Вестхайде В.,Ригер Р.( ред.) Зоология беспозвоночных в двух томах. Том 1: от простейших до моллюсков и артропод
Подождите немного. Документ загружается.


J
Илл. 664.
Palpigradi.
Eukoenenia
sp.
Длина 2 мм. Из
Weygoldt
(1972).
модифицированы. Размножение, развитие и число
линек у нимф не изучены; редкая встречаемость сам-
цов у некоторых видов указывает на партеногенез.
Систематика
Архаичные признаки, такие как трёхчленико-
вые хелицеры и наличие хвостовой нити, указывают
на то, что безлёгочные кенении ответвились от об-
щего ствола Lipoctena ещё до отделения от него Me-
goperculata. С этой точки зрения Palpigradi представ-
ляют собой сестринскую группу всех безлёгочных
арахнид (альтернативное мнение — см. илл. 613).
*Eukoenenia
mirabilis (Eukoeneniidae), кенения уди-
вительная, длина
2
мм. Обитает в просветах между части-
цами почвы и под камнями в Южной Европе до Австрии.
Другие виды распространены
в
тропиках и субтропиках, а
в Центральной Европе встречаются в оранжереях. Неко-
торые виды живут между частицами грунта на морских
берегах.
Arthropoda
борозды
на
спинной
[
стороне
'
просомы
педипальпа
-\^скутум
I
тергит
генитальное
отверстие
X
тергит
анальный
сегмент
коксальныи
мешочек
стернит
IV
стернит
X
стернит
Илл.
665.
Chelifer cancroides
(Pseudoscorpiones). Внешнее строение.
А — вид со спинной стороны. Б — вид с брюш-
ной стороны. По Beier из Weygoldt (1969).
3.2.6.
Pseudoscorpiones (Chelonethi),
Ложноскорпионы
Более 3
ООО
видов мелких (1-7 мм) животных,
напоминающих по габитусу скорпионов (илл. 665)
из-за крупных педипальп с клешнями. Опистосома
у них, однако, цельная, а не разделённая на мезо- и
метасому. Во время движения педипальпы вытяги-
ваются вперёд и служат щупиками. На них находят-
ся такие важные органы чувств, как 12 трихоботрии,
расположение которых видоспецифично. Кроме них,
у ложноскорпионов есть такие органы чувств, как
механо- и хеморецепторные щетинки, щелевидные
органы, немногочисленные лировидные органы и
одна-две пары простых боковых глаз.
Ложноскорпионы живут на почве в подстилке,
под корой и в других узких щелях. Они населяют все
сухопутные местообитания, включая морскую суп-
ралитораль. Некоторые виды распространяются за
счёт их переноса мухами, жуками и другими насеко-
мыми, а также мелкими млекопитающими. Некото-
рые виды сожительствуют с тропическими жуками,
под надкрыльями которых они могут не только пе-
реноситься, но и спариваться.
Добыча — мелкие членистоногие — схватыва-
ется и умерщвляется клешнями педипальп. На кон-
цах одной или обеих створок клешней открываются
каналы ядовитых желёз (илл. 666). Виды с больши-
ми хелицерами (Chthoniidae, Neobisiidae) мацериру-
ют свою пищу, виды же с маленькими хелицерами
(Chernetidae, Cheliferidae) лишь прогрызают отвер-
стие и высасывают жертву. Преддверие рта образо-
вано выростами тазиков педипальп. Маленький рот
ведёт в сосательную глотку с Х-образным внутрен-
ним просветом и далее в узкий пищевод. В передний
отдел средней кишки впадают протоки объёмистой
печени. Задний отдел средней кишки образует пет-
лю из трёх колен, проходящих параллельно друг дру-
гу, расширяется в ректальный пузырь и переходит в
короткую заднюю кишку с анусом на конце тела.
http://jurassic.ru/
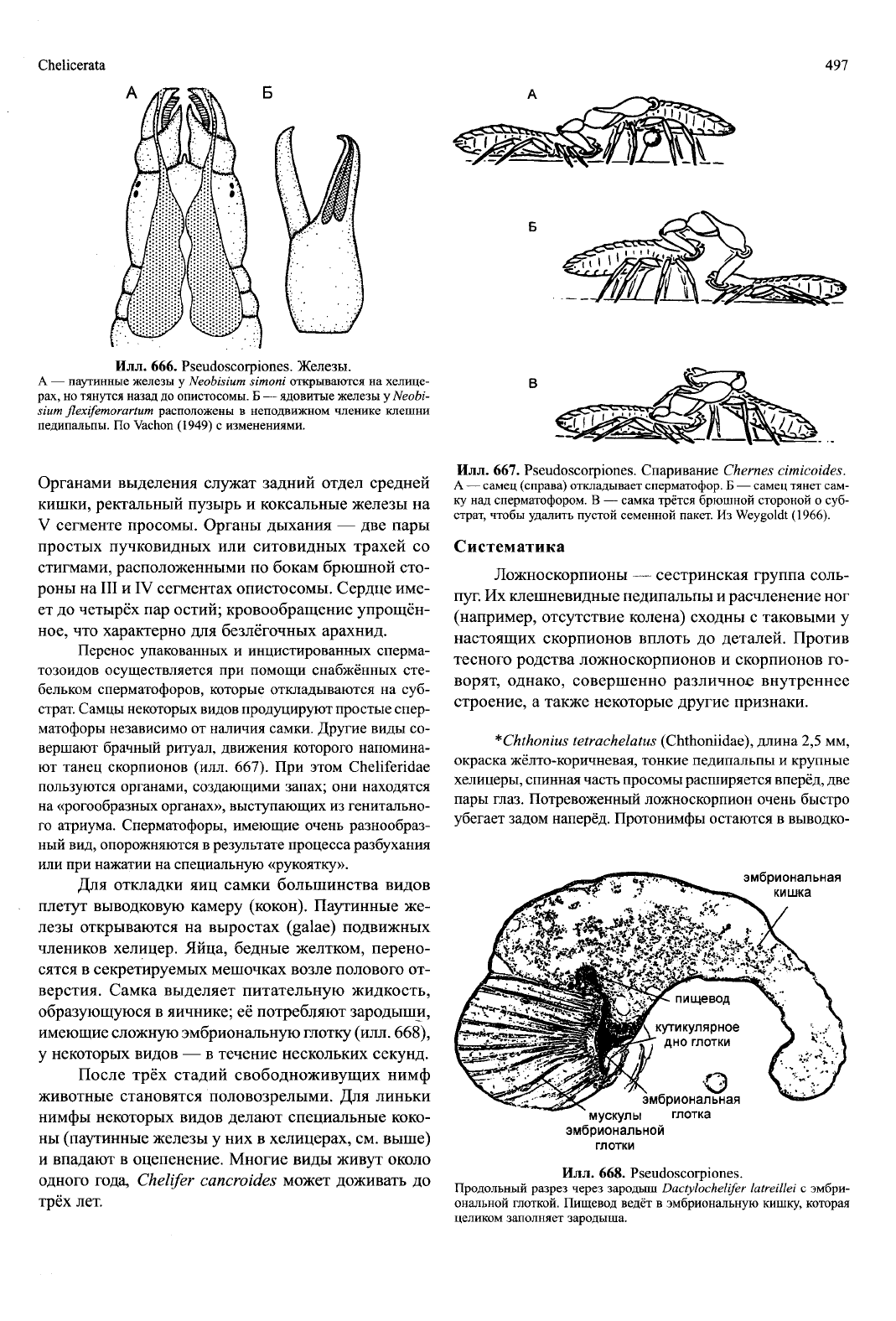
Chelicerata
Илл.
