Вестхайде В.,Ригер Р.( ред.) Зоология беспозвоночных в двух томах. Том 1: от простейших до моллюсков и артропод
Подождите немного. Документ загружается.

426
Arthropoda
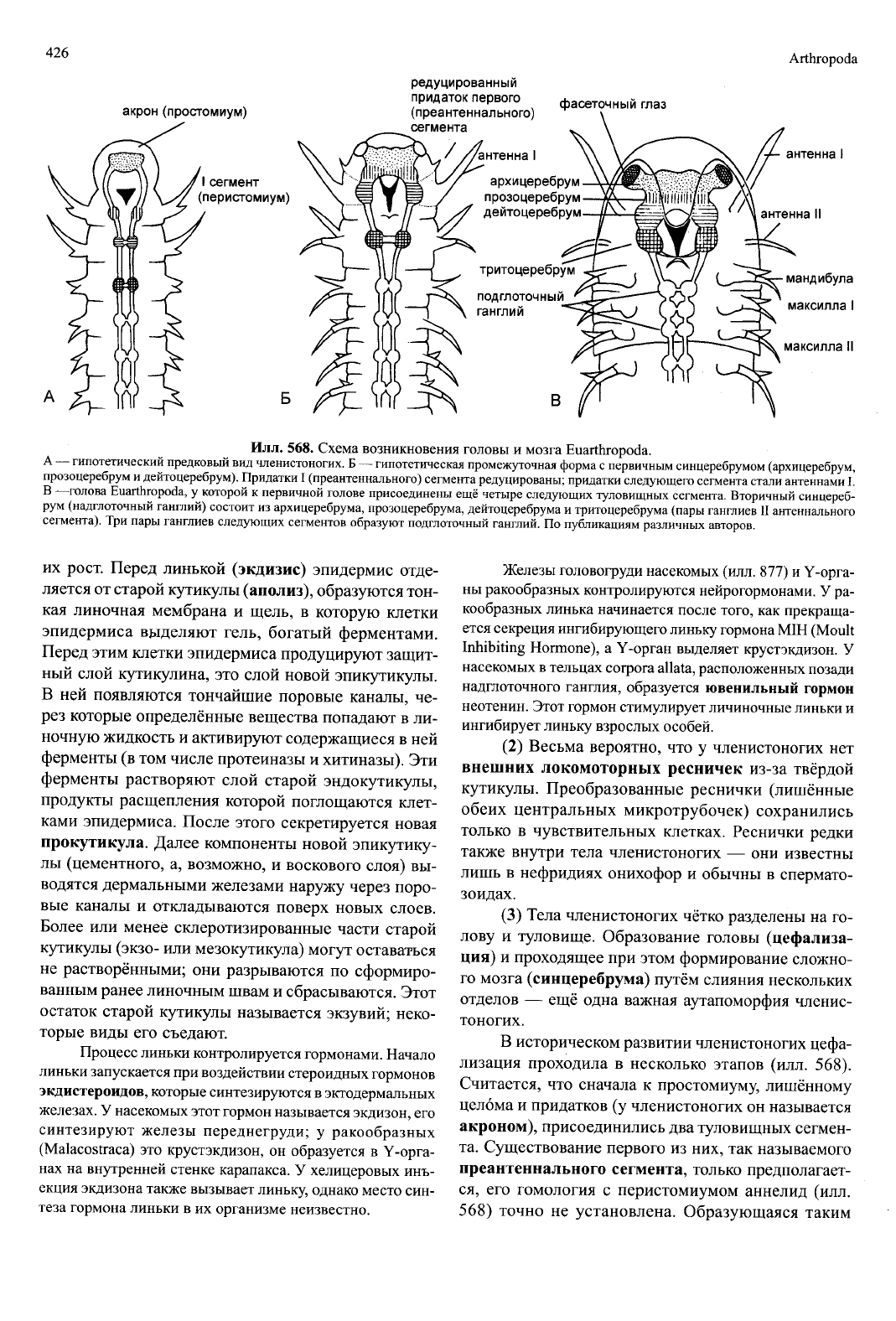
Илл.
568. Схема возникновения головы и мозга Euarthropoda.
А — гипотетический предковый вид членистоногих. Б — гипотетическая промежуточная форма с первичным синцеребрумом (архицеребрум,
прозоцеребрум и дейтоцеребрум). Придатки I (преантеннального) сегмента редуцированы; придатки следующего сегмента стали антеннами L
В —голова Euarthropoda, у которой к первичной голове присоединены ещё четыре следующих туловищных сегмента. Вторичный синцереб-
рум (надглоточный ганглий) состоит из архицеребрума, прозоцеребрума, дейтоцеребрума и тритоцеребрума (пары ганглиев II антеннального
сегмента). Три пары ганглиев следующих сегментов образуют подглоточный ганглий. По публикациям различных авторов.
их рост. Перед линькой (экдизис) эпидермис отде-
ляется от старой кутикулы (аполиз), образуются тон-
кая линочная мембрана и щель, в которую клетки
эпидермиса выделяют гель, богатый ферментами.
Перед этим клетки эпидермиса продуцируют защит-
ный слой кутикулина, это слой новой эпикутикулы.
В ней появляются тончайшие поровые каналы, че-
рез которые определённые вещества попадают в ли-
ночную жидкость и активируют содержащиеся в ней
ферменты (в том числе протеиназы и хитиназы). Эти
ферменты растворяют слой старой эндокутикулы,
продукты расщепления которой поглощаются клет-
ками эпидермиса. После этого секретируется новая
прокутикула. Далее компоненты новой эпикутику-
лы (цементного, а, возможно, и воскового слоя) вы-
водятся дермальными железами наружу через поро-
вые каналы и откладываются поверх новых слоев.
Более или менее склеротизированные части старой
кутикулы (экзо- или мезокутикула) могут оставаться
не растворёнными; они разрываются по сформиро-
ванным ранее линочным швам и сбрасываются. Этот
остаток старой кутикулы называется экзувий; неко-
торые виды его съедают.
Процесс линьки контролируется гормонами. Начало
линьки запускается при воздействии стероидных гормонов
экдистероидов, которые синтезируются
в
эктодермальных
железах.
У
насекомых этот гормон называется экдизон, его
синтезируют железы переднегруди; у ракообразных
(Malacostraca) это крустэкдизон, он образуется в Y-opra-
нах на внутренней стенке карапакса. У хелицеровых инъ-
екция экдизона также вызывает линьку, однако место син-
теза гормона линьки в их организме неизвестно.
Железы головогруди насекомых (илл. 877) и Y-opra-
ны ракообразных контролируются нейрогормонами. У ра-
кообразных линька начинается после того, как прекраща-
ется секреция ингибирующего линьку гормона МШ (Moult
Inhibiting Hormone), а Y-орган выделяет крустэкдизон. У
насекомых
в
тельцах corpora allata, расположенных позади
надглоточного ганглия, образуется ювенильный гормон
неотении. Этот гормон стимулирует личиночные линьки и
ингибирует линьку взрослых особей.
