Вестхайде В.,Ригер Р.( ред.) Зоология беспозвоночных в двух томах. Том 1: от простейших до моллюсков и артропод
Подождите немного. Документ загружается.

406
Annelida
A
Б
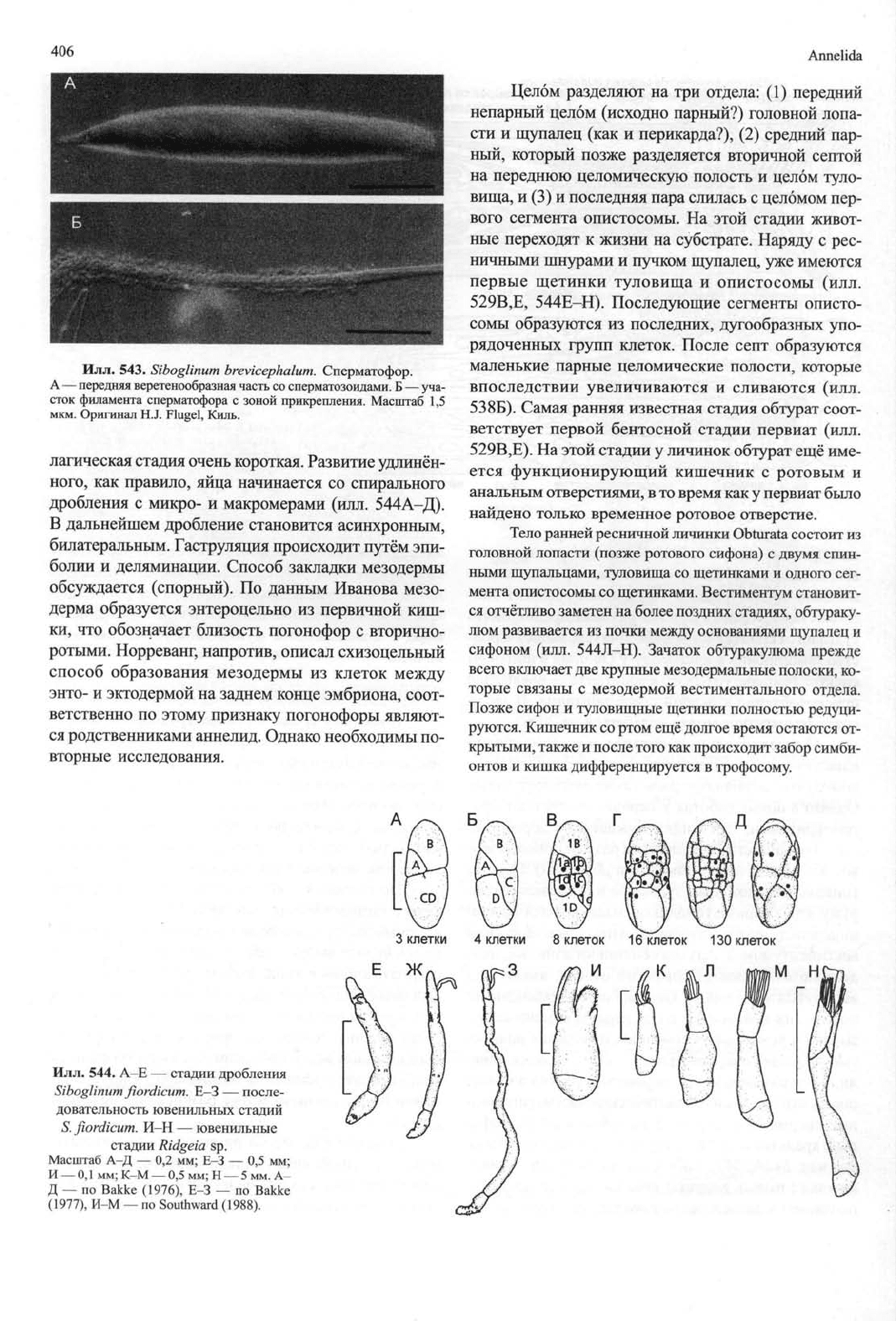
Илл.
543.
Siboglinum
brevicephalum.
Сперматофор.
A — передняя веретенообразная часть со сперматозоидами. Б — уча-
сток филамента сперматофора с зоной прикрепления. Масштаб 1,5
мкм.
Оригинал H.J. Flugel, Киль.
лагическая стадия очень короткая. Развитие удлинён-
ного,
как правило, яйца начинается со спирального
дробления с микро- и макромерами (илл. 544А-Д).
В дальнейшем дробление становится асинхронным,
билатеральным. Гаструляция происходит путём эпи-
болии и деляминации. Способ закладки мезодермы
обсуждается (спорный). По данным Иванова мезо-
дерма образуется энтероцельно из первичной киш-
ки,
что обозначает близость погонофор с вторично-
ротыми. Норреванг, напротив, описал схизоцельный
способ образования мезодермы из клеток между
энто-
и эктодермой на заднем конце эмбриона, соот-
ветственно по этому признаку погонофоры являют-
ся родственниками аннелид. Однако необходимы по-
вторные исследования.
Целом разделяют на три отдела: (1) передний
непарный целом (исходно парный?) головной лопа-
сти и щупалец (как и перикарда?), (2) средний пар-
ный, который позже разделяется вторичной септой
на переднюю целомическую полость и целом туло-
вища, и (3) и последняя пара слилась с целомом пер-
вого сегмента опистосомы. На этой стадии живот-
ные переходят к жизни на субстрате. Наряду с рес-
ничными шнурами и пучком щупалец, уже имеются
первые щетинки туловища и опистосомы (илл.
529В,Е, 544Е-Н). Последующие сегменты описто-
сомы образуются из последних, дугообразных упо-
рядоченных групп клеток. После септ образуются
маленькие парные целомические полости, которые
впоследствии увеличиваются и сливаются (илл.
538Б).
Самая ранняя известная стадия обтурат соот-
ветствует первой бентосной стадии первиат (илл.
529В,Е). На этой стадии у личинок обтурат ещё име-
ется функционирующий кишечник с ротовым и
анальным отверстиями,
в
то время как у первиат было
найдено только временное ротовое отверстие.
Тело ранней ресничной личинки Obturata состоит из
головной лопасти (позже ротового сифона) с двумя спин-
ными щупальцами, туловища со щетинками и одного сег-
мента опистосомы со щетинками. Вестиментум становит-
ся отчётливо заметен на более поздних стадиях, обтураку-
люм развивается из почки между основаниями щупалец и
сифоном (илл. 544JT-H). Зачаток обтуракулюма прежде
всего включает две крупные мезодермальные полоски, ко-
торые связаны с мезодермой вестиментального отдела.