666.
Pseudoscorpiones. Железы.
A — паутинные железы у Neobisium simoni открываются на хелице-
рах, но тянутся назад до опистосомы. Б — ядовитые железы у Neobi-
sium flexifemorartum расположены в неподвижном членике клешни
педипальпы. По Vachon (1949) с изменениями.
Органами выделения служат задний отдел средней
кишки, ректальный пузырь и коксальные железы на
V сегменте просомы. Органы дыхания — две пары
простых пучковидных или ситовидных трахей со
стигмами, расположенными по бокам брюшной сто-
роны на III и IV сегментах опистосомы. Сердце име-
ет до четырёх пар остий; кровообращение упрощён-
ное,
что характерно для безлёгочных арахнид.
Перенос упакованных и инцистированных сперма-
тозоидов осуществляется при помощи снабжённых сте-
бельком сперматофоров, которые откладываются на суб-
страт. Самцы некоторых видов продуцируют простые спер-
матофоры независимо от наличия самки. Другие виды со-
вершают брачный ритуал, движения которого напомина-
ют танец скорпионов (илл. 667). При этом Cheliferidae
пользуются органами, создающими запах; они находятся
на «рогообразных органах», выступающих из генитально-
го атриума. Сперматофоры, имеющие очень разнообраз-
ный вид, опорожняются в результате процесса разбухания
или при нажатии на специальную «рукоятку».
Для откладки яиц самки большинства видов
плетут выводковую камеру (кокон). Паутинные же-
лезы открываются на выростах (galae) подвижных
члеников хелицер. Яйца, бедные желтком, перено-
сятся в секретируемых мешочках возле полового от-
верстия. Самка выделяет питательную жидкость,
образующуюся в яичнике; её потребляют зародыши,
имеющие сложную эмбриональную глотку (илл. 668),
у некоторых видов — в течение нескольких секунд.
После трёх стадий свободноживущих нимф
животные становятся половозрелыми. Для линьки
нимфы некоторых видов делают специальные коко-
ны (паутинные железы у них в хелицерах, см. выше)
и впадают в оцепенение. Многие виды живут около
одного года, Chelifer cancroides может доживать до
трёх лет.
497
Илл.
667. Pseudoscorpiones. Спаривание Cherries cimicoides.
А — самец (справа) откладывает сперматофор. Б — самец тянет сам-
ку над сперматофором. В — самка трётся брюшной стороной о суб-
страт, чтобы удалить пустой семенной пакет. Из Weygoldt (1966).
Систематика
Ложноскорпионы — сестринская группа соль-
пуг. Их клешневидные педипальпы и расчленение ног
(например, отсутствие колена) сходны с таковыми у
настоящих скорпионов вплоть до деталей. Против
тесного родства ложноскорпионов и скорпионов го-
ворят, однако, совершенно различное внутреннее
строение, а также некоторые другие признаки.
*Chthonius tetrachelatus
(Chthoniidae), длина 2,5 мм,
окраска жёлто-коричневая, тонкие педипальпы и крупные
хелицеры, спинная часть просомы расширяется вперёд, две
пары глаз. Потревоженный ложноскорпион очень быстро
убегает задом наперёд. Протонимфы остаются в выводко-
эмбриональной
глотки
Илл.
668.
Pseudoscorpiones.
Продольный разрез через зародыш Dactylochelifer latreillei с эмбри-
ональной глоткой. Пищевод ведёт в эмбриональную кишку, которая
целиком заполняет зародыша.
http://jurassic.ru/

498
Arthropoda
вой камере и кормятся питательной жидкостью, которой
их снабжает мать. Обитают в опавшей листве широколи-
ственных лесов или под камнями. —
*Neobisium muscorum
(Neobisiidae), длина
4 мм,
блестящий буровато-чёрный лож-
носкорпион, живёт в подстилке лесов (в Германии). Хели-
церы крупные, боковые края просомы параллельны между
собой, две пары глаз. —
N.
maritimum
обитает на морской
литорали. —
Gary-pus beauvoisi
(Garypidae), длина
до 7
мм,
хелицеры мелкие. Обитает на супралиторали Средиземно-
го моря. —
*Cheiridium
museorum (Cheiridiidae), длина 1
мм,
одна пара глаз, живёт в сараях и гнёздах воробьев. —
*Lasiochernes
pilosus (Chernetidae), длина 4 мм, глаз нет.
Обитает в норах кротов и полёвок, расселяется, прикреп-
ляясь к шерсти хозяина. —
*Chernes
cimicoides, длина 3
мм,
живёт под корой
деревьев.
— *Chelifer
cancroides
(Che-
liferidae), книжный ложноскорпион, длина
4-5 мм,
распро-
странён всесветно, живёт в постройках человека, в ульях
и под корой.
3.2.7.
Solifugae (Solpugida),
Сольпуги
Около 900 видов сольпут, длина которых варьи-
рует от 10 до 70 мм, живут в основном в засушливых
областях, пустынях и степях. В облике этих паукооб-
разных примечательны очень крупные двучлениковые
хелицеры, подвижная просома
с
пропельтидием и дву-
мя-тремя пряжковидными склеритами, а также ци-
линдрическая опистосома с мягкими покровами и
одиннадцатью мелкими тергитами (илл. 669).
Сольпуги подолгу и быстро бегают на
трёх задних парах ходильных ног; ноги I
пары развиты слабее
и
служат
щупиками.
На
отвесных гладких поверхностях используют-
ся педипальпы, на которых терминально рас-
положен клейкий орган. Благодаря нему
животные могут передвигаться, находясь в
подвешенном состоянии. Вес виды хорошо
копают, используя при этом также и силь-
ные хелицеры.
Органами чувств служат средин-
ные глаза на пропельтидии, одна-две
пары уменьшенных боковых глаз, а так-
же длинные осязательные щетинки на
ногах и педипальпах, погружённые чув-
ствительные щетинки (sensilla ampu-
lacea) и по пять крупных молотовидных
органов (malleoli, маллеолы, привес-
ки) на вентральной стороне прокси-
мального членика последней пары хо-
дильных ног с многочисленными (до
72 ООО) сенсорными нервными оконча-
ниями; вероятно, они используются как
хеморецепторы.
При опасности сольпуга поднимает
переднюю часть просомы, угрожая больши-
ми хелицерами. Некоторые виды при этом
стрекочут, потирая хелицеры друг о друга.
Илл. 669.
Galeodes
sp. (Solifugae). Длина около 40 мм.
Оригинал P. Weygoldt, Фрейбург.
Многие виды — крупные подвижные хищни-
ки,
часть из них активны днём. Добычей сольпугам
служат крупные членистоногие и мелкие позвоноч-
ные.
Жертва, схваченная педипальпами, быстро из-
мельчается и мацерируется на хелицерах. Ядовитых
желёз нет. Рот расположен на роструме между верх-
ней и нижней губой; преддверие рта отсутствует.
Органами выделения служат одна пара мальпигие-
вых трубочек, а также нефроциты и коксальные же-
лезы, каналы которых открываются позади тазиков
педипальп. Для дыхания служит система трахей, уни-
кальная для арахнид и сходная с таковой у насеко-
мых (илл. 670). Парные стигмы между III и IV пара-
пропельтидий
метапельтидии
мезопельтидий
сегмент
опистосомы
стигма
на
просоме
стигмы
на I
опистосомном
сегменте
непарная
стигма
на V
опистосомном
стигмы
на IV
сегменте
опистосомном
сегменте
Илл. 670. Solifugae.