(2)
Весьма вероятно, что у членистоногих нет
внешних локомоторных ресничек из-за твёрдой
кутикулы. Преобразованные реснички (лишённые
обеих центральных микротрубочек) сохранились
только в чувствительных клетках. Реснички редки
также внутри тела членистоногих — они известны
лишь в нефридиях онихофор и обычны в спермато-
зоидах.
(3) Тела членистоногих чётко разделены на го-
лову и туловище. Образование головы (цефализа-
ция) и проходящее при этом формирование сложно-
го мозга (синцеребрума) путём слияния нескольких
отделов — ещё одна важная аутапоморфия членис-
тоногих.
В историческом развитии членистоногих цефа-
лизация проходила в несколько этапов (илл. 568).
Считается, что сначала к простомиуму, лишённому
целбма и придатков (у членистоногих он называется
акроном), присоединились два туловищных сегмен-
та. Существование первого из них, так называемого
преантеннального сегмента, только предполагает-
ся,
его гомология с перистомиумом аннелид (илл.
568) точно не установлена. Образующаяся таким
http://jurassic.ru/
Arthropoda 427
Надглоточный
ганглий
tTrilobita
Chelicerata Crustacea
«Myriapoda»
Insecta
Акрон Архицеребрум
Iпреантеннальный Прозоцеребрум
сегмент
?
II головной сегмент Дейтоцеребрум
III
головной сегмент Тритоцеребрум
Прото-
церебрум
IV
головной сегмент
V
головной сегмент
VI
головной сегмент
Подглоточные ганглии
Лабрум
?
Лабрум
?
Лабрум
?
Лабрум
?
Лабрум
?
Антенны
(1)
—
Антенны
1
Антенны
1
Антенны
1
Ходильные
Хелицеры Антенны
II
— —
ноги
1
Ходильные
Педипальпы Мандибулы Мандибулы Мандибулы
ноги
II
Ходильные
Ходильные Максиллы
1
Максиллы
1
Максиллы
(1)
ноги
III
ноги
1
Ходильные
Ходильные Максиллы
II
Максиллы
II
Лабиум
ноги
IV
ноги
II
(нижняя
губа)
образом голова (цефалон), вероятно, имела верхнюю
губу (лабрум) и одну пару антенн. Предполагают, что
губа произошла из конечностей первого сегмента.
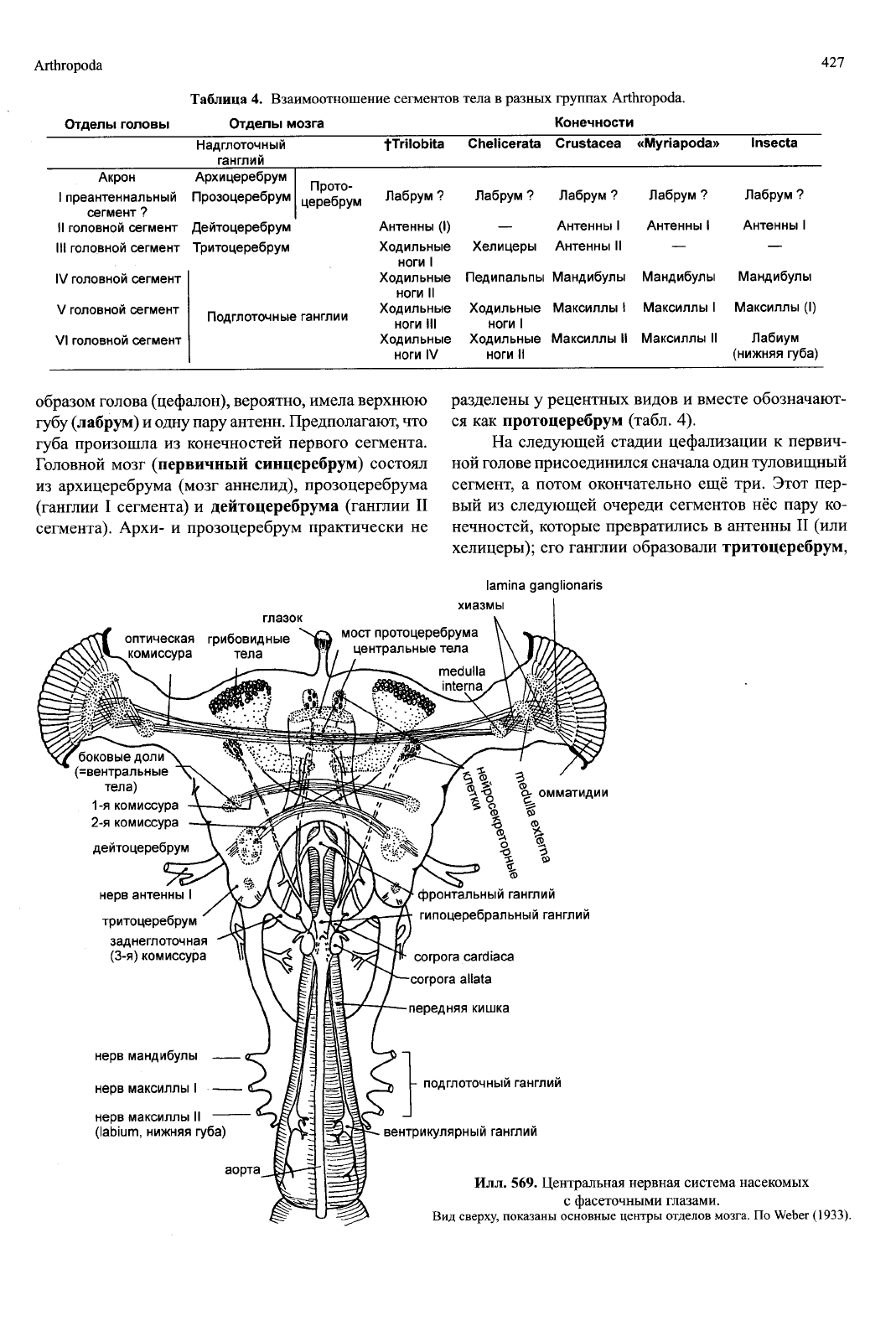
Головной мозг (первичный синцеребрум) состоял
из архицеребрума (мозг аннелид), прозоцеребрума
(ганглии I сегмента) и дейтоцеребрума (ганглии II
сегмента). Архи- и прозоцеребрум практически не
разделены у рецентных видов и вместе обозначают-
ся как протоцеребрум (табл. 4).