Позже сифон и туловищные щетинки полностью редуци-
руются. Кишечник со ртом ещё долгое время остаются от-
крытыми, также
и
после
того
как происходит забор симби-
онтов и кишка дифференцируется в трофосому.
http://jurassic.ru/
Annelida
407
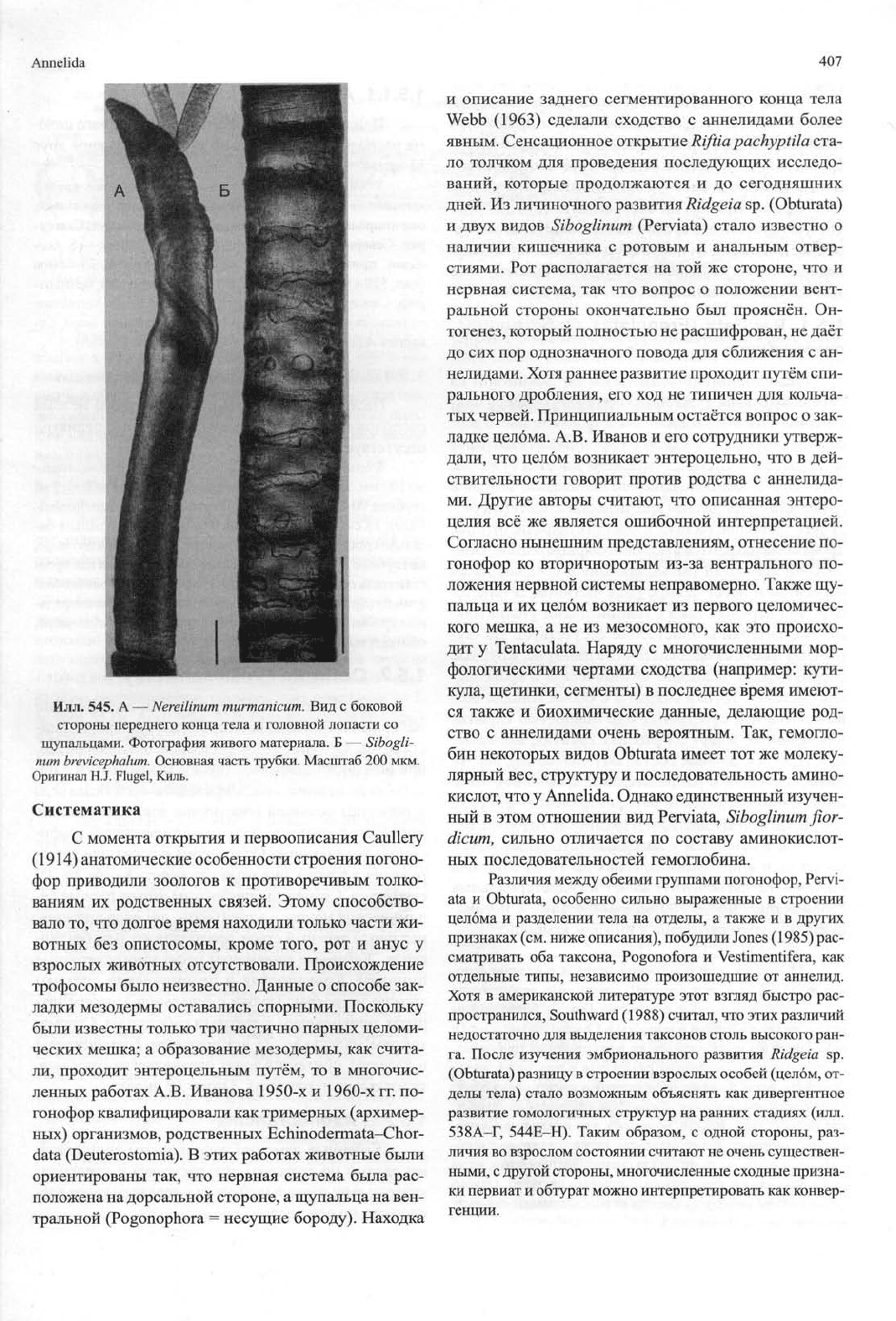
Илл.
545.
A —
Nereilinum
murmanicum.
Вид с боковой
стороны переднего конца тела и головной лопасти со
щупальцами. Фотография живого материала. Б —
Sibogli-
num
brevicephalum.
Основная часть трубки. Масштаб
200
мкм.
Оригинал H.J. Flugel, Киль.
Систематика
С момента открытия и первоописания Caullery
(1914) анатомические особенности строения погоно-
фор приводили зоологов к противоречивым толко-
ваниям их родственных связей. Этому способство-
вало то, что долгое время находили только части жи-
вотных без опистосомы, кроме того, рот и анус у
взрослых животных отсутствовали. Происхождение
трофосомы было неизвестно. Данные о способе зак-
ладки мезодермы оставались спорными. Поскольку
были известны только три частично парных целоми-
ческих мешка; а образование мезодермы, как счита-
ли,
проходит энтероцельным путём, то в многочис-
ленных работах А.В. Иванова 1950-х и 1960-х гг. по-
гонофор квалифицировали как тримерных (архимер-
ных) организмов, родственных Echinodermata-Chor-
data (Deuterostomia). В этих работах животные были
ориентированы так, что нервная система была рас-
положена на дорсальной стороне, а щупальца на вен-
тральной (Pogonophora = несущие бороду). Находка
и описание заднего сегментированного конца тела
Webb (1963) сделали сходство с аннелидами более
явным. Сенсационное открытие Riftia pachyptila ста-
ло толчком для проведения последующих исследо-
ваний, которые продолжаются и до сегодняшних
дней. Из личиночного развития Ridgeia sp. (Obturata)
и двух видов Siboglinum (Perviata) стало известно о
наличии кишечника с ротовым и анальным отвер-
стиями. Рот располагается на той же стороне, что и
нервная система, так что вопрос о положении вент-
ральной стороны окончательно был прояснён. Он-
тогенез, который полностью не расшифрован, не даёт
до сих пор однозначного повода для сближения с ан-
нелидами. Хотя раннее развитие проходит путём спи-
рального дробления, его ход не типичен для кольча-
тых червей. Принципиальным остаётся вопрос о зак-
ладке целома. А.В. Иванов и его сотрудники утверж-
дали, что целом возникает энтероцельно, что в дей-
ствительности говорит против родства с аннелида-
ми.
Другие авторы считают, что описанная энтеро-
целия всё же является ошибочной интерпретацией.
Согласно нынешним представлениям, отнесение по-
гонофор ко вторичноротым из-за вентрального по-
ложения нервной системы неправомерно. Также щу-
пальца и их целом возникает из первого целомичес-
кого мешка, а не из мезосомного, как это происхо-
дит у Tentaculata. Наряду с многочисленными мор-
фологическими чертами сходства (например: кути-
кула, щетинки, сегменты) в последнее время имеют-
ся также и биохимические данные, делающие род-
ство с аннелидами очень вероятным. Так, гемогло-
бин некоторых видов Obturata имеет тот же молеку-
лярный вес, структуру и последовательность амино-
кислот, что у Annelida. Однако единственный изучен-
ный в этом отношении вид Perviata, Siboglinum fior-
dicum,
сильно отличается по составу аминокислот-
ных последовательностей гемоглобина.
Различия между обеими группами погонофор, Pervi-
ata и Obturata, особенно сильно выраженные в строении
целома и разделении тела на отделы, а также и в других
признаках
(см.
ниже описания), побудили Jones (1985) рас-
сматривать оба таксона, Pogonofbra и Vestimentifera, как
отдельные типы, независимо произошедшие от аннелид.
Хотя в американской литературе этот взгляд быстро рас-
пространился, Southward (1988) считал, что этих различий
недостаточно для выделения таксонов столь высокого ран-
га. После изучения эмбрионального развития Ridgeia sp.
