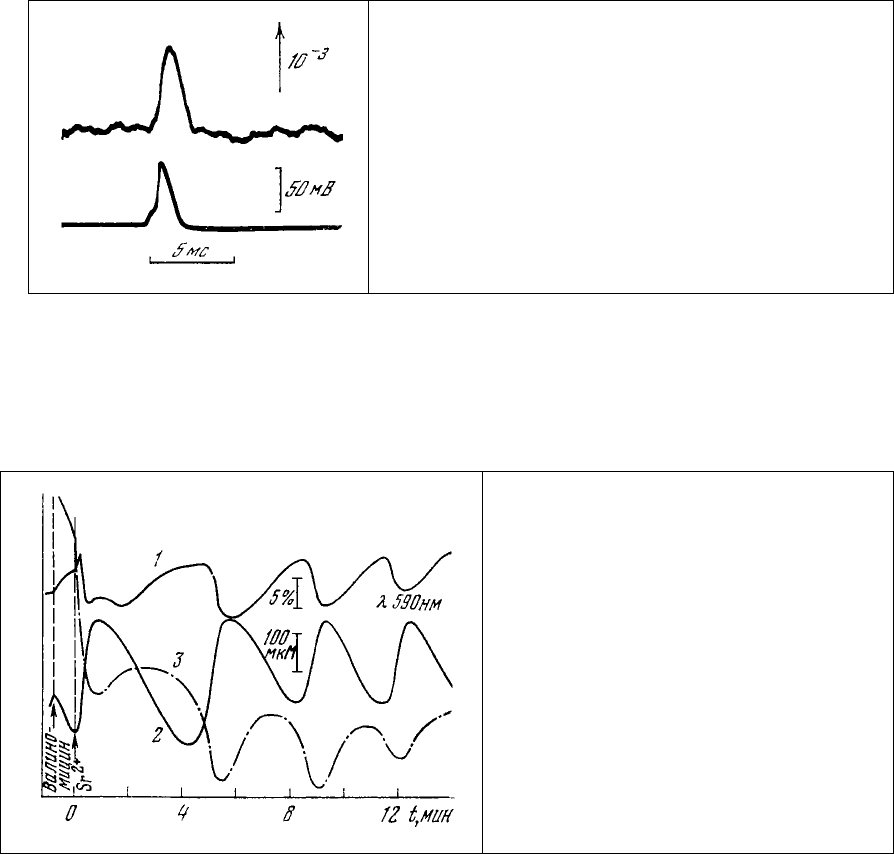
Существенным недостатком АНС в этих экспериментах является весьма малая величина
изменения интенсивности его люминесценции в ответ на электрическую стимуляцию объекта,
лежащая значительно ниже уровня шумов. Поэтому оказывается необходимым применение сложных
методов синхронного накопления для выделения полезного сигнала из-под шума. Более того,
окончательно не ясно, отражает ли потенциалзависимое изменение люминесценции АНС
динамику
структурных перестроек в мембране или оно связано с прямым действием электрического поля на
молекулы красителя или их окружение [324].
Наилучшие результаты, были получены при использовании мероцианиновых красителей,
обладающих высоким соотношением сигнал/шум на аксоне. Изменение люминесценции аксона,
окрашенного мероцианином, оказалось возможным наблюдать во время одиночного потенциала
действия без применения накопления
сигнала [325-327].
«Мероцианин 540»
Применявшийся краситель слабо растворим в воде, и потому готовили его спиртовой раствор,
разбавлявшийся затем морской водой. Конечное содержание спирта - 1%. Данные, полученные на
аксоне кальмара диаметром 0,4 мм, окрашенном в течение 10 мин при температуре 14° С, приведены
на рис. 88.
Рис. 88. Изменение интенсивности люминесценции
аксона кальмара, флуорохромированного
мероцианиновым красителем, во время потенциала
действия [325]. Прямые измерения.
В живых клетках мероцианин окрашивает в основном мембранные структуры препарата,
взаимодействуя при этом, по-видимому, с липидной областью мембраны, свободной от
функциональных белковых групп. Во всяком случае, окраска мероцианином не вызывает нарушений
работы механизмов ионного транспорта, не препятствует генерации потенциала действия и не
нарушает, например, колебательного режима работы митохондриальных мембран (
рис.89).
Рис. 89. Влияние мероцианина на
функциональное состояние
митохондрий. 1 – интенсивность
люминесценции «мероцианина-540»,
связанного с мембраной митохондрий; 2
– концентрация калия в среде; 3 –
скорость потребления кислорода.