На следующей стадии цефализации к первич-
ной голове присоединился сначала один туловищный
сегмент, а потом окончательно ещё три. Этот пер-
вый из следующей очереди сегментов нёс пару ко-
нечностей, которые превратились в антенны II (или
хелицеры); его ганглии образовали тритоцеребрум,
lamina ganglionaris
хиазмы
гипоцеребральный ганглий
нерв мандибулы
нерв максиллы
нерв максиллы
(labium, нижняя
губа)
подглоточныи ганглии
вентрикулярныи ганглии
Илл. 569. Центральная нервная система насекомых
с фасеточными глазами.
Вид сверху, показаны основные центры отделов мозга. По Weber (1933).
Таблица
4. Взаимоотношение сегментов тела в разных группах Arthropoda.
Отделы головы Отделы мозга Конечности
http://jurassic.ru/
который объединился с расположенным перед ним
мозгом во вторичный синцеребрум. Конечности
трёх последующих сегментов стали у мандибулят
частями ротового аппарата, а ганглии этих сегмен-
тов слились в компактный подглоточный ганглий
(илл. 569, 570).
В ходе дальнейшей цефализации у многих Eu-
arthropoda с подглоточным ганглием слились и пос-
ледующие туловищные ганглии. У крабов (Вга-
chyura) вообще имеются только компактный надгло-
точный ганглий и такой же компактный подглоточ-
ный ганглий со всеми туловищными ганглиями. И,
наконец, у тенетных пауков вся центральная нервная
система представляет собой одно большую гангли-
озную массу, протянутую от пищевода
(илл.
621,622).
Итак, вся центральная нервная система возникает из
парных ганглиев. От них отходят нервы к конечнос-
тям, мышцам и внутренним органам; все вместе эти
нервы образуют периферическую нервную систему.
В мозгу находятся ассоциативные центры. Прото-
церебрум, иннервирующий глаза, состоит их двух крупных
полушарий. По бокам полушария переходят в оптические
доли (lobus opticus); у ракообразных со стебельковыми
глазами оптические доли перенесены
в
глазные стебельки.
У Chelicerata, Crustacea, Chilopoda и Progoneata зритель-
ные доли содержат два зрительных центра. У Eumalacost-
гаса и насекомых имеются даже три зрительных скопле-
ния: lamina ganglionaris, medulla externa и medulla interna
(lobula).
От
medulla interna волокна оптических нервов идут
у высших Malacostraca к medulla terminalis либо прямо к
грибовидным телам (corpora pedunculata), если таковые
имеются. Это ассоциативные центры удлинённой формы,
состоящие из плотного нейропиля. Плотно упакованные,
мелкие, хорошо прокрашиваемые тела их нервных клеток
расположены сверху и напоминают грибы. Нижние края
грибовидных тел образуют поперечное соединение в виде
балки. Corpora pedunculata имеется и у аннелид, но, воз-
можно они не гомологичны одноимённым структурам на-
секомых. Другие ассоциативные центры в протоцеребру-
ме — центральные тела, мост протоцеребрума и зритель-
ные скопления медиальных глаз. Вероятно, все эти цент-
ры являются структурами архицеребрума.
Кроме того, в состав протоцеребрума входят боко-
вые доли (у ракообразных) или вентральные тела (у насе-
комых) и их комиссура, которую считают остатком прозо-
церебрума
(1-й
комиссуры). К протоцеребруму сзади при-
http://jurassic.ru/

Arthropoda 429
мыкает дейтоцеребрум со своей комиссурой (2-я комиссу-
ра).
Он иннервирует антенны I. В тех случаях, когда эти
придатки являются важнейшими органами чувств, дейто-
церебрум образует
в
качестве ассоциативных центров круп-
ные
антеннальные клубочки (гломерулы),
в
которых соеди-
нены сенсорные нервы антеннальных органов
чувств.
Три-
тоцеребрум иннервирует антенны
И.
Если они становятся
главными органами чувств (например, у некоторых изопод
и амфипод), здесь также могут быть соответствующие ан-
теннальные
клубочки.
Это означает, что такие ассоциатив-
ные центры являются функциональными структурами, ко-
торые не всегда
гомологичны.
Например, похожие клубоч-
ки могут возникать у Amblypygi в ганглии сенсорных ко-
нечностей,
а у
лишённых антенн Protura
— в
ганглии
I
пары
ног.
У
хелицеровых, не имеющих антенн, дейтоцеребрум
не обнаружен. Их первые конечности, хелицеры, иннерви-
руются
от
тритоцеребрума
и
считаются гомологами антенн
II (см. ниже).
Внутренностная, или висцеральная нервная
система состоит из (1) каудальной части, которая от-
ходит от брюшной нервной цепочки и неодинакова
у различных таксонов, и (2) стоматогастрической
нервной системы, отходящей от тритоцеребрума
(илл. 570). Поскольку центральные части стоматога-
стрической нервной системы, фронтальный ганглий
с nervus procurrens и nervus recurrens занимают оди-
наковое положение у всех членистоногих, они пред-
ставляют собой решающий критерий для гомологи-
зации тритоцеребрума, а вместе с ним и второго ан-
теннального сегмента мандибулят, с хелицерным сег-
ментом у Chelicerata.
(4) Возможно, что вследствие образования эк-
зоскелета (у онихофор — из тонкой кутикулы и мощ-
ного базального матрикса, а у Euarthropoda—из скле-
ротизированной кутикулы), мог стать излишним гид-
роскелет в виде парных сегментарных целомических
полостей, который стабилизировал тело примитив-
ных артикулят. Первичная и вторичная полости тела
объединились друг с другом; возникшая таким спо-
собом «третичная» полость тела названа миксоце-
лем (гемоцелем) (илл. 571). Миксоцель — отличи-
тельный признак всех без исключения членистоно-
гих, так же как функционально связанная с ним не-
замкнутая кровеносная система.
В процессе эмбрионального развития ещё зак-
ладываются мезодермальные полоски и целомичес-
кие полости. Однако они рано резорбируются при
образовании туловищной и кишечной мускулатуры,
кровеносных сосудов с мускулистой сердечной труб-
кой, жировых тел и перикардиальной перегородки;
в качестве остатков целома сохраняются только го-
нады с их выводными отверстиями и целомические
мешочки нефридиев (см. ниже).
На самом деле образование полостей тела у артро-
под происходит не всегда одинаково. Например, у некото-
рых ракообразных целомические полости не исчезают, а
сжимаются и уменьшаются, прежде чем из них образуют-
ся названные выше структуры.
В результате разрушения целома кровь и цело-
мическая жидкость объединяются и образуют гемо-
лимфу. Она циркулирует в системе сосудов и полос-
ти тела, где протекает по лакунам.
От дорсомедианного сердца берёт начало ар-
териальная система, которая хорошо развита у при-
митивных и особенно у большинства крупных форм.
