Вестхайде В.,Ригер Р.( ред.) Зоология беспозвоночных в двух томах. Том 1: от простейших до моллюсков и артропод
Подождите немного. Документ загружается.

376
Annelida
ментов
у
многих видов сразу
же
образуется зона
ро-
ста,
в
которой формируются новые сегменты (иног-
да столько
же,
сколько было утеряно).
Тело полихеты
Clymenella
torquata,
у
которой посто-
янное число сегментов равно
22,
может полностью вос-
станавливаться
из
13 сегментов. Во время опыта были вы-
резаны участки
из
различных отделов тела. Утраченные
сегменты
из
передней
и
задней частей тела были восста-
новлены
во
всех случаях
в
соответствии
со
своей исход-
ной длиной. У
Chaetopterus variopedatus
(Polychaeta) один-
единственный сегмент может наращивать недостающие
участки тела
как
спереди,
так и
сзади,
так что
животное
восстанавливается полностью. Напротив,
Sabella
(Polycha-
eta) никогда не восстанавливает более трёх сегментов; если
же утеряно более трёх передних сегментов,
то
сегменты
из задней части модифицируются в сегменты передней то-
ракальной части. Особенно быстрой регенерацией обла-
дает крона щупалец, которая крайне необходима при сбо-
ре пищи. Именно
эта
часть тела зачастую становится
до-
бычей рыб.
У дождевых червей регенерацию, вероятно, контро-
лирует нейросекреция передних ганглиев. Таким образом,
передний конец тела может быть восстановлен, только
в
том случае, если один из этих ганглиев сохранился. Задняя
часть тела восстанавливается полностью, однако чаще все-
го число новых сегментов не совпадает с первоначальным.
Восстанавливающиеся животные становятся неподвижны-
ми;
кроты пользуются таким поведением: они откусывают
передние сегменты
и
складывают
на
хранение неподвиж-
ных дождевых червей
в
свои хранилища, расположенные
в норах.
Высокая способность тела аннелид
к
репара-
тивной регенерации используется
во
многих случа-
ях
при бесполом размножении.
Каудальная автото-
мия
и
последующая регенерация являются одним
из
этапов жизненного цикла дождевых червей. Деление
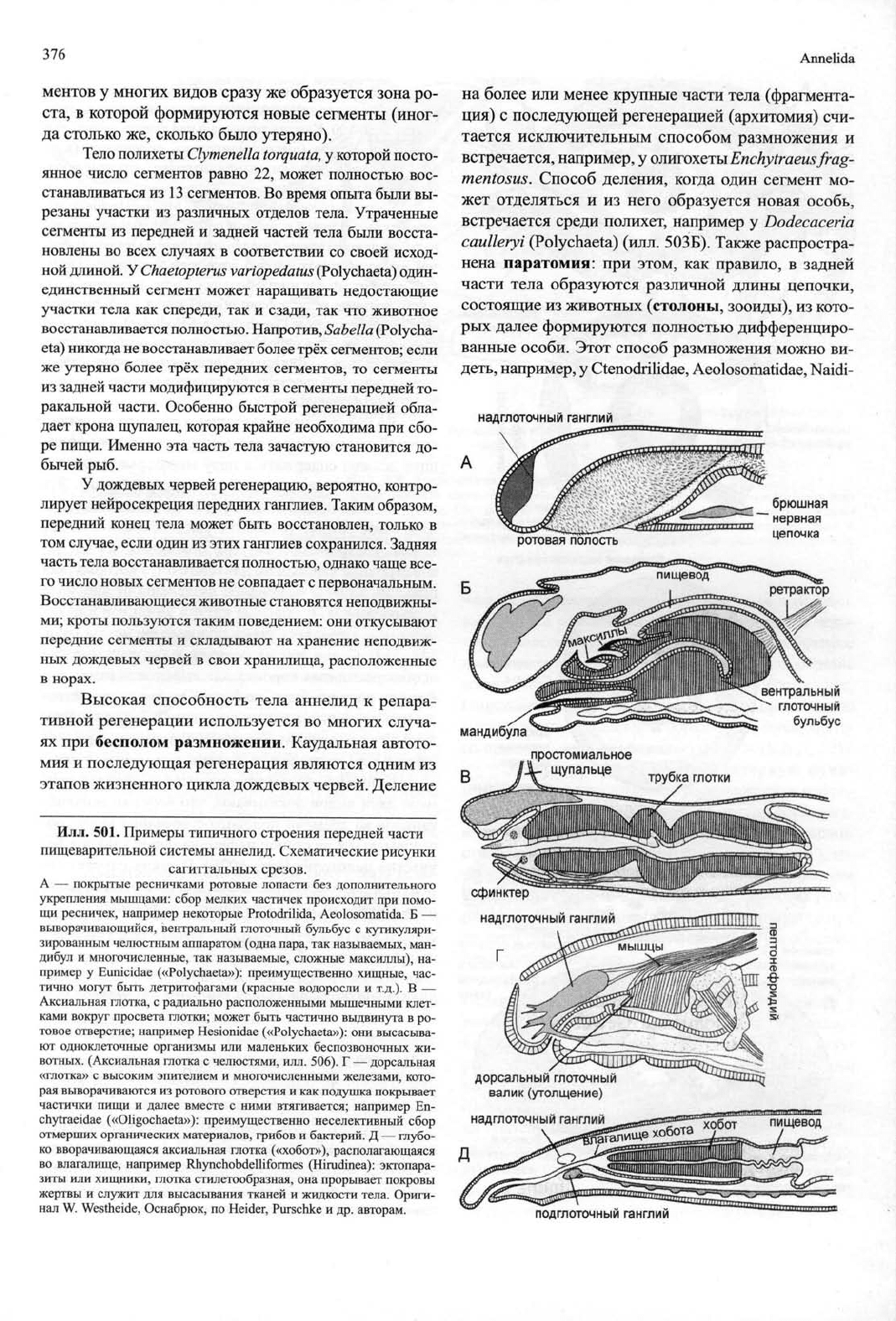
Илл. 501.
Примеры типичного строения передней части
пищеварительной системы аннелид. Схематические рисунки
сагиттальных срезов.
А
—
покрытые ресничками ротовые лопасти
без
дополнительного
укрепления мышцами: сбор мелких частичек происходит
при
помо-
щи ресничек, например некоторые Protodrilida, Aeolosomatida.
Б —
выворачивающийся, вентральный глоточный бульбус
с
кутикуляри-
зированным челюстным аппаратом (одна пара, так называемых, ман-
дибул
и
многочисленные,
так
называемые, сложные максиллы),
на-
пример
у
Eunicidae («Polychaeta»): преимущественно хищные, час-
тично могут быть детритофагами (красные водоросли
и
т.д.).
В —
Аксиальная глотка,
с
радиально расположенными мышечными клет-
ками вокруг просвета глотки; может быть частично выдвинута
в ро-
товое отверстие; например Hesionidae («Polychaeta»):
они
высасыва-
ют одноклеточные организмы
или
маленьких беспозвоночных
жи-
вотных. (Аксиальная глотка
с
челюстями, илл. 506).
Г —
дорсальная
«глотка»
с
высоким эпителием
и
многочисленными железами, кото-
рая выворачиваются
из
ротового отверстия
и как
подушка покрывает
частички пищи
и
далее вместе
с
ними втягивается; например
Еп-
chytraeidae («01igochaeta»): преимущественно неселективный сбор
отмерших органических материалов, грибов
и
бактерий.