А — передний конец тела Galeodes graecus, вид сбоку. Педипальпы и ноги удале-
ны.
Б — схема трахейной системы. По Kaestner (1932).
http://jurassic.ru/
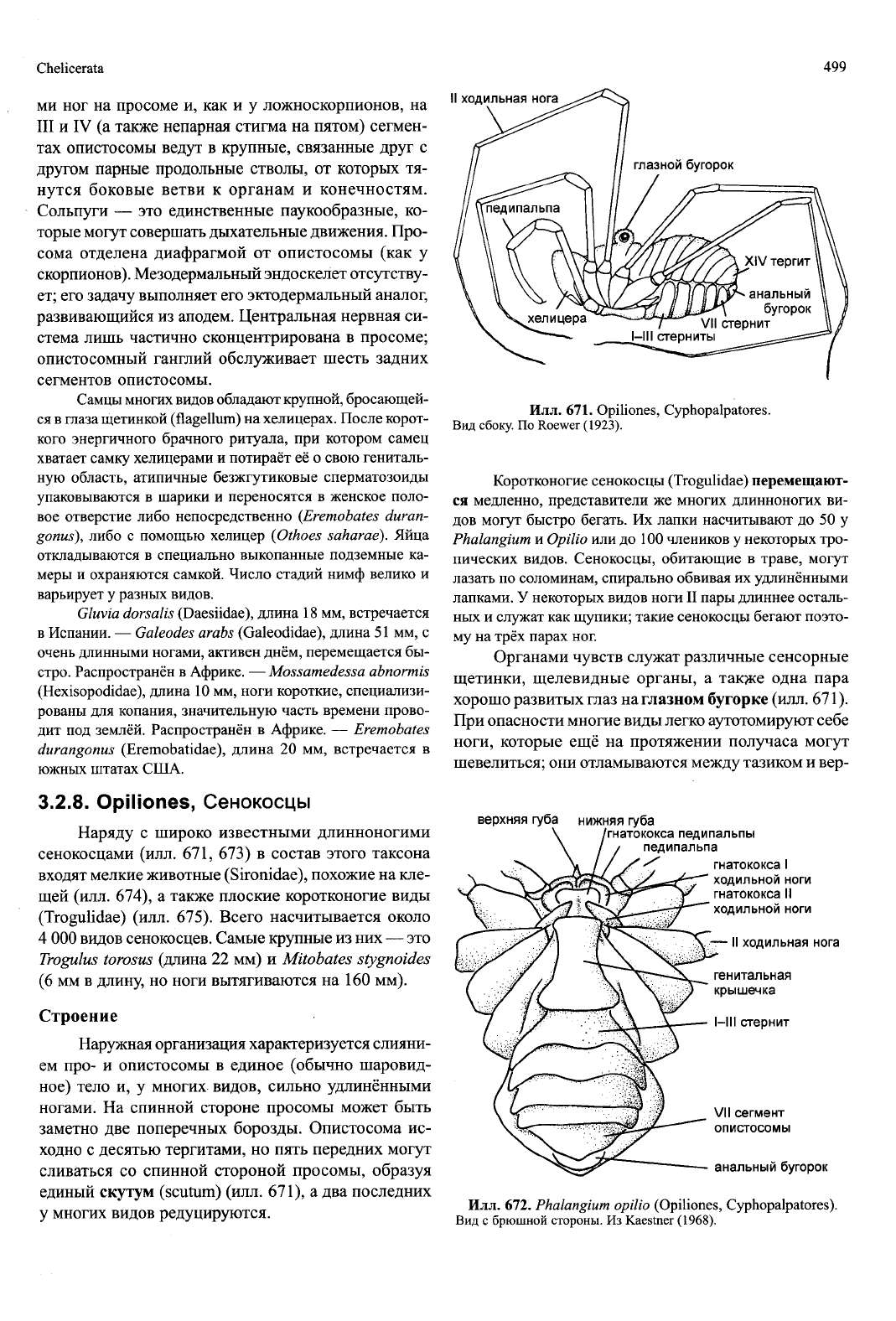
Chelicerata
499
ми ног на просоме и, как и у ложноскорпионов, на
III и IV (а также непарная стигма на пятом) сегмен-
тах опистосомы ведут в крупные, связанные друг с
другом парные продольные стволы, от которых тя-
нутся боковые ветви к органам и конечностям.
Сольпуги — это единственные паукообразные, ко-
торые могут совершать дыхательные движения. Про-
сома отделена диафрагмой от опистосомы (как у
скорпионов). Мезодермальный эндоскелет отсутству-
ет; его задачу выполняет его эктодермальный аналог,
развивающийся из аподем. Центральная нервная си-
стема лишь частично сконцентрирована в просоме;
опистосомный ганглий обслуживает шесть задних
сегментов опистосомы.
Самцы многих
видов
обладают крупной, бросающей-
ся
в
глаза щетинкой (flagellum) на
хелицерах.
После корот-
кого энергичного брачного ритуала, при котором самец
хватает самку хелицерами и потирает её о свою гениталь-
ную область, атипичные безжгутиковые сперматозоиды
упаковываются в шарики и переносятся в женское поло-
вое отверстие либо непосредственно (Eremobates duran-
gonus),
либо с помощью хелицер (Othoes saharae). Яйца
откладываются в специально выкопанные подземные ка-
меры и охраняются самкой. Число стадий нимф велико и
варьирует у разных видов.
Gluvia dorsalis
(Daesiidae), длина
18
мм, встречается
в Испании. —
Galeodes arabs
(Galeodidae), длина
51
мм, с
очень длинными ногами, активен днём, перемещается бы-
стро.
Распространён в Африке. —Mossamedessa
abnormis
(Hexisopodidae), длина 10 мм, ноги короткие, специализи-
рованы для копания, значительную часть времени прово-
дит под землёй. Распространён в Африке. — Eremobates
durangonus (Eremobatidae), длина 20 мм, встречается в
южных штатах США.
3.2.8.
Opiliones,
Сенокосцы
Наряду с широко известными длинноногими
сенокосцами (илл. 671, 673) в состав этого таксона
входят мелкие животные (Sironidae), похожие на кле-
щей (илл. 674), а также плоские коротконогие виды
(Trogulidae) (илл. 675). Всего насчитывается около
4
ООО
видов сенокосцев. Самые крупные из них — это
Trogulus torosus (длина 22 мм) и Mitobates stygnoides
(6 мм в длину, но ноги вытягиваются на 160 мм).
Строение
Наружная организация характеризуется слияни-
ем про- и опистосомы в единое (обычно шаровид-
ное) тело и, у многих видов, сильно удлинёнными
ногами. На спинной стороне просомы может быть
заметно две поперечных борозды. Опистосома ис-
ходно с десятью тергитами, но пять передних могут
сливаться со спинной стороной просомы, образуя
единый скутум (scutum) (илл. 671), а два последних
у многих видов редуцируются.
Илл. 671. Opiliones, Cyphopalpatores.
Вид сбоку. По Roewer (1923).