Она состоит из: (1) парных сегментарных боковых
сосудов, которые образуются в зачатках септ между
сегментами, (2) передней аорты и задней артерии и
(3) часто также из брюшного сосуда, идущего вдоль
брюшной нервной цепочки, дорсально или вентраль-
но от неё. Сердце, которое первоначально распола-
галось вдоль всего тела, всё больше концентрирует-
ся в той части туловища, где находятся органы дыха-
ния. Как правило, гемолимфа поступает в аорты и
артерии вследствие сокращения кольцевых мышц
сердца (систола). Диастола происходит за счёт со-
кращений крыловидных мышц, которые находятся
между сегментами в перегородке перикарда и про-
тянуты от сердца к стенке тела. Клапаны сосудов,
отходящих от сердца, не дают гемолимфе попадать
обратно в сердце. Направляющаяся в туловище ге-
молимфа по лакунам подходит к органам, далее
(обычно пройдя через органы дыхания) попадает по
боковым протокам обратно в перикардиальный си-
нус и оттуда через сегментарные щели (остии) —
обратно в сердце. Остии снабжены полулунными
клапанами, которые закрываются во время систолы
и открываются при диастоле. В гемолимфе могут
содержаться клетки семи различных типов (гемоци-
ты),
которые, однако, не у всех групп членистоногих
есть в полном наборе.
Дыхательными пигментами служат гемоциа-
нин, реже гемоглобин (например, у красных личи-
нок Chironomus и некоторых видов Daphnia); пиг-
менты не связаны с клетками крови.
С появлением трахей (см. ниже) гемолимфа утрачи-
вает функцию переноса газов (за исключением Notostig-
mophora), в связи с чем сильно редуцируются артерии (на-
пример, у Antennata). Однако артериальная система также
редуцируется независимо от формы дыхания, так что ос-
таётся сердце в форме мешочка с одной или немногими
парами остий. Многие мелкие формы, например копепо-
ды,
остракоды, клещи, вообще лишены сердца. Циркуля-
ция гемолимфы обеспечивается у них движениями кишеч-
ника и туловища. Чтобы снабжать кровью конечности,
имеются различные дополнительные органы в виде сер-
дец, антенн, крыльев, церок (у Plecoptera) и иногда диаф-
рагм в ногах.
http://jurassic.ru/
430
Arthropoda
спинные продольные мышцы перикардиальный синус
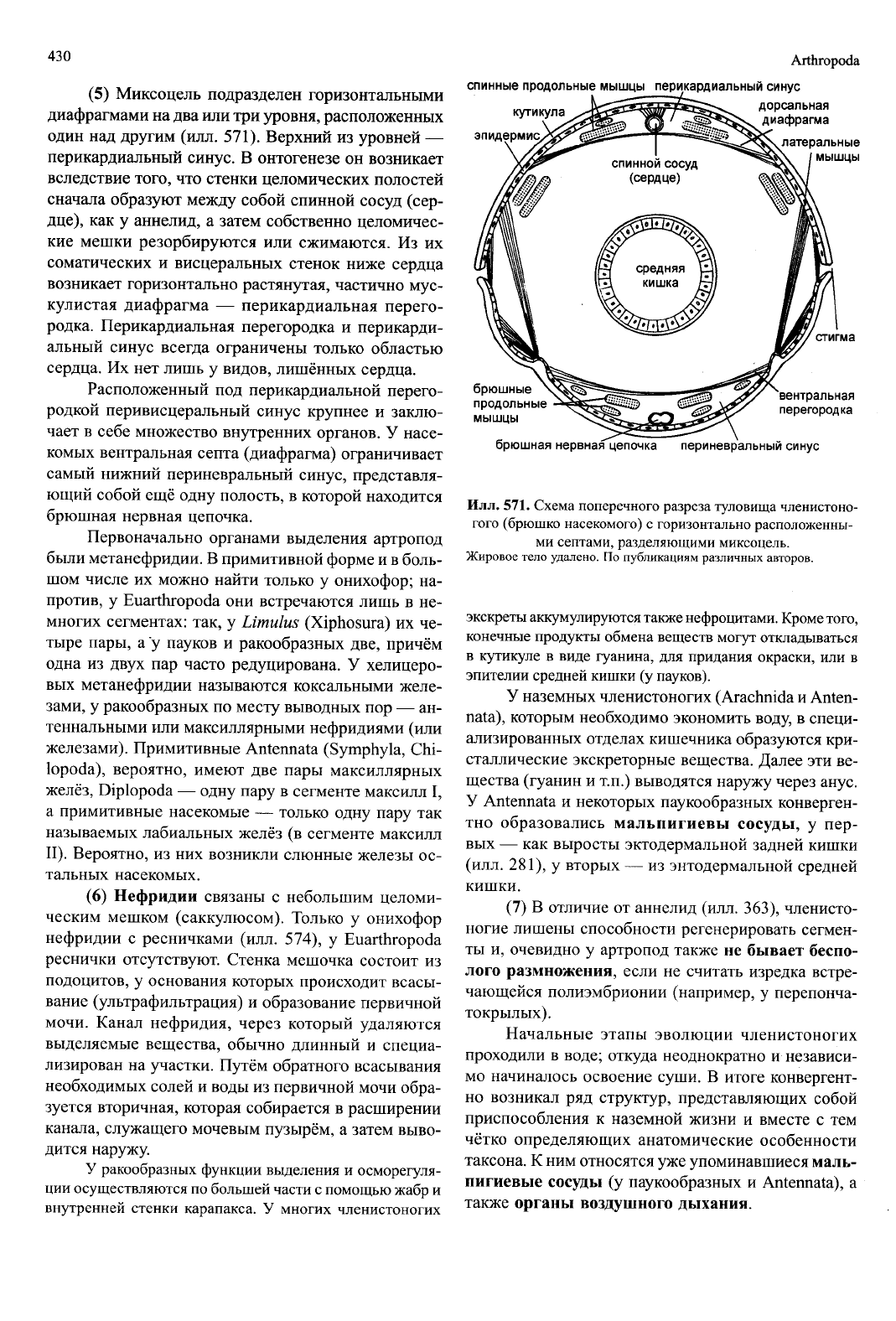
Илл.
571.
Схема поперечного разреза туловища членистоно-
гого (брюшко насекомого) с горизонтально расположенны-
ми септами, разделяющими миксоцель.
Жировое тело удалено. По публикациям различных авторов.
экскреты аккумулируются также нефроцитами. Кроме того,
конечные продукты обмена веществ могут откладываться
в кутикуле в виде гуанина, для придания окраски, или в
эпителии средней кишки (у пауков).
У
наземных членистоногих (Arachnida и Anten-
nata),
которым необходимо экономить воду, в специ-
ализированных отделах кишечника образуются кри-
сталлические экскреторные вещества. Далее эти ве-
щества (гуанин и т.п.) выводятся наружу через анус.
У
Antennata и некоторых паукообразных конверген-
тно образовались мальпигиевы сосуды, у пер-
вых — как выросты эктодермальной задней кишки
(илл.
281),
у вторых — из энтодермальной средней
кишки.