Другим перспективным подходом, аналогичным рассмотренному выше на примере
взаимодействия белков в саркомере мышцы, является использование для изучения мембран пары
красителей, квантовый выход которых меняется за счет миграции энергии возбуждения при
сближении меток. Используя такую пару люминесцентных красителей, Кун [328] весьма изящно
исследовал сближение липидных монослоев при их контакте. Монослои формировались из
арахидоновой кислоты на твердой подложке (поливиниловая пленка, стекло) или на поверхности
воды. Поверхность одного монослоя окрашивали красителем-донором (Д), поверхность второго -
красителем-акцептором (А), в качестве которых были использованы люминесцентные метки с
приведенными ниже структурными формулами.
Используя монохроматическое излучение, возбуждали только голубую люминесценцию
метки-донора. Приближение второго монослоя с адсорбированной на нем меткой-акцептором на
расстояние менее 200 Å приводило к уменьшению интенсивности голубой люминесценции донора и
появлению желтой люминесценции акцептора. Миграция энергии возбуждения
от метки-донора к
метке-акцептору наблюдалась в интервале расстояний между этими молекулами от 50 до 200 Å.
Аналогичные результаты были получены и в случае использования в качестве акцептора
нелюминесцирующего гемоглобина, безызлучательно рассеивающего принятую от метки-донора
энергию. Естественно, что в этом случае о миграции энергии возбуждения судили только по
уменьшению интенсивности голубой
люминесценции. Расчеты показывают [328], что
взаимодействие красителей в данном случае может быть описано как взаимодействие
фосфоресцирующих электрических диполей.
Миграцию энергии между различными парами таких соединений, как хлорофиллы а и в, и
некоторыми красителями исследовали на бислойных липидных мембранах [329]. Пигменты вводили
в мембрану и определяли ту концентрацию их, при которой по изменению спектра
люминесценции
обнаруживалась миграция энергии. Было показано, в частности, что миграция энергии между
хлорофиллом а и в осуществляется на расстоянии до 100 Å.По-видимому, методические подходы с
использованием явления миграции энергии окажутся наиболее перспективными при исследовании
структурных изменений в клеточных мембранах в процессе выполнения ими функций
.
4.4. Взаимодействие белков и нуклеиновых кислот
Явление миграции энергии от одного красителя к другому может служить основой для
разработки чрезвычайно чувствительных методов исследования взаимодействия различных
макромолекул в процессе функционирования их в клетке. Некоторые особенности такого подхода
могут быть рассмотрены на примере разработки люминесцентного аналога известной реакции Грама
[330—332].
Отношение микроорганизмов к окраске по Граму является одним из
важнейших
таксономических признаков. Однако классический вариант реакции Грама [333] представляет собой
довольно длительный и трудоемкий процесс, требующий предварительной фиксации исследуемых
клеток. В этой связи представлялось полезным разработать быстрый способ прижизненного
определения грамположительных и грамотрицательных микроорганизмов на основе использования
некоторых специфических свойств люминесцентных красителей.
Как неоднократно указывалось, отношение к окраске по Граму
связано с особенностями
клеточной оболочки микроорганизмов. Предполагается, что в то время как поверхность
грамотрицательных клеток покрыта слоем белков и липопротеидов, в состав клеточной оболочки
грамположительных микроорганизмов входит рибонуклеиновая кислота [32, 333, 334]. Поэтому для
разработки метода были выбраны люминесцентные красители, один из которых - АНС -обладает
высоким сродством к гидрофобным группам белковых молекул, а другой
- этидиум бромид -
характеризуется крайне высокой специфичностью связывания с нуклеиновыми кислотами.