Коротконогие сенокосцы (Trogulidae) перемещают-
ся медленно, представители же многих длинноногих ви-
дов могут быстро бегать. Их лапки насчитывают до 50 у
Phalangium
и
Opilio
или до
100
члеников у некоторых тро-
пических видов. Сенокосцы, обитающие в траве, могут
лазать по соломинам, спирально обвивая их удлинёнными
лапками. У некоторых видов ноги II пары длиннее осталь-
ных и служат как щупики; такие сенокосцы бегают поэто-
му на трёх парах ног.
Органами чувств служат различные сенсорные
щетинки, щелевидные органы, а также одна пара
хорошо развитых глаз наглазном бугорке (илл. 671).
При опасности многие виды легко аутотомируют себе
ноги, которые ещё на протяжении получаса могут
шевелиться; они отламываются между тазиком и вер-
Илл.
672.
Phalangium opilio
(Opiliones, Cyphopalpatores).
Вид с брюшной стороны. Из Kaestner (1968).
http://jurassic.ru/

500
Arthropoda
Илл.
673.
Oligolophus tridens
(Opiliones).
Самка с яйцекладом откладывает яйца в полость в почве. Длина тела
6 мм. По Silhavy из Kaestner (1968).
лове друг напротив друга, и самец вводит пенис между хе-
лицерами самки
в
женское половое отверстие. Яйца откла-
дываются
в
полости и щели в почве
с
помощью яйцеклада.
У некоторых южноамериканских Gonyleptidae известна
забота о потомстве
—
охрана яиц
и
нимф.
Число нимфаль-
ных стадий варьирует:
у Trogulus их
шесть,
у
Nemastoma
—
семь.
Взрослые особи больше не линяют. Многие виды
живут один год, некоторые дольше
(Т.
nepaeformis до
трёх,
a
Siro rubens
— до девяти лет).
Систематика
Из-за слияния отделов тела сенокосца (особен-
но мелкие Sironidae) напоминают клещей. Выясне-
ние их филогенетических связей с другими паукооб-
разными, однако, затруднено из-за того, что наряду с
типичными архаичными признаками (например,
трёхчленными хелицерами) у Opiliones есть ряд од-
нозначно продвинутых черт (атипичные спермин,
пенис, яйцеклад). Как и у Ricinulei, II пара ходиль-
ных ног во многих случаях служит щупиками. Шульц
(Shultz) (илл. 613) считает Opiliones сестринской
группой по отношению к Scorpiones и Haplocnemata
(Pseudoscorpiones и Solifugae), но эта точка зрения в
настоящей работе не принимается (илл. 612).
3.2.8.1.
Cyphopalpatores
(Виды из этого таксона раньше подразделяли на
Cyphophthalmi и Palpatores; последние, однако, представ-
ляют собой парафилетическую группу).
Sim
rubens
(Sironidae = Cyphophthalmi), длина 1,7 мм,
похожи
на
клещей
(илл.
674),
ноги
короткие.
Спинная часть
просомы и первые восемь тергитов срастаются в единый
твёрдый скутум. Педипальпы щупальцевидные, половое
отверстие неприкрытое. Распространён в Испании и юж-
ной Франции, обычно встречаются под камнями или
в
под-
стилке. Питаются коллемболами и им подобными членис-
тоногими. При опасности ногами запускает в противника
каплю
защитного пачкающего
секрета.
Продолжительность
жизни до 9 лет. Около 25 сходных видов встречаются в
южной Европе и Северной Америке.
тлугом. Для защиты служат также защитные желе-
зы,
протоки которых выводятся в передней части
просомы. Их секрет, содержащий хинон, обладаю-
щий антибиотическим действием, выводится по кап-
лям или распыляется в виде аэрозоля.
Питание сенокосцев разнообразно. Многие виды
специализируются
на
определённой
пище,
другие питаются
мелкими членистоногими, падалью, гнилыми плодами и
(при содержании в неволе) варёным картофелем и хлебом.
Педипальпы многих сенокосцев по строению
сходны с ногами, у Laniatores они образуют ловчий
аппарат, проламывающий покровы жертвы. Рот срав-
нительно широкий, поэтому сенокосцы могут питать-
ся мелкими частицами. Рот находится в преддверии,
образованном мягкими, частично даже членистыми
коксэндитами педипальп и ходильных ног I (а у
Phalangioidea — также и II) пары. Органами выде-
ления служит одна пара коксальных желёз, протоки
которых открываются на тазиках III пары ходильных
ног. Вторая пара желёз закладывается у зародышей
в сегменте, несущем I пару ходильных ног. Кроме
того,
имеются нефроциты и периневральные орга-
ны — узловидные скопления экскреторных клеток
на нервах и коннективах, тянущихся в опистосому.
Органы дыхания — одна пара трахей, малозаметные
стигмы которых расположены вблизи стернита III
сегмента опистосомы или, как у Phalangiidae, поза-
ди последних тазиков. От них крупные разветвлён-
ные трахеи идут в просому. Дополнительные мелкие
стигмы (200^00 мкм в диаметре) на концах голеней
характерны для Phalangiidae. Сердце короткое, с дву-
мя парами остий.
У многих гонилептид (Laniatores) самцы более
склеротизированы и скульптурированы, нежели сам-
ки,
и имеют многочисленные шипы на задних ногах.
Парные гонады U-образно соединены внизу, а дис-
тальные гениталии сильно модифицированы. От зад-
него края полового отверстия в сторону головы про-
должается крышечка, которая не гомологична гени-
тальным крышечкам (operculum) других паукообраз-
ных. Крышечка смещает половое отверстие в про-
межуток между тазиками IV и III пар ходильных ног,
а у Phalangioidea — сильно вперёд, прямо под ниж-
нюю губу. Внутри генитального атриума находится
длинная трубка, подвижная благодаря мускулам и
телескопически выдвигаемая вперёд за счёт повы-
шения давления крови; у самок она служит яйцек-
ладом (илл. 673), у самцов — пенисом. Сперматозо-
иды атипичные безжгутиковые, лишь у Sironoidea
они некоторое время имеют жгутик.
Размножение и развитие
Перенос семени осуществляется путём прямой ко-
пуляции. При спаривании партнёры встают головой к го-
http://jurassic.ru/

Chelicerata
501
Илл.
674.
Sim
duricorius
(Opiliones, Cyphopalpatores).
Длина около
1,7 мм. По
Martens (1978).
*Trogulus nepaeformis
(Trogulidae, «Palpatores»), дли-
на
10 мм
(илл. 675). Тело сильно склеротизированое,
уп-
лощённое. На переднем конце просомы выступ (капюшон),
прикрывающий ротовые
части;
спинной щит просомы сра-
стается
с
пятью тергитами
в
единый скутум. Живут
в по-
чве, активны ночью.
II
пара ног функционирует
в
качестве
щупиков.
При
опасности принимают характерную «позу
трупа». Питаются улитками
с
диаметром раковины
до 8
мм.
Яйца откладывают
в
пустые раковины улиток, кото-
рые затем закупориваются специальным секретом
и
пере-
носятся
в
укрытие.
—
*Paranemastoma quadripunctatum
(Nematostomatidae, «Palpatores»), длина
4,5 мм,
окраска
чёрная. Обитатели почвы,
с
умеренно удлинёнными нога-
ми.
Питаются коллемболами, которые прилипают
к
клей-
ким шаровидным щетинкам
на
педипальпах, похожих
на
ноги.
Яйца
в
студенистых коконах, прикрепляемых на ниж-
ней стороне камней.
—
*Ischyropsalis hellwigi
(Ischyropsa-
lidae, «Palpatores»), длина
7 мм.
