Zelditch M.L. (и др.) Geometric Morphometrics for Biologists: a primer
Подождите немного. Документ загружается.

chap-11 4/6/2004 17: 27 page 268
268 GEOMETRIC MORPHOMETRICS FOR BIOLOGISTS
(A)
(B)
1
2
3
12
13
14
15
16
10
9
4
5
6
7
8
11
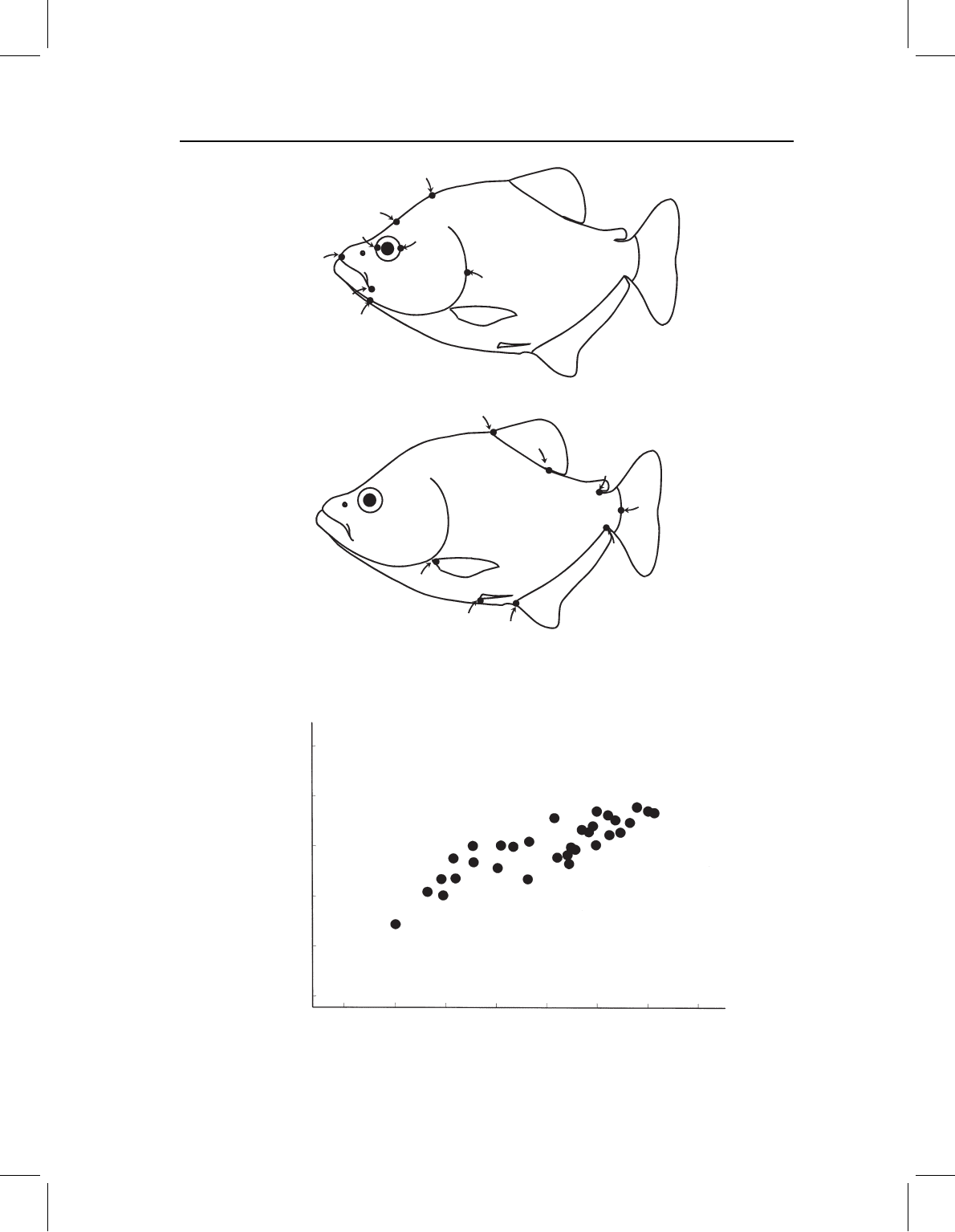
Figure 11.1 Subdivision of landmarks into two blocks: (A) cranial configuration; (B) postcranial
configuration.