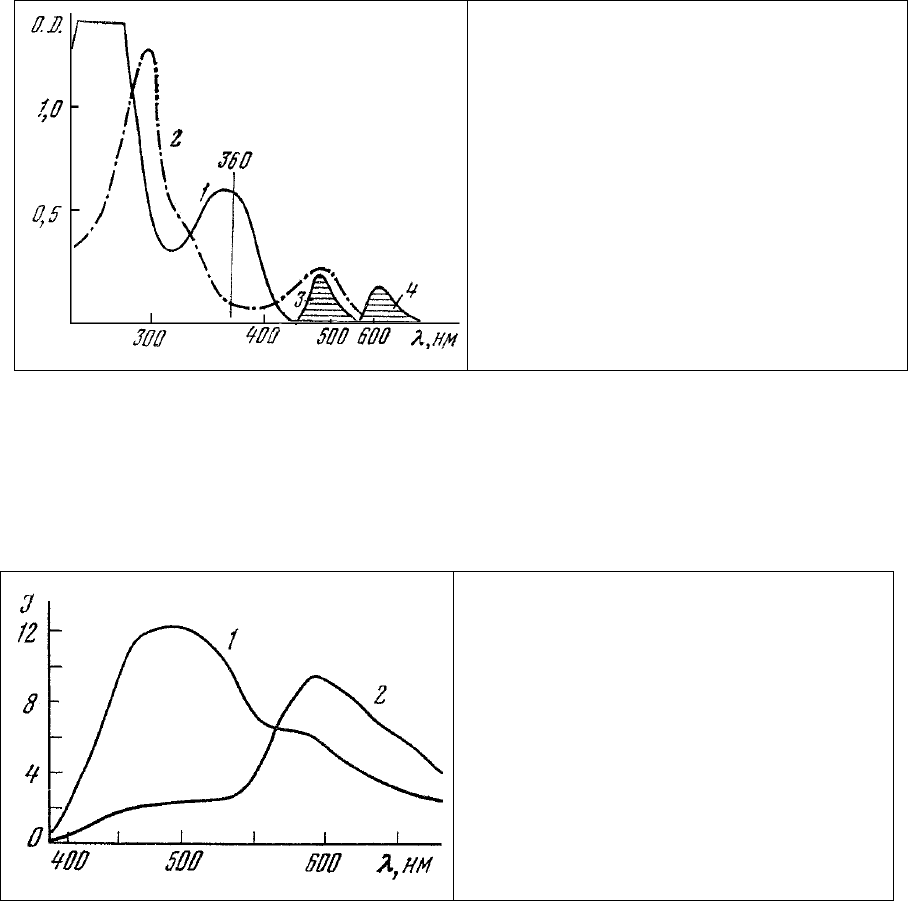
Другим особым свойством, общим для обоих красителей, определившим их выбор для
разработки метода, является способность резко (в 50-100 раз) увеличивать квантовый выход
люминесценции при связывании с субстратом. Это обстоятельство исключительно полезно в
методическом отношении, так как позволяет наблюдать окрашенные клетки без
предварительной
отмывки избытка несвязавшихся красителей. В связанном виде АНС и этидиум бромид обладают
люминесценцией с максимумами 490 и 610 нм соответственно, как это схематически показано на рис.
90. При этом полоса излучения АНС перекрывается практически полностью полосой поглощения
этидиум бромида, что создает особо благоприятные предпосылки для миграции энергии с АНС на
этидиум бромид.
Для разработки метода были использованы 150 заведомо определенных по Граму штаммов
микроорганизмов, полученных из лаборатории коллекционных культур ИБФМ АН СССР.
Микроорганизмы окрашивали водными растворами этидиум бромида в концентрации 10
-5
г/мл и
АНС в концентрации 10
-4
г/мл (Serva, ФРГ). Каплю окрашиваемой бактериальной суспензии
наносили на предметное стекло и добавляли к ней каплю раствора этидиум бромида. Затем препарат
накрывали покровным стеклом, излишки жидкости удаляли фильтровальной бумагой, после чего на
край покровного стекла наносили каплю раствора АНС и оттягивали жидкость фильтровальной
бумагой с противоположной стороны, ускоряя таким образом
распределение АНС в препарате под
покровным стеклом. Препарат окантовывали парафином для предохранения от высыхания и
исследовали микроспектрофлуориметрически при возбуждении люминесценции в максимуме
поглощения АНС (365 нм).
Рис. 90. Спектры поглощения АНС в
концентрации 10
-4
г/мл (1) и этидиум
бромида в концентрации 2,5⋅10
-4
г/мл (2).
Схематически показано положение полос
излучения АНС (3) и этидиум бромида (4) в
спектральном диапазоне.
После окраски этидиум бромидом и АНС по описанной выше методике живые
грамотрицательные микроорганизмы резко отличаются от грамположительных по цвету
люминесценции. Живые грамотрицательные микроорганизмы разных видов приобретают голубую
люминесценцию с максимумом излучения в области 490 нм (рис. 91, 1), в то время как
грамположительные бактерии вне зависимости от их видовой принадлежности обладают однотипной
красной люминесценцией с максимумом излучения в области 605-610 нм (рис. 91, 2).
Рис. 91. Спектры люминесценции
грамотрицательных (1 – Serratia marcescens
B-63) и грамположительных (2 – Bacilus
subtilis В-580) микроорганизмов,
окрашенных АНС и этидиум бромидом.
Длина волны возбуждения 365 нм.
Следует отметить, что в культурах грамотрицательных микроорганизмов, выращенных в
жидких средах, встречаются единичные клетки, обладающие люминесценцией в красной области
спектра. Количество таких клеток увеличивается в препаратах из культур, выращенных на твердых
средах. Можно было предположить, что это мертвые или поврежденные клетки. Контрольные опыты
с окрашиванием суспензий грамотрицательных микроорганизмов, подвергнутых нагреву
до 100° С,
показали, что при увеличении времени нагревания суспензии относительное количество клеток,
приобретающих красную люминесценцию, пропорционально увеличивается.