(7)
В отличие от аннелид (илл. 363), членисто-
ногие лишены способности регенерировать сегмен-
ты и, очевидно у артропод также не бывает беспо-
лого размножения, если не считать изредка встре-
чающейся полиэмбрионии (например, у перепонча-
токрылых).
Начальные этапы эволюции членистоногих
проходили в воде; откуда неоднократно и независи-
мо начиналось освоение суши. В итоге конвергент-
но возникал ряд структур, представляющих собой
приспособления к наземной жизни и вместе с тем
чётко определяющих анатомические особенности
таксона. К ним относятся уже упоминавшиеся маль-
пигиевые сосуды (у паукообразных и Antennata), а
также органы воздушного дыхания.
(5) Миксоцель подразделен горизонтальными
диафрагмами на два или три уровня, расположенных
один над другим (илл. 571). Верхний из уровней —
перикардиальный синус. В онтогенезе он возникает
вследствие того, что стенки целомических полостей
сначала образуют между собой спинной сосуд (сер-
дце),
как у аннелид, а затем собственно целомичес-
кие мешки резорбируются или сжимаются. Из их
соматических и висцеральных стенок ниже сердца
возникает горизонтально растянутая, частично мус-
кулистая диафрагма — перикардиальная перего-
родка. Перикардиальная перегородка и перикарди-
альный синус всегда ограничены только областью
сердца. Их нет лишь у видов, лишённых сердца.
Расположенный под перикардиальной перего-
родкой перивисцеральный синус крупнее и заклю-
чает в себе множество внутренних органов. У насе-
комых вентральная септа (диафрагма) ограничивает
самый нижний периневральный синус, представля-
ющий собой ещё одну полость, в которой находится
брюшная нервная цепочка.
Первоначально органами выделения артропод
были метанефридии. В примитивной форме и в боль-
шом числе их можно найти только у онихофор; на-
против, у Euarthropoda они встречаются лишь в не-
многих сегментах: так, у Limulus (Xiphosura) их че-
тыре пары, а
"у
пауков и ракообразных две, причём
одна из двух пар часто редуцирована. У хелицеро-
вых метанефридии называются коксальными желе-
зами, у ракообразных по месту выводных пор — ан-
теннальными или максиллярными нефридиями (или
железами). Примитивные Antennata (Symphyla, Chi-
lopoda), вероятно, имеют две пары максиллярных
желёз, Diplopoda — одну пару в сегменте максилл I,
а примитивные насекомые — только одну пару так
называемых лабиальных желёз (в сегменте максилл
II).
Вероятно, из них возникли слюнные железы ос-
тальных насекомых.
(6) Нефридии связаны с небольшим целоми-
ческим мешком (саккулюсом). Только у онихофор
нефридии с ресничками (илл. 574), у Euarthropoda
реснички отсутствуют. Стенка мешочка состоит из
подоцитов, у основания которых происходит всасы-
вание (ультрафильтрация) и образование первичной
мочи. Канал нефридия, через который удаляются
выделяемые вещества, обычно длинный и специа-
лизирован на участки. Путём обратного всасывания
необходимых солей и воды из первичной мочи обра-
зуется вторичная, которая собирается в расширении
канала, служащего мочевым пузырём, а затем выво-
дится наружу.
У ракообразных функции выделения и осморегуля-
ции осуществляются по большей части с помощью жабр и
внутренней стенки карапакса. У многих членистоногих
http://jurassic.ru/

Arthropoda
431
Илл. 572.
Родственные отношения групп членистоногих
(кроме
Tardigrada).
Апоморфии:
[1]
Кутикула
из
а-хитина
и
протеинов, линька; утрата
внешних локомоторных ресничек; цефализация
и
образование
син-
церебрума; миксоцель
и
незамкнутая кровеносная система; перикар-
диальная перегородка; нефридии
с
целомический мешочком.
[2] Ко-
роткие культевидные ноги
без
суставов; нерегулярно распределён-
ные пучки трахей;
на
один сегмент приходится несколько комиссур
между двумя продольными брюшными стволами; защитные железы
с ротовыми папиллами.
[3]
Скелет
из
склеритов; цефалон состоит
из
акрона
и,
вероятно, пяти сегментов
— с
одной парой предротовых
членистых антенн
и
тремя парами других конечностей; одна пара
латеральных фасеточных глаз
и
четыре медианных глазка; нефридии
в четырёх головных
и
двух туловищных сегментах.
[4]
Разделение
туловища
на
просому (вероятно,
из
семи сегментов)
с
шестью пара-
ми конечностей
и
опистосому (двенадцать сегментов); сегмент
ан-
тенн
II
несёт хелицеры
в
качестве первой пары конечностей.
[5] Го-
лова
из
акрона
и,
вероятно, шести сегментов,
с
пятью парами конеч-
ностей (антенны
I и II,
мандибулы, максиллы
I и
II); особое строение
омматидиев фасеточных глаз; железы гормона линьки
и
т.д.
[6]
науп-
лиальный глаз;
две
пары нефридиев
в
сегменте антенн
II и
сегменте
максилл
II;
науплиус
как
первая постэмбриональная стадия; другие
аутапоморфии ракообразных.
[7]
Сегмент антенн
II
(интеркалярный
сегмент)
без
конечностей; другие апоморфии
см.
Обсуждение.
У водных форм дыхание осуществляется всей
поверхностью тела
или
ограничено специальными
участками утончённой кутикулы (жабры, внутренняя
стенка карапакса).
У
наземных членистоногих раз-
вились следующие органы воздушного дыхания:
(1)
удлинённые наружу выросты жабр морских предков
преобразовались
у
наземных хелицеровых
в
склад-
чатые веерные лёгкие
или
лёгочные мешки
(илл.
625,
626); (2)
у
некоторых наземных декапод
в
подо-
бие лёгких превращена полость карапакса;
(3) у
многих сухопутных мокриц (ракообразные) есть пле-
оподные лёгкие
или
псевдотрахеи,
это
впячивания
экзоподитов плеоподов (илл. 802);
(4)
многократно
и независимо происходило образование трахей
—
трубковидных впячиваний поверхности тела.
У
они-
хофор это нерегулярно распределённые по телу оди-
ночные трахеи
или
пучки трахей,
без
замыкающих
аппаратов
(илл.
577).
У
паукообразных трахеи воз-
никают
из
веерных лёгких или энтапофизов
и
могут
располагаться посегментно (Araneae, Solifugae)
или
как-то иначе
(у
Opiliones, Acari).
Для
Antennata
(их
второе название Tracheata!) характерны первично
сегментарные
или
расположенные попарно трахеи,
открывающиеся
в
стигмы, которые
у
большинства
видов имеют устройства для закрывания
и
экономии
воды.
У водных насекомых имеются вторичные трахейные
жабры
или
кровеносные жабры, образованные либо
из
преобразованных конечностей абдомена (например,
у
Ме-
galoptera, Coleoptera: Gyrinidae, Ephemeroptera), либо
из
простых выпячиваний кожи (Plecoptera, Trichoptera).