⫺0.20 ⫺0.15 ⫺0.10 ⫺0.05 0 0.05 0.10 0.15
⫺0.15
⫺0.10
⫺0.05
0
0.05
0.10
SA1 (cranial)
SA1 (postcranial)
Figure 11.2 Scores of postcranial SA1 on cranial SA1 for P. denticulata.

chap-11 4/6/2004 17: 27 page 269
PARTIAL LEAST SQUARES ANALYSIS 269
1
2
3
4
5
6
7
8
9
10
11
12
13
14 15
16
Figure 11.3 Singular axis expressing the covariance between cranial and postcranial landmarks of
P. denticulata.
of ontogenetic integration using regression because that technique allows us to find the
shape variable correlated with size, which ought to explain the covariance between cranial
and postcranial landmarks. We would also expect PCA to yield the same result because
size is the dominant component of the variance in an ontogenetic series.
Despite such apparently realistic expectations, we find that regression and PCA give
similar results (Figures 11.4A, 11.4B), but they appear to differ from those yielded by
PLS (Figure 11.4C). However, the visual comparison of graphics is complicated for three
reasons. First, the analysis by PLS examines two partly overlapping parts rather than a
single whole, in contrast to PCA and regression. Second, when analyzed by PLS, each half
is separately scaled to unit centroid size rather than the whole, so the relative sizes of each
block are portrayed differently in the two analyses. Third, when analyzed by PLS, the
magnitude of change within each part is calculated and scaled separately rather than in
relation to the whole as done by methods that analyze the entire configuration of landmarks
comprising both blocks.
Proceeding with the visual comparison nonetheless, one apparent difference is the bal-
ance between cranial and postcranial changes. Regression and PCA (of the whole fish)
both suggest that there is a large change in the orbital region relative to the change of the
posterior body, but PLS suggests that they are more equally balanced. A more troubling
difference is that the posterior covariate of size found by regression or PCA looks quite

chap-11 4/6/2004 17: 27 page 270
270 GEOMETRIC MORPHOMETRICS FOR BIOLOGISTS
(A)
(B)
(C)
Figure 11.4 Comparing the direction of ontogenetic change in P. denticulata as determined by:
(A) regression of whole body shape on size; (B) PC1 of whole body shape; (C) SA1 of the axis of
covariance between cranial and postcranial shape extracted by PLS.
different than posterior SA1 – it appears that the most caudal region (the caudal peduncle)