Д —
глубо-
ко вворачивающаяся аксиальная глотка («хобот»), располагающаяся
во влагалище, например Rhynchobdelliformes (Hirudinea): эктопара-
зиты
или
хищники, глотка стилетообразная,
она
прорывает покровы
жертвы
и
служит
для
высасывания тканей
и
жидкости тела. Ориги-
нал
W.
Westheide, Оснабрюк,
по
Heider, Purschke
и др.
авторам.
на более или менее крупные части тела (фрагмента-
ция) с последующей регенерацией (архитомия) счи-
тается исключительным способом размножения и
встречается, например, у олигохеты Enchytraeus frag-
mentosus. Способ деления, когда один сегмент мо-
жет отделяться и из него образуется новая особь,
встречается среди полихет, например у Dodecaceria
caulleryi (Polychaeta) (илл. 503Б). Также распростра-
нена паратомия: при этом, как правило, в задней
части тела образуются различной длины цепочки,
состоящие из животных (столоны, зооиды), из кото-
рых далее формируются полностью дифференциро-
ванные особи. Этот способ размножения можно ви-
деть,
например, у Ctenodrilidae, Aeolosomatidae, Naidi-
надглоточный ганглий
брюшная
нервная
цепочка
вентральный
глоточный
бульбус
дорсальный глоточный
валик
(утолщение)
надглоточный ганглий^ хобот
подглоточныи ганглии
http://jurassic.ru/
Annelida
377
соленоциты
/ •"' m
целотелий
|
экстрацеллюлярныи
матрикс
целотелий
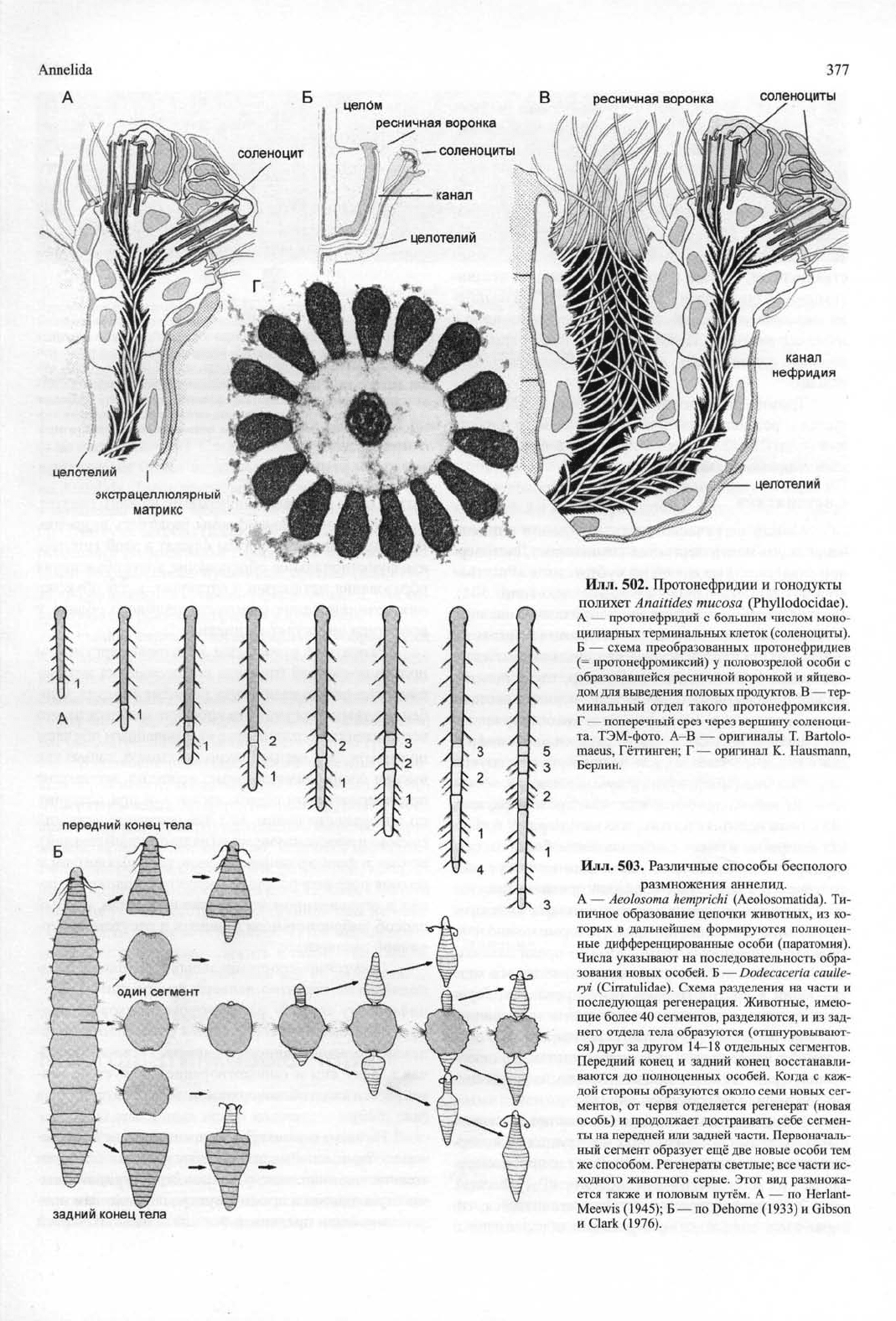
Илл.
502. Протонефридии и гонодукты
полихет
Anaitides mucosa
(Phyllodocidae).
А — протонефридии с большим числом моно-
цилиарных терминальных клеток (соленоциты).
Б — схема преобразованных протонефридиев
(= протонефромиксий) у половозрелой особи с
образовавшейся ресничной воронкой и яйцево-
дом для выведения половых продуктов. В — тер-
минальный отдел такого протонефромиксия.
Г — поперечный срез через вершину соленоци-
та.
ТЭМ-фото. А-В — оригиналы Т. Bartolo-
maeus, Гёттинген; Г — оригинал К. Hausmann,
Берлин.
задний
конец
тела
Илл.
503.
Различные способы бесполого
размножения аннелид.
А — Aeolosoma hemprichi (Aeolosomatida). Ти-
пичное образование цепочки животных, из ко-
торых в дальнейшем формируются полноцен-
ные дифференцированные особи (паратомия).
Числа указывают на последовательность обра-
зования новых особей. Б — Dodecaceria caulle-
ryi (Cirratulidae). Схема разделения на части и
последующая регенерация. Животные, имею-
щие более 40 сегментов, разделяются, и из зад-
него отдела тела образуются (отшнуровывают-
ся) друг за другом 14-18 отдельных сегментов.
Передний конец и задний конец восстанавли-
ваются до полноценных особей. Когда с каж-
дой стороны образуются около семи новых сег-
ментов, от червя отделяется регенерат (новая
особь) и продолжает достраивать себе сегмен-
ты на передней или задней части. Первоначаль-
ный сегмент образует ещё две новые особи тем
же способом. Регенераты светлые; все части ис-
ходного животного серые. Этот вид размножа-
ется также и половым путём. А — по Herlant-
Meewis (1945); Б — по Dehorne (1933) и Gibson
и Clark (1976).
http://jurassic.ru/

378
Annelida
dae (илл. 503A, 554). Такого рода образовавшиеся
животные становятся носителями половых клеток,
например, Syllidae (Polychaeta); и поэтому особи, раз-
множающиеся бесполым и половым путями у этих
видов, обязательно связаны между собой.