Спинная часть просомы
не срастается
с
тергитами
в
скутум, ноги удлинённые,
хе-
лицеры очень крупные. Питаются только улитками, кото-
рых
они
вытягивают
из
раковин
при
помощи хелицер.
У
самцов
на
хелицерах имеются железистые поля, секрет
которых оповещает самку
о
готовности
к
спариванию.
Обитает на почве в умеренно влажных прохладных лесах;
другие виды встречаются
в
южной Европе.
—
*Phalangium
opdio
(Phalangiidae, «Palpatores»), длина самцов
до 7 мм,
самок
—
до
9
мм. Ноги II пары достигают
у
самцов
54
мм
в длину,
у
самок
—
до
38
мм.
У
самцов на хелицерах име-
ется дорсально направленный отросток. Живёт
в
садах,
лесах, на лугах;
на
почве,
но
также
и
на деревьях
и в
кус-
тарнике, проводит часть дневного времени на сухих участ-
ках.
—
*Opdio
parietinus,
длина
7,5 мм.
Живёт, главным
образом, вблизи человеческих построек,
в том
числе
и в
городах. Яйца развиваются лишь весной, после того,
как
они подверглись действию холода. Всего около 1
ООО
ос-
тальных видов, некоторые
из них
распространены все-
светно.
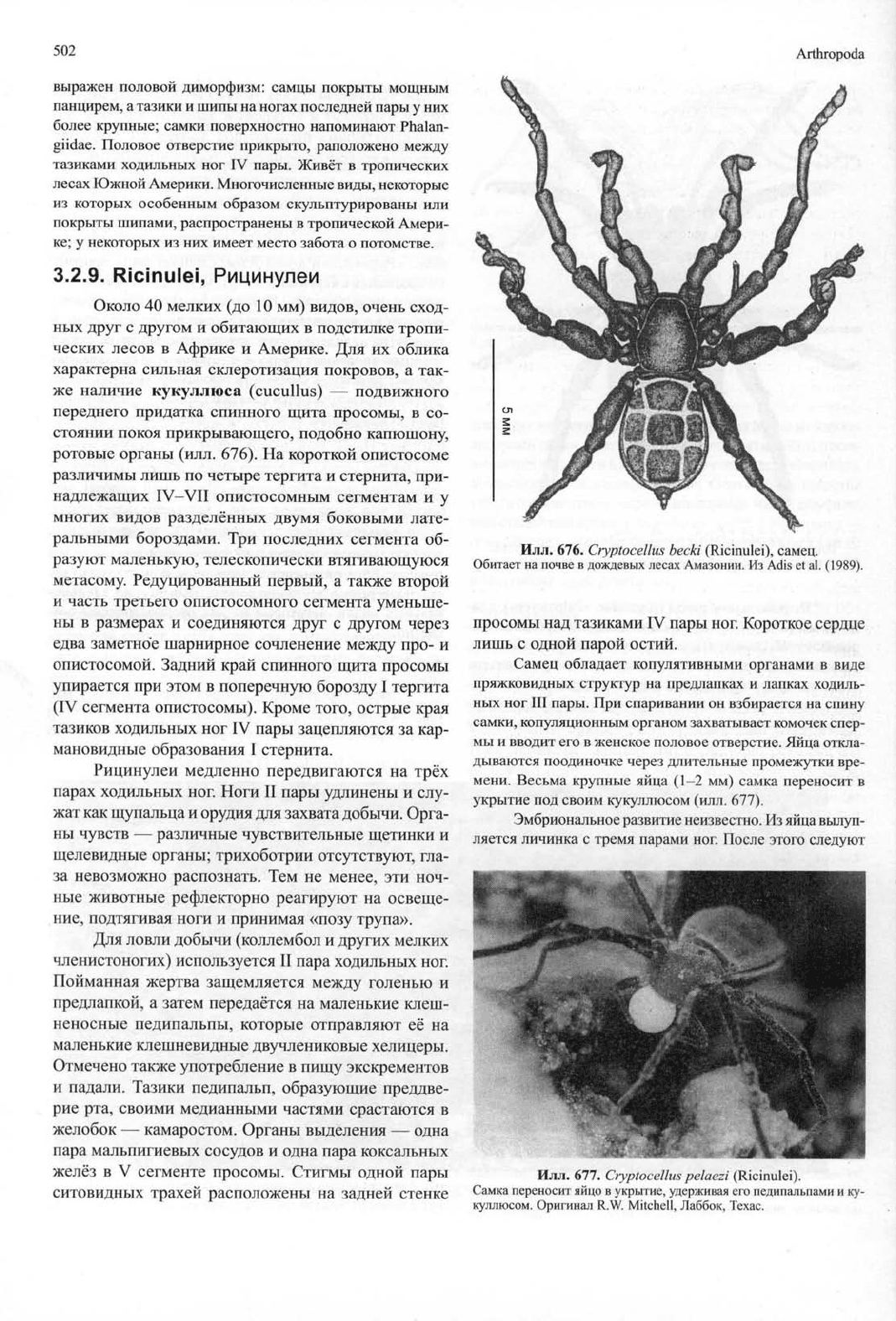
3.2.8.2.
Laniatores
Discocyrtus prospicus
(Gonyleptidae), длина
7 мм.
Педипальпы устроены как хватательные конечности, ноги
II пары преобразованы
в
сильно удлинённые щупики. Ярко
Илл.
675.
Trogulus
sp.
(Opiliones, Cyphopalpatores).
Сенокосец, поедает улитку. Оригинал
В.
Knoflach, Инсбрук.
http://jurassic.ru/
502
Arthropoda
Илл.
676.
Cryptocellus becki (Ricinulei), самец.
Обитает на почве в дождевых лесах Амазонии. Из Adis et al. (1989).
просомы над тазиками IV пары ног. Короткое сердце
лишь с одной парой остий.
Самец обладает копулятивными органами в виде
пряжковидных структур на предлапках и лапках ходиль-
ных ног III пары. При спаривании он взбирается на спину
самки, копуляционным органом захватывает комочек спер-
мы и вводит его
в
женское половое отверстие. Яйца откла-
дываются поодиночке через длительные промежутки вре-
мени. Весьма крупные яйца (1-2 мм) самка переносит в
укрытие под своим кукуллюсом (илл. 677).
Эмбриональное развитие неизвестно.
Из
яйца вылуп-
ляется личинка с тремя парами ног. После этого следуют
Илл. 677. Cryptocellus pelaezi (Ricinulei).
Самка переносит яйцо в укрытие, удерживая его педипальпами и ку-
куллюсом. Оригинал R.W. Mitchell, Лаббок, Техас.
выражен половой диморфизм: самцы покрыты мощным
панцирем, а тазики
и
шипы на ногах последней пары у них
более крупные; самки поверхностно напоминают Phalan-
giidae. Половое отверстие прикрыто, раположено между
тазиками ходильных ног IV пары. Живёт в тропических
лесах Южной
Америки.
Многочисленные виды, некоторые
из которых особенным образом скульптурированы или
покрыты шипами, распространены в тропической Амери-
ке;
у некоторых из них имеет место забота о потомстве.