is integrated with the posterior anal fin, and more change is localized here (relative to the
remainder of the posterior body).
The consequence of analyzing each half separately can be appreciated by regressing each
block separately on size, and also by using PCA to extract the dominant component of
variance of each separate half. Regression of each part separately on size yields a result
chap-11 4/6/2004 17: 27 page 271
PARTIAL LEAST SQUARES ANALYSIS 271
(A)
(B)
(C)
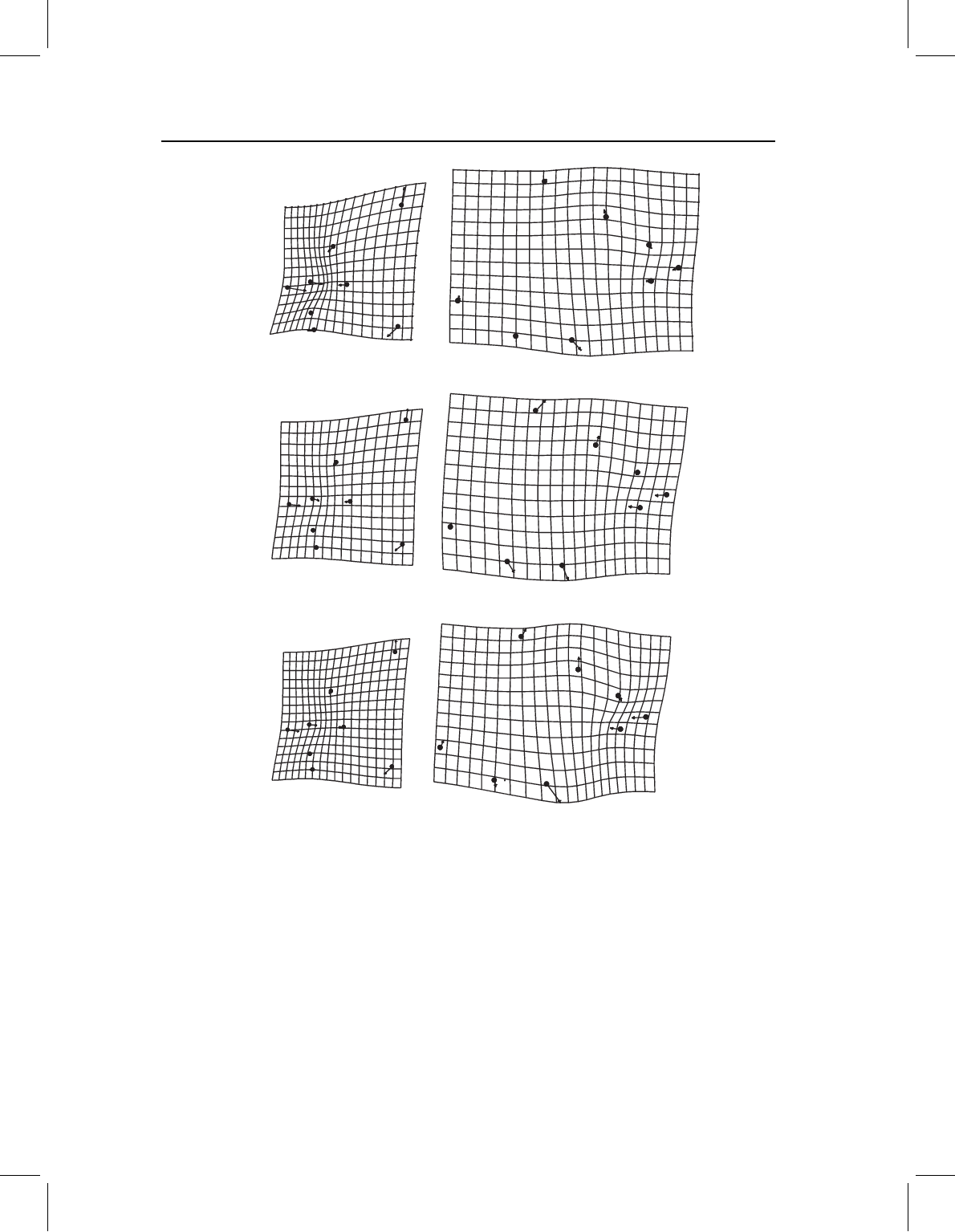
Figure 11.5 Comparing results of three methods for analyzing the two blocks of landmarks: (A)
regression of P. denticulata cranial and postcranial landmarks separately on size; (B) PC1 of cranial
and postcranial landmarks analyzed separately; (C) cranial and postcranial SA1.
very similar to that of regression of the whole (compare Figures 11.4A and 11.5A). Each
block appears to be nearly linearly related to size, and the correlations between each block
and size equal the correlations between each SA1 and size (i.e. 0.89, 0.72 for cranial and
postcranial blocks, respectively). There is, however, a notable difference between the two
blocks in the magnitude of change in relation to size. Measuring the rate at which the
Procrustes distance away from the smallest specimen increases with size gives strikingly
different estimates for the two blocks: 0.08 for the cranial block and only 0.03 for the
postcranial block (Figure 11.6). Thus, over a given change in size, the cranial region
undergoes far more change than the postcranial body. Such information is captured by

chap-11 4/6/2004 17: 27 page 272
272 GEOMETRIC MORPHOMETRICS FOR BIOLOGISTS
3.0 3.5 4.0 4.5 5.0 5.5 6.
0
LCS
LCS
0
0.05
0.10
0.15
0.20
0.25
D (cranial)D (postcranial)
3.0 3.5 4.0 4.5 5.0 5.5 6.0
0.01
0.02
0.03
0.04
0.05
0.06
0.07
0.08
0.09
0.10
0.11
Figure 11.6 Relative rates of cranial and postcranial development analyzed by regressing the
Procrustes distance away from the average of the smallest specimens (D) against log centroid size
(LCS).
regression even when the parts are analyzed separately, because the changes are calibrated
in relation to size. Going back to the plot of the SA1 scores (Figure 11.2), it is now possible
to appreciate the effect of these different magnitudes of response by noting the larger range
of scores for cranial SA1 scores compared to postcranial SA1 scores.