Исходным типом полового размножения яв-
ляется выбрасывание гамет в воду и оплодотворе-
ние вне тела животного, как, например, у большин-
ства полихет. Однако в процессе эволюции, незави-
симыми путями среди Polychaeta и Clitellata возник-
ли механизмы прямого переноса сперматозоидов в
полового партнера. Некоторые виды имеют внешние
половые органы, такие как пенис у Pisionidae (Poly-
chaeta).
Трохофорная личинка (илл. 268А,Б, 521) обра-
зуется в результате типичного спирального дробле-
ния (илл. 286, 288); ряд Polychaeta и все Clitellata
имеют прямое развитие.
Систематика
Монофилетическое происхождение аннелид
чаще всего не подвергается сомнениям. Достовер-
ной синапоморфии для обоих субтаксонов «Polycha-
eta» и Clitellata однако же неизвестно (илл. 504).
Многие их общие характерные признаки являются
предковыми, первичными структурами (симплезио-
морфия). Они должны быть уже представлены или у
прототипа всех Articulata (например, такие призна-
ки как, метамерное деление тела, нервная система
лестничного типа, специфическая косоисчерченная
мускулатура, парные целомические мешки и нефри-
дии в каждом сегменте), или возникнуть уже внутри
Articulata (например, кутикулярные щетинки, состо-
ящие из тонких трубочек илл. 494, трохофора, илл.
521).
Если исходить из того, что метамерия у Articu-
lata возникла в связи с образованием боковых сег-
ментарных придатков, то параподии не могут рас-
сматриваться как аутапоморфный признак аннелид.
Аутапоморфиями таксона аннелид являются
только дискуссионные признаки, которые можно най-
ти всего у некоторых групп. Можно предположить,
что эти признаки были у прототипа аннелид и в ходе
эволюции в других группах редуцировались; веро-
ятно,
именно этим можно подтвердить монофилию
таксона Annelida. Это доказательство связано с дис-
куссией о направлении эволюции признаков и связа-
но непосредственно с представлением о филогене-
тическом ряде субтаксонов аннелид.
1.
Традиционная систематика считает производ-
ными группами пресноводных и живущих на повер-
хности почвы поясковых (Clitellata) и противопос-
тавляет их всем другим аннелидам (= «Polychaeta»).
2.
По противоположной гипотезе считается, что
первичные аннелиды вели роющий образ жизни и
Annelida
о
о.
га
Илл. 504. Родственные связи внутри Annelida.
Апоморфии: [1] Простомиум с придатками (антенны, пальпы); ну-
хальные органы. [?] Аутапоморфия полихет неизвестна, вероятно, бес-
поясковые аннелиды образуют парафилетическую группу. [2] Поясок
и формирование кокона, особенности онтогенеза, перемещение моз-
га из простомиума, гермафродитизм, положение гонад только в опре-
делённых сегментах тела и т.д.
были схожи с дождевыми червями; из этого следует,
что полихетоподобные формы развились вторично.
Роющие жизненные формы служат в этой гипотезе
как функциональное обоснование и для объяснения
образования метамерии у артикулят, и для объясне-
ния возникновения олигохетоподобного облика у
прототипа артикулят и аннелид.
Ранее уже приведены некоторые аргументы
против последней гипотезы возникновения метаме-
рии и выведения из неё пути развития аннелид. Осо-
бенно важным аргументом «против» является то, что
все клителляты отличаются чрезвычайным обилием
признаков. Эти черты можно трактовать только как
высоко специализированные, вероятно, возникшие
при пресноводном образе жизни или при ползании
по поверхности почвы. Вот эти признаки: лежащий
глубоко в простомиуме мозг (надглоточный ганглий),
поясок и формирование коконов, гермафродитизм и
прямая передача половых продуктов, положение го-
над в ограниченном количестве сегментов, особый
способ эмбрионального развития и отсутствие пер-
вичной личинки.
Отсутствие простомиальных придатков и пара-
подий также, вероятно, является предпосылкой к спе-
цифичному способу размножения поясковых: про-
цесс образования и сбрасывания с тела плотно при-
лежащего, наполненного жидкостью кокона, (вклю-
чая кладку яиц и оплодотворение) требует по воз-
можности гладкой поверхности тела, без придатков
(илл. 549).
Из этого можно предположить, что не олигохе-
топодобные, а полихетоподобные формы с большим
количеством щетинок, с хорошо структурированны-
ми параподиями и простомиумом, снабжённым при-
датками, были предковой формой кольчатых червей
http://jurassic.ru/

Annelida
379
Илл.
505.
Nereis diversicolor (Nereididae) в движении.
Переход между медленными и быстрыми извиваниями тела. В задней части тела сокращаются продольные мышечные валики поочередно с
левой и правой сторон, так что тело образует горизонтальную волну. Эти волны пробегают по телу в направлении движения, сзади вперёд. На
выпуклой стороне изгиба тела параподий опираются о субстрат, и при дальнейшем движении волны они перемещаются назад, так что тело
переносится параподиями вперёд. В следующем вогнутом изгибе волны продольные мышцы максимально сжаты; соответствующие парапо-
дий сильно укорочены, они отрываются от субстрата и снова переносятся вперёд. Тёмная полоса на спине животного — спинной сосуд.
Оригинал W. Westheide, Оснабрюк.
(а не всех Articulata). Следовательно, простомиум с
придатками может являться аутапоморфией таксо-
на Annelida. Такое представление предполагает, что
у предковой формы артропод не было придатков на
простомиуме (акрон) (илл. 568). По меньшей мере,
рецентные (современные) формы не противоречат
такой теории. Также нухальные органы (илл. 498)
вероятно являются аутапоморфией аннелид; они от-
сутствуют, вероятно, вторично у клителлят.
Однако снова остаётся открытым вопрос: если
Arthropoda не сестринская группа всех Annelida, тогда
являются ли членистоногие лишь подчинённым так-
соном более низкого ранга. Это требует дальнейше-
го изучения. Здесь аннелид, несмотря на сомнения,
мы рассматриваем как монофилетическую группу.
Высокоспециализированные поясковые про-
шли через многие синапоморфии (см. выше) и сфор-
мировались как монофилетическая группа, у всех
полихет не могут найти подходящих, вторичных при-
знаков: «Polychaeta», вероятно, являются парафиле-
тической группой, собранием всех аннелид, которых
нельзя однозначно определить как Clitellata; Clitellata
тогда могут быть сестринской группой ещё не изве-
стной части таких «Polychaeta». Такое определение
позволяет также присоединить к полихетам таксон,
традиционно рассматриваемый как Pogonophora.
1.
«Polychaeta»,
Многощетинковые черви
К многощетинковым червям относятся поряд-
ка 10
ООО
описанных видов кольчатых червей, име-
ющих чрезвычайно разнообразное строение. Важ-
ным отличием полихет является обязательное отсут-
ствие пояска (clitellum). В море они занимают все воз-
можные ниши. В морях полихеты доминируют в бен-
тосных сообществах и формируют кормовую базу
многих организмов, прежде всего рыб. Некоторые
виды живут в солоноватой (опреснённой) воде, име-
ются виды, обитающие в пресной воде или воде, на-
ходящейся в капиллярах почвы. В тропиках во влаж-
ном грунте живут дышащие воздухом Nereididae. Ряд
многощетинковых червей являются облигатными
комменсалами, экто- и эндопаразитами.