Смену цвета люминесценции грамотрицательных микроорганизмов с голубого на красный
удобно наблюдать на одиночной клетке у видов, обладающих ярко выраженной двигательной
активностью. Если в свежеприготовленном препарате грамотрицательных бактерий Enterobacter
cloacae клетки характеризуются высокой двигательной активностью
и имеют голубую
люминесценцию (рис. 92, 1), то через некоторое время их движение замедляется и в спектре
люминесценции появляется красная компонента (рис. 92, 2), которая становится доминирующей (рис.
92, 3) после прекращения подвижности клеток. При этом форма спектра люминесценции
поврежденных или мертвых грамотрицательных микроорганизмов (рис. 92, 3) становится такой же,
как и у заведомо
грамположительных клеток (рис. 92, 4).
Следует отметить, что в то время как вегетативные клетки грамположительных
микроорганизмов приобретают красную люминесценцию с максимумом излучения 605-610 нм, их
споры обладают голубой люминесценцией с максимумом излучения 490 нм.
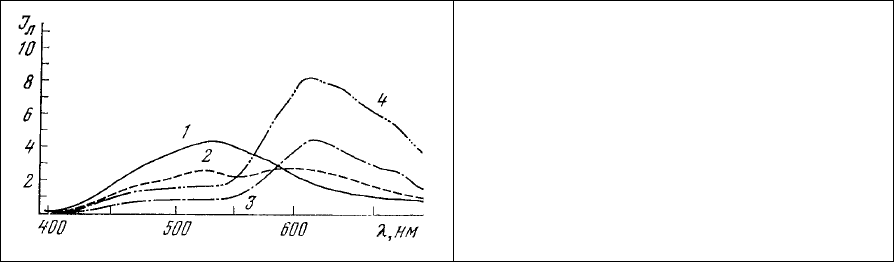
Рис. 92. Изменение спектров
люминесценции грамотрицательных
бактерий Enterobacter cloacae В-69 (1-3)
при повреждении, сопровождающемся
прекращением движения (3). Для сравнения
приведен спектр люминесценции
грамположительных микроорганизмов
Staphylococcus aureus В-83 (4). Длина волны
365 нм.
Имея в виду упомянутые специфические свойства использованных красителей, можно
предполагать, что именно различие в организации клеточной оболочки грамотрицательных и
грамположительных микроорганизмов является причиной наблюдаемого различия в их
люминесцентной окраске. Действительно, появление у живых грамотрицательных микроорганизмов
голубой люминесценции с максимумом излучения 490 нм указывает на то, что только АНС имеет
возможность связываться
с субстратом (гидрофобные области белков) на поверхности этих клеток.
Повреждение белково-липопротеиновой оболочки клеток грамотрицательных микроорганизмов
открывает доступ этидиум бромиду к местам его специфического взаимодействия с молекулами
нуклеиновых кислот, что приводит к появлению характерной для связанного этидиум бромида
дополнительной полосы люминесценции в красной области (рис. 92, 2). При этом по мере
увеличения
интенсивности полосы люминесценции этидиум бромида (605-610 нм) наблюдается
уменьшение интенсивности излучения АНС с максимумом в области 490 нм (рис. 96, 3). По-
видимому, причиной этого является безызлучательная миграция энергии с АНС на этидиум бромид
благодаря почти полному перекрытию полосы излучения АНС (рис. 90, 3) с полосой поглощения
этидиум бромида (рис. 90, 2).
По той же
причине (непроницаемость внешней оболочки для этидиум бромида и отсутствие
специфических для него мест связывания на поверхности) наблюдается голубая люминесценция спор
грамположительных микроорганизмов за счет абсорбции на их поверхности АНС.
Иная ситуация возникает при взаимодействии двух красителей с оболочкой
грамположительных микроорганизмов. В этом случае этидиум бромид имеет доступ к
специфическим для него
местам связывания с рибонуклеиновыми кислотами. Благодаря этому
грамположительные клетки приобретают красную люминесценцию с максимумом излучения в
области 605-610 нм. Возможно, что и АНС адсорбируется на гидрофобных группах белков этих
клеток, однако, упомянутая выше миграция энергии с АНС на этидиум бромид приводит к резкому
ослаблению голубой люминесценции АНС в области 490 нм (рис. 91,
2).