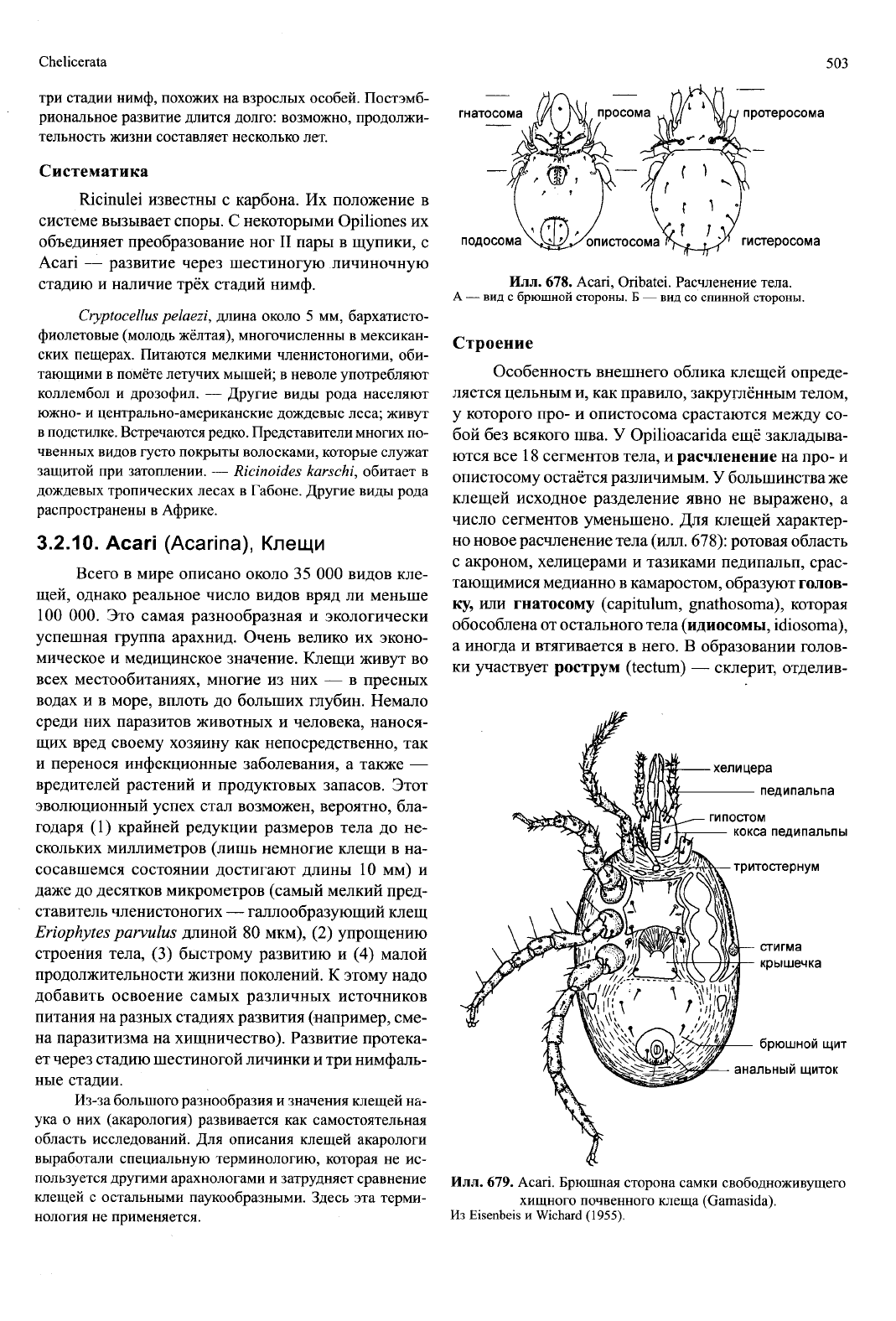
3.2.9.
Ricinulei,
Рицинулеи
Около 40 мелких (до 10 мм) видов, очень сход-
ных друг с другом и обитающих в подстилке тропи-
ческих лесов в Африке и Америке. Для их облика
характерна сильная склеротизация покровов, а так-
же наличие кукуллюса (cucullus) — подвижного
переднего придатка спинного щита просомы, в со-
стоянии покоя прикрывающего, подобно капюшону,
ротовые органы (илл. 676). На короткой опистосоме
различимы лишь по четыре тергита и стернита, при-
надлежащих IV—VII опистосомным сегментам и у
многих видов разделённых двумя боковыми лате-
ральными бороздами. Три последних сегмента об-
разуют маленькую, телескопически втягивающуюся
метасому. Редуцированный первый, а также второй
и часть третьего опистосомного сегмента уменьше-
ны в размерах и соединяются друг с другом через
едва заметное шарнирное сочленение между про- и
опистосомой. Задний край спинного щита просомы
упирается при этом в поперечную борозду I тергита
(IV сегмента опистосомы). Кроме того, острые края
тазиков ходильных ног IV пары зацепляются за кар-
мановидные образования I стернита.
Рицинулеи медленно передвигаются на трёх
парах ходильных ног. Ноги II пары удлинены и слу-
жат как щупальца и орудия для захвата добычи. Орга-
ны чувств — различные чувствительные щетинки и
щелевидные органы; трихоботрии отсутствуют, гла-
за невозможно распознать. Тем не менее, эти ноч-
ные животные рефлекторно реагируют на освеще-
ние,
подтягивая ноги и принимая «позу трупа».
Для ловли добычи (коллембол и других мелких
членистоногих) используется II пара ходильных ног.
Пойманная жертва защемляется между голенью и
предлапкой, а затем передаётся на маленькие клеш-
неносные педипальпы, которые отправляют её на
маленькие клешневидные двучлениковые хелицеры.
Отмечено также употребление в пищу экскрементов
и падали. Тазики педипальп, образующие преддве-
рие рта, своими медианными частями срастаются в
желобок — камаростом. Органы выделения — одна
пара мальпигиевых сосудов и одна пара коксальных
желёз в V сегменте просомы. Стигмы одной пары
ситовидных трахей расположены на задней стенке
http://jurassic.ru/
Chelicerata
503
три стадии нимф, похожих на взрослых особей. Постэмб-
риональное развитие длится долго: возможно, продолжи-
тельность жизни составляет несколько лет.
Систематика
Ricinulei известны с карбона. Их положение в
системе вызывает споры. С некоторыми Opiliones их
объединяет преобразование ног II пары в щупики, с
Acari — развитие через шестиногую личиночную
стадию и наличие трёх стадий нимф.
Cryptocellus
pelaezi,
длина около 5 мм, бархатисто-
фиолетовые (молодь жёлтая), многочисленны в мексикан-
ских пещерах. Питаются мелкими членистоногими, оби-
тающими в помёте летучих мышей; в неволе употребляют
коллембол и дрозофил. — Другие виды рода населяют
южно- и центрально-американские дождевые леса; живут
в
подстилке.
Встречаются
редко.
Представители многих
по-
чвенных видов густо покрыты волосками, которые служат
защитой при затоплении. — Ricinoides karschi, обитает в
дождевых тропических лесах в Габоне. Другие виды рода
распространены в Африке.
3.2.10.
Acari (Acarina),
Клещи
Всего в мире описано около 35
ООО
видов кле-
щей, однако реальное число видов вряд ли меньше
100 ООО. Это самая разнообразная и экологически
успешная группа арахнид. Очень велико их эконо-
мическое и медицинское значение. Клещи живут во
всех местообитаниях, многие из них — в пресных
водах и в море, вплоть до больших глубин. Немало
среди них паразитов животных и человека, нанося-
щих вред своему хозяину как непосредственно, так
и перенося инфекционные заболевания, а также —
вредителей растений и продуктовых запасов. Этот
эволюционный успех стал возможен, вероятно, бла-
годаря (1) крайней редукции размеров тела до не-
скольких миллиметров (лишь немногие клещи в на-
сосавшемся состоянии достигают длины 10 мм) и
даже до десятков микрометров (самый мелкий пред-
ставитель членистоногих — галлообразующий клещ
Eriophytes parvulus длиной 80 мкм), (2) упрощению
строения тела, (3) быстрому развитию и (4) малой
продолжительности жизни поколений. К этому надо
добавить освоение самых различных источников
питания на разных стадиях развития (например, сме-
на паразитизма на хищничество). Развитие протека-
ет через стадию шестиногой личинки и три нимфаль-
ные стадии.
Из-за большого разнообразия и значения клещей на-
ука о них (акарология) развивается как самостоятельная
область исследований. Для описания клещей акарологи
выработали специальную терминологию, которая не ис-
пользуется другими арахнологами и затрудняет сравнение
клещей с остальными паукообразными. Здесь эта терми-
нология не применяется.
Илл. 678. Acari, Oribatei. Расчленение тела.
А — вид с брюшной стороны. Б — вид со спинной стороны.
Строение
Особенность внешнего облика клещей опреде-
ляется цельным и, как правило, закруглённым телом,
у которого про- и опистосома срастаются между со-
бой без всякого шва. У Opilioacarida ещё закладыва-
ются все 18 сегментов тела, и расчленение на про- и
опистосому остаётся различимым. У большинства же
клещей исходное разделение явно не выражено, а
число сегментов уменьшено. Для клещей характер-
но новое расчленение тела
(илл.
678): ротовая область
с акроном, хелицерами и тазиками педипальп, срас-
тающимися медианно в камаростом, образуют голов-
ку, или гнатосому (capitulum, gnathosoma), которая
обособлена от остального тела (идиосомы, idiosoma),
а иногда и втягивается в него. В образовании голов-
ки участвует рострум (tectum) — склерит, отделив-
-хелицера
педипальпа
ги
постом
—
кокса педипальпы
Илл. 679. Acari. Брюшная сторона самки свободноживущего
хищного почвенного клеща (Gamasida).
Из Eisenbeis и Wichard (1955).
http://jurassic.ru/
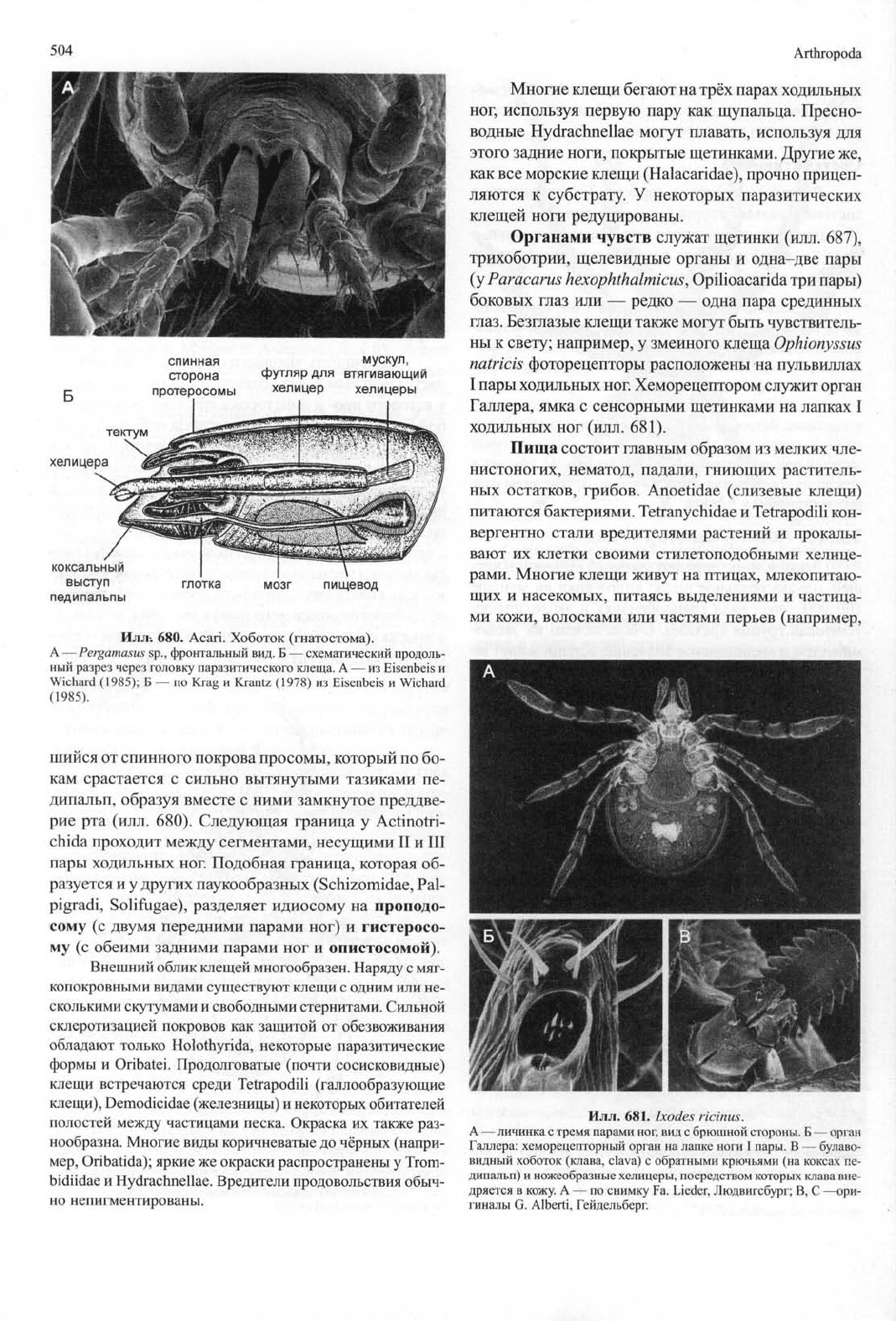
504
выступ
глотка мозг пищевод
педипальпы
Илл-.
680. Acari. Хоботок (гнатостома).
А — Pergamasus sp., фронтальный вид. Б — схематический продоль-
ный разрез через головку паразитического клеща. А — из Eisenbeis и
Wichard (1985); Б — по K_rag и Krantz (1978) из Eisenbeis и Wichard
(1985).
шийся от спинного покрова просомы, который по бо-
кам срастается с сильно вытянутыми тазиками пе-
дипальп, образуя вместе с ними замкнутое преддве-
рие рта (илл. 680). Следующая граница у Actinotri-
chida проходит между сегментами, несущими II и III
пары ходильных ног. Подобная граница, которая об-
разуется и у других паукообразных (Schizomidae, Pal-
pigradi, Solifugae), разделяет идиосому на проподо-
сому (с двумя передними парами ног) и гистеросо-
му (с обеими задними парами ног и опистосомой).
Внешний облик клещей многообразен. Наряду с мяг-
копокровными видами существуют клещи с одним или не-
сколькими скутумами и свободными стернитами. Сильной
склеротизацией покровов как защитой от обезвоживания
обладают только Holothyrida, некоторые паразитические
формы и Oribatei. Продолговатые (почти сосисковидные)
клещи встречаются среди Tetrapodili (галлообразующие
клещи), Demodicidae (железницы) и некоторых обитателей
полостей между частицами песка. Окраска их также раз-
нообразна. Многие виды коричневатые до чёрных (напри-
мер,
Oribatida); яркие же окраски распространены у Trom-
bidiidae и Hydrachnellae. Вредители продовольствия обыч-
но непигментированы.
Arthropoda
А
Илл.
681. Ixodes ricinus.
А — личинка с тремя парами ног, вид с брюшной стороны. Б — орган
Галлера: хеморецепторный орган на лапке ноги I пары. В — булаво-
видный хоботок (клава, clava) с обратными крючьями (на коксах пе-
дипальп) и ножеобразные хелицеры, посредством которых клава вне-
дряется в кожу. А — по снимку Fa. Lieder, Людвигсбург; В, С —ори-
гиналы G. Alberti, Гейдельберг.
Многие клещи бегают на трёх парах ходильных
ног, используя первую пару как щупальца. Пресно-
водные Hydrachnellae могут плавать, используя для
этого задние ноги, покрытые щетинками. Другие же,
как все морские клещи (Halacaridae), прочно прицеп-
ляются к субстрату. У некоторых паразитических
клещей ноги редуцированы.
Органами чувств служат щетинки (илл. 687),
трихоботрии, щелевидные органы и одна-две пары
(у Paracarus hexophthalmicus, Opilioacarida три пары)
боковых глаз или — редко — одна пара срединных
глаз.
Безглазые клещи также могут быть чувствитель-
ны к свету; например, у змеиного клеща Ophionyssus
natricis фоторецепторы расположены на пульвиллах
I пары ходильных
ног.
Хеморецептором служит орган
Галлера, ямка с сенсорными щетинками на лапках I
ходильных ног (илл. 681).
Пища состоит главным образом из мелких чле-
нистоногих, нематод, падали, гниющих раститель-
ных остатков, грибов. Anoetidae (слизевые клещи)
питаются бактериями. Tetranychidae и Tetrapodili кон-
вергентно стали вредителями растений и прокалы-
вают их клетки своими стилетоподобными хелице-
рами. Многие клещи живут на птицах, млекопитаю-
щих и насекомых, питаясь выделениями и частица-
ми кожи, волосками или частями перьев (например,
http://jurassic.ru/

Chelicerata
505
Илл.
682.
Caparinia tripdis (Acaridida, Acaridei), вызывает
чесотку у ежей.
Спаривание. Оригинал H. Mehlhorn, Бохум.
переносятся в самку газом, освобождаемым под действи-
ем энзимов. Самцы многих Actinotrichida образуют стебель-
ковые сперматофоры: некоторые — поодиночке, другие —
попарно. Самец водяного клеща Arrenurus приклеивает
самку к своему спинному отростку, выделяет сперматофор
и переносит его
в
женское половое отверстие. Копуляция с
помощью пенисообразных структур возникала многократ-
но,
причём при ней партнёры занимают различные поло-
жения в зависимости от того, где находится у них половое
отверстие (илл. 682). При вентральном его расположении
спаривание осуществляется «животом
к
животу», при зад-
нем же самец взбирается на самку или же партнёры раз-
вёрнуты головами в противоположные стороны. У Poda-
polipidae и Cheyletidae самец подползает сзади под самку,
пока его половое отверстие, находящееся на дорсальной
стороне, щ достигает полового отверстия самки. У неко-
торых клещей самец носит несозревшую самку до копуля-
ции: у Tarsonemini — на стадии личинки, у Sarcoptifor-
mes —дейтонимфы. Предшествующий этап эволюции этой
формы поведения демонстрируют Tetranychidae, у которых
самец созревает раньше самок и находится возле женских
дейтонимф, ожидая их линьки. Самцы Pyemotidae, нако-
нец, ожидают рождения самок (обычно их сестёр), с кото-
рыми они тут
же
спариваются, на физогастрической самке
(обычно на собственной матери, паразитируя на ней).
Распространён партеногенез. У многих видов имеет
место арренотокный партеногенез (самцы выходят из нео-
плодотворённых гаплоидных яиц); вероятно, это вообще
самый обычный способ определения пола. Телитокный
партеногенез (неоплодотворённые яйца после различных
регуляторных воздействий становятся диплоидными
и
дают
самок) распространён как географический вариант. У не-
которых Gamasida и Oribatida самцы, по-видимому, отсут-
ствуют.
Яйца откладываются поодиночке или группа-
ми.
Исходно из них выходит личинка лишь с тремя
парами ног (илл. 681), но зачатки IV пары заметны
уже у зародыша. У Holothyrida, Oribatei и некоторых
Acaridia за личинкой следуют три стадии прото-, дей-
то-
и тритонимфы. У многих Anactinotrichida отсут-
ствуют тритонимфы, у Trombidiidae и Hydrachnelli-
угревая железница Demodex folliculorum) (илл. 688).
Паразитизм у клещей возникает многократно и кон-
вергентно; у многих из них паразитический образ
жизни характерен только для одной или нескольких
стадий. Клешневидные (исходно трёхчлениковые)
хелицеры могут вытягиваться в результате повыше-
ния кровяного давления. У хищных видов они круп-
ные,
у многих паразитов — тонкие стилетовидные.
Педипальпы обычно щупикообразные; у некоторых
видов они служат как очень маленький, а у других —
как крупный ловчий аппарат. Их медианно сросшие-
ся тазики образуют на хоботке (гнатосоме) преддве-
рие рта, в которое открываются протоки слюнных
или паутинных (Tetranychidae) желёз. Рот через со-
сательную глотку (илл. 680) ведёт в среднюю кишку
с одной-семью парами слепых выростов. У Actine-
dida средняя кишка оканчивается слепо, а задняя
кишка функционирует как орган выделения. Дру-
гими экскреторными органами служат одна пара кок-
сальных желёз на сегменте I ходильных ног (Holo-
thyridae, Ixodidae), одна-две пары мальпигиевых со-
судов, а также клетки средней кишки.
Исходно (у Anactinotrichida) органами дыха-
ния служат четыре пары трахей с боковыми стигма-
ми (на IX-XII сегментах у Opilioacarida), у большин-
ства же клещей образуется лишь одна их пара. У
Actinotrichida трахеи отсутствуют (многие Sarcopti-
formes) или возникают вторично, начинаясь в облас-
ти хелицер, полового отверстия или тазиков ходиль-
ных ног (аподемы у Oribatei). Большинство назем-
ных клещей нуждаются в высокой влажности возду-
ха; некоторые могут извлекать воду из воздуха. Ма-
ленькое сердце с двумя парами остий ещё сохраня-
ется у многих Anactinotrichida.
Гонады, исходно представляющие собой пар-
ные трубки, у многих групп U-образно срастаются
внизу, в пределе становясь непарной гонадой. Виды,
для которых характерно образование сперматофоров,
имеют сложные гениталии; у других имеются струк-
туры, подобные пенису. Исходно половое отверстие
расположено вентрально в середине тела; оно может
перемещаться вперёд или назад, переходить через
задний конец на дорсальную сторону и даже распо-
лагаться непосредственно позади гнатосомы (у сам-
цов Podapolipidae). Сперматозоиды безжгутиковые.
Размножение и развитие
Перенос семени у Anactinotrichida осуществля-
ется через бесстебельчатые сперматофоры, которые
при помощи хелицер (у Gamasida — соответствую-
щим образом модифицированных) вводятся в жен-
ское половое отверстие.
Особенно сложные сперматофоры имеют иксодовые
клещи; сперматозоиды и симбиотические бактерии в них
http://jurassic.ru/