PCA of the separate blocks yields one distinct eigenvalue for the cranial landmarks and
none for the postcranial block, which is not surprising in light of their different magnitudes
of change. Because of the larger ontogenetic change in the cranial region there is a larger
distance between the shapes of the smallest and largest specimens, and consequently varia-
tion is more elliptical because ontogeny produces a long axis of variation (correlated with
size). In contrast, the ontogenetic change of the postcranial region is subtle, so the distance
between the smallest and largest specimen is not as large, and the postcranial variation is

chap-11 4/6/2004 17: 27 page 273
PARTIAL LEAST SQUARES ANALYSIS 273
(A)
(B)
(
C
)
Figure 11.7 Comparing results of three methods for analyzing the two blocks of landmarks, with
PLS results rescaled in light of relative rates of development. (A) Regression of P. denticulata cranial
and postcranial landmarks separately on size; (B) PC1 of cranial and postcranial landmarks analyzed
separately; (C) SA1 of cranial and postcranial shape.
more nearly spherical. There is no dominant size axis, although size is still a factor explain-
ing variation within the postcranium. When analyzed as part of the whole, we can see the
impact of size on the postcranium in the context of its effect on the cranium, an effect
that becomes more ambiguous when the blocks are analyzed separately (compare Figures
11.4B, 11.5B). However, that ambiguity is again partly due to the greater magnitude of
change undergone by the head.
We can rescale the plots of SA1, amplifying the deformation of the head to reflect
its greater developmental rate (Figure 11.7C). That does not fully reconcile the graphical
results of regression, PLS and PCA, but it removes one major discrepancy among them. The

chap-11 4/6/2004 17: 27 page 274
274 GEOMETRIC MORPHOMETRICS FOR BIOLOGISTS
(
A
)(
B
)
10
9
4
5
6
7
8
11
1
2
3
12
13
14
15
16
Figure 11.8 Comparing SA1 (solid lines) to PC1 (dotted lines).
changes within the head appear to be similar now, regardless of method. The similarity
between PCA and PLS is even more evident if we diagram PC1 and SA1 on the same
plot (Figure 11.8). The difference is most striking for the posterior block, where it is most
notable in the orientation of the vectors at the two anal fin landmarks (landmarks 8 and 9).
To check the generality of the conclusions we drew from the analysis of P. denti-
culata, we can (more briefly) analyze two other species. One, S. gouldingi, differs from
P. denticulata in that there are two distinct eigenvalues for the variation of the head
(rather than one) and there is one for the variation of the postcranial landmarks (rather
than none). In the analysis of S. gouldingi the first singular value is very high (0.2379),
accounting for 89% of the covariance between the two blocks, and the correlation is
also remarkably high (0.968). Not surprisingly, the covariance explained by the paired
SA1 axes is significant (p < 0.01), as is the correlation (p < 0.01). No other SA explains
more covariance than expected by chance, so these data, like those of P. denticulata, pro-
duce a one-dimensional solution. Having already found that differences in relative rates
of development between the blocks can produce apparent discrepancies among the results
of the three methods, we show the results taking the difference in cranial and postcranial
developmental rates into account. The primary discrepancy among the results of the three
methods is in their descriptions of posterior head deepening (Figure 11.9). There is virtu-
ally no difference between the results of PCA and PLS, to the point that we cannot visually
compare them by superimposing the two sets of vectors on the same plot – they entirely
overlap each other. The results of PCA and PLS do differ from those of regression, albeit
subtly.