Результаты описанных выше исследований позволяют полагать, что донорно-акцепторная
пара АНС - этидиум бромид представляет собой тонкий инструмент для изучения взаимодействия
белков и нуклеиновых кислот в клетке. Как и следовало ожидать, имея в виду высокую
специфичность взаимодействия каждого из этих красителей, люминесценция не возникает при
добавлении ДНК или РНК к раствору
АНС, так же как и в случае добавления гистона к этидиум
бромиду. При окрашивании ДНК (или РНК) парой АНС - этидиум бромид возникает только
характерная красная люминесценция этидиум бромида. Обработка этой парой меток раствора
гистона приводит к появлению только голубой люминесценции АНС.
Другая картина [335, 336] наблюдается при окрашивании коацерватных капель [337],
образованных системой
ДНК-гистон. Добавление к препарату АНС приводит к появлению голубой
люминесценции его комплексов с гистоном (рис. 93, 1). Дополнительное окрашивание этих же
капель этидиум бромидом вызывает резкое изменение спектра: голубая люминесценция АНС гасится
и появляется красная люминесценция комплексов этидиум бромида с ДНК (рис. 93, 2). Несомненно,
что миграция энергии с АНС на этидиум
бромид лежит в основе регистрируемого эффекта, как и в
описанном выше случае изменения окраски при гибели грамотрицательных клеток.
Интересно отметить, что гашение люминесценции АНС за счет миграции энергии на этидиум
бромид наблюдается и в том случае, когда коацерватные капли (система ДНК - гистон)
приготавливаются из предварительно окрашенных компонентов ДНК + этидиум бромид
и гистон +
АНС. Присутствие меток на макромолекулах не препятствует образованию ими коацерватных
капель, имеющих люминесценцию только в красной области спектра. По-видимому, это означает, что
миграция энергии с АНС, закрепленного на гистоне, на этидиум бромид, связанный с ДНК,
осуществляется вдоль молекулы белка и нуклеиновой кислоты по системе взаимодействующих π-
электронных орбит
этих макромолекул.
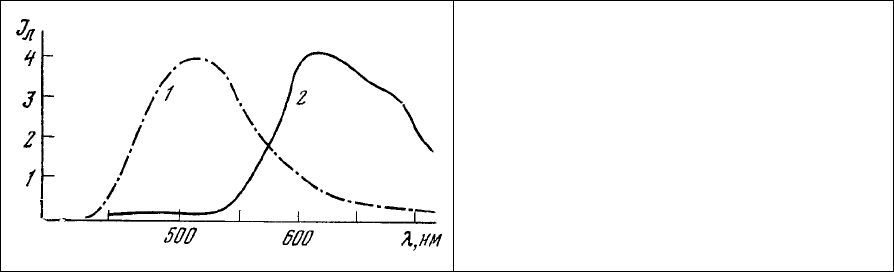
Рис. 93. Спектры люминесценции
коацерватных капель (система ДНК-
гистон), окрашенных вначале АНС (1), а
затем дополнительно этидиум бромидом
(2).
Другим примером использования этого же методического подхода может служить работа
Бекера и соавт. [338], в которой исследовалась полимеризация белков микротрубочек и их связь с
мембранными структурами клетки. При этом один компонент тубулина (белок микротрубочек) или
мембраны окрашивали флуоресцеинизотиоцианатом (донор), а другой - родаминизотиоцианатом
(акцептор). Длина волны возбуждения соответствовала поглощению метки-донора. При
смешивании
приготовленных таким способом растворов в спектре люминесценции помимо полосы излучения
флуоресцеинизотиоцианата (желто-зеленый цвет) появлялась дополнительно и полоса
люминесценции родаминизотиоцианата (красный цвет) за счет миграции энергии от молекулы-
донора к молекуле-акцептору при образовании комплексов тубулин - тубулин и тубулин - мембраны.
Заканчивая данную главу, имеет смысл обратить внимание на возможность использования
некоторых
люминесцентных меток с высокой специфичностью взаимодействия для выяснения молекулярного
состава соответствующих функциональных механизмов клетки. Приведенные выше данные
свидетельствуют о высокой специфичности взаимодействия этидиум бромида именно с
нуклеиновыми кислотами. Оказалось, что этидиум бромид, кроме того, обладает способностью в
малых концентрациях подавлять спайковую электрическую активность нервных клеток,
накапливаясь в их мембранах.
Этот эффект трудно объяснить, не предполагая наличия «скелета» из
нуклеиновой кислоты в составе электрогенных каналов клеточной мембраны, имея в виду и то
обстоятельство, что рибонуклеопротеиновые комплексы из клеточной стенки грамположительных
микроорганизмов послужили одной из первых моделей электрогенных каналов в искусственных
липидных мембранах [339-341].

ПРИЛОЖЕНИЕ
Приборы и техника люминесцентного спектрального
анализа клеток
Введение
Какие бы заманчивые перспективы не открывались в биологии клетки в связи с внедрением в
практику лабораторного эксперимента методов люминесцентного спектрального анализа,
возможность их реализации зависит прежде всего от наличия в распоряжении исследователя
соответствующей техники эксперимента. Важнейшими приборами, необходимыми для этой цели,
являются микроспектрофлуориметры.