Раннее эмбриональное развитие
в
типичном
случае формировалось, вероятно, также конвергент-
но,
независимо
от
перехода
к
наземной жизни.
Как
правило, развитие меробластическое. Центролеци-
тальные, богатые желтком яйца дробятся поверхно-
стно;
этот способ дробления присущ всем группам
членистоногих (илл. 870).
Но у
онихофор наблюда-
ются многочисленные переходы между полным рав-
номерным
и
поверхностным дроблением.
У
ракооб-
разных (Anostraca, Phyllopoda, Ostracoda, Copepoda,
Cirripedia
и
некоторые Decapoda) тоже бывают бед-
ные желтком яйца
и
полное дробление (голобласти-
ческое развитие),
а
также полный спектр переходов
к поверхностному дроблению. Это полное, детерми-
нированное дробление демонстрирует различные
генеалогии клеток, напоминающие спиральное дроб-
ление. Представляет
ли
такое полное дробление
ис-
ходный модус начального эмбриогенеза членистоно-
гих, остаётся непонятным; возможно, поверхностное
дробление возникало многократно
и
конвергентно.
У многих насекомых (паразитические Hymenoptera)
и паукообразных (живородящие скорпионы
и лож-
носкорпионы) также может быть полное дробление.
Но
в
этих случаях
оно с
высокой вероятностью
вы-
водится
из
поверхностного дробления; яйца этих
организмов стали вторично олиголецитальными,
приспособившись
к
особым условиям развития.
Систематика
Вопрос
о
монофилии
или
полифилии членис-
тоногих
как
целого таксона
не
имеет сейчас одно-
значного решения.
В
немецкоязычной литературе
(и
в этом учебном пособии) таксон традиционно счи-
тается монофилетическим, а приведённые выше при-
http://jurassic.ru/

432
Arthropoda
Илл.
573. Реконструкция особей \Anomalocaris из сланцев
Берджес (Burgess Shele) Британской Колумбии
(возраст 530 млн. лет).
Это загадочные ископаемые организмы относят к вымершей группе
членистоногих или, по крайней мере, к группе, близкой к членисто-
ногим. К ним относят также находки из Австралии и Китая. По-на-
стоящему крупные (60 см) животные, возможно, умели плавать. Круг-
лую структуру на нижней стороне тела считают ротовым отверсти-
ем.
Большие передние придатки служили, вероятно, для добывания
пищи. Из Gould (1989).
другим группам, не относящимся к Articulata, чем
группам членистоногих. Наконец, гипотеза незави-
симого происхождения таксонов членистоногих оз-
начает, что лишённая ресничек кутикула из хитина и
белков, её специфическая линька, миксоцель с пери-
кардиальной перегородкой, дорсально расположен-
ным сердцем и незамкнутой кровеносной системой,
нефридии с целомическим мешочком и сложный
мозг не являются у этих групп гомологичными, а
должны были возникнуть независимо. Этому нет
никаких свидетельств или, по крайней мере, — убе-
дительных. Также конвергентным образованием дол-
жны быть в этом случае и фасеточные глаза различ-
ных таксонов Euarthropoda. В действительности, их
составные части можно однозначно гомологизиро-
вать на основе сходной ультраструктуры (илл. 600).
Поэтому «унирамная» гипотеза должна быть призна-
на весьма маловероятной
Дополнения (с. 896).
знаки — хорошо обоснованными апоморфиями, воз-
никавшими в предковой линии членистоногих толь-
ко один раз (илл. 572). Наличие этих апоморфии ука-
зывает на высокую вероятность происхождения всех
членистоногих только от одного общего предково-
го вида.
Полифилетическое происхождение членистоно-
гих предполагается, прежде всего, в англосаксонс-
кой литературе (Manton, Tiegs, Anderson). Согласно
этой точке зрения членистоногие возникли незави-
симо несколько раз от предков, похожих на аннелид.
Одна из линий должна вести к онихофорам и
«Myriapoda» и от них — к насекомым; все эти груп-
пы объединяются в составе «Uniramia». При этом
развитие шло в направлении увеличения числа ко-
нечностей, входящих в состав ротового аппарата: от
моногнатии (только одна пара челюстей) у онихофор,
через дигнатию (мандибулы и гнатохиларий) Diplo-
poda и Pauropoda к тригнатии (мандибулы, максил-
лы I и максиллы II) Chilopoda и насекомых. В каче-
стве важного общего признака этих групп выделяют
«унирамные» (одноветвистые) конечности и так на-
зываемые «мандибулы целой ноги» (это мандибулы,
которые соответствуют всей преобразованной конеч-
ности, где режущая часть находится дистально). Со-
гласно этим представлениям ракообразные должны
были пройти независимую эволюцию, поскольку их
мандибулы происходят из базальных отделов ног
(илл. 598).
«Челюсти» онихофор действительно являются
мандибулами целой ноги, но мандибулы Chilopoda и
насекомых устроены иначе, как видно из сравнения
коксальных мышечных тяжей — таким образом, эта
аргументация лишена оснований. Впрочем, эти ко-
нечности у онихофор и Antennata находятся на раз-
личных головных сегментах и никак не могут быть
гомологичными. «Дигнатия» Diplopoda основана на
редукции максилл II, таким образом, это вторичное
явление.
Другие независимые линии должны вести к
хелицеровым и f Trilobita. Однако ни в одном из этих
случаев невозможно показать синапоморфиями, что
эти «линии» ближе к каким-либо аннелидам или даже
http://jurassic.ru/

1.
Onychophora,
Онихофоры,
или
Первичнотрахейные
К онихофорам относится около
160
внешне
очень сходных между собой рецентных видов. Хотя
эта группа маленькая,
она
очень важна
в
плане
фи-
логенетики
и
зоогеографии.
Как
эволюционно древ-
ние формы они занимают ключевое положение меж-
ду аннелидами
и
Euarthropoda. Часто онихофоры
противопоставляются эуартроподам
в
качестве Prot-
arthropoda или Pararthropoda. Организация этих орга-
низмов характеризуется мозаичным распределением
признаков аннелид
и
членистоногих,
но
вместе с тем
отмечена высокой степенью своеобразия. Подобно
другим сухопутным членистоногим, онихофоры
ха-
рактеризуются прямым развитием
с
периодически-
ми линьками. Похожие
на
онихофор ископаемые
ос-
танки известны из нижнего кембрия, преимуществен-
но
из
морских отложений;
в
отличие
от
них, рецепт-
ные онихофоры
— все без
исключения наземные.
Поскольку
они
плохо защищены
от
испарения
(от-
верстия
их
трахей
не
закрываются,
а
кутикула
не
имеет воскового слоя), онихофоры приурочены
к
биотопам
с
высокой влажностью (около 95%)
и по-
стоянной (обычно 18°С) температурой.
Эти
живот-
ные распространены в тропической, субтропической
и умеренной зонах, преимущественно, Южного
по-
лушария (илл. 585).
Они
активны
в
ночное время
и
прячутся во влажной
и
мягкой почве, внутри трухля-
вых стволов деревьев и под ними, под плоскими кам-
нями,
в
листовом опаде, мхе или
в
грунте береговых
откосов.
Строение
Гомономно расчленённое червеобразное тело,
сильно выпуклое со спинной стороны
и
уплощённое
вентрально (илл. 574). Голова чётко
не
выделяется.
Hilke Puhberg, Гамбург
надглоточный
ганглий
антенна
слизевая
(защитная)
железа
пищевод
На
ней
расположены одна пара мелких глаз
и три
пары видоизмененных придатков:
(1)
антенны,
(2)
ротовые крючья или челюсти («мандибулы»)
в
каче-
стве единственной части ротового аппарата
и (3)
бо-
ковые оральные папиллы,
на
которых открываются
протоки мощных защитных (слизевых) желёз, тяну-
щихся почти вдоль всего тела. Далее
с
равными про-
межутками следуют остальные конечности сегмен-
тов:
13-43 пары ходильных ног (ножки, онкоподии).
Прикреплённые
к
туловищу вентролатерально,
ноги состоят
из
конического основания
и
узкой тер-
минальной лапки (илл.
576,
578). Лапка состоит
из
колючих кольцевых подошвенных утолщений
и од-
ной пары подвижных коготков.
В
основании ноги
вентрально располагаются выпячиваемые коксаль-
ные мешочки
и
поры большинства нефридиев. У сам-
цов
там же
находятся круральные папиллы.
Поверхность тела кольчатая; однако истинная
сегментация прослеживается снаружи только по рас-
положению конечностей. Анус терминальный, поло-
вое отверстие находится перед анусом.
Мягкая, бархатистая, способная растягиваться
кутикула онихофор (отсюда английское название
—
«вельветовые черви»)
в
течение всей жизни сменя-
ется раз
в
две-три недели. Она состоит из тонкой
(1—
3
мкм)
прокутикулы,
с
комплексом
из
а-хитина
и
протеина,
и
лежащей
над ней
четырёхслойной
эпи-
кутикулы.
Под эпидермисом расположен необычно мощ-
ный (до
30
мкм!) базальный матрикс
с
коллагеновы-
ми волокнами; они являются важной составной час-
тью хорошо развитого кожно-мускульного мешка
и
в основном определяют внешнюю форму
и
проч-
ность животного.
В то
время
как у
Euarthropoda
из-
менения формы тело жёстко ограничены сильно скле-
ротизированной экзокутикулой,
у
Onychophora,
на-
спинной сосуд
(сердце)
матка
с
эмбрионами
околоротовои
сосочек
рот
нефридии
Илл. 574. Анатомия самки Onychophora, вид сбоку. По Pearse и Buchsbaum (1987), с изменениями.
мешочек
слюнной
железы
брюшной
нервный
ствол
http://jurassic.ru/

434 Arthropoda
оборот, форма тела может меняться очень сильно, что
обеспечивает тонкая кутикула вместе
с
эластичным
базальным матриксом.
Так,
Peripatopsis,
имеющий длину
3
см
в
состоянии
покоя,
при
ходьбе легко растягивается вдвое
и,
соответ-
ственно, становится
уже;
местами диаметр тела может
уменьшаться до
1/9
части от первоначального значения.
Вся поверхность тела онихофор густо усажена
папиллами, которые покрыты чешуйками
и
различа-
ются окраской, формой
и
размерами. Большинство
этих папилл снабжены чувствительными щетинка-
ми и служат механо- или хеморецепторами (илл. 577).
Мускулатура
не
разделена посегментно,
в от-
личие
от
типичного (исходного) состояния
у
анне-
лид. Она состоит из внешнего слоя кольцевых мышц
и расположенных
под
ними косых
и
сильных про-
дольных мышц.
В
ходильные ноги заходят
и
дорсо-
вентральные,
и
поперечные мышцы (илл.
576).
Согласно новым данным электронной микроскопии
мышечные волокна покровов тела онихофор отличаются
от поперечнополосатых мышц Euarthropoda
и от
косо ис-
черченных мышц аннелид: они демонстрируют специфи-
ческую форму поперечной исчерченности, которые обес-
печивают более сильное изменение длины, чем обычные
поперечнополосатые мышцы.
Полость тела
—
миксоцель (гемоцель), кото-
рая закладывается
у
эмбриона как вторичная полость
тела (целом),
но в
процессе дальнейшего развития
она преобразуется или резорбируется
и
объединяет-
ся
с
первичной полостью тела. Остатки целома
со-
храняются
в
виде мешочков нефридиев
и
слюнных
желёз,
а
также полости гонад.
На
поперечном срезе
тела можно различить типичную анатомическую
организацию онихофор: боковые дорсовентральные
мышцы разграничивают большую медианную
по-
лость
и
меньшие парные боковые камеры (латераль-
ные синусы) (илл. 576).
В
спинной части перикар-
диальная септа (дорсальная диафрагма) отделяет пе-
рикардиальный синус
от
висцерального.
Нервная система состоит
из
большого парно-
го надглоточного ганглия
над
пищеводом,
и
брюш-
ной нервной цепочки (илл. 574).
Два
вентральных
продольных ствола цепочки широко отделены друг
от друга
и в
каждом метамере соединены девятью-
десятью тонкими комиссурами.
В
каждом сегменте
от продольных стволов отходят только восемь
не-
рвов,
включая
два
ножных нерва. Таким образом,
число комиссур
не
соответствует числу нервов
в од-
ном сегменте. Также отсутствуют указания
на
лате-
ральные
и
дорсальные продольные нервы.
Только
в
сердце онихофор проходит либо один дор-
сальный нерв (у Peripatidae), либо три (у Peripatopsidae).
Клеточные тела вентральных нейронов распре-
делены более
или
менее регулярно
над
продольны-
ми стволами;
над
ними находится система нервных
волокон, среди которых
и
гигантские волокна.
По-
этому ганглии чётко
не
различаются.
Тем не
менее,
нервная система состоит не из нервных тяжей,
а
раз-
делена
на
нейромеры.
В онтогенезе нервная система формируется
из
парных сегментарных эктодермальных утолщений.
Они отчётливо видны
и у
взрослых животных
—
посредине между ходильными ногами (так называе-
мые «вентральные органы»),
а
также
и
под мозгом
в
качестве инфрацеребрального органа (гипоцереб-
рального тела);
эти
органы выполняют,
в
том числе,
и нейросекреторную функцию. Мозг (надглоточный
ганглий) состоит
из
трёх отделов: прото-, дейто-
и
тритоцеребрума. Имеются непарное центральное
тело,
три
глобулы грибовидных
тел
(типичные
для
артропод ассоциативные центры) и мост протоцереб-
рума. Большой дейтоцеребрум образует второй
от-
дел головного мозга;
от
него отходят нервы антенн,
которые
в
онтогенезе закладываются позади
рта.
Сзади
от
дейтоцеребрума расположен существенно
более мелкий тритоцеребрум. От него идут нервы
ко
рту
и к
передней кишке, образующие стоматогаст-
рическую нервную систему.
Под
глоткой проходят
комиссуры тритоцеребрума.
Два пузыревидных глаза с линзой (диаметром
0,2-0,3
мм) расположены дорсально
у
основания ан-
тенн (илл. 581
А).
Они иннервируются архицеребру-
мом.
В
онтогенезе глаза возникают
из
эктодермы:
зачатки впячиваются
и
отшнуровываются.
Глаза онихофор служат, главным образом,
для
вос-
приятия направления
и
интенсивности
освещения,
посколь-
ку,
будучи
ночными животными, онихофоры избегают
днев-
ного света.
Typhloperipatus williamsoni
и
Tasmanipatus
http://jurassic.ru/
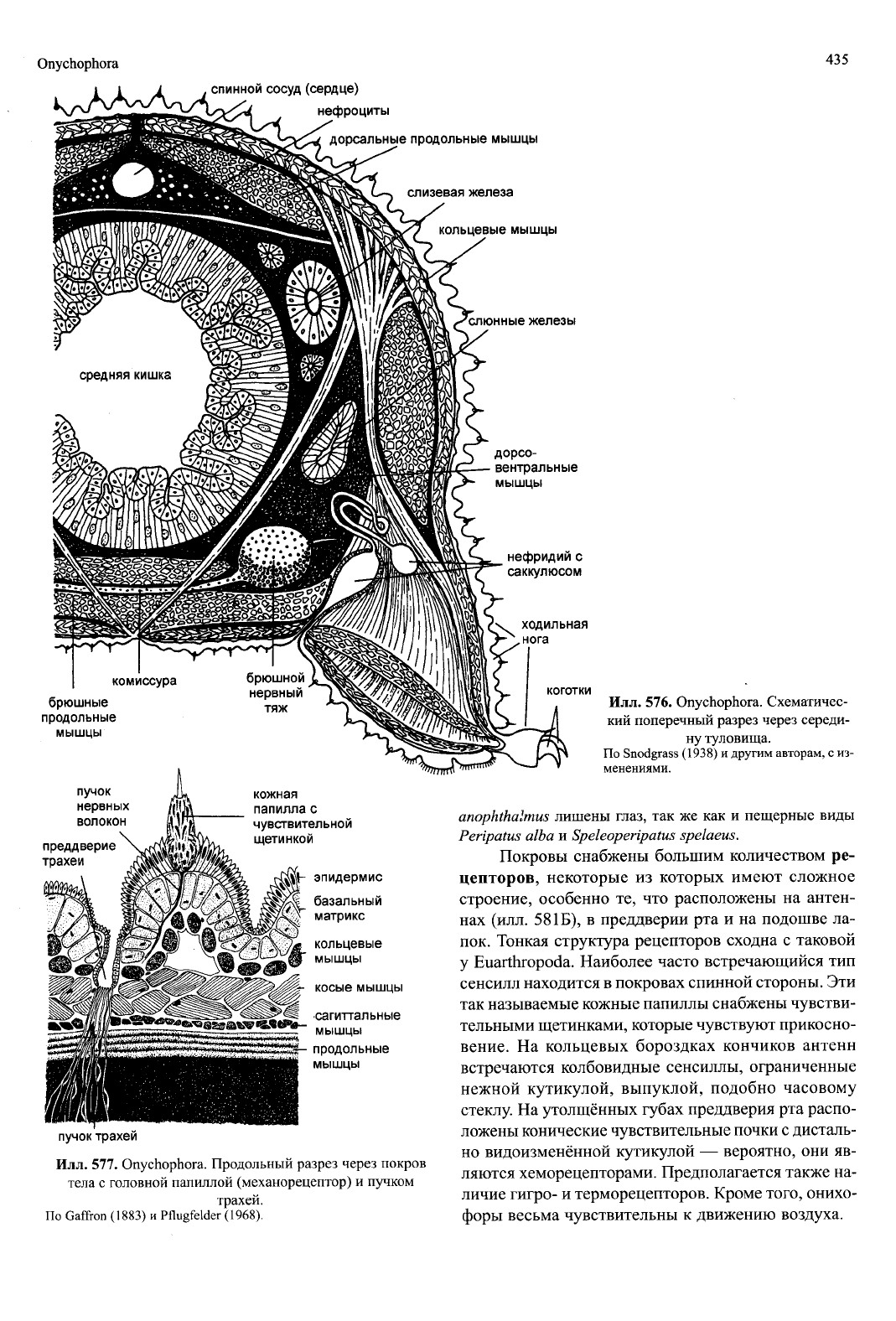
Onychophora
435
,
спинной сосуд
(сердце)
нефроциты
дорсальные продольные мышцы
слизевая
железа
кольцевые мышцы
слюнные железы
комиссура
брюшные
продольные
мышцы
пучок
нервных
волокон
дорсо-
вентральные
мышцы
нефридий
с
саккулюсом
ходильная
нога
коготки
преддверие
трахеи
кожная
папилла
с
чувствительной
щетинкой
•ИЙ- эпидермис
|щ_ базальный
матрикс
кольцевые
мышцы
косые мышцы
сагиттальные
мышцы
продольные
мышцы
пучок
трахеи
Илл.
577. Onychophora. Продольный разрез через покров
тела с головной папиллой (механорецептор) и пучком
трахей.
По Gaffron (1883) и Pflugfelder (1968).
Илл.
576. Onychophora. Схематичес-
кий поперечный разрез через середи-
ну туловища.
По Snodgrass (1938) и другим авторам, с из-
менениями.
anophthalmus
лишены глаз, так же как и пещерные виды
Peripatus alba и
Speleoperipatus
spelaeus.
Покровы снабжены большим количеством ре-
цепторов, некоторые из которых имеют сложное
строение, особенно те, что расположены на антен-
нах (илл.
581 Б),
в преддверии рта и на подошве ла-
пок. Тонкая структура рецепторов сходна с таковой
у Euarthropoda. Наиболее часто встречающийся тип
сенсилл находится в покровах спинной стороны. Эти
так называемые кожные папиллы снабжены чувстви-
тельными щетинками, которые чувствуют прикосно-
вение. На кольцевых бороздках кончиков антенн
встречаются колбовидные сенсиллы, ограниченные
нежной кутикулой, выпуклой, подобно часовому
стеклу. На утолщённых губах преддверия рта распо-
ложены конические чувствительные почки с дисталь-
но видоизменённой кутикулой — вероятно, они яв-
ляются хеморецепторами. Предполагается также на-
личие гигро- и терморецепторов. Кроме того, онихо-
форы весьма чувствительны к движению воздуха.
http://jurassic.ru/