(Obturata) разницу
в
строении взрослых особей (целом, от-
делы тела) стало возможным объяснять как дивергентное
развитие гомологичных структур на ранних стадиях (илл.
538А-Г, 544Е-Н). Таким образом, с одной стороны, раз-
личия во взрослом состоянии считают
не
очень существен-
ными,
с
другой стороны, многочисленные сходные призна-
ки первиат
и
обтурат можно интерпретировать как конвер-
генции.
http://jurassic.ru/

408
Annelida
Вероятно, Perviata
и
Obturata всего лишь ука-
зывают
нам на
существование общего предкового
таксона. Даже без дополнительных исследований эм-
брионального развития сегодняшнее состояние
на-
ших знаний позволяет отнести погонофор
к
кольча-
тым червям. Причём
их
сестринскую группу, веро-
ятно,
следует искать среди сидячих «Polychaeta» (Sa-
bellida?), обитающих в трубках и несущих венчик щу-
палец.
На это
указывают структуры аналогичные
крючковидным щетинкам (илл. 493Ж, 533)*.
1.5.1. Perviata
(Frenulata),
Собственно
погонофоры
Тело состоит (номенклатура
по
Southward)
из
переднего отдела, туловища и сегментированной опи-
стосомы. Головная лопасть несёт одно или много дор-
сальных щупалец
и
уздечку (френулюм).
На
грани-
це головной лопасти
с
туловищем имеется сужение.
Туловище
с
многочисленными папиллами; поясок
с
двумя или более рядами крючковидных щетинок раз-
деляет туловище
на
пре-
и
постаннулярные отделы.
Трофосома находится в постаннулярном отделе. Опи-
стосома
как
орган рытья, состоит
в
среднем
из 20
сегментов
и
несёт четыре ряда щетинок.
В
описто-
соме
три
нервных тяжа
с
ганглиозными утолщения-
ми.
Мезентерий
в
опистосоме отсутствует.
На щу-
пальцах имеются одноклеточные пиннулы. Крупные
сперматофоры. Сложные целомическая
и
кровенос-
ная системы. Трубки открыты
с
обоих концов, воз-
вышаются
над
субстратом
или
полностью скрыты.
Как правило, живут
на
рыхлых грунтах, насыщен-
ных
H
2
S
(исключение составляет только
вид
Sclero-
linum,
живущий
в
разлагающейся древесине). Име-
ют космополитическое распространение: фьорды,
континентальные шельфы
и
океанские глубины
25-
10 700
м; в
основном
в
холодных глубинных водах,
но также
и в
прибрежных водах
до
18°С; один
вид
(Siphonobrachia lauensis) живёт
в
гидротермальных
сообществах. Более
100
видов. Сходные
с
погоно-
форами трубки отмечены
в
нижнем кембрии.
* Сходства лишь в строении щетинок
и
наличие рото-
вых щупалец
(не
всегда венчика!) совершенно недостаточно
для сближения погонофор
с
Sabellida. Гораздо более сходны
крючковидные щетинки погонофор
и
Chaetopteridae, много
у
них общего и
в
других признаках,
в
частности их трубки час-
то практически невозможно различить:
у
некоторых видов
Spiochaetopterus
полупрозрачные трубки имеют правильную
кольчатость, как
и
трубки погонофор, например изображен-
ная
на
илл. 532В. Вполне возможно,
что
некоторые Chaeto-
pteridae, например
Spiochaetopterus,
питаются сходно
с
пого-
нофорами,
образуя
с ними
иногда смешанные
поселения,
виды
этого рода имеют пару длинных ротовых щупалец. Однако
Chaetopteridae почему-то никогда не рассматривают как сест-
ринскую группу
(прим.
ред.).
1.5.1.1.
Athecanephria
Целомодукты переднего мешкообразного целб-
ма расположены латерально
и
сильно удалены друг
от друга.
Siboglinum
ekmani
(Siboglinidae),
40-100
мм;
как и
у
всех видов данного рода имеется только одно щупальце,
оно спирально закручено и несёт два ряда
пиннул;
Скагер-
рак, Северная Атлантика, глубина 300-1
250 м. —
S.
fior-
dicum,
примерно
80 мм;
Скагеррак, Норвегия, 25-560
м
(илл.
529А).
—
S.
poseidoni,
64
мм, гермафродит, метилот-
роф;
Скагеррак, обитает
на
глубине
300 м. —
Nereilinum
murmanicum, 105
мм;
два
щупальца, Баренцево море,
Се-
верная Атлантика; глубина 170-1
300 м
(илл. 545А).
1.5.1.2.
Thecanephria
Целомодукты переднего U-образного целбма
расположены близко
к
медианной линии, перикард
отсутствует.
Sclerolinum
brattstroemi
(Sclerolinidae),
приблизитель-
но 100 мм, два щупальца без пиннул; Норвегия, обитает на
глубине 90-1 300
м.
—
Lamellisabella zachsi
(Lamellisabel-
lidae),
18
см; 29-31 щупальце
с
пиннулами, щупальца
ба-
зально сросшиеся; 220 мм, Охотское море, Берингово море,
на глубине
2
900-3 500 м, впервые была описана как пред-
ставитель семейства полихет (!) Sabellidae.
—Spirobrachia
grandis
(Spirobrachiidae), фрагменты до 80
мм;
на спираль-
ном гребне
до
225 щупалец
с
пиннулами; Берингово море,
обитают на глубине
3
250
м.
1.5.2.
Obturata (Vestimentifera)
Тело состоит
из
обтуракулюма, щупалец
или
жаберных пластинок, вестиментума, туловища и опи-
стосомы. Обтуракулюм расширен бокалообразно,
при опасности закрывает трубку. Вестиментум
с бо-
ковыми складками, они покрывают второй отдел тела
и образуют большой воротничок вокруг основания
жаберных пластинок. Туловище
с
массивной трофо-
сомой,
без
щетинок. Опистосома служит
для
заяко-
ривания
в
трубке,
она
может включать
в
себя
до 100
сегментов. Сегменты опистосомы имеют парные це-
ломические мешки
и
мезентерии; два ряда щетинок,
непарный нервный ствол, предположительно без ган-
глиев. Жаберные филаменты несут многоклеточные
пиннулы. Кровеносная
и
целомические системы
сложно устроены. Трубки
с
базальной стороны зак-
рыты, прикреплены
к
твёрдым субстратам. Сперма-
тофоры маленькие.
За
исключением Lamellibrachia
и Escarpia, обитают
в
зонах океанических разломов,
термальных источниках. Около
10
видов.
1.5.2.1.
Axonobranchia
Жаберные ламеллы расположены восходящи-
ми рядами
по
сторонам обтуракулюма
и
перпенди-
кулярно ему; дорсальный сосуд
в
переднем конце рас-
положен
по оси
тела.
http://jurassic.ru/

Annelida
409
мозг
фронтальные
железы
мужская
паренхима
пора
семенник яичник кишка
семяприёмник
задняя
кишка
анус
преоральныи
ганглий
Илл. 546. Lobatocerebrida.
Вид сбоку, Lobatocerebrumpsammicola. Длина: 3 мм. По Rieger (1980).
Только
Riftia pachyptila
1,5 м длиной, диаметр 4 см;
обитает в Тихом океане: Галапагосский рифт, восточная
возвышенность Тихого океана, глубина обитания 2 500 м;
максимальное число жаберных ламелл достигает 334 пар;
это самый большой вид среди погонофор и самый боль-
шой вид среди глубоководной фауны термальных источ-
ников (илл. 529Г).