The analysis of S. manueli, like that of the other two species, yields a single significant
dimension of covariation between blocks. SA1 accounts for 76% of the covariance between
chap-11 4/6/2004 17: 27 page 275
PARTIAL LEAST SQUARES ANALYSIS 275
(B)
(A)
(C)
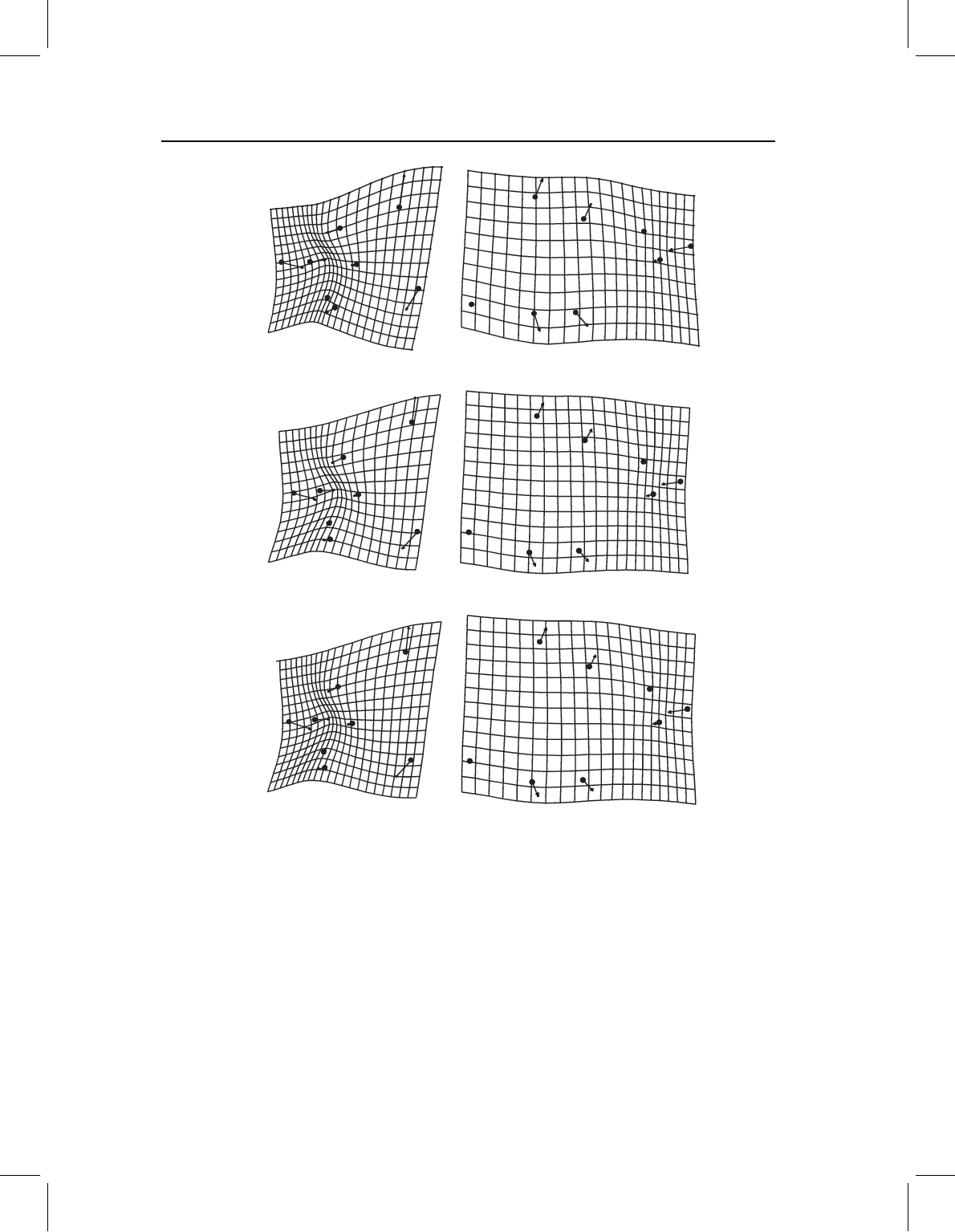
Figure 11.9 Comparing results of three methods for analyzing the two blocks of landmarks after
rescaling plots to reflect the magnitude of the change of each block relative to a unit change in
size. (A) Regression of S. gouldingi cranial and postcranial landmarks separately on size; (B) PC1 of
cranial and postcranial landmarks analyzed separately; (C) cranial and postcranial SA1.
blocks, which is significantly greater than expected by chance (p < 0.01). The correlation
between blocks is 0.92, which is also significant (p < 0.01). In drawing the SAs, we again
scale the magnitudes of the cranial and postcranial blocks in accordance with their relative
rates of development (Figure 11.10). As in the case of S. gouldingi, regression provides a
somewhat different picture of the ontogenetic change in shape than do PLS and PCA, but
the results of PCA and PLS are consistent with each other.
The possibility that the three methods can give different results underscores the
importance of deciding which methods ought to be used.