В настоящее время большинство исследований на клеточном
уровне выполняется на
приборах, изготовленных непосредственно в лаборатории. Основная причина, определяющая эту
ситуацию, заключается в том, что промышленные приборы обычно весьма консервативны и трудно
поддаются приспособлению их для наиболее оптимального решения конкретных задач.
Поэтому, несмотря на некоторые трудности самостоятельного изготовления
микроспектрофлуориметров, многие исследователи как за рубежом [342-346, 122, 196], так и в нашей
стране
[347-357) идут по пути самостоятельного создания приборов, наиболее полно отвечающих
требованиям поставленной задачи. Такого типа приборы обладают в то же время значительной
гибкостью и легко могут быть модернизированы при смене объекта или цели исследования.
Наиболее рациональным подходом при этом является, по-видимому, максимальное
использование стандартных блоков и узлов [21, 168, 358-365], выпускаемых промышленностью.
Рациональное сочленение стандартных блоков позволяет получить установки, способные решать
достаточно широкий круг задач.
Такой подход может быть проиллюстрирован описанием серии
микроспектрофлуориметров, разработанных и эксплуатировавшихся в течение ряда лет в
Институте биологической физики АН СССР. Важной особенностью этих приборов, как и
большинства отечественных микроспектрофлуориметров, является использование
интерференционных светоделительных пластинок в качестве опак-иллюминатора. Именно
это позволяет применять специальные высокоапертурные микрообъективы и получать
высокую яркость люминесценции микрообъекта, что является особенно важным для
устойчивости работы спектроанализирующего устройства.
Приложение 1
П1. Микроспектрофлуориметры
П1.1. Микроспектрофлуориметр на базе стандартных деталей и узлов
Микроспектрофлуориметр на базе стандартных деталей и узлов [168] является наиболее
простым в изготовлении и заслуживает описания, так как его принципиальная схема, приведенная на
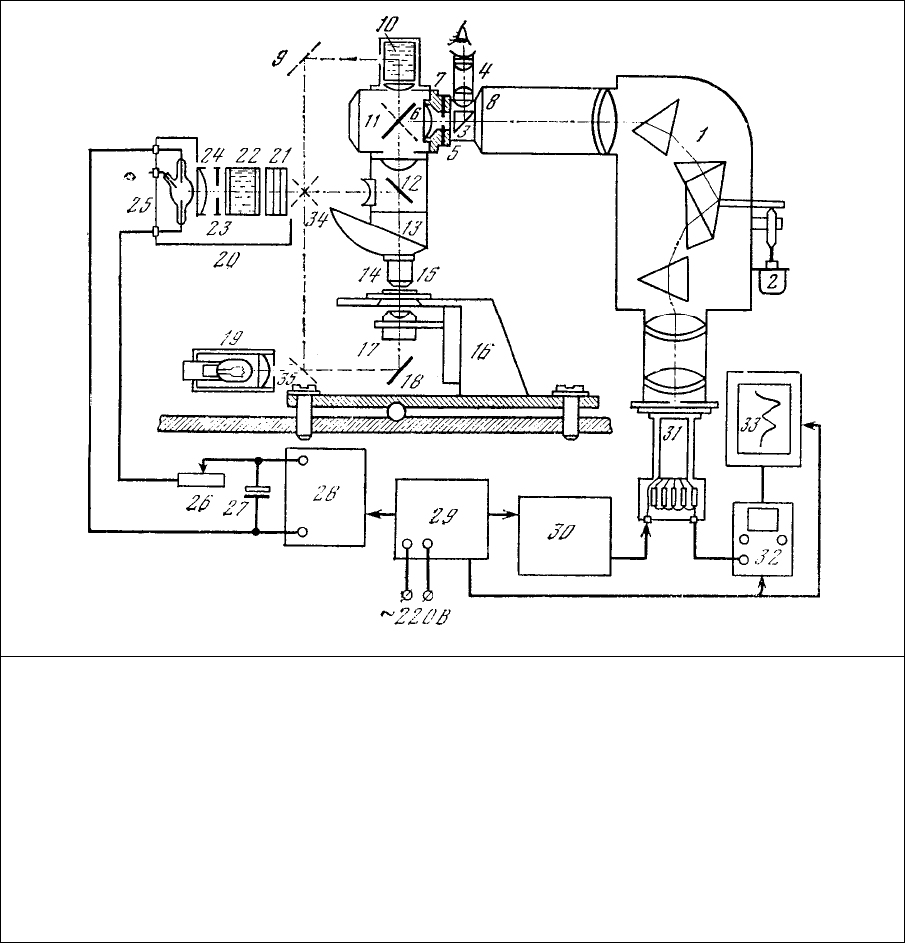
рис. 1.1, послужила основой нескольких интересных модификаций, описываемых ниже.