1.5.2.2.
Basibranchia
Жаберные ламеллы расположены рядами вок-
руг основания обтуракулюма и ориентированы па-
раллельно; в переднем конце тела спинной сосуд рас-
положен базально.
Lamellibrachia barhami
(Lamellibrachiidae), 48 см в
длину и 5 мм в диаметре; обитают в холодных сипах, со-
держащих сульфиды, углеводороды и водород, на конти-
нентальных цоколях; Западная Пацифика, на глубине
1 100-2 000 м; 25 пар жаберных филаментов и две-четы-
ре пары периферических покрывающих пластинок. — L.
luymesi,
55 см; Атлантика. —Ridgeiapiscesae (Ridgeiidae),
65 см; Тихий океан.
1.6.
Lobatocerebrida
Небольшие животные, обитающие на морских
песчаных или заиленных грунтах. Внешне похожи
на турбеллярий (полностью покрыты ресничками,
целом отсутствует) (илл. 546). Имеется большой ло-
пастной мозг. Гермафродиты. Филогенетический ана-
лиз признаков показал, что Lobatocerebrida, вероят-
но,
произошли от каких-то ювенильных аннелид, в
условиях интерстициали и в результате прогенеза.
Мужские половые канальцы схожи с целомодукта-
ми,
они открываются ресничной воронкой в мешко-
видные гонады. Протонефридий парные, открывают-
ся отдельно, имеются задняя кишка и анус, рудимен-
тарная кровеносная система. Как у аннелид и дру-
гих целомических Spiralia, тело покрыто кутикулой.
Ультраструктура сперматозоидов указывает на воз-
можную близость к малощетинковым червям.
Описан только один вид: Lobatocerebrum psammicola
(Lobatocerebridae), 4 мм; животные медленно скользят по
поверхности при помощи ресничек, также как мелкие ин-
терстициальные брюхоногие моллюски. В верхней субли-
торали
(1-3 м) в
окрестностях морской биологической стан-
ции в Эйлате, Израиль, был найден ещё один близкий вид.
2. Clitellata,
Поясковые
кольчецы*
Поясковые аннелиды — более чем 8 000 ви-
дов — преимущественно пресноводные или назем-
ные организмы; морские виды живут прежде всего
на литорали. Дождевые черви являются самой изве-
стной группой среди червеобразных беспозвоноч-
ных. Наряду с большинством мелких водных видов
миллиметрового размера, имеются многочисленные
наземные формы, размеры которых варьируют от не-
больших до более 1 метра (несколько видов) в дли-
ну. Преимущественно эктопаразитами являются Hiru-
dinea; к ним относятся так называемые медицинс-
кие пиявки, которые сосут кровь также и у людей.
Среди них находится Hirudo medicinalis, которая дол-
гое время в Европе имела исключительное значение
для донаучной медицины.
* Wilfried Westheide, Оснабрюк.
щетинка
железа
Илл. 547. Половые органы олигохет.
Плезиопорное положение у Tubificidae: органы расположены только
в двух сегментах. Вид сбоку. Рисунок изменён, по Erseus (1980).
http://jurassic.ru/

410
Annelida
Структура, которая дала название группе, —
клителлюм (поясок) — отчётливо выраженное воз-
вышение участка эпидермиса, пронизанное много-
численными железами (илл. 547, 550, 553А). Поясок
включает несколько расположенных друг за другом
сегментов передней части тела и выглядит как коль-
цевой валик или седло. Поясок есть у всех без ис-
ключения Clitellata. Его функции (см. ниже) опреде-
ляют особенности биологии размножения, органи-
зацию половых органов и внешний облик этих анне-
лид.
Строение
Форма тела поясковых червей довольно едино-
образна и определяется отчётливой гомономной сег-
ментацией. Параподии отсутствуют, щетинки про-
стые — по большей части они короткие, волосовид-
ные или в виде штырьков (илл. 500Б); некоторые так-
соны щетинок не имеют, например род Achaeta (Еп-
chytraeidae), Branchiobdellida и все Euhirudinea.
Простомиум чаще всего редуцирован до ма-
ленькой лопасти без придатков или простомиум вовсе
отсутствует (Branchiobdellida, Acanthobdellida). Мозг
из-за маленького размера простомиума смещён на-
зад в перистомиум и в последующие сегменты. Ган-
глии передних метамеров образуют вентральный
подглоточный ганглий (илл. 496Б). У олигохет нерв-
ный тяж без отчётливых ганглиев. Напротив, у пия-
вок тела клеток нейронов объединяются по шесть и
формируют чётко выраженную цепочку ганглиев
(илл. 496В, 556Б, 557). Нухальные органы отсутству-
ют. Простые глаза в виде пигментных бокалов-оцел-
лей; они только иногда имеются на простомиуме.
Глазки олигохет нельзя гомологизировать с глазами
полихет. Часто встречаются одиночные фоторецеп-
торы, расположенные в эпидермисе. Фоторецепто-
ры эпидермиса имеют центральную вакуоль, окайм-
лённую микровиллями, совершенно так же как со-
ответствующие клетки в глазах (фаосомы, Phaosom)
(илл. 559).
Олигохеты имеют маленький пигидиум, чаще
всего без придатков, несущий анальное отверстие. У
Hirudinea пигидиум вычленить невозможно, т.к. он
входит в состав каудальной присоски.
В качестве органов выделения имеются ме-
танефридии, которые у пиявок сильно видоизмене-
ны.
Все Clitellata — синхронные гермафродиты.
Положение их половых органов ограничено только
несколькими сегментами. Впереди расположены два
сегмента с семенниками, далее следуют два сегмен-
та с яичниками, в каждом по паре гонад (октогонад-
ный вариант, например у Haplotaxidae).
У пиявок парные семенные пузырьки в несколь-
ких сегментах (илл. 558Б) — это вторичное преоб-
разование: исходно единственная пара семенных
мешков находится в 10-м сегменте, как это демонст-
рирует Acanthobdella (илл. 558А).
У «01igochaeta» гаметы попадают наружу че-
рез парные ресничные воронки и присоединенные
к ним гонодукты (семяпроводы или яйцеводы). По-
ложение их наружных половых отверстий является
очень важным таксономическим признаком. У
Tubificida (илл. 547) и Haplotaxida мужские отверстия
(у Tubificidae могут быть пенисы) расположены на
следующем метамере от сегмента с семенным пу-
зырьком (плезиопорный вариант). У Lumbriculida
семяпроводы открываются в том же сегменте, где рас-
пложены семенные пузырьки (прозопорный вари-
ант).
У Lumbricida семенные поры отнесены далеко
назад (опистопорный вариант) (илл. 551). У Hiru-
dinea мужские половые выводящие канальцы перед
открытием наружу объединяются в одну непарную
пору. Эта пора, как правило, расположена на вент-
ральной стороне IX сегмента. Таким же образом не-
парное женское половое отверстие располагается на
X сегменте. Только у пиявок семенные пузырьки вто-
рично смещены назад, за яичники (илл. 558). К жен-
ским половым органам олигохет относятся одна или
более пара сперматек (receptacula seminis) (илл. 547,
551).