chap-11 4/6/2004 17: 27 page 276
276 GEOMETRIC MORPHOMETRICS FOR BIOLOGISTS
(A)
(B)
(C)
Figure 11.10 Comparing results of three methods for analyzing the two blocks of landmarks after
rescaling plots to reflect the magnitude of the change of each block relative to a unit change in size.
(A) Regression of S. manueli cranial and postcranial landmarks separately on size; (B) PC1 of cranial
and postcranial landmarks analyzed separately; (C) cranial and postcranial SA1.
Interspecific comparisons of ontogenetic integration
We can use PLS to compare ontogenetic integration among species. This is another case
where we could also use a regression-based approach (as we did in Chapter 10), so we
focus on the distinction between the results of the two methods. To make the analyses as
similar as possible, we subdivide the landmarks into cranial and postcranial landmarks in
the analyses based on regression, just as we do for the analyses based on PLS.
In the comparison between P. denticulata and S. gouldingi, the interspecific angle
between cranial SV1s is small (13.8
◦
), suggesting that these species are virtually indis-
tinguishable in cranial ontogeny; not surprisingly, that angle is not statistically significant.
More surprisingly, the larger angle of 30.9
◦
between postcranial SA1s is also not significant

chap-11 4/6/2004 17: 27 page 277
PARTIAL LEAST SQUARES ANALYSIS 277
(owing to the larger range of angles obtained by resampling within S. gouldingi – 32.8
◦
).
Comparing the cranial regressions also yields a modest angle (12.8
◦
) that is not statisti-
cally significant. However, the interspecific angle between postcranial ontogenies is larger
(44.0
◦
), and this is statistically significant. Thus we can conclude that the two species
share a common cranial ontogeny, but the results for the postcranial landmarks are more
ambiguous. Based on PLS, we cannot say that the patterns of postcranial integration differ
between species. However, if we analyze the relationship between the postcranial land-
marks and size, we do find a significant difference between species. In this case, it is
important to decide whether the hypothesis ought to be formulated in terms of PLS or
regression.
In the comparison between P. denticulata and S. manueli, we find a relatively large
angle of 39.2
◦
between SA1 of the cranial landmarks, which is statistically significant,
and an equally large angle for the postcranial landmarks of 40.2
◦
, which is also sig-
nificant. Turning to the comparison of their cranial ontogenetic allometries based on
regression, we find an interspecific angle of 45.4
◦
between cranial landmarks, and an
angle of 51.1
◦
for the postcranial landmarks. In this case, both methods detect statis-
tically significant differences between species in both blocks of landmarks. Similarly, in
the comparison between S. manueli and S. gouldingi, the results from both methods are
consistent. Comparing cranial SV1s between species yields an angle of 43.4
◦
, which is
statistically significant, and a comparably large angle between postcranial SV1s of 30.3
◦
,
which is also significant. The analysis based on vectors of allometric coefficients yields
an interspecific cranial angle of 46.8
◦
and postcranial angle of 32.7
◦
, both of which are
statistically significant. Unfortunately we cannot assume that the results will always be
consistent, as they were not in the comparison between P. denticulata and S. gouldingi.
Thus, as in the analyses of intraspecific integration, it is important to decide whether the
analysis ought to be based on regression or on PLS.
Using PLS to test competing hypotheses of integration
Our objective now is to formulate competing hypotheses of integration and use PLS to
test them. Specifically, we ask whether the integration between the cranial and caudalmost
landmarks is greater than that between the cranial and median fin landmarks (the three
blocks are depicted in Figure 11.11). We might expect that this would be the case, because
the head and caudal body usually develop earlier than the midbody, and the deepening
of the midbody occurs fairly late in development. Any factors, both genetic and environ-
mental, that affect larval development are likely to affect both these cranial and caudal
regions, but might have little impact on midbody depth (or anteroposterior locations of
the median fins). Therefore, if the timing of development explains integration, we might
anticipate a greater correlation between parts that develop at the same time. We will test
this hypothesis using two species. In one, S. gouldingi, the general expectations appear
to be met (for reasons that will not be evident until Chapter 13, when we discuss the
relationship between allometric coefficients and developmental timing). In the other, the
caudalmost part of the body seems to develop unusually late in relation to the head, so
we would expect that this species would not evince greater integration between head and
tail than between head and fin/midbody landmarks.