При сохранении основного плана строения по-
лихеты дали адаптивную радиацию, выразившуюся
в конструкциях простомиума и перистомиума, пара-
подий и пищедобывательного аппарата. Так возник-
ло многообразие морфотипов питания и движения,
настолько различных, что его изначальное состояние
теперь можно представить только по некоторым пред-
ставителям семейств «традиционных» многощетин-
ковых червей. Несколько разделов главы посвяще-
ны Aeolosomatida, Potammodrilida, Myzostomida, Po-
gonophora и Lobatocerebrida. Организация Myzosto-
mida и Pogonophora сильно отличается, однако в них
хорошо различимы признаки аннелид. Принадлеж-
ность к аннелидам Lobatocerebrida, тело которых не
сегментировано, удалось определить только благо-
даря их ультраструктуре.
Строение
Вероятно, первичные полихеты были длинным,
по большей части хищными формами с многочис-
ленными, одинаковой формы (гомономными) сегмен-
тами тела. Nereididae (илл. 505) несут хорошо раз-
витый простомиум с мозгом, одной парой антенн и
одной парой двучлениковых пальп с многочислен-
ными рецепторами (илл. 506), четыре глаза и два ну-
хальных органа. Следующий за ним перистомиум
сливается с первым щетинконосным сегментом и
несёт четыре пары длинных, направленных вперёд
щупалец — перистомиальных усиков.
Часть передней кишки преобразована в акси-
альную мускулистую трубку (глотку), вооружённую
http://jurassic.ru/
380
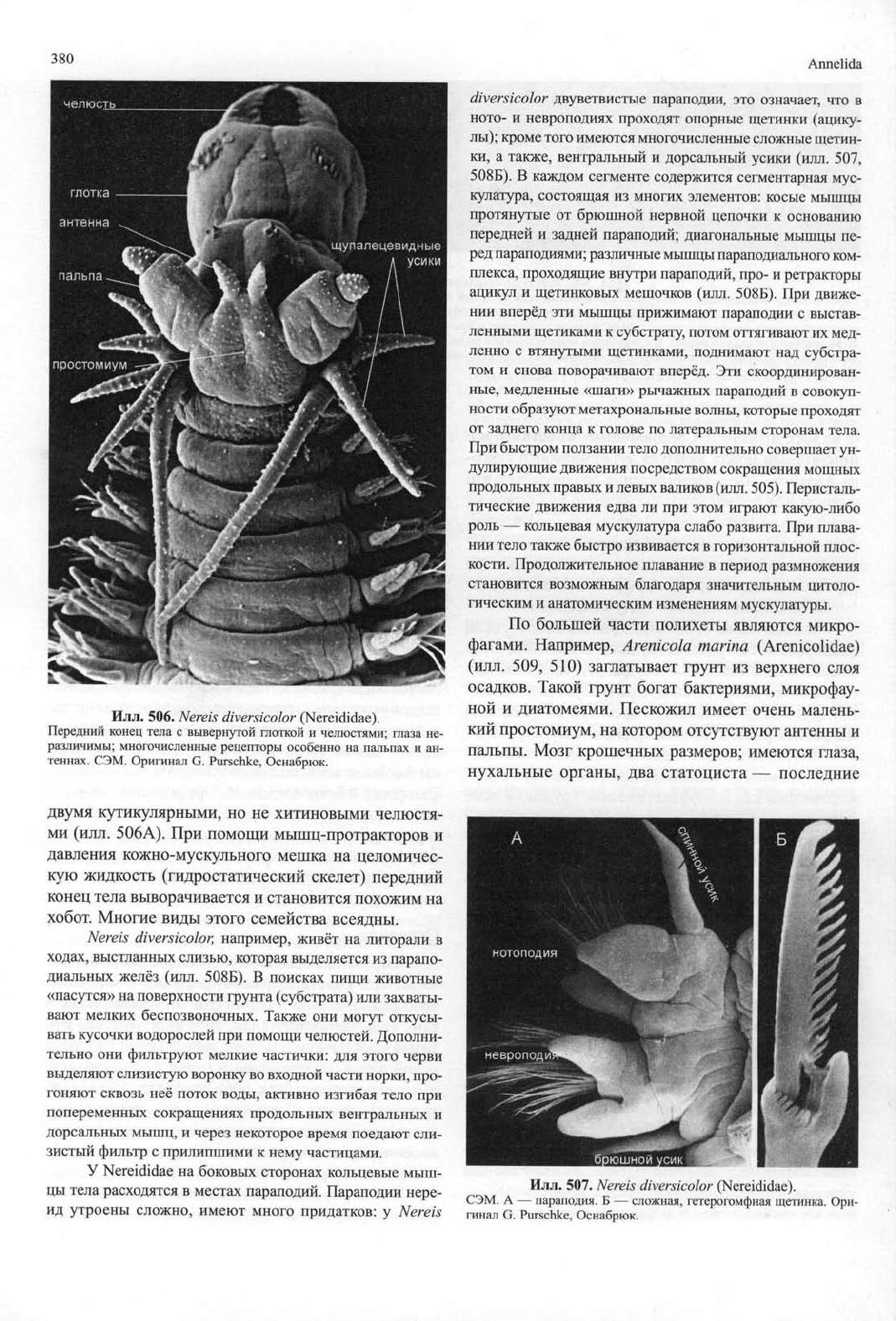
Илл.
506.
Nereis diversicolor
(Nereididae).
Передний конец тела
с
вывернутой глоткой
и
челюстями; глаза
не-
различимы; многочисленные рецепторы особенно
на
пальпах
и ан-
теннах. СЭМ. Оригинал
G.
Purschke, Оснабрюк.
двумя кутикулярными, но не хитиновыми челюстя-
ми (илл. 506А). При помощи мышц-протракторов и
давления кожно-мускульного мешка на целомичес-
кую жидкость (гидростатический скелет) передний
конец тела выворачивается и становится похожим на
хобот. Многие виды этого семейства всеядны.
Nereis
diversicolor,
например, живёт
на
литорали
в
ходах, выстланных слизью, которая выделяется из парапо-
диальных желёз
(илл.
508Б).
В
поисках пищи животные
«пасутся» на поверхности грунта (субстрата) или захваты-
вают мелких беспозвоночных. Также
они
могут откусы-
вать кусочки водорослей при помощи челюстей. Дополни-
тельно
они
фильтруют мелкие частички:
для
этого черви
выделяют слизистую воронку во входной части норки, про-
гоняют сквозь
неё
поток воды, активно изгибая тело
при
попеременных сокращениях продольных вентральных
и
дорсальных мышц,
и
через некоторое время поедают сли-
зистый фильтр
с
прилипшими к нему частицами.
У Nereididae
на
боковых сторонах кольцевые мыш-
цы тела расходятся
в
местах параподии. Параподии нере-
ид утроены сложно, имеют много придатков:
у Nereis
Annelida
Илл.
507.
Nereis diversicolor
(Nereididae).
СЭМ.
А —
параподия.
Б —
сложная, гетерогомфная щетинка. Ори-
гинал
G.
Purschke, Оснабрюк.
diversicolor
двуветвистые параподии,
это
означает,
что в
ното-
и
невроподиях проходят опорные щетинки (ацику-
лы);
кроме того имеются многочисленные сложные щетин-
ки,
а
также, вентральный
и
дорсальный усики (илл.
507,
508Б).
В
каждом сегменте содержится сегментарная мус-
кулатура, состоящая
из
многих элементов: косые мышцы
протянутые
от
брюшной нервной цепочки
к
основанию
передней
и
задней параподии; диагональные мышцы
пе-
ред параподиями; различные мышцы параподиального
ком-
плекса, проходящие внутри параподии, про- и ретракторы
ацикул
и
щетинковых мешочков (илл. 508Б). При движе-
нии вперёд
эти
мышцы прижимают параподии
с
выстав-
ленными щетиками
к
субстрату, потом оттягивают их мед-
ленно
с
втянутыми щетинками, поднимают
над
субстра-
том
и
снова поворачивают вперёд.
Эти
скоординирован-
ные, медленные «шаги» рычажных параподии
в
совокуп-
ности образуют метахрональные волны, которые проходят
от заднего конца
к
голове
по
латеральным сторонам тела.
При быстром ползании тело дополнительно совершает ун-
дулирующие движения посредством сокращения мощных
продольных правых и левых
валиков
(илл.
505). Перисталь-
тические движения едва
ли при
этом играют какую-либо
роль
—
кольцевая мускулатура слабо развита. При плава-
нии тело также быстро извивается в горизонтальной плос-
кости. Продолжительное плавание
в
период размножения
становится возможным благодаря значительным цитоло-
гическим
и
анатомическим изменениям мускулатуры.
По большей части полихеты являются микро-
фагами. Например, Arenicola marina (Arenicolidae)
(илл. 509, 510) заглатывает грунт из верхнего слоя
осадков. Такой грунт богат бактериями, микрофау-
ной и диатомеями. Пескожил имеет очень малень-
кий простомиум, на котором отсутствуют антенны и
пальпы. Мозг крошечных размеров; имеются глаза,
нухальные органы, два статоциста — последние
http://jurassic.ru/
Annelida
381
пальпа
задний
корешок
окологлоточной
коннективы
кишка
спинная
продольная
мускулатура
брюшная
продольная мускулатура
мускулатура
параподий
кольцевая
мускулатура
мышцы
ацикулы
параподиальныи
ганглий
вентральный
ганглий
латеральный
нервный
тяж
ретрактор
протрактор
косые
мышцы
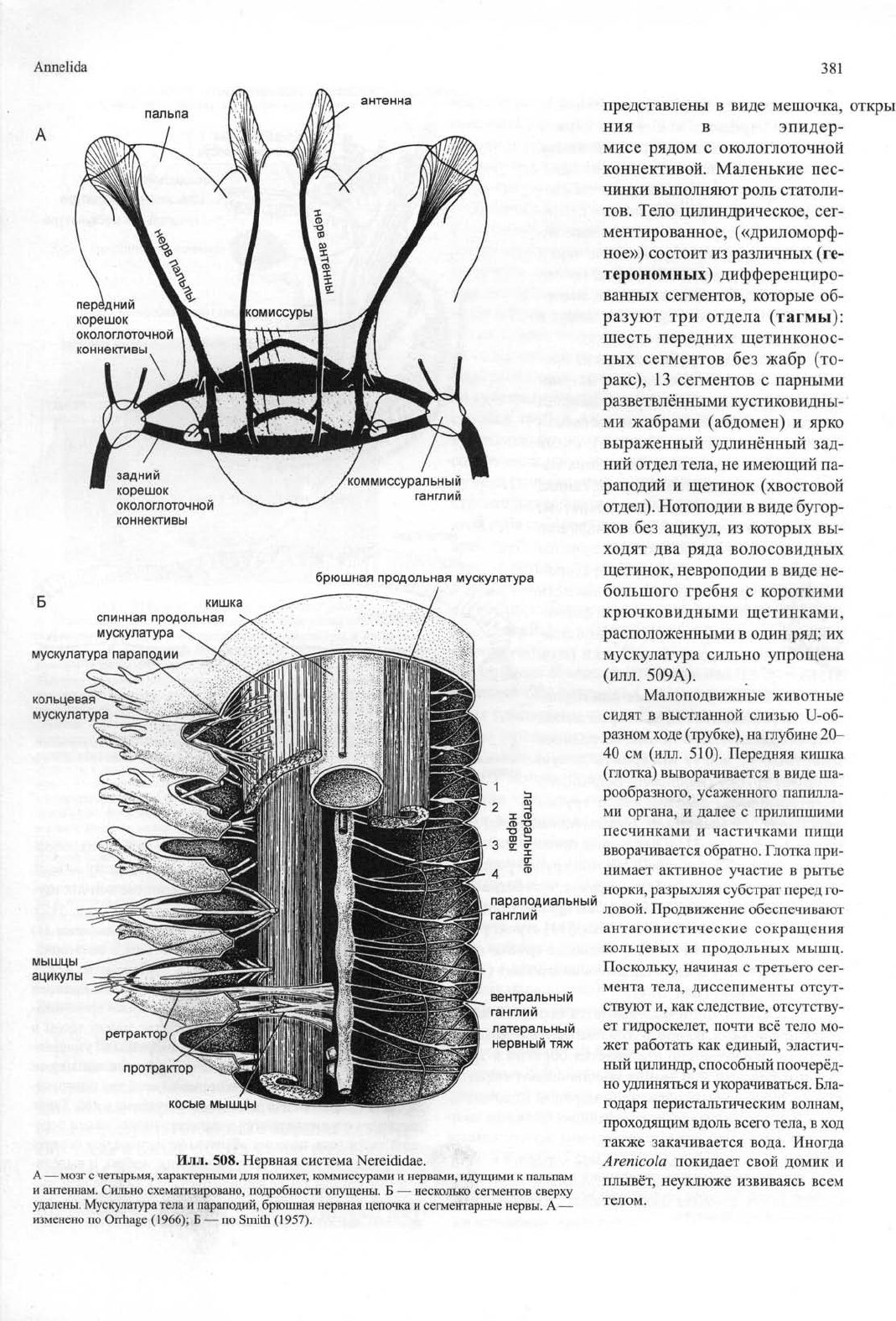
Илл. 508. Нервная система Nereididae.
А — мозг с четырьмя, характерными для полихет, коммиссурами и нервами, идущими к пальпам
и антеннам. Сильно схематизировано, подробности опущены. Б — несколько сегментов сверху
удалены. Мускулатура тела и параподий, брюшная нервная цепочка и сегментарные нервы. А —
изменено по Orrhage (1966); Б — по Smith (1957).
представлены в виде мешочка, откры
ния в эпидер-
мисе рядом с окологлоточной
коннективой. Маленькие пес-
чинки выполняют роль статоли-
тов.
Тело цилиндрическое, сег-
ментированное, («дриломорф-
ное») состоит из различных (ге-
терономных) дифференциро-
ванных сегментов, которые об-
разуют три отдела (тагмы):
шесть передних щетинконос-
ных сегментов без жабр (то-
ракс),
13 сегментов с парными
разветвлёнными кустиковидны-
ми жабрами (абдомен) и ярко
выраженный удлинённый зад-
ний отдел тела, не имеющий па-
раподий и щетинок (хвостовой
отдел).
Нотоподии в виде бугор-
ков без ацикул, из которых вы-
ходят два ряда волосовидных
щетинок, невроподии в виде не-
большого гребня с короткими
крючковидными щетинками,
расположенными в один ряд; их
мускулатура сильно упрощена
(илл. 509А).
Малоподвижные животные
сидят в выстланной слизью U-об-
разном ходе (трубке), на глубине 20-
40 см (илл. 510). Передняя кишка
(глотка) выворачивается в виде ша-
рообразного, усаженного папилла-
ми органа, и далее с прилипшими
песчинками и частичками пищи
вворачивается обратно. Глотка при-
нимает активное участие в рытье
норки, разрыхляя субстрат перед го-
ловой. Продвижение обеспечивают
антагонистические сокращения
кольцевых и продольных мышц.
Поскольку, начиная с третьего сег-
мента тела, диссепименты отсут-
ствуют и, как следствие, отсутству-
ет гидроскелет, почти всё тело мо-
жет работать как единый, эластич-
ный цилиндр, способный поочерёд-
но удлиняться
и
укорачиваться. Бла-
годаря перистальтическим волнам,
проходящим вдоль всего тела,
в
ход
также закачивается вода. Иногда
Arenicola
покидает свой домик и
плывёт, неуклюже извиваясь всем
телом.
http://jurassic.ru/
382
Annelida
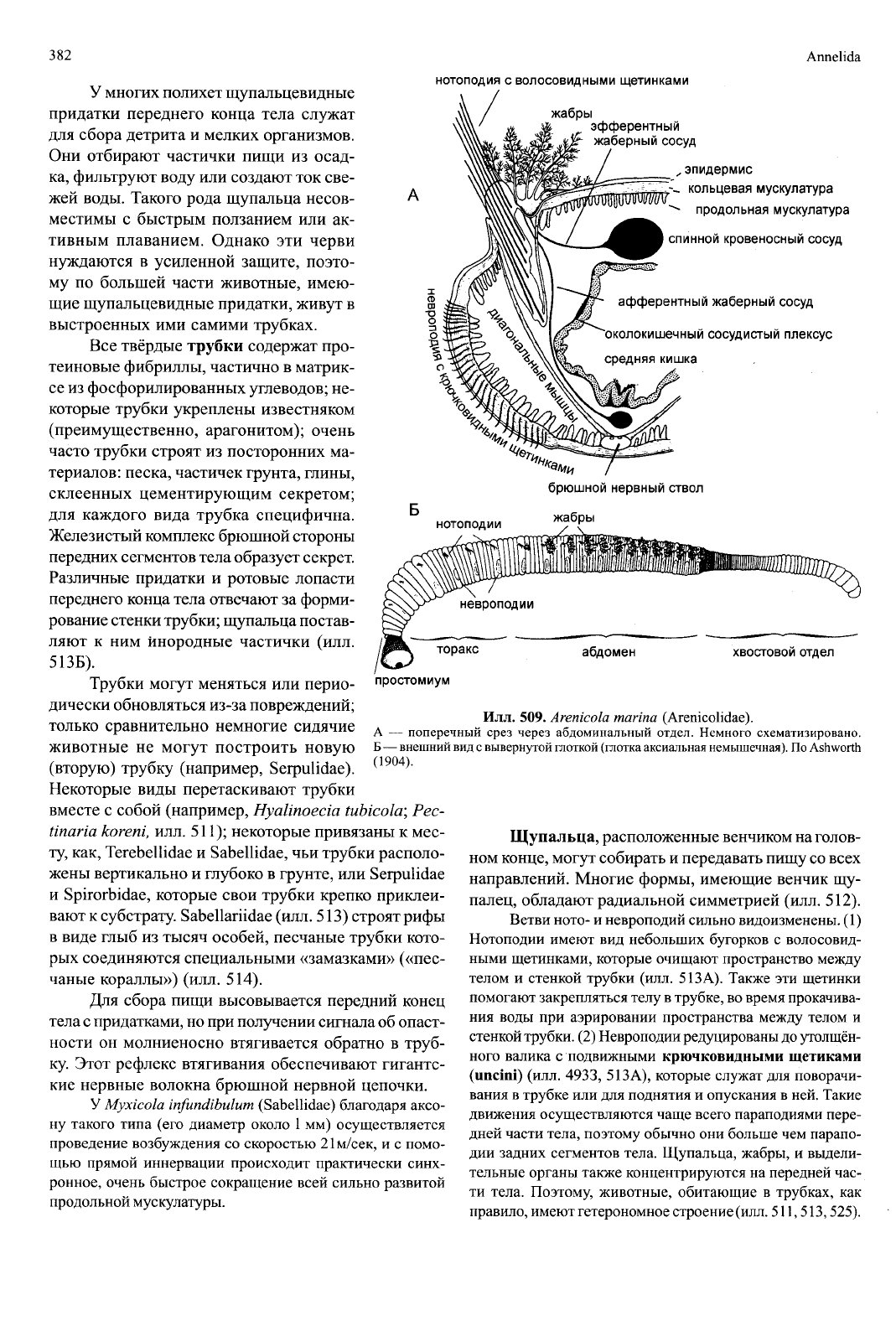
У многих полихет щупальцевидные
придатки переднего конца тела служат
для сбора детрита
и
мелких организмов.
Они отбирают частички пищи
из
осад-
ка, фильтруют воду или создают ток све-
жей воды. Такого рода щупальца несов-
местимы
с
быстрым ползанием
или ак-
тивным плаванием. Однако
эти
черви
нуждаются
в
усиленной защите, поэто-
му
по
большей части животные, имею-
щие щупальцевидные придатки, живут
в
выстроенных
ими
самими трубках.
Все твёрдые трубки содержат про-
теиновые фибриллы, частично
в
матрик-
се из фосфорилированных углеводов; не-
которые трубки укреплены известняком
(преимущественно, арагонитом); очень
часто трубки строят
из
посторонних
ма-
териалов: песка, частичек грунта, глины,
склеенных цементирующим секретом;
для каждого вида трубка специфична.
Железистый комплекс брюшной стороны
передних сегментов тела образует секрет.
Различные придатки
и
ротовые лопасти
переднего конца тела отвечают
за
форми-
рование стенки трубки; щупальца постав-
ляют
к ним
инородные частички
(илл.
513Б).
Трубки могут меняться
или
перио-
дически обновляться из-за повреждений;
только сравнительно немногие сидячие
животные
не
могут построить новую
(вторую) трубку (например, Serpulidae).
Некоторые виды перетаскивают трубки
вместе
с
собой (например, Hyalinoecia tubicola;
Рес-
tinaria koreni, илл. 511); некоторые привязаны
к
мес-
ту, как, Terebellidae
и
Sabellidae, чьи трубки располо-
жены вертикально
и
глубоко
в
грунте, или Serpulidae
и Spirorbidae, которые свои трубки крепко приклеи-
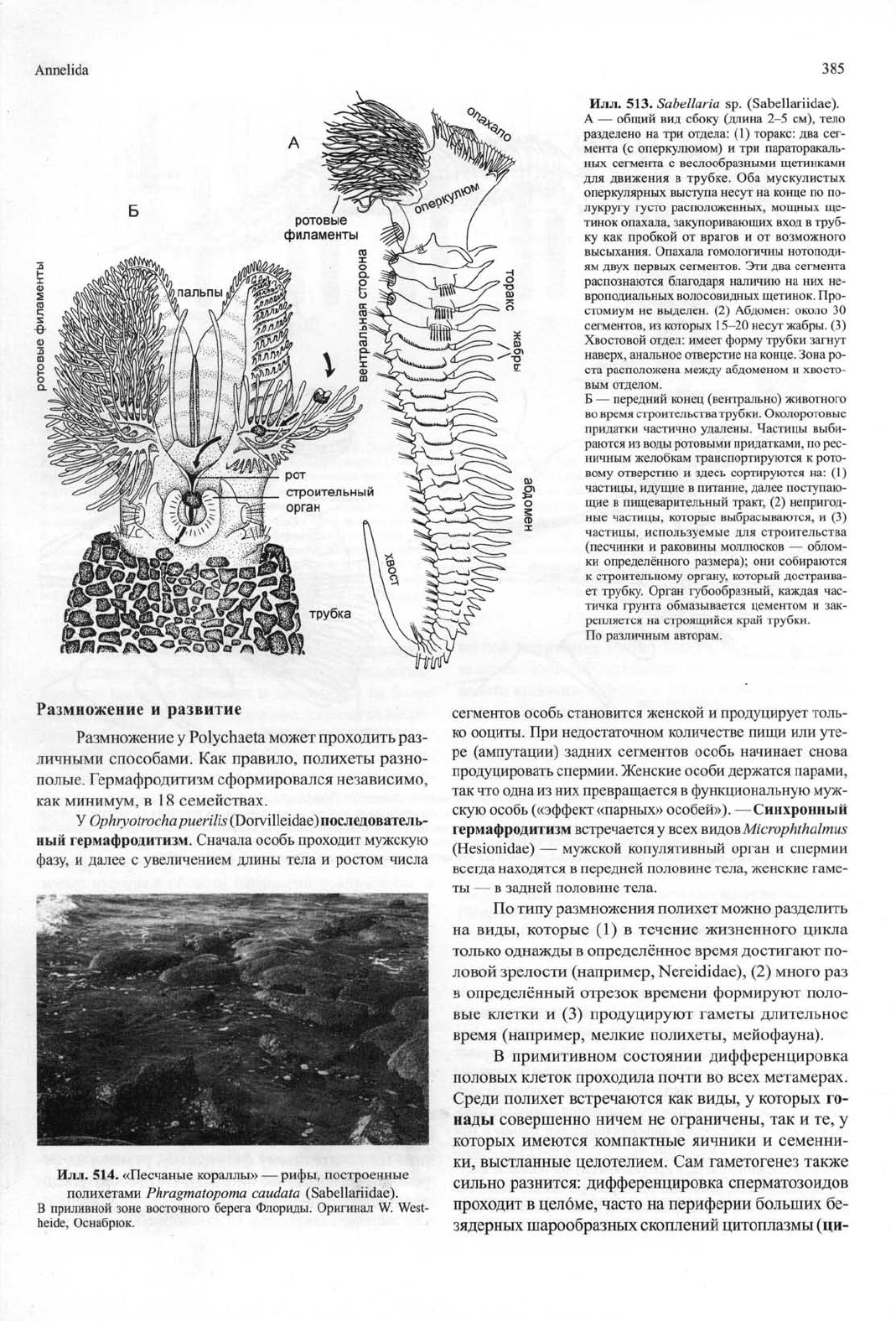
вают к субстрату. Sabellariidae (илл. 513) строят рифы
в виде глыб
из
тысяч особей, песчаные трубки кото-
рых соединяются специальными «замазками» («пес-
чаные кораллы») (илл.
514).
Для сбора пищи высовывается передний конец
тела
с
придатками, но при получении сигнала об опаст-
ности
он
молниеносно втягивается обратно
в
труб-
ку. Этот рефлекс втягивания обеспечивают гигантс-
кие нервные волокна брюшной нервной цепочки.
У
Myxicola infundibulum
(Sabellidae) благодаря аксо-
ну такого типа
(его
диаметр около
1
мм)
осуществляется
проведение возбуждения
со
скоростью 21м/сек,
и с
помо-
щью прямой иннервации происходит практически синх-
ронное, очень быстрое сокращение всей сильно развитой
продольной мускулатуры.
нотоподия
с
волосовидными
щетинками
жабры
эфферентный
жаберный сосуд
,
эпидермис
--.
кольцевая мускулатура
продольная мускулатура
спинной кровеносный сосуд
афферентный жаберный сосуд
'околокишечный сосудистый плексус
средняя кишка
нотоподии
*****
брюшной нервный ствол
жабры
хвостовой отдел
простомиум
Илл.
509.
Arenicola marina
(Arenicolidae).
А
—
поперечный срез через абдоминальный отдел. Немного схематизировано.
Б
—
внешний вид
с
вывернутой глоткой (глотка аксиальная немышечная). По Ashworth
(1904).
Щупальца, расположенные венчиком на голов-
ном конце, могут собирать и передавать пищу со всех
направлений. Многие формы, имеющие венчик
щу-
палец, обладают радиальной симметрией (илл. 512).
Ветви ното- и невроподий сильно видоизменены. (1)
Нотоподии имеют
вид
небольших бугорков
с
волосовид-
ными щетинками, которые очищают пространство между
телом
и
стенкой трубки (илл. 513А). Также
эти
щетинки
помогают закрепляться телу
в
трубке, во время прокачива-
ния воды
при
аэрировании пространства между телом
и
стенкой трубки. (2) Невроподий редуцированы до утолщён-
ного валика
с
подвижными
крючковидными щегиками
(uncini)
(илл. 4933, 513А), которые служат для поворачи-
вания в трубке или для поднятия
и
опускания в ней. Такие
движения осуществляются чаще всего параподиями пере-
дней части тела, поэтому обычно они больше чем парапо-
дии задних сегментов тела. Щупальца, жабры,
и
выдели-
тельные органы также концентрируются на передней час-
ти тела. Поэтому, животные, обитающие
в
трубках,
как
правило, имеют гетерономное строение
(илл.
511,513,525).
http://jurassic.ru/

Annelida
383
сокращение сокращение
^
продольных мышц кольцевых мышц
ход
(«галерея»)
Илл.
510.
Arenicola marina
(Arenicolidae).
А — зарывание в грунт: (1) сжатый передний конец тела набухает
(расширяется) (сокращение продольной мускулатуры и нагнетание
целомической жидкости из середины тела), он служит для заякори-
вания, в то время как задняя часть проталкивается через субстрат. (2)
Вывернутая глотка разрыхляет песок перед головным концом; далее
вытянутый передний конец закрепляется при помощи кольцевых
«фланцев» на передних сегментах. После этого глотка вворачивается
обратно, благодаря чему песок выталкивается назад, а в образовав-
шийся таким образом проход проникает вода. Благодаря растяжению
всего тела (сокращение кольцевой мускулатуры) животное протал-
кивается в разрыхленную часть осадка. Б — срез норки. Животное
лежит в горизонтальном отделе (галерее) L-образного хода, чьи стен-
ки укреплены слизью. Волны, проходящие через всё тело, создают
поток воды, который входит через заднее отверстие норки (около 190
мл/час). Вода из норки также выходит через вертикальную, запол-
ненную песком часть «шахты» (тонкие стрелки!), расположенную над
головой червя. В основании этой «шахты» животное постоянно по-
едает грунт, из-за чего грунт просачивается сверху вниз (толстые
стрелки!). В результате на поверхности субстрата образуется ворон-
ка, в которую собирается особенно много органического материала,
после чего попадает к червю. Одновременно с этим взвешенный ма-
териал отфильтровывается животным с током воды из песка и посту-
пает в пищу. В течение около 40 мин животное поднимается на по-
верхность и выделяет непереваренный песок в виде фекалий. Кисло-
род поступает через древовидные жабры, которые располагаются над
дорсальными пучками щетинок в середине тела (илл. 509). А — по
Trueman (1966); Б — по различным авторам.
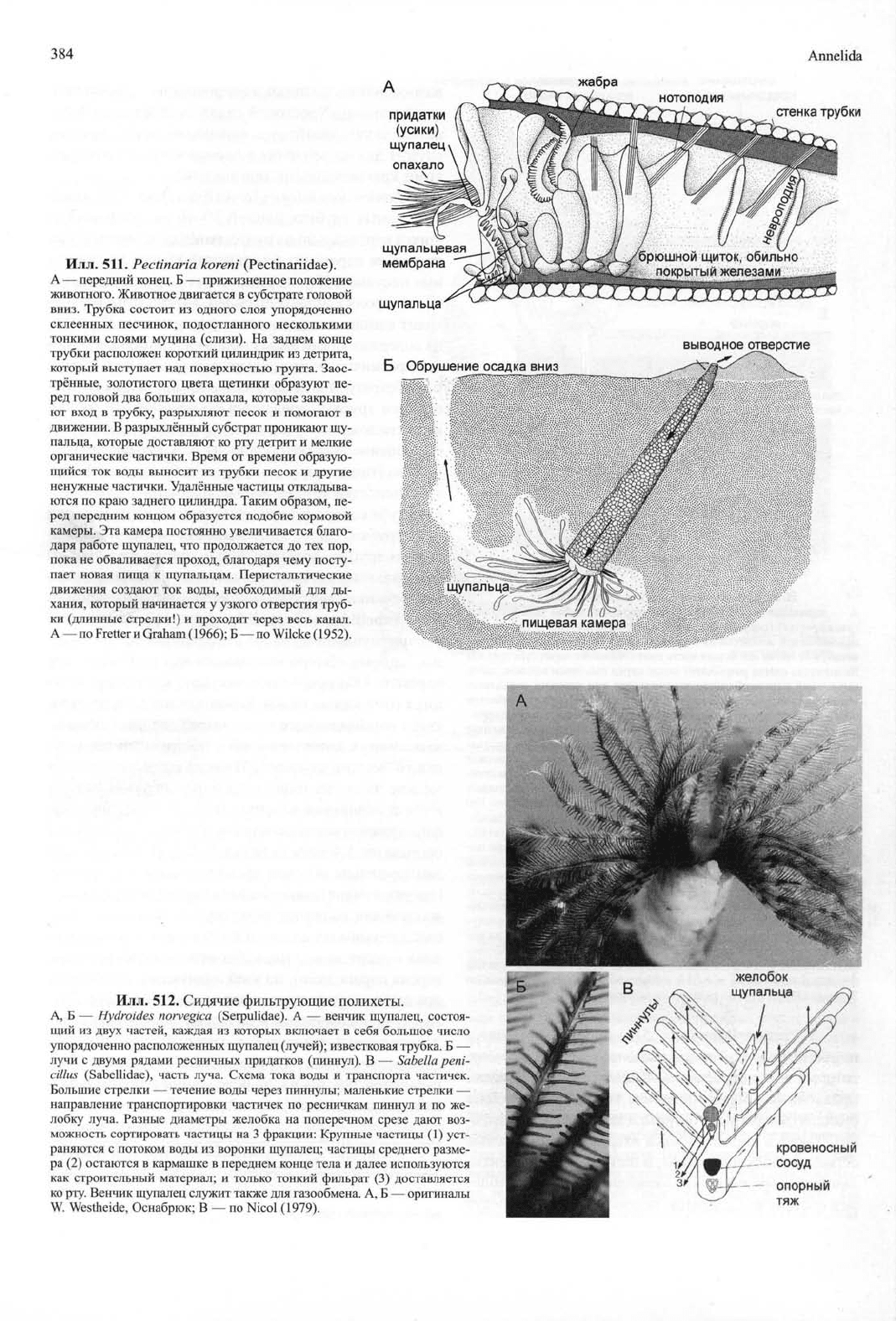
У Pectinariidae (илл. 511) простомиум слит с
перистомиумом. Торакс включает в себя сегмент,
который несёт особенно большие гребневидные, на-
правленные вперёд щетинки, это так называемые
опахала (для рытья субстрата и закрытия входа труб-
ки).
Также в состав торакса входят бесщетинковые
сегменты, несущие жабры, и щетинковые сегменты,
лишённые брюшных крючковидных щетинок. Абдо-
мен состоит из сегментов, нотоподии которых несут
волосовидные щетинки, а невроподии — крючковид-
ные щетинки. Хвостовой отдел преобразован в ло-
патовидную, лишённую параподий скафу, которая
служит для закрепления в песчаной трубке несколь-
кими крючковидными щетинками*.
Lanice conchilega (Terebellidae, илл. 527) живёт
в песчаных трубках длиной 30—40 см, располагаю-
щихся вертикально в грунте, наверху, в верхней час-
ти трубка переходит в ветвистый кустик из склеен-
ных песчинок. Перистомиум слит с простомиумом и
несёт корону длинных щупалец, внутри которых про-
ходит единое целомическое пространство. Щупаль-
ца содержат много желёз и хеморецепторов, с помо-
щью ресничного эпителия щупальца могут «ползать»
по субстрату. Частички пищи или материалы для по-
стройки трубки передаются по ресничному желоб-
ку, расположенному вдоль посередине щупальца, или
сокращение щупалец подносит их к переднему кон-
цу тела (тип питания — посредством щупалец). При
строительстве трубки особая капюшоновидная вер-
хняя губа собирает песчинки и кладёт их на верхний
край трубки. После этого животное немного выпол-
зает из трубки, проводит вдоль стенки железистым
вентральным органом и склеивает (зашпаклёвыва-
ет) песчинки быстрозастывающим свежим секретом.
Sabellidae и Serpulidae (илл. 382, 512) имеют
венчик щупалец и (жабр) разделённых на две груп-
пы,
которые обычно выставлены над трубкой в виде
воронки. Оба основания, несущих щупальца, явля-
ются гомологами пальп. Каждое щупальце, по стро-
ению напоминающее перо, имеет два ряда реснич-
ных пиннул, которые создают постоянный ток воды
сквозь венчик щупалец. Из тока воды отбираются
мелкие частички пищи и сложным образом переда-
ются в основание щупалец (илл. 512В) (например,
фильтрационная способность Pomatoceros triqueter
составляет 1,4 мл/ч-мг веса). У Serpulidae одно или
два щупальца обычно преобразованы в крышечку
(оперкулюм) на ножке, часто кальцинированную, слу-
жащую для закрытия входа трубки. В тораксе длин-
ные щетинки находятся в нотоподиях, а в невропо-
диях — крючковидные, в абдомене — наоборот («ин-
версия параподий»); по всей видимости, такой поря-
док щетинок упрощает поворачивание в трубке. Дру-
гим приспособлением к сидячему образу жизни яв-
ляется ресничный желобок, который проходит в аб-
доминальной части на вентральной стороне, а в то-
ракальной тянется вдоль дорсальной стороны и слу-
жит для выведения фекалий из трубки.
* Согласно другой трактовке сегментов тела скафа со-
ответствует абдомену, а сегменты с нотоподиями — тораксу,
«крючковидные» щетинки скафы являются видоизменённы-
ми нотохетами
(прим.
ред.).
http://jurassic.ru/
384
Annelida
жабра
Илл.
511.
Pectinaria koreni
(Pectinariidae).
A — передний конец. Б — прижизненное положение
животного. Животное двигается в субстрате головой
вниз. Трубка состоит из одного слоя упорядоченно
склеенных песчинок, подостланного несколькими
тонкими слоями муцина (слизи). На заднем конце
трубки расположен короткий цилиндрик из детрита,
который выступает над поверхностью грунта. Заос-
трённые, золотистого цвета щетинки образуют пе-
ред головой два больших опахала, которые закрыва-
ют вход в трубку, разрыхляют песок и помогают в
движении. В разрыхлённый субстрат