Основным элементом оптической схемы прибора служит трехпризменный светосильный
спектрограф ИСП-51 со стеклянной оптикой на область спектра от 370 до 1000 нм. Сочленение
спектрографа 1 с микрообъективом 14, достигается тем, что входная щель 7 спектрографа помещена
в плоскость промежуточного изображения микроскопа, состоящего из микрообъектива 14 и окуляра
Гюйгенса 4 (линзы 6 и 8).
Оптическая система микроспектрофлуориметра при регистрации спектров люминесценции
клеток работает следующим образом. Источник света - ртутная дуговая лампа 25 типа ДРШ-250 (или
ДРШ-100) - освещает коллекторную линзу 24, действующие размеры которой ограничиваются
диафрагмой 23. После этого световой поток проходит через водный теплофильтр 22 (4%-ный раствор
СuSO
4
) и светофильтры 21, выделяющие необходимую для возбуждения люминесценции линию
излучения ртути. Все эти детали представляют собой единый узел 20, в качестве которого
используется система освещения люминесцентного осветителя ОСЛ-1 [21]. Возбуждающее
люминесценцию излучение отражается опак-иллюминатором ОИ-17 с интерференционной
светоделительной пластинкой 12 и микрообъективом 14, установленным в стандартном
револьверном держателе от любого
биологического микроскопа, направляется на микрообъект 15.
При этом в плоскость микрообъекта проецируется изображение диафрагмы 23, являющейся, таким
образом, полевой диафрагмой.
Рис. 1.1. Блок-схема микроспектрофлуориметра на базе стандартных деталей и узлов.
1 – призменный монохроматор (ИСП-51);
2 – мотор развертки спектра;
3 – поворотная призма;
4 – визирная трубка;
5 – узел входной щели;
6 – 8 – линзы окуляра Гюйгенса ×4;
7 – входна
я
щель монохроматора (полевая
диафрагма окуляра Гюйгенса);
9, 11, 18, 34, 35 – зеркала;
10 – кювета;
12 – светоделительная пластинка с
интерференционным покрытием;
13 – револьверный держатель
микрообъективов;
14 – объектив;
15 – препарат;
16 – предметный столик;
17 – конденсор;
19 – осветитель с лампой накаливания;
20 – блок осветителя ОСЛ-1 в сборе;
21 – светофильтры;
22 – жидкостный теплофильтр;
23 – ирисовая диафрагма;
24 – коллекторная линза;
25 – дуговая лампа ДРШ-250;
26 – реостат;
27 – блок
электролитических
конденсаторов фильтра;
28 – выпрямитель;
29 – электронный стабилизатор сетевого
напряжения;
30 – стабилизированный источник
высокого напряжения;
31 – фотоумножитель;
32 – милливольтметр (рН-метр ЛПУ-01;
33 – самописцы.
Свет люминесценции препарата, собранный высокоапертурным микрообъективом 14 и
прошедший через пластинку 12 с интерференционным покрытием, поворотным зеркалом 11 (деталь
осветителя ОСЛ-1) и линзой 6, направляется в спектрограф ИСП-51. При этом в плоскости входной
щели 7 спектрографа находится промежуточное люминесцентное изображение микрообъекта. Таким
образом, входная щель 7 спектрографа вырезает требуемый участок увеличенного изображения
исследуемой клетки. Этот участок и его расположение во входной щели спектрографа можно
визуально наблюдать при введении в ход лучей поворотной призмы 3. В этом положении призма 3
вместе с линзами 6 и 8 представляет собой видоизмененный окуляр Гюйгенса ×4, в плоскости
полевой диафрагмы которого размещена входная щель спектрографа.
Промежуточное изображение объекта
исследования в плоскости полевой диафрагмы окуляра
Гюйгенса (входной щели 7 спектрографа) рассматривается с помощью визирной трубки 4 от
фотонасадки типа МФН-1. При этом окончательно выбирается для исследования участок
микрообъекта. Затем поворотная призма 3 выводится из хода лучей и световой поток из входной
щели поступает в колиматор спектрографа. В качестве узла поворотной призмы 3 может быть
использован с незначительной переделкой узел призм насадки сравнения ОКС-1. Свет
люминесценции выбранного участка объекта исследования, ограниченного входной щелью,
разлагается призмами в спектр, изображение которого фокусируется выходным объективом
фотокамеры в плоскость кассеты для фотопластинок, где устанавливается выходная щель
спектрографа
. Непосредственно за ней находится фотокатод фотоэлектронного умножителя типа
ФЭУ-51 со спектральной чувствительностью от 400 до 800 нм.