Сперматеки — эпидермальные мешкообразные
Илл. 548. Откладывание кокона у Clitellata.
Erpobdella octoculata
(Hirudinea).
A — клителлюм из пояска с гладкой слизевой муф-
той (тёмный отдел); под ним расположены запаса-
ющая (питательная) жидкость и яйца. Б-Д — пи-
явка стягивает муфту и секрет через переднюю
часть тела, после чего муфта в виде кокона будет
прикреплена к субстрату. Е — дополнительное
формирование кокона при помощи ротовой при-
соски. Ж — законченный кокон, длина 3-6 мм;
плоская нижняя сторона крепко прикрепляется,
верхняя сторона плоскосферическая. Яйца бедны
желтком, плавают в питательной жидкости. Раз-
витие проходит через особую стадию вторичной
личинки (илл. 549 3). По Westheide (1980).
http://jurassic.ru/
Annelida
411
эктотел
облает
мезотелобласт
средняя
желточный кишка
эктодермальный
М
1
меш01
<
полоска
эктотел
oблacтoв
,
желточный
эктодермальный
мешок
эктотел
облает
м
1
средняя
кишка
эктотелобласт
зародышевая
пластинка
целомические мешочки
брюшная нервная цепочка
личиночный
эпидермис
личиночные
органы
выделения
эктотелобласт
мезотелобласт
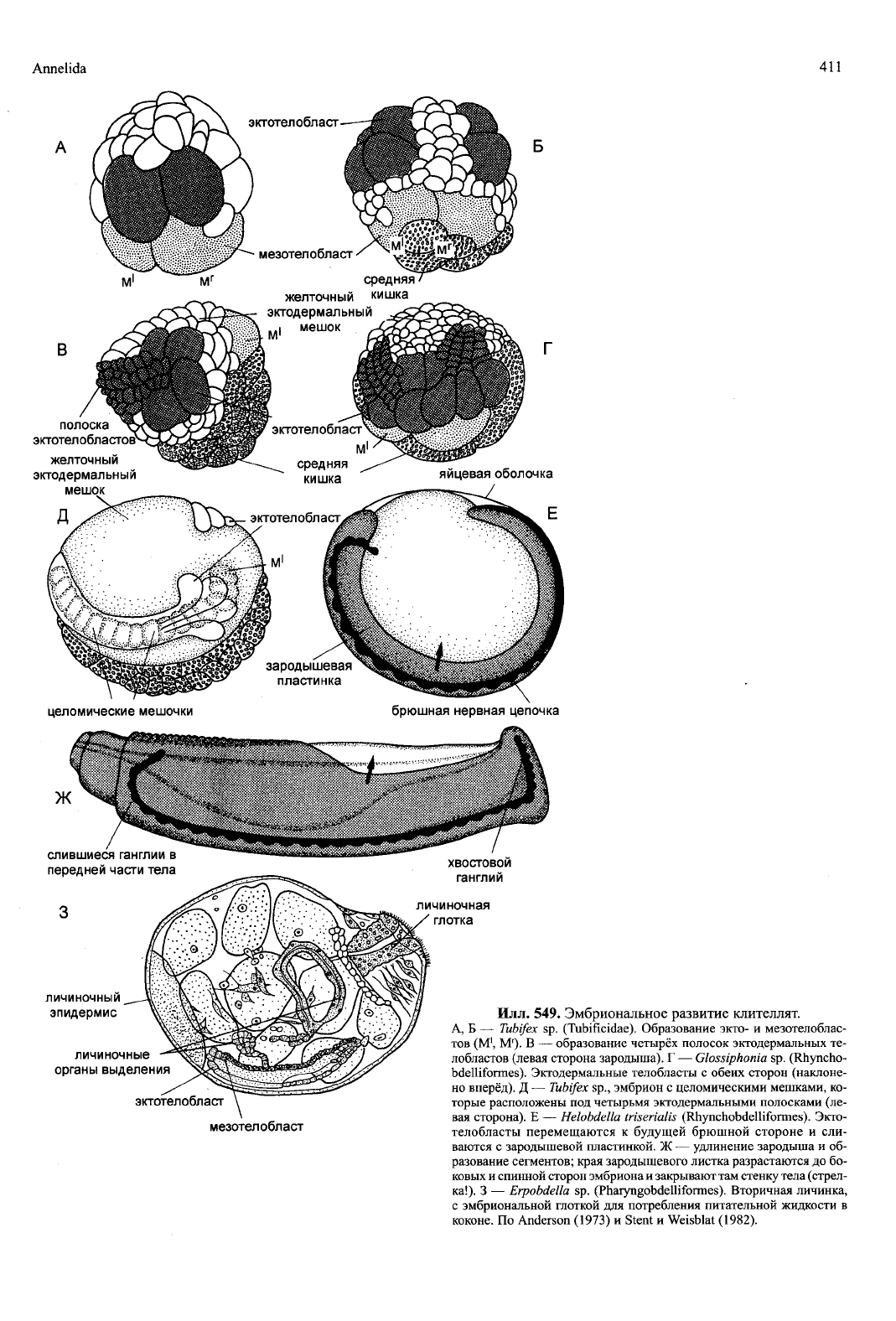
Илл. 549. Эмбриональное развитие клителлят.
А, Б — Tubifex sp. (Tubificidae). Образование экто- и мезотелоблас-
тов (М
1
, М
г
). В — образование четырёх полосок эктодермальных те-
лобластов (левая сторона зародыша). Г — Glossiphonia sp. (Rhyncho-
bdelliformes). Эктодермальные телобласты с обеих сторон (наклоне-
но вперёд). Д — Tubifex sp., эмбрион с целомическими мешками, ко-
торые расположены под четырьмя эктодермальными полосками (ле-
вая сторона). Е — Helobdella triserialis (Rhynchobdelliformes). Экто-
телобласты перемещаются к будущей брюшной стороне и сли-
ваются с зародышевой пластинкой. Ж — удлинение зародыша и об-
разование сегментов; края зародышевого листка разрастаются до бо-
ковых и спинной сторон эмбриона и закрывают там стенку тела (стрел-
ка!).
3 — Erpobdella sp. (Pharyngobdelliformes). Вторичная личинка,
с эмбриональной глоткой для потребления питательной жидкости в
коконе. По Anderson (1973) и Stent и Weisblat (1982).
http://jurassic.ru/

412
Annelida
выросты, которые служат для забора спермы другой
особи. Сперматеки первично отделены от полости
тела. Только у мелких олигохет избыточное количе-
ство спермы может поступать через канальцы в ки-
шечник.
Размножение и развитие
У олигохет при копуляции две особи прижима-
ются брюшными сторонами друг к другу в противо-
положных направлениях таким образом, чтобы по-
ясок одного животного соответствовал сперматеке
другого. В результате каждая сперматека наполняет-
ся спермой другой особи (илл. 551).
У пиявок отсутствуют такого рода накопитель-
ные органы: сперматозоиды передаются (инъециру-
ются) партнёру посредством сперматофоров (илл.
562) или переносятся при помощи пениса
(илл.
556А)
в женскую пору. Таким образом осуществляется
внутреннее оплодотворение.
Строение и функция пояска таковы, что отвер-
стия сперматек и женские половые отверстия (отвер-
стия яйцеводов) находятся на пояске перед ним. При
размножении клителлюм отделяет слизистую муфту
с жидкостью под ней. Далее животное вытягивает
передний конец из этой муфты, в неё откладывают-
ся яйца (илл. 548); муфта превращается в защитный
кокон. У олигохет в муфте проходят наружное опло-
дотворение и полный цикл эмбрионального разви-
тия.
Эпидермис пояска, например у Lumbricidae, в три-
четыре раза выше, чем в других частях тела. Он содержит
(1) наряду
с
обычными эпидермальными клетками особен-
но много железистых клеток; (2) слизеотделяющие клет-
ки,
дающие тонкий матрикс для кокона; (3) клетки, секре-
тирующие стенку
кокона,
и (4)
обычно несколько типов же-
лезистых клеток, которые выделяют секрет для жидкости,
в которой плавают яйца. У видов с бедными желтком яй-
цами этот секрет содержит много протеинов и липидов. В
основании пояска располагается
хорошо
развитый нервный
плексус.
Ход развития — характерная аутапоморфия так-
сона поясковых кольчецов. Раннее развитие изначаль-
но богатых желтком яиц происходит по типу спираль-
ного дробления, однако сильно модифицированно-
го.
У Tubifex («01igochaeta») и Glossiphonia (Eu-
hirudinea) яйца снабжены желтком. У них закладка
клеток презумптивной средней кишки происходит
путём эпиболии анимальных микромеров внутрь
эмбриона (илл. 549). Из бластомера 2d образуются
восемь больших эктотелобластов. Из них на каж-
дой стороне зародыша формируются по четыре по-
лоски, состоящие, в свою очередь, из четырёх рядов
клеток (илл. 549В). Вместе с шапочкой из микроме-
ров (провизорная или желточная эктодерома) полос-
ки клеток формируют, прежде всего, эктодермальную
оболочку зародыша. В дальнейшем они срастаются
друг с другом и объединяются в одну вентральную
вогнутую зародышевую пластинку, которая, разрас-
таясь, охватывает весь эмбрион (илл. 549Е). По мере
растяжения зародыша дифференцируются ганглии
нервной системы, эпителий поверхности тела и ще-
тинковые мешки. Из
4d-mieTOK
возникают мезоте-
лобласты (М
1
, М
г
); они формируют две мезодермаль-
ные полоски клеток внутри зародыша. Из мезодер-
мальных клеток, как у полихет, отпочковываются ти-
пичные попарно упорядоченные целомические меш-
ки.
Ювенильные животные покидают кокон, имея
внешний вид взрослых червей.
У
таксонов
с
вторично бедными желтком
яйцами,
на-
пример
Lumbricus
(«OHgochaeta»),
Piscicola
(Euhirudinea),
внутри кокона развивается личинка (не гомологичная тро-
хофоре!). Эта личинка имеет эмбриональную глотку, при
помощи которой потребляет питательную жидкость (илл.
549 3).
Систематика
Монофилия клителлят уверенно подтверждает-
ся многочисленными признаками (поясок и образо-
вание кокона, специфический онтогенез, смещение
мозга из простомиума, гермафродитизм, размещение
гонад в ограниченном числе сегментов) (илл. 504,
547).
Мы будем придерживаться общеупотребитель-
ного подразделения на «OHgochaeta» и Hirudinea.
Однако для таксона «01igochaeta» неизвестно ника-
ких синапоморфий. Малощетинковые черви образу-
ют парафилетическую группу. Какие олигохеты яв-
ляются сестринской группой пиявок? Этот вопрос
открыт, но чаще такой группой считают Lumbriculida.
2.1.
«Oligochaeta»,
Малощетинковые черви
Подавляющее большинство олигохет — пре-
сноводные или почвенные обитатели. Морские виды
встречаются только среди очень мелких («микродри-
лидных») форм, прежде всего у Enchytraeidae и Ти-
bificidae, которые поселяются в основном на лито-
рали. В то время как водные таксоны распростране-
ны по всему миру, многие наземные группы олиго-
хет, к которым относятся самые крупные («мегадри-
лидные») виды, имеют географически ограниченные
ареалы, или широко распространены с помощью че-
ловека.
Множество пресноводно-наземных видов живут за
счёт субстрата,
обогащенного органическими веществами,
детритом, грибами, диатомеями, бактериями и другими
микроорганизмами. Многие виды питаются к тому же и
частями растений на определённых этапах распада (сап-
http://jurassic.ru/

Annelida
413
Clitellata
Илл. 550.
Родственные отношения внутри Clitellata.
Апоморфии:
[1]
Поясок
и
образование кокона, особенности онтоге-
неза, смещение мозга
из
простомиума, гермафродитизм, размещение
гонад
в
ограниченном числе сегментов
и т.д. [?]
Аутапоморфии
«01igochaeta» неизвестны, вероятно группа является парафилитичес-
кой.
[2]
Число сегментов постоянно
(32,
плюс простомиум
и
перис-
томиум), щетинки только
у
пяти первых сегментов, задняя присоска
состоит
из
семи сегментов, пигидиум утрачен, особые мышечные
клетки, паренхимная соединительная ткань, редукция мезентериев
и
целомических мешков, особое строение нефридиев, слияние мужс-
ких
и
женских половых отверстий
в
одну пору, утрата сперматек
и
внутреннего оплодотворения
и
т.д..[3] Постоянное число внешне
оп-
ределяемых сегментов равно
29,
задняя присоска состоит только
из
четырёх сегментов
и
т.
д. [4]
Утрата всех щетинок; передняя присос-
ка включает
в
себя четыре сегмента, целом редуцирован
до
системы
канальцев, семенные мешки подразделены особым образом.
По Pur-
schke, Westheide, Rohde
и
Brinkhurst (1993).
робионты). Таким образом, олигохеты играют важную роль
в разложении высших растений. Только некоторые виды
являются фильтраторами, например
Ripistes parasita
(Nai-
didae), хищниками, например виды рода
Chaetogaster
(Nai-
didae), «падалыциками», например некоторые Enchytrae-
idae,
а
также эктопаразитами, например немногочислен-
ные Branchiobdellida.
Строение
Для олигохет характерен единообразный вне-
шний
вид
(габитус): хорошо выраженные, чётко
от-
делённые друг
от
друга, одинаковые, округлые сег-
менты
без
придатков. Число щетинок обычно неве-
лико (отсюда название!) (например,
у
Lumbricus
ter-
restris
—
восемь
на
сегмент). Форма щетинок про-
стая
—
преимущественно
это
длинные капиллярные
волосовидные щетинки
или
простые шиловидные
(илл.
500Б), которые только немного выдаются
из
тела.
Первоначальным состоянием
является наличие
по
одной паре латеро-дорсальных
и
латеро-вентральных
ще-
тинковых мешков
на
сегмент
(илл.
500Б), движение кото-
рых осуществляется
за
счёт мышц. Однако
у
многих
Megascolecidae каждый сегмент окружён практически пол-
ным кольцом
из
щетинок,
их
может быть
от 50
до
100.
По-
ловые щетинки
имеются
у
представителей различных
се-
мейств (илл. 493Г). Щетинки могут отсутствовать, напри-
мер,
у
рода
Achaeta
(Enchytraeidae)
и у
Branchiobdellida.
Внутренняя сегментация также чёткая
и
регу-
лярная.
Мускулистые, мощные, полные диссепимен-
ты являются важным условием
для
роющего образа
жизни
и,
прежде всего,
для
наземных форм. Также
для роющего образа жизни важно иметь мускульный,
заострённый,
не
имеющий придатков передний
ко-
нец тела
и
мощно развитый кожно-мускульный
ме-
шок
(илл.
500Б).
Для
продвижения вперёд
у
Lumbricidae
сокраща-
ются сначала кольцевые мышцы (илл. 552). Благодаря чему
создаётся сильное давление целомической жидкости,
из-
за чего тело становится твёрдым
и
продвигается вперёд
со
втянутыми щетинками. Далее,
при
сокращении продоль-
ных мышц этот участок
с
выставленными наружу щетин-
ками сильно утолщается
и
закрепляется
в
грунте. Таким
образом, утоныпение тела
со
сжатыми кольцевыми мыш-
цами и утолщения
с
сокращёнными продольными мышца-
ми проходят вдоль всего тела спереди назад («ретроград-
но») перистальтическими волнами. Каждый сегмент в
оп-
ределенный момент служит
для
закрепления
в
грунте
вы-
тянутого участка тела. Благодаря полным диссепиментам,
целомическая полость каждого метамера является мощным
независимым гидравлическим компартментом (отделом).
Заострённый передний конец тела при рытье сначала про-
двигается
в
имеющееся пространство между частицами
грунта, после чего
это
пространство увеличивается благо-
даря расширению первых сегментов.
Система парафилетических «OUgochaeta» явля-
ется спорной. Названные здесь таксоны включают
в
себя намного больше известных видов,
чем
перечис-
лено.
2.1.1.
Haplotaxida
Очень мелкие, часто обитают
в
грунтовых
во-
дах,
в
древних водоёмах ледникового происхожде-
ния.
Географическое распространение дискретно,
много эндемичных видов. Положение
пор
прозопор-
ное;
чаще всего имеется
по два
сегмента
с
семенни-
ками
и
яичниками (октогонадные). Вероятно пара-
филетические.
*Haplotaxis gordioides
(Haplotaxidae),
180^100
мм,
тонкие нитевидные. Обитают прежде всего
в
континенталь-
ных грунтовых водах.
http://jurassic.ru/
414
Annelida
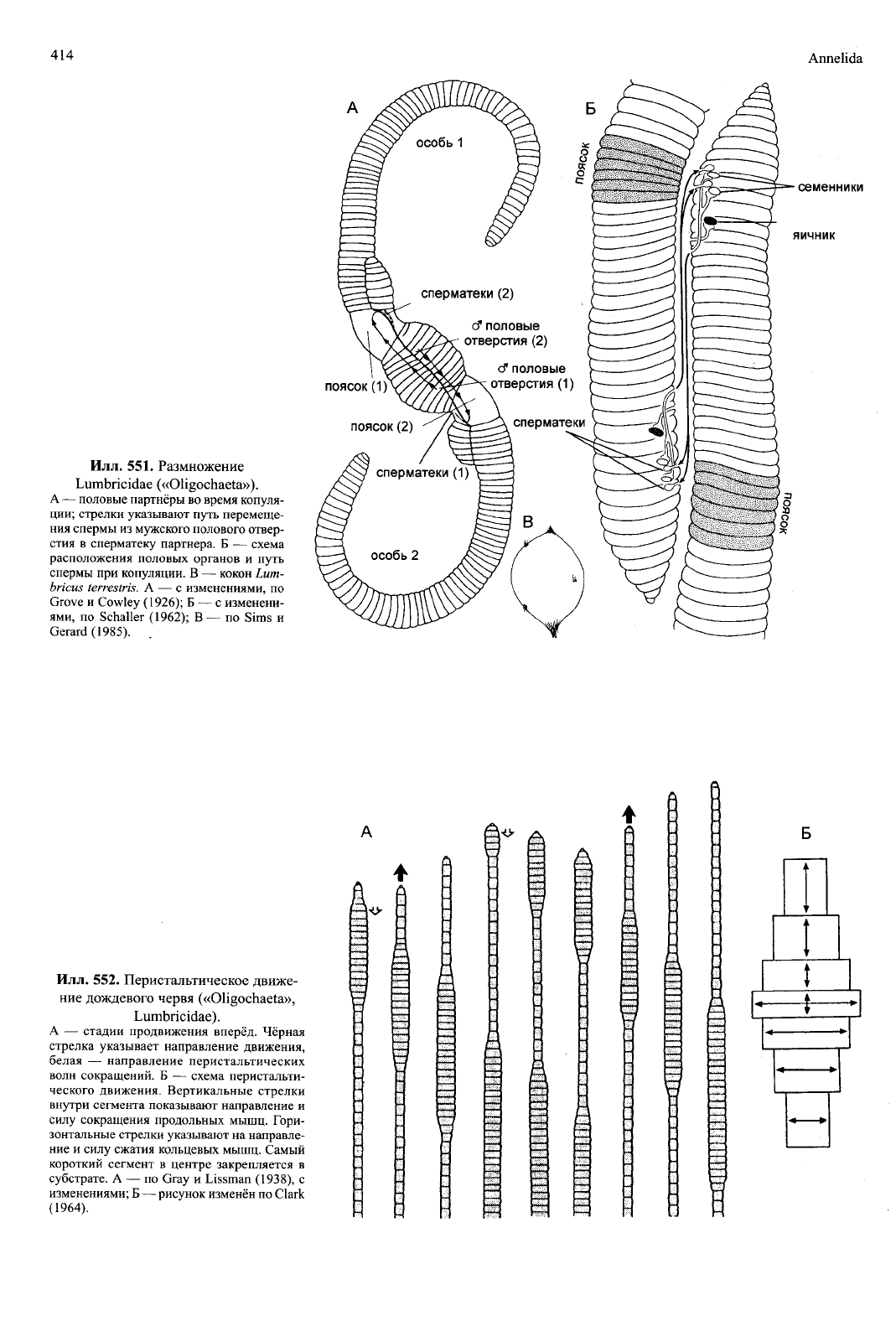
Илл.
551. Размножение
Lumbricidae («OHgochaeta»).
A — половые партнёры во время копуля-
ции;
стрелки указывают путь перемеще-
ния спермы из мужского полового отвер-
стия в сперматеку партнера. Б — схема
расположения половых органов и путь
спермы при копуляции. В — кокон Lum-
bricus terrestris. А — с изменениями, по
Grove и Cowley (1926); Б — с изменени-
ями,
по Schaller (1962); В — по Sims и
Gerard (1985).
семенники
Илл.
552. Перистальтическое движе-
ние дождевого червя («01igochaeta»,
Lumbricidae).
А — стадии продвижения вперёд. Чёрная
стрелка указывает направление движения,
белая — направление перистальтических
волн сокращений. Б — схема перистальти-
ческого движения. Вертикальные стрелки
внутри сегмента показывают направление и
силу сокращения продольных мышц. Гори-
зонтальные стрелки указывают на направле-
ние и силу сжатия кольцевых мышц. Самый
короткий сегмент в центре закрепляется в
субстрате. А — по Gray и Lissman (1938), с
изменениями; Б — рисунок изменён по Clark
(1964).
Л
*
Д
I л
А
Д
http://jurassic.ru/
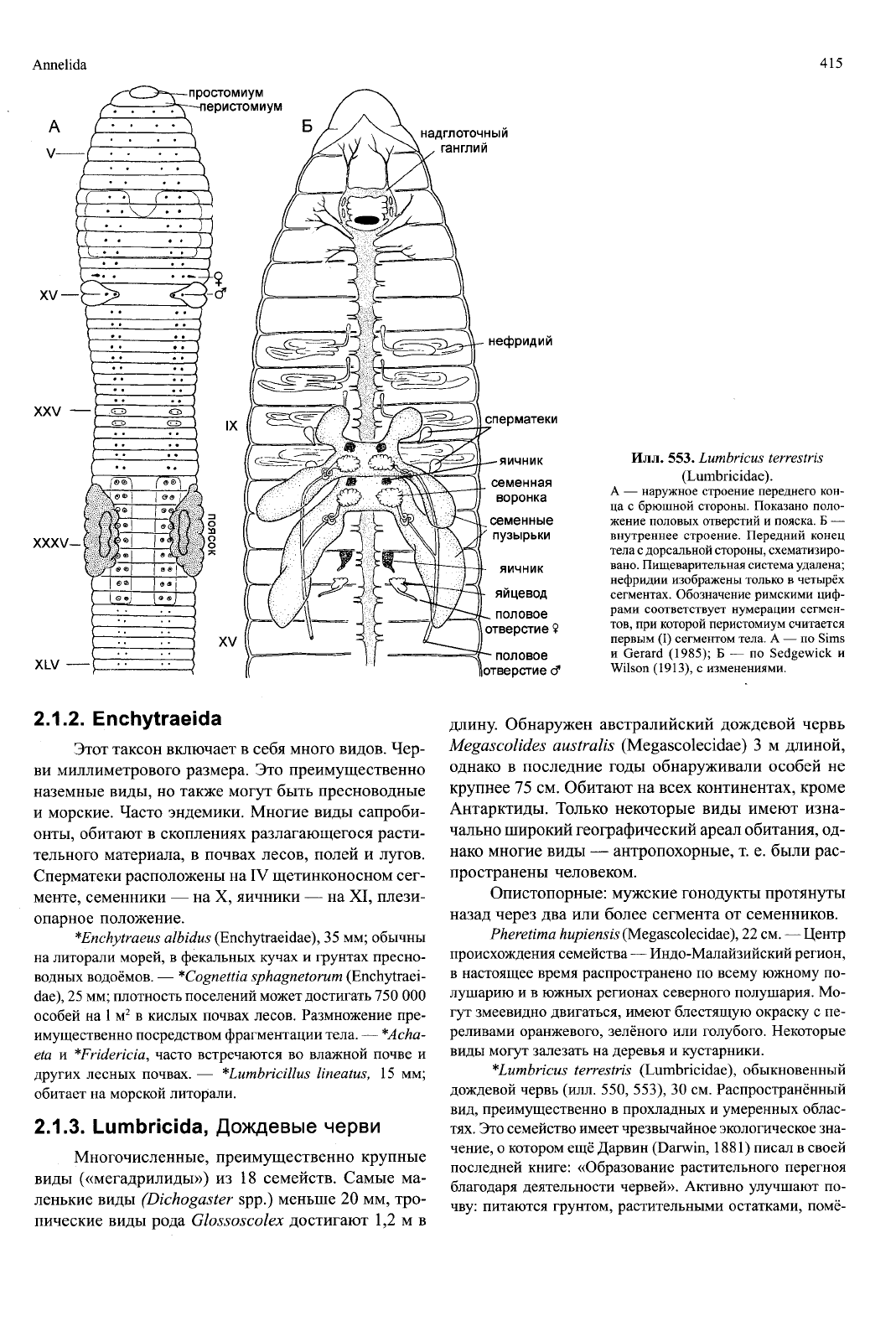
Annelida
415
-простомиум
r—перистомиум
A
V-
надглоточныи
ганглий
XV —
нефридий
XXV
XXXV-
XLV
половое
отверстие
d
1
Илл. 553. Lumbricus terrestris
(Lumbricidae).
А — наружное строение переднего кон-
ца с брюшной стороны. Показано поло-
жение половых отверстий и пояска. Б —
внутреннее строение. Передний конец
тела с дорсальной стороны, схематизиро-
вано.
Пищеварительная система удалена;
нефридии изображены только в четырёх
сегментах. Обозначение римскими циф-
рами соответствует нумерации сегмен-
тов,
при которой перистомиум считается
первым (I) сегментом тела. А — по Sims
и Gerard (1985); Б — по Sedgewick и
Wilson (1913), с изменениями.
2.1.2.
Enchytraeida
Этот таксон включает в себя много видов. Чер-
ви миллиметрового размера. Это преимущественно
наземные виды, но также могут быть пресноводные
и морские. Часто эндемики. Многие виды сапроби-
онты,
обитают в скоплениях разлагающегося расти-
тельного материала, в почвах лесов, полей и лугов.
Сперматеки расположены на IV щетинконосном сег-
менте,
семенники — на X, яичники — на XI, плези-
опарное положение.
*Enchytraeus albidus
(Enchytraeidae),
35
мм;
обычны
на литорали морей, в фекальных кучах и грунтах пресно-
водных водоёмов. —
*Cognettia sphagnetorum
(Enchytraei-
dae),
25 мм; плотность поселений может достигать 750
ООО
особей на
1
м
2
в кислых почвах лесов. Размножение пре-
имущественно посредством фрагментации тела. —
*Acha-
eta и
*Fridericia,
часто встречаются во влажной почве и
других лесных почвах. —
*Lumbricillus
lineatus,
15 мм;
обитает на морской литорали.
2.1.3.
Lumbricida,
Дождевые черви
Многочисленные, преимущественно крупные
виды («мегадрилиды») из 18 семейств. Самые ма-
ленькие виды (Dichogaster spp.) меньше 20 мм, тро-
пические виды рода Glossoscolex достигают 1,2 м в
длину. Обнаружен австралийский дождевой червь
Megascolides australis (Megascolecidae) 3 м длиной,
однако в последние годы обнаруживали особей не
крупнее 75 см. Обитают на всех континентах, кроме
Антарктиды. Только некоторые виды имеют изна-
чально широкий географический ареал обитания, од-
нако многие виды — антропохорные, т. е. были рас-
пространены человеком.
Опистопорные: мужские гонодукты протянуты
назад через два или более сегмента от семенников.
Pheretima hupiensis
(Megascolecidae),
22
см.
—
Центр
происхождения семейства — Индо-Малайзийский регион,
в настоящее время распространено по всему южному по-
лушарию и в южных регионах северного полушария. Мо-
гут змеевидно двигаться, имеют блестящую окраску с пе-
реливами оранжевого, зелёного или голубого. Некоторые
виды могут залезать на деревья и кустарники.
*Lumbricus terrestris
(Lumbricidae),
обыкновенный
дождевой червь (илл. 550, 553), 30 см. Распространённый
вид,
преимущественно в прохладных и умеренных облас-
тях. Это семейство имеет чрезвычайное экологическое зна-
чение, о котором ещё Дарвин (Darwin, 1881) писал в своей
последней книге: «Образование растительного перегноя
благодаря деятельности червей». Активно улучшают по-
чву: питаются грунтом, растительными остатками, помё-
http://jurassic.ru/