Вестхайде В.,Ригер Р.( ред.) Зоология беспозвоночных в двух томах. Том 1: от простейших до моллюсков и артропод
Подождите немного. Документ загружается.

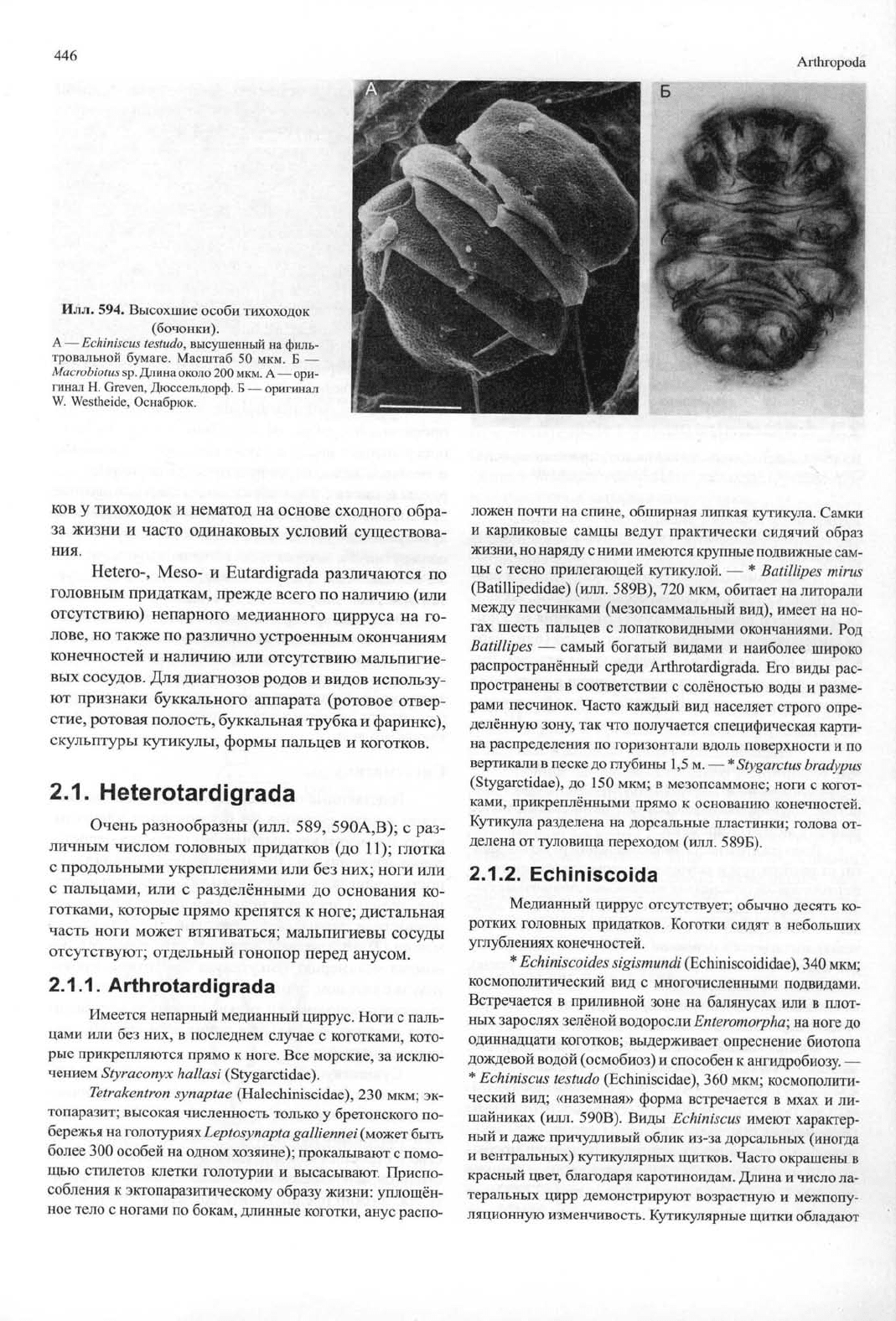
446
Arthropoda
Илл. 594. Высохшие особи тихоходок
(бочонки).
А — Echiniscus testudo, высушенный на филь-
тровальной бумаге. Масштаб 50 мкм. Б —
Macrobiotus sp. Длина около 200 мкм. А — ори-
гинал Н. Greven, Дюссельдорф. Б — оригинал
W. Westheide, Оснабрюк.
ков у тихоходок и нематод на основе сходного обра-
за жизни и часто одинаковых условий существова-
ния.
Hetero-, Meso- и Eutardigrada различаются по
головным придаткам, прежде всего по наличию (или
отсутствию) непарного медианного цирруса на го-
лове, но также по различно устроенным окончаниям
конечностей и наличию или отсутствию мальпигие-
вых сосудов. Для диагнозов родов и видов использу-
ют признаки буккального аппарата (ротовое отвер-
стие,
ротовая полость, буккальная трубка и фаринкс),
скульптуры кутикулы, формы пальцев и коготков.
2.1.
Heterotardigrada
Очень разнообразны (илл. 589, 590А,В); с раз-
личным числом головных придатков (до 11); глотка
с продольными укреплениями или без них; ноги или
с пальцами, или с разделёнными до основания ко-
готками, которые прямо крепятся к ноге; дистальная
часть ноги может втягиваться; мальпигиевы сосуды
отсутствуют; отдельный гонопор перед анусом.
2.1.1. Arthrotardigrada
Имеется непарный медианный циррус. Ноги с паль-
цами или без них, в последнем случае с коготками, кото-
рые прикрепляются прямо к ноге. Все морские, за исклю-
чением
Styraconyx
hallasi (Stygarctidae).
Tetrakentron
synaptae (Halechiniscidae), 230 мкм; эк-
топаразит; высокая численность только у бретонского по-
бережья на голотуриях
Leptosynapta galliennei
(может быть
более 300 особей на одном хозяине); прокалывают с помо-
щью стилетов клетки голотурии и высасывают. Приспо-
собления к эктопаразитическому образу жизни: уплощён-
ное тело с ногами по бокам, длинные коготки, анус распо-
ложен почти на спине, обширная липкая кутикула. Самки
и карликовые самцы ведут практически сидячий образ
жизни, но наряду
с
ними имеются крупные подвижные сам-
цы с тесно прилегающей кутикулой. — *
Batillipes
minis
(Batillipedidae) (илл. 589В), 720 мкм, обитает на литорали
между песчинками (мезопсаммальный вид), имеет на но-
гах шесть пальцев с лопатковидными окончаниями. Род
Batillipes — самый богатый видами и наиболее широко
распространённый среди Arthrotardigrada. Его виды рас-
пространены в соответствии с солёностью воды и разме-
рами песчинок. Часто каждый вид населяет строго опре-
делённую зону, так что получается специфическая карти-
на распределения по горизонтали вдоль поверхности и по
вертикали
в
песке
до
глубины 1,5
м.
—
*
Stygarctus bradypus
(Stygarctidae), до 150 мкм; в мезопсаммоне; ноги с когот-
ками, прикреплёнными прямо к основанию конечностей.
Кутикула разделена на дорсальные пластинки; голова от-
делена от туловища переходом (илл. 589Б).
2.1.2.
Echiniscoida
Медианный циррус отсутствует; обычно десять ко-
ротких головных придатков. Коготки сидят в небольших
углублениях конечностей.
*
Echiniscoides sigismundi
(Echiniscoididae), 340 мкм;
космополитический вид с многочисленными подвидами.
Встречается в приливной зоне на балянусах или в плот-
ных зарослях зелёной водоросли
Enteromorpha;
на ноге до
одиннадцати коготков; выдерживает опреснение биотопа
дождевой водой (осмобиоз) и способен к ангидробиозу. —
*
Echiniscus testudo
(Echiniscidae), 360 мкм; космополити-
ческий вид; «наземная» форма встречается в мхах и ли-
шайниках (илл. 590В). Виды Echiniscus имеют характер-
ный и даже причудливый облик из-за дорсальных (иногда
и вентральных) кутикулярных щитков. Часто окрашены в
красный цвет, благодаря каротиноидам. Длина и число ла-
теральных цирр демонстрируют возрастную и межпопу-
ляционную изменчивость. Кутикулярные щитки обладают
http://jurassic.ru/

Tardigrada
447
сложившейся системой полостей (возможно, эти ёмкости
жидкости служат для замедления испарения).
2.2. Mesotardigrada
Вокруг ротового отверстия расположены боко-
вые цирры и папиллы; глотка с плакоидами; коготки
одинаковые, мальпигиевы сосуды имеются.
Thermozodium
esakii (Thermozodiidae), 490 мкм; об-
наружен
в
Японии,
в
водорослевых матах по краям горячих
источников (около 40°С). Со времени открытия в 1930 г.
не было ни одной повторной находки: типовое местона-
хождение было разрушено землетрясением. По своей мор-
фологии занимает промежуточное положение между Hete-
rotardigrada и Eutardigrada.
2.3.
Eutardigrada
Довольно однородная группа; преимуществен-
но,
без головных придатков; мальпигиевы сосуды
имеются; анальное и половое отверстия соединены
в клоаку; на каждой ноге два двойных коготка, каж-
дый из которых состоит из главной и дополнитель-
ной ветвей; глотка обычно с плакоидами.
2.3.1.
Parachela
Головных придатков нет; главная и дополнительная
ветви коготка соединены.
Macrobiotus
hufelandi (Macrobiotidae), длина до 1,2
мм;
космополитический эврибионтный
вид;
назван
в
честь
врача Хуфеланда, опубликовавшего классическую работу
по долгожительству (макробиозу) «Искусство продления
человеческой жизни» (1797). У самцов наружная сторона
ног четвёртой пары с уплощённым краем. Как и большин-
ство видов Macrobiotus, откладывает одиночные яйца со
скульптурной оболочкой (илл. 593). — * Pseudobiotus
megalonyx
(Hypsibiidae), 900 мкм; широко распространён-
ный вид, населяет только пресные водоёмы, весной и осе-
нью достигает высокой численности; самка откладывает
до
60
яиц
в
старую кутикулу, которую после линьки долгое
время носит с собой (забота о потомстве?). —
*
Ramazzot-
tius oberhaeuseri
(Hypsibiidae), 500
мкм;
очень часто встре-
чается во мхе на крышах; отличается девятью коричневы-
ми поперечными поясами; у самцов внешняя сторона ног
четвёртой пары
с
уплощенным краем (илл. 590Б). —
*
На-
lobiotus crispae (Hypsibiidae), 665 мкм; морской (!) вид,
встречается на бурых водорослях в приливной зоне о-ва
Диско (Гренландия); хорошо выраженный цикломорфоз
зависит от времени года. Эти вторично перешедшие
к
жиз-
ни в море Eutardigrada отличаются предельно увеличен-
ными мальпигиевыми сосудами.
Ископаемые представители:
Beorn leggi
(Beornidae),
300 мкм; единственный вид, хорошо сохранившийся в те-
чение 60 млн. лет в канадском янтаре; очень похож на ре-
центные виды Hypsibius.
2.3.2.
Apochela
Вокруг ротового отверстия расположены иннерви-
руемые папиллы; глотка без плакоидов; главная и допол-
нительная ветви коготка разделены.
*
Milnesium tardigradum
(Milnesiidae),
1
мм, эвриби-
онтный космополитический вид; его пища, преимуще-
ственно, животного происхождения.
http://jurassic.ru/

Euarthropoda,
Настоящие членистоногие
За исключением малой группы Onychophora и,
вероятно, Tardigrada (чьё систематическое положе-
ние трудно установить из-за их мелких размеров, все
другие членистоногие относятся к Euarthropoda, т.е.
настоящим членистоногим. Такой уровень организа-
ции («вторая ступень артроподизации») был достиг-
нут ими, вероятно, уже в кембрии. К этому выводу
можно прийти, рассмотрев общие признаки и уро-
вень организации и формы, признанные самыми при-
митивными. Поэтому для Euarthropoda характер-
ны прежде всего:
1.
скелет, состоящий из соединённых друг с
другом склеритов (пластинок, щитков) и связанная
с этим редукция кожно-мускулыюго мешка,
2.
цефалон (голова) из акрона и, вероятно, пяти
первичных сегментов, с которыми позднее сливает-
ся шестой; первый из головных сегментов, так назы-
ваемый преантеннальный сегмент лишён парных
придатков (возможно, остатки его конечностей сли-
лись с образованием лабрума — верхней губы); за
ним следует одна пара предротовых членистых ан-
тенн и три (четыре) пары других конечностей (илл.
568,
607),
3.
членистые конечности в виде расщеплён-
ных ног (двуветвистых) (илл. 596),
4.
одна пара боковых фасеточных глаз (илл.
599,
602) и исходно четыре медианных глаза,
5.
нефридии исходно, видимо, в четырёх го-
ловных сегментах и двух следующих за ними туло-
вищных сегментах.
Кутикула состоит из участков, часть из которых
сильно склеротизирована, часть — слабее или вооб-
ще не склеротизирована. В результате возникает ске-
лет, состоящий из склеритов или щитков. Начиная
с мандибулят, каждый сегмент имеет одну сильно
склеротизированную дорсальную пластинку (тер-
гит),
одну вентральную (стернит) и слабо склеро-
тизированные мягкие плевры по бокам, в которых
могут быть твёрдые плевриты (отчётливо выражен-
ные,
например, у многих насекомых). Между сегмен-
тами расположены мягкие кутикулярные сочленов-
ные мембраны (межсегментные покровы), обеспечи-
вающие подвижность или вытягивание туловища при
заглатывании пищи и при образовании большого
количества половых клеток. Эти сочленовные мем-
браны не маркируют, как правило, границы первич-
ных сегментов, а расположены впереди или позади
этих границ. В тех местах, которые должны быть
эластичными, например, в сочленениях крыльев или
Hannes Paulus, Вена
дорсальная
продольная дорсовентральная
вентральное поперечное сухожилие
Илл.
595. Распад кожно-мускульного мешка у Euarthropoda
в отдельные пучки мышц.
Схематическое изображение сегмента с конечностями. По публика-
циям различных авторов.
суставах ног для прыгания (у блох), откладывается
резилин — протеин, напоминающий резину.
Одним из следствий образования щиткового
скелета была редукция кожно-мускульного меш-
ка до отдельных пучков поперечнополосатых мышц
(илл. 595). Первоначально тело состояло из равно-
ценных (гомономных) сегментов, следующих один
за другим. Более или менее отчётливо такое состоя-
ние наблюдается у многих вымерших трилобитов, у
Progoneata, Chilopoda и у некоторых ракообразных
(Remipedia и Cephalocarida). Однако у большинства
Euarthropoda сегментация тела гетерономная. Сег-
менты объединены в несколько функциональных
единиц —тагм. На переднем конце акрона и несколь-
ко (пять?) следующих за ним сегментов всегда сли-
ваются с образованием головы (цефалон). Вероятно,
голова является гомологичным у всех Euarthropoda
отделом, однако в сформированном состоянии в ней
уже нельзя найти следов первичной сегментации.
Туловище также делится на тагмы, которые возник-
ли у различных групп независимо и объединяют в
своем составе различное число сегментов.
У хелицеровых всего две тагмы: (1) просома, обра-
зованная из головы и двух туловищных сегментов, и (2)
опистосома. У большинства ракообразных и насекомых
туловище состоит из торакса и абдомена. Тагмы могут объе-
http://jurassic.ru/
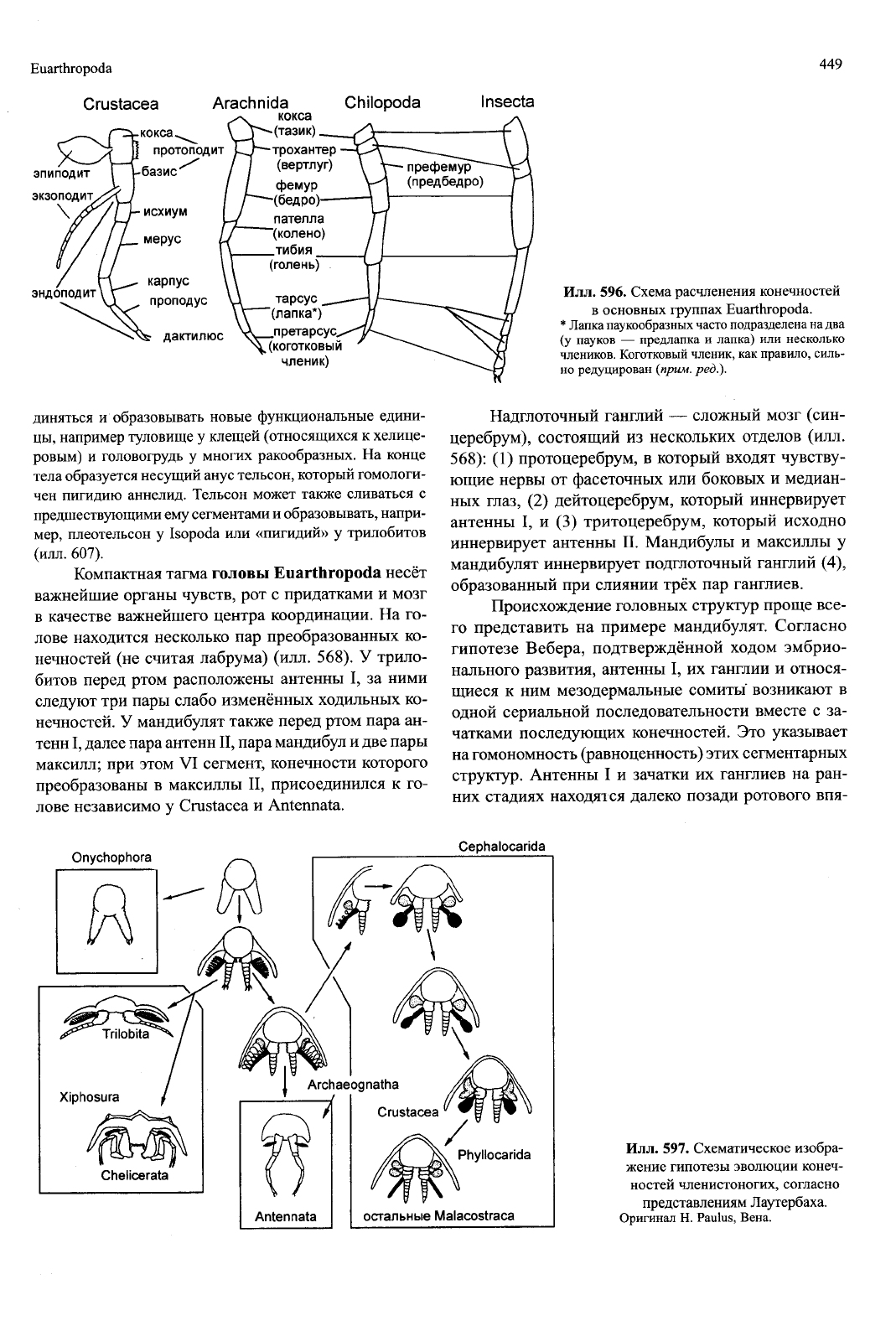
Euarthropoda
449
Crustacea
Arachnida
кокса
Chilopoda
Insecta
ЭПИПОДИТ
ЭКЗОПОДИТ
\
'
ЭНДОПОДИТ
кокса
^
протоподит
базис^
исхиум
мерус
карпус
проподус
дактилюс
-(тазик)
-трохантер
(вертлуг)
фемур
-(бедро)
пателла
"(колено)
_тибия
(голень)
тарсус
"(лапка*)
_претарсус.
ч
(коготковый
членик)
диняться и образовывать новые функциональные едини-
цы,
например туловище у клещей (относящихся к хелице-
ровым) и головогрудь у многих ракообразных. На конце
тела образуется несущий анус тельсон, который гомологи-
чен пигидию аннелид. Тельсон может также сливаться с
предшествующими ему сегментами
и
образовывать, напри-
мер,
плеотельсон у Isopoda или «пигидий» у трилобитов
(илл. 607).
Компактная тагма головы Euarthropoda несёт
важнейшие органы чувств, рот с придатками и мозг
в качестве важнейшего центра координации. На го-
лове находится несколько пар преобразованных ко-
нечностей (не считая лабрума) (илл. 568). У трило-
битов перед ртом расположены антенны I, за ними
следуют три пары слабо изменённых ходильных ко-
нечностей. У мандибулят также перед ртом пара ан-
тенн
I,
далее пара антенн II, пара мандибул и две пары
максилл; при этом VI сегмент, конечности которого
преобразованы в максиллы II, присоединился к го-
лове независимо у Crustacea и Antennata.
Onychophora
Илл.
596. Схема расчленения конечностей
в
основных группах Euarthropoda.
*
Лапка
паукообразных
часто подразделена на два
(у
пауков — предлапка и лапка) или несколько
члеников.
Коготковый членик, как правило, силь-
но
редуцирован
(прим.
ред.).
Надглоточный ганглий — сложный мозг (син-
церебрум), состоящий из нескольких отделов (илл.
568):
(1) протоцеребрум, в который входят чувству-
ющие нервы от фасеточных или боковых и медиан-
ных глаз, (2) дейтоцеребрум, который иннервирует
антенны I, и (3) тритоцеребрум, который исходно
иннервирует антенны II. Мандибулы и максиллы у
мандибулят иннервирует подглоточный ганглий (4),
образованный при слиянии трёх пар ганглиев.
Происхождение головных структур проще все-
го представить на примере мандибулят. Согласно
гипотезе Вебера, подтверждённой ходом эмбрио-
нального развития, антенны I, их ганглии и относя-
щиеся к ним мезодермальные сомиты' возникают в
одной сериальной последовательности вместе с за-
чатками последующих конечностей. Это указывает
на гомономность (равноценность) этих сегментарных
структур. Антенны I и зачатки их ганглиев на ран-
них стадиях находятся далеко позади ротового впя-
Cephalocarida
Archaeognatha
Crustacea
остальные
Malacostraca
Илл.
597. Схематическое изобра-
жение
гипотезы эволюции конеч-
ностей
членистоногих, согласно
представлениям
Лаутербаха.
Оригинал
Н. Paulus, Вена.
http://jurassic.ru/

450
Arthropoda
чивания. В процессе дальнейшего развития рот и
верхняя губа смещаются в направлении, противопо-
ложном движению зачатков антенн I и их пар ганг-
лиев,
в результате рот оказывается перемещённым
вниз и назад. Комиссура дейтоцеребрума возникает
только тогда, когда зачатки ганглиев антенн I отодви-
гаются ото рта. Отсюда следует, что антенны I по
происхождению являются настоящими сегментарны-
ми конечностями.
На ещё более ранней стадии, перед зачатками
антенн I появляется другая пара сомитов, которая
(прежде всего, у ракообразных) тоже может иметь
целомические полости. Эта мезодерма обеспечива-
ет мускулатурой верхнюю губу и стомодеум и уча-
ствует в образовании передней аорты в области го-
ловы. Её считают признаком наличия ещё одного, так
называемого преантеннального сегмента. У него
ныне нет конечностей; однако, возможно, из них про-
изошла верхняя губа (лабрум), которая многократно
образуется из парных или двухлопастных закладок.
Если это предположение правильное, протоцеребрум
также образуется из двух частей: из отдела мозга ак-
рона, архицеребрума и из пары ганглиев преантен-
нального сегмента, прозоцеребрума. Вместе с тем, у
рецентных Euarthropoda чёткое разграничение двух
этих отделов невозможно. Они образуют единый
функциональный блок; но вполне возможно, что бо-
ковые лопасти со своими комиссурами являются ос-
татками прозоцеребрума.
Согласно этой точке зрения, которая также по-
ложена в основу описания сложного мозга, голова
мандибулят возникла из акрона и, вероятно, шести
настоящих, расположенных исходно позади рта сег-
ментов, которые сливались вместе, обычно не остав-
ляя следов швов (илл. 568, таблица 4).
Вероятно, тело предкового вида Euarthropoda
состояло из многочисленных, одинаковых (гомоном-
ных) сегментов с такими же одинаковыми конечнос-
тями. Исходно они были мультифункциональными
и служили для передвижения, добывания пищи и
дыхания. Но возможно, что пара конечностей второ-
го сегмента (антенны I) уже была специализирована
в качестве органа
осязания:
такие преоральные (пред-
ротовые) антенны были у всех трилобитов (илл. 607).
У большинства рецентных Euarthropoda туловище
разными способами разделено на функциональные
отделы (тагмы). Поэтому конечности также преоб-
разованы по-разному и служат для хождения, плава-
ния, прыгания, лазания, хватания, ловли или копа-
ния (илл. 846). Остальные конечности несут органы
чувств, служат для жевания, хватания и чистки. Для
размножения служат гоноподии, конечности для вы-
нашивания яиц и т.п.
Многообразие структур и конструкций конеч-
ностей Euarthropoda является следствием склеритно-
го построения кутикулы. Это членистые конечнос-
ти (артроподии), части которых образованы из проч-
ных трубок (экзокутикулы). На концах членики снаб-
жены суставами для прикрепления к туловищу и свя-
заны между собой мягкими, не склеротизированны-
ми участками кутикулы. Членики приводятся в дви-
жение мышцами, сгибателями и разгибателями, ко-
торые начинаются в туловище или в проксимальных
частях члеников конечностей и сухожилия которых
прикрепляются прямо за соответствующим суставом.
Число члеников конечностей может быть увеличено
вторично путём образования ложных суставов. У таких
суставов нет собственных мышц; сухожилия проходят
сквозь них (без прикрепления) и заканчиваются у ближай-
шего настоящего сустава. Лапки насекомых и некоторых
паукообразных,
а
также многие экзоподиты ракообразных,
часто подразделены вторично таким образом. У некоторых
хелицеровых и губоногих
в
определённых суставах ног нет
разгибателей; распрямление таких конечностей достига-
ется путём повышения давления гемолимфы. Последний
членик эндоподита первоначально заканчивался в виде
простого коготка (дактилюс или дактилоподит). У насеко-
мых, пауков и симфил он сильно редуцирован и снабжён
двумя коготками (у кивсяков — одним), а часто ещё и при-
крепительными лопастями (аролий, пульвиллы) (илл. 844).
Традиционно членики ног Euarthropoda назы-
вают по-разному в разных группах (илл. 596).
Гомология этих члеников не всегда доказана:
неизвестно, например, какой членик отсутствует у
ракообразных и насекомых в сравнении с паукооб-
разными или «Myriapoda» (возможно, пателла). По-
этому основной план строения членистой ноги Euar-
thropoda не общепризнан, а его возникновение в фи-
логенезе — предмет различных гипотез.
Исходный пункт рассуждений — прежде все-
го,
решение вопроса о том, были ли у последнего
общего предка аннелид и членистоногих конечнос-
ти.
(1) Если предположить, что у предкового вида
конечностей не было, то ноги членистоногих и пара-
подий аннелид следует считать конвергентными об-
разованиями (возникшими независимо). (2)
Если всё же у предкового вида уже были конечнос-
ти,
напоминающие параподий, тогда можно считать
примитивные параподий аннелид и первичные чле-
нистые ноги гомологичными образованиями.
Мы считаем вероятным, что предковый вид
членистых уже обладал конечностями, напоминаю-
щими параподий. Далее, у полихет (аннелид) обра-
зовались типичные латеральные параподий, которые
были затем модифицированы и редуцированы у Cli-
tellata. Эти параподий служили полихетам уклонив-
шейся ветви для ползания, плавания и дыхания. Из
них у предкового вида членистоногих образовались
http://jurassic.ru/
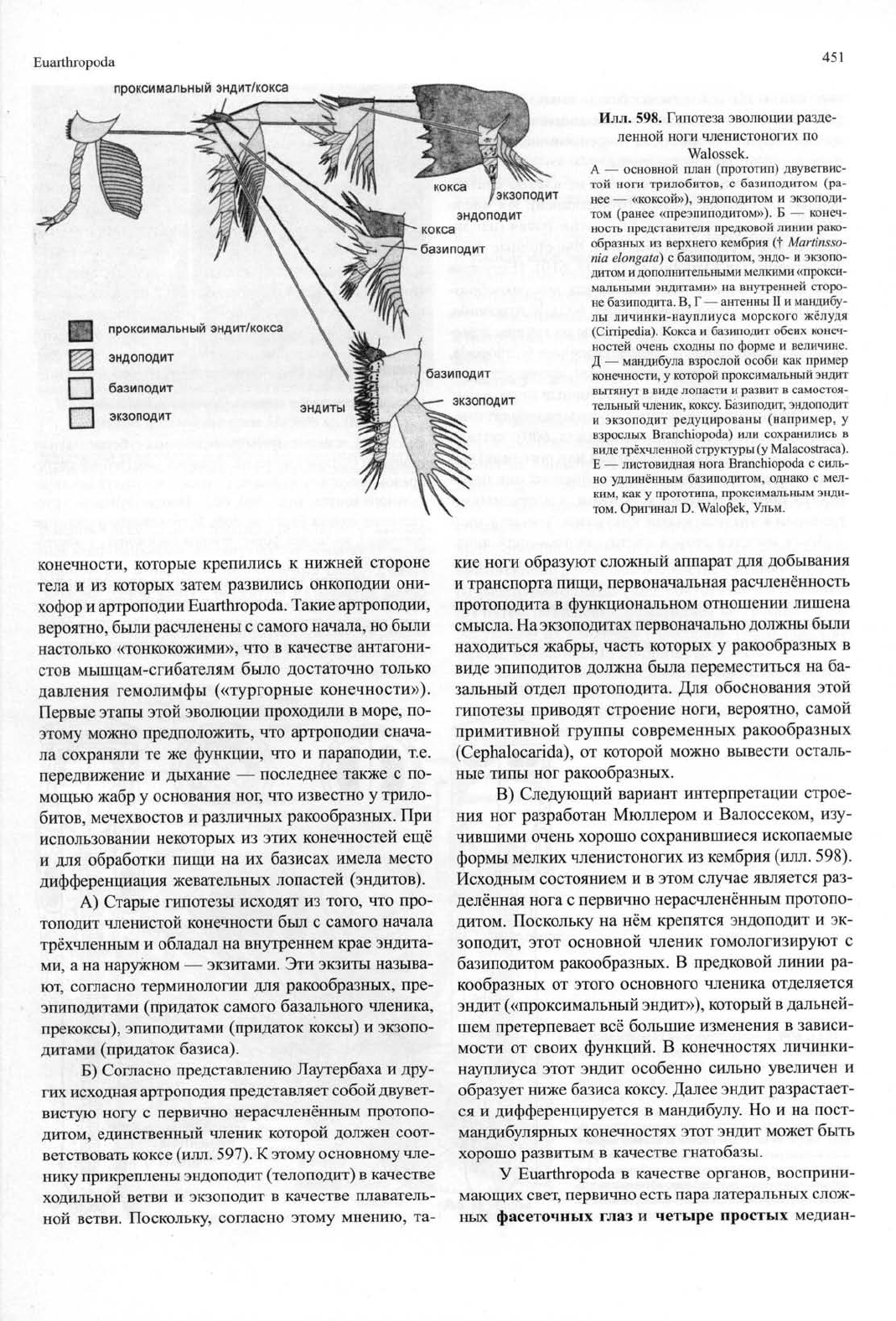
Euarthropoda
проксимальный эндит/кокса
проксимальный эндит/кокса
эндоподит
j \ базиподит
| | экзоподит
451
Илл. 598. Гипотеза эволюции разде-
ленной ноги членистоногих по
Walossek.
А — основной план (прототип) двуветвис-
той ноги трилобитов, с базиподитом (ра-
нее — «коксой»), эндоподитом и экзоподи-
том (ранее «преэпиподитом»). Б — конеч-
ность представителя предковой линии рако-
образных из верхнего кембрия (t Martinsso-
nia elongate) с базиподитом, эндо- и экзопо-
дитом и дополнительными мелкими «прокси-
мальными эндитами» на внутренней сторо-
не базиподита. В, Г — антенны II и мандибу-
лы личинки-науплиуса морского жёлудя
(Cirripedia). Кокса и базиподит обеих конеч-
ностей очень сходны по форме и величине.
Д — мандибула взрослой особи как пример
конечности, у которой проксимальный эндит
вытянут в виде лопасти и развит в самостоя-
тельный членик, коксу. Базиподит, эндоподит
и экзоподит редуцированы (например, у
взрослых Branchiopoda) или сохранились в
виде трёхчленной структуры (у Malacostraca).
Е — листовидная нога Branchiopoda с силь-
но удлинённым базиподитом, однако с мел-
ким,
как у прототипа, проксимальным энди-
том.
Оригинал D. Walo(3ek, Ульм.
конечности, которые крепились к нижней стороне
тела и из которых затем развились онкоподии они-
хофор и артроподии Euarthropoda. Такие артроподии,
вероятно, были расчленены с самого начала, но были
настолько «тонкокожими», что в качестве антагони-
стов мышцам-сгибателям было достаточно только
давления гемолимфы («тургорные конечности»).
Первые этапы этой эволюции проходили в море, по-
этому можно предположить, что артроподии снача-
ла сохраняли те же функции, что и параподии, т.е.
передвижение и дыхание — последнее также с по-
мощью жабр у основания ног, что известно у трило-
битов, мечехвостов и различных ракообразных. При
использовании некоторых из этих конечностей ещё
и для обработки пищи на их базисах имела место
дифференциация жевательных лопастей (эндитов).
А) Старые гипотезы исходят из того, что про-
топодит членистой конечности был с самого начала
трёхчленным и обладал на внутреннем крае эндита-
ми,
а на наружном — экзитами. Эти экзиты называ-
ют, согласно терминологии для ракообразных, пре-
эпиподитами (придаток самого базального членика,
прекоксы), эпиподитами (придаток коксы) и экзопо-
дитами (придаток базиса).
Б) Согласно представлению Лаутербаха и дру-
гих исходная артроподия представляет собой двувет-
вистую ногу с первично нерасчленённым протопо-
дитом, единственный членик которой должен соот-
ветствовать коксе (илл. 597). К этому основному чле-
нику прикреплены эндоподит (телоподит) в качестве
ходильной ветви и экзоподит в качестве плаватель-
ной ветви. Поскольку, согласно этому мнению, та-
кие ноги образуют сложный аппарат для добывания
и транспорта пищи, первоначальная расчленённость
протоподита в функциональном отношении лишена
смысла. На экзоподитах первоначально должны были
находиться жабры, часть которых у ракообразных в
виде эпиподитов должна была переместиться на ба-
зальный отдел протоподита. Для обоснования этой
гипотезы приводят строение ноги, вероятно, самой
примитивной группы современных ракообразных
(Cephalocarida), от которой можно вывести осталь-
ные типы ног ракообразных.
В) Следующий вариант интерпретации строе-
ния ног разработан Мюллером и Валоссеком, изу-
чившими очень хорошо сохранившиеся ископаемые
формы мелких членистоногих из кембрия (илл. 598).
Исходным состоянием и в этом случае является раз-
делённая нога с первично нерасчленённым протопо-
дитом. Поскольку на нём крепятся эндоподит и эк-
зоподит, этот основной членик гомологизируют с
базиподитом ракообразных. В предковой линии ра-
кообразных от этого основного членика отделяется
эндит («проксимальный эндит»), который в дальней-
шем претерпевает всё большие изменения в зависи-
мости от своих функций. В конечностях личинки-
науплиуса этот эндит особенно сильно увеличен и
образует ниже базиса коксу. Далее эндит разрастает-
ся и дифференцируется в мандибулу. Но и на пост-
мандибулярных конечностях этот эндит может быть
хорошо развитым в качестве гнатобазы.
У Euarthropoda в качестве органов, восприни-
мающих свет, первично есть пара латеральных слож-
ных фасеточных глаз и четыре простых медиан-
http://jurassic.ru/

452
Arthropoda
Илл. 599.
Фасеточный
глаз Syrphidae (Insecta, Diptera).
А,
Б
—
части поверхности
(секторы).
Между роговицами
омматиди-
ев
расположены тонкие
щетинки.
Оригинал
Н.
Paulus,
Вена.
ри
клеток),
псевдоконусом (внеклеточная субстанция)
или
аконусом
(четыре клетки не
образуют собственной
свето-
преломляющей
субстанции,
поэтому
омматидии
без хрус-
тального
конуса) (илл. 600, 602). Иногда функцию хрус-
тального
конуса берёт на себя кутикулярное удлинение
роговицы,
имеющее форму шишки (экзоконус, у некото-
рых
жуков).
К клеткам хрустального конуса присоединяют-
ся обычно восемь сенсорных ретинулярных кле-
ток (клетки сетчатки, или ретинулы), которые обра-
зуют в центре палочковидный проводник света -
Crustacea
Илл.
600. Схема строения омматидиев.
Продольные
и поперечные срезы на различных
уровнях.
А — Crustacea. Б — Insecta. Обе груп-
пы
имеют гомологичные клетки каждого типа:
две
корнеагенные клетки
(ядра
отмечены
чёрным
цветом)
ракообразных
соответствуют
двум глав-
ным
пигментным клеткам насекомых; хрусталь-
ный
конус из четырёх клеток и восемь первич-
ных
клеток ретинулы имеют место в обеих груп-
пах,
последние образуют у ракообразных так
называемый
слоистый рабдом, который встре-
чается
и у некоторых насекомых. Оригинал Н.
Paulus,
Вена.
две
корнеагенные
клетки
четыре
клетки
хрустального
конуса
Insecta
ных глаза. Их считают особенно важной аутомор-
фией таксона; свидетельство гомологии глаз у раз-
личных групп Euarthropoda — решающий аргумент
против «унирамной» гипотезы.
Фасеточные глаза состоят из плотно состав-
ленных отдельных глазков (омматидиев). Их число
изменяется от нескольких до многих тысяч (напри-
мер,
20
ООО
у стрекоз) (илл. 897). Фасеточные глаза
характерны для "jTrilobita (илл. 607, 610), tEuryptera
и Xiphosura (илл. 614А), так же как для ракообраз-
ных (илл. 702) и насекомых (илл. 602). У Arachnida,
Chilopoda и Diplopoda они разбиты на группы изме-
нённых омматидиев. У Notostigmophora (Chilopoda,
Scutigera) из них возникают новые, иначе устроен-
ные так называемые псевдофасеточные глаза.
Омматидии различных групп мандибулят име-
ют сходное до деталей строение (илл. 600): кутику-
лярная корнеальная линза (cornea или роговица) вы-
деляется двумя клетками: у ракообразных они назы-
ваются корнеагенными клетками, у насекомых —
главными пигментными клетками. Только у ман-
дибулят имеется второй светоулавливающий аппа-
рат— хрустальный конус. Первоначально его об-
разуют всегда четыре клетки (илл. 600).
У
насекомых хрустальный конус называют в
зависи-
мости
от внешнего вида
настоящим
конусом или эукону-
сом
(светопреломляющая конусовидная
субстанция внут-
http://jurassic.ru/
Euarthropoda
453
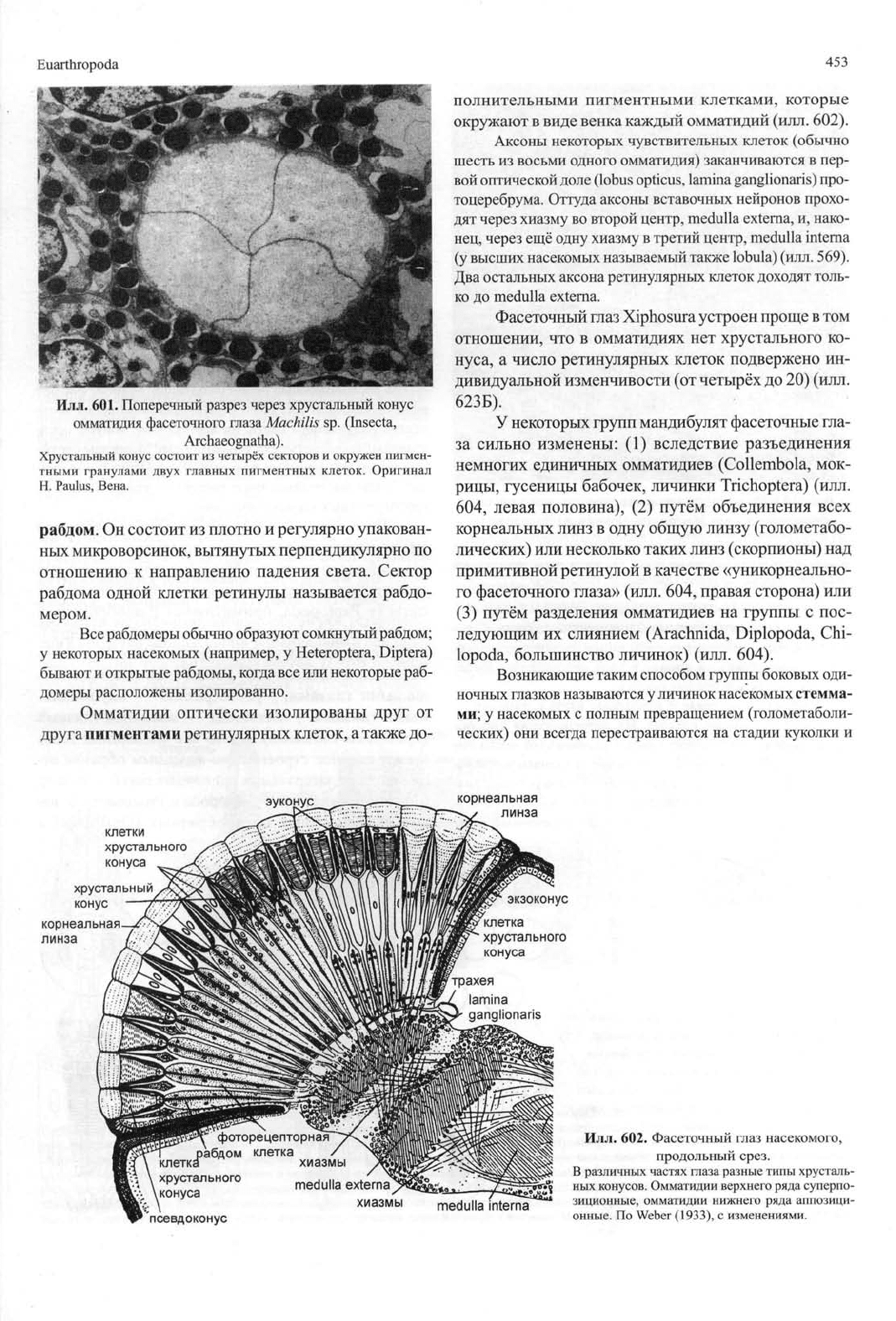
Илл.
601. Поперечный разрез через хрустальный конус
омматидия фасеточного глаза Machilis sp. (Insecta,
Archaeognatha).
Хрустальный конус состоит из четырёх секторов и окружен пигмен-
тными гранулами двух главных пигментных клеток. Оригинал
Н. Paulus, Вена.
рабдом. Он состоит из плотно и регулярно упакован-
ных микроворсинок, вытянутых перпендикулярно по
отношению к направлению падения света. Сектор
рабдома одной клетки ретинулы называется рабдо-
мером.
Все рабдомеры обычно образуют сомкнутый рабдом;
у некоторых насекомых (например, у Heteroptera, Diptera)
бывают
и
открытые рабдомы, когда все или некоторые раб-
домеры расположены изолированно.
Омматидии оптически изолированы друг от
друга пигментами ретинулярных клеток, а также до-
полнительными пигментными клетками, которые
окружают в виде венка каждый омматидии (илл. 602).
Аксоны некоторых чувствительных клеток (обычно
шесть из восьми одного омматидия) заканчиваются в пер-
вой оптической доле (lobus opticus, lamina ganglionaris) про-
тоцеребрума. Оттуда аксоны вставочных нейронов прохо-
дят через хиазму во второй центр, medulla externa, и, нако-
нец,
через ещё одну хиазму
в
третий центр, medulla interna
(у высших насекомых называемый также lobula) (илл. 569).
Два остальных аксона ретинулярных клеток доходят толь-
ко до medulla externa.
Фасеточный глаз Xiphosura устроен проще в том
отношении, что в омматидиях нет хрустального ко-
нуса, а число ретинулярных клеток подвержено ин-
дивидуальной изменчивости (от четырёх до 20) (илл.
623Б).
У некоторых групп мандибулят фасеточные гла-
за сильно изменены: (1) вследствие разъединения
немногих единичных омматидиев (Collembola, мок-
рицы, гусеницы бабочек, личинки Trichoptera) (илл.
604,
левая половина), (2) путём объединения всех
корнеальных линз в одну общую линзу (голометабо-
лических) или несколько таких линз (скорпионы) над
примитивной ретинулой в качестве «уникорнеально-
го фасеточного глаза» (илл. 604, правая сторона) или
(3) путём разделения омматидиев на группы с пос-
ледующим их слиянием (Arachnida, Diplopoda, Chi-
lopoda, большинство личинок) (илл. 604).
Возникающие таким способом группы боковых оди-
ночных глазков называются у личинок насекомых
стемма-
ми; у насекомых с полным превращением (голометаболи-
ческих) они всегда перестраиваются на стадии куколки и
эуконус
корнеапьная
линза
клетки
хрустального
конуса
хрустальный
конус
экзоконус
корнеальная
линза
medulla interna
псевдоконус
Илл.
602. Фасеточный глаз насекомого,
продольный срез.
В различных частях глаза разные типы хрусталь-
ных конусов. Омматидии верхнего ряда суперпо-
зиционные, омматидии нижнего ряда аппозици-
онные. По Weber (1933), с изменениями.
http://jurassic.ru/

454
Илл.
603.
Поперечный срез через несколько омматидиев
фасеточного глаза Machilis sp. (Insecta, Archaeognatha).
На этом уровне рабдом состоит
из
кромок микроворсинок (рабдоме-
ров) семи клеток ретинулы.
В
клетках
4 и 6
видны ядра. Рабдомер
клетки
8
находится дистально выше плоскости среза, разрезанным
оказался отросток
его
нерва. Оригинал
Н.
Paulus, Вена.
замещаются имагинальными фасеточными глазами. Толь-
ко
у
некоторых-насекомых
с
неполным превращением
(ге-
миметаболических),
а
именно
у
тлей, оба типа глаз встре-
чаются вместе
у
взрослых особей.
Фасеточные глаза
и
некоторые глаза
с
линзами
—
высокоэффективные органы.
Их
разрешающая способ-
ность зависит (при прочих равных условиях) от числа ом-
матидиев
и
угла входящего пучка света. Однако разреша-
ющая способность фасеточных глаз намного уступает гла-
зам
с
хрусталиком
у
головоногих
и
позвоночных. Непод-
вижные кутикулярные линзы лишают фасеточные глаза
Arthropoda
Илл.
604.
Отдельные омматидии сливают-
ся
в
двойные омматидии
(у
личинок
насекомых
они
называются двойными
стеммами, поскольку
все
клетки
в них в
двойном наборе)
или
путём дальнейшего
слияния
— в
сложный глаз более высоко-
го порядка.
Глаза таких типов встречаются
у
всех Chilopoda,
Diplopoda
и
многих личинок насекомых
с
пол-
ным превращением.
В
нижнем ряду показаны
продольные срезы приведённых
в
верхнем ряду
глаз
с
линзами, условно обозначенных кружка-
ми.
Оригинал
Н.
Paulus, Вена.
также способности
к
аккомодации. Некоторые виды все же
могут смещать свои ретинулы
с
помощью мышечной
ре-
тиномоторики (Salticidae /Агапеае/
и
Phyllopoda). Особен-
ное достоинство рабдомов членистоногих
— их
способ-
ность воспринимать поляризованный свет. Некоторые вы-
сокоорганизованные членистоногие (многие насекомые)
имеют хорошо развитое цветовое зрение, часто
с
охватом
ультрафиолетовой области спектра.
Для повышения количества получаемого света рако-
образные
с
ночной активностью
и
высшие насекомые вы-
работали так называемые
суперпозиционные глаза
(илл.
602).
В
отличие
от
первичных аппозиционных глаз, каж-
дый омматидии которых отделён от соседнего пигментом,
у суперпозиционных глаз отсутствуют
эти
препятствия
прохождению света.
В
таких глазах лучи света проникают
сквозь расположенные рядом омматидии. Многие высшие
раки выработали
ещё
один «трюк» для повышения коли-
чества получаемого света:
их
хрустальные конусы пред-
ставляют собой четырёхугольные полые цилиндры
с
отра-
жающими внутренними поверхностями (зеркальные лин-
зы).
Такие фасеточные глаза состоят
из
квадратных,
а не
шестиугольных корнеальных линз.
Второй тип глаз членистоногих представлен
медианными глазами. Они иннервирутся от цент-
ральной части протоцеребрума (центр науплиальных
глаз и глазков). Первично имеется четыре отдельных
глаза (у Pantopoda, примитивных ракообразных и
Insecta/Collembola), которые многократно конверген-
тно уменьшались в числе до трёх и менее. Эти меди-
анные глаза называются у пауков главными или сре-
динными глазами, у ракообразных — науплиаль-
Hi.iMii
глазами, у насекомых — лобными глазами
(простыми глазками). Часто все медианные глаза
имеют сходное строение, но коренным образом от-
личаются от латеральных одиночных глаз (см. выше).
Медианных глаз нет у Chilopoda и Progoneata. У не-
которых взрослых паукообразных (Opiliones) и
http://jurassic.ru/
Euarthropoda
455
Pantopoda, а также у науплиальных личинок ракооб-
разных
(илл.
706) они представляют собой единствен-
ные органы зрения.
Функции медианных глаз не выяснены до сих пор.
Возможно, наряду
с
регулировкой фотопериодизма живот-
ного,
они используются как «фотометр», который путём
определения окружающей освещённости регулирует чув-
ствительность ретинулярных клеток в омматидиях.
Кроме того, у эуартропод известны и другие фото-
чувствительные органы, которые часто называют фрон-
тальными (лобными) органами.
У
ракообразных (напри-
мер,
Notostraca)
это
расположенная выше (дорсальный лоб-
ный орган) или ниже науплиального глаза (вентральный
лобный орган) пара светочувствительных клеток, у кото-
рых нет кутикулярной
линзы.
Фронтальные органы встре-
чаются и у примитивных насекомых (Collembola) и, пред-
положительно, у Xiphosura (у Limulus, так называемые
брюшные глаза), но их не следует путать с негомологич-
ными синусными железами или Х-органом ракообразных
или с церебральными железами Chilopoda, которые тоже
называют фронтальными органами.
Другими широко распространёнными органа-
ми чувств являются расположенные по всему телу
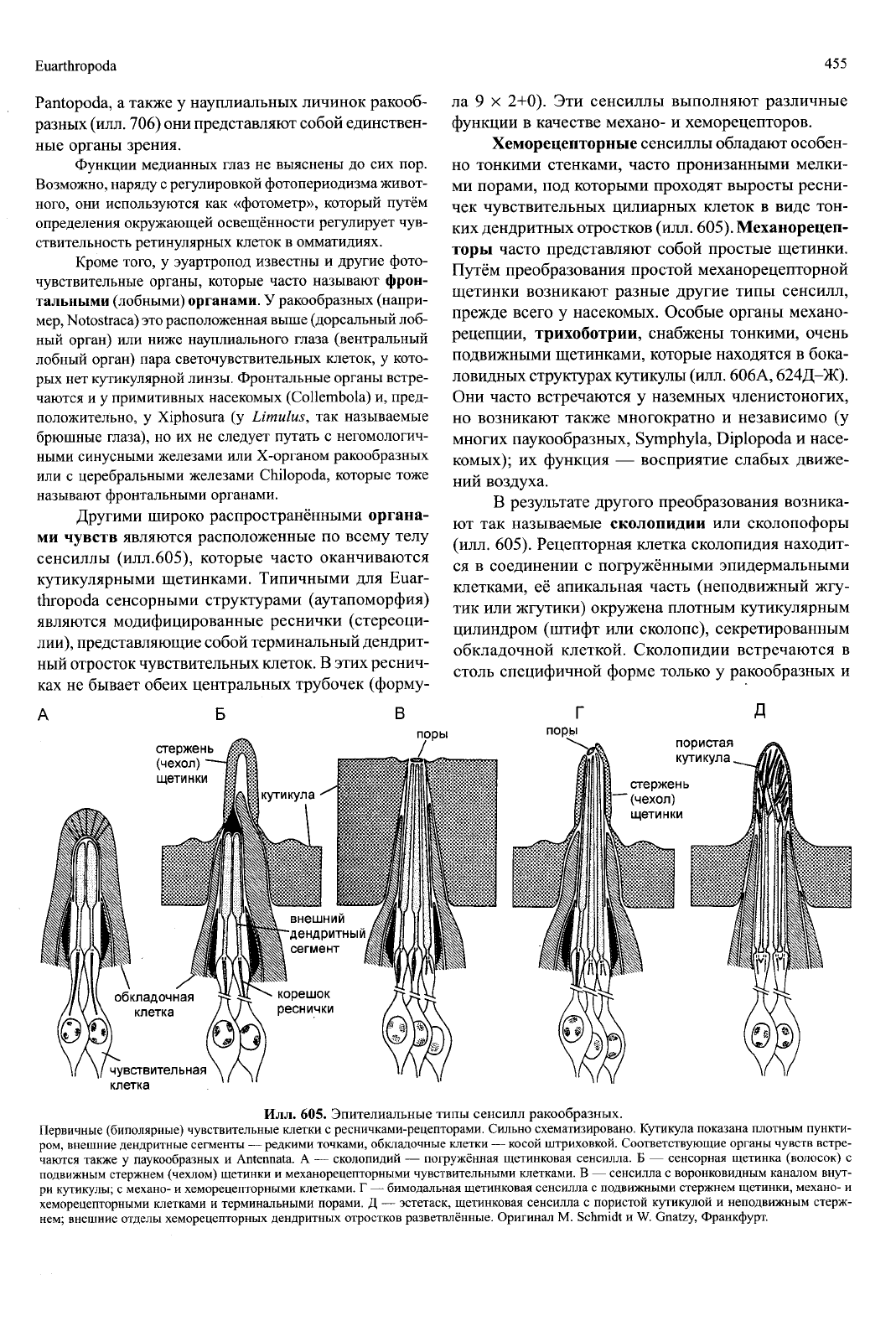
сенсиллы (илл.605), которые часто оканчиваются
кутикулярными щетинками. Типичными для Euar-
thropoda сенсорными структурами (аутапоморфия)
являются модифицированные реснички (стереоци-
лии),
представляющие собой терминальный дендрит-
ный отросток чувствительных клеток. В этих реснич-
ках не бывает обеих центральных трубочек (форму-
ла 9 х 2+0). Эти сенсиллы выполняют различные
функции в качестве механо- и хеморецепторов.
Хеморецепторные сенсиллы обладают особен-
но тонкими стенками, часто пронизанными мелки-
ми порами, под которыми проходят выросты ресни-
чек чувствительных цилиарных клеток в виде тон-
ких дендритных отростков (илл. 605). Механорецеп-
торы часто представляют собой простые щетинки.
Путём преобразования простой механорецепторной
щетинки возникают разные другие типы сенсилл,
прежде всего у насекомых. Особые органы механо-
рецепции, трихоботрии, снабжены тонкими, очень
подвижными щетинками, которые находятся в бока-
ловидных структурах кутикулы (илл. 606А, 624Д-Ж).
Они часто встречаются у наземных членистоногих,
но возникают также многократно и независимо (у
многих паукообразных, Symphyla, Diplopoda и насе-
комых); их функция — восприятие слабых движе-
ний воздуха.
В результате другого преобразования возника-
ют так называемые сколопидии или сколопофоры
(илл. 605). Рецепторная клетка сколопидия находит-
ся в соединении с погружёнными эпидермальными
клетками, её апикальная часть (неподвижный жгу-
тик или жгутики) окружена плотным кутикулярным
цилиндром (штифт или сколопс), секретированным
обкладочной клеткой. Сколопидии встречаются в
столь специфичной форме только у ракообразных и
В
Д
поры
стержень
(чехол)
щетинки
обкладочная
клетка
чувствительная
клетка
Илл. 605. Эпителиальные типы сенсилл ракообразных.
Первичные (биполярные) чувствительные клетки с ресничками-рецепторами. Сильно схематизировано. Кутикула показана плотным пункти-
ром,
внешние дендритные сегменты — редкими точками, обкладочные клетки — косой штриховкой. Соответствующие органы чувств встре-
чаются также у паукообразных и Antennata. А — сколопидии — погружённая щетинковая сенсилла. Б — сенсорная щетинка (волосок) с
подвижным стержнем (чехлом) щетинки и механорецепторными чувствительными клетками. В — сенсилла с воронковидным каналом внут-
ри кутикулы; с механо- и хеморецепторными клетками. Г — бимодальная щетинковая сенсилла с подвижными стержнем щетинки, механо- и
хеморецепторными клетками и терминальными порами. Д — эстетаск, щетинковая сенсилла с пористой кутикулой и неподвижным стерж-
нем;
внешние отделы хеморецепторных дендритных отростков разветвлённые. Оригинал М. Schmidt и W. Gnatzy, Франкфурт.
http://jurassic.ru/