Таким образом, регистрирующий узел вместе с выходной щелью и кожухом ФЭУ и его
делитель напряжения может быть установлен вместо обычной кассеты для фотопластинок.
Разрешающая способность спектрографа при этом несколько ухудшается, однако для широких линий
люминесценции
биологических объектов это улучшение не столь существенно.
Препарат 15 размещается на предметном столике 16 от микроскопа МС-51. С помощью
конденсора 17, зеркала 18 и осветителя 19 (типа ОИ-19) возможно освещение препарата проходящим
светом от лампы накаливания. Такое освещение может быть полезным для предварительного выбора
участка клетки без излишнего освещения его лучами
сине-фиолетовой или УФ-области спектра.
Излучение этого же источника света, выделенное узкополосным интерференционным
светофильтром, может быть использовано для контроля рассеивающих свойств изучаемой области
объекта, как это упоминалось выше. Для облегчения юстировки столика относительно оптической
оси микроскопа он имеет одну точку опоры (стальной шарик) и три регулировочных винта.
Описываемый прибор
позволяет изучать также спектры люминесценции раствора в кюветах.
Для этого возбуждающее люминесценцию излучение может направляться дополнительными
зеркалами 34 и 9 на кювету 10 с исследуемым веществом. Поворотное зеркало 11 переключается при
этом в верхнее положение и направляет свет люминесценции кюветы во входную щель 7
спектрографа. На практике, однако, особенно при наличии контактных
микрообъектов [21], удобнее
бывает производить эти исследования в микрокюветах, размещенных на предметном столике вместо
микропрепарата 15. Развертка спектра в описываемой первой модели микроспектрофлуориметра
осуществлялась мотором 2 типа РД-0,9, на валу которого были закреплены контакты метчика длин
волн.
Электронная схема микроспектрофлуориметра была построена также по принципу
максимального использования доступных стандартных приборов. Регистрация
спектров
люминесценции производилась фотоумножителем 31 типа ФЭУ-51, питающимся от
стабилизированного источника напряжения 30 типа ВС-22. Сигнал с сопротивления нагрузки ФЭУ
подавался на вход рН-метра 32 типа ЛПУ-01, используемого в режиме высокоомного нуль-
индикатора, на выходные клеммы которого подключался электронный самопишущий
милливольтметр 33 типа ЭПП-09 (КСП-4). Таким образом, в этой системе
регистрации рН-метр
ЛПУ-01 играет роль буферного каскада для согласования высокого выходного сопротивления
фотоумножителя с низким входным сопротивлением самописца.
Поступающий на вход ЛПУ-01 сигнал постоянного тока с выходного сопротивления ФЭУ
модулируется вибропреобразователем, и дальнейшее усиление его по мощности осуществляется
усилителями переменного тока, лишенными такой неприятной особенности, как дрейф нуля.
Регистрирующая система
на базе ЛПУ-01 работает исключительно стабильно и надежно.
Компенсация темнового тока ФЭУ осуществляется ручкой «настройка но буферному раствору» на
лицевой панели прибора после предварительной установки пределов компенсации ручкой «рН» под
откидной крышкой на боковой панели прибора. Ручка «крутизна» может быть использована для
регулировки усиления сигнала после предварительной калибровки ее положения,
однако более
удобно изменять чувствительность, используя набор калиброванных сопротивлений нагрузки ФЭУ
при установленной в крайнее по часовой стрелке положение ручки «крутизна» на лицевой панели
ЛПУ-01. Одна из выходных клемм ЛПУ-01, к которым подключается самописец 33, периодически
замыкается на землю контактами метчика длин волн. Ртутная дуговая лампа 25 типа ДРШ-250
питается постоянным
током от выпрямителя 28 типа ВСА-III через реостат 26 (R =30 Ω). На выходе
выпрямителя включен электролитический конденсатор 27 емкостью 800 мкФ на рабочее напряжение
300 В для сглаживания пульсаций напряжения.
Существенным элементом схемы является электронный стабилизатор сетевого напряжения 29
типа St = 2000 (Tesla), от которого питаются все элементы электронной схемы
микроспектрофлуориметра. Применение этого стабилизатора позволяет снизить, по
крайней мере, на
порядок пульсации светового потока ртутной лампы без использования сложных систем
стабилизации светового потока.
Ценным качеством описанной выше базовой модели является простота ее изготовления из
узлов по сути дела снятых с вооружения современных лабораторий оптических приборов. Обладая
высокой чувствительностью, эта модель микроспектрофлуориметра может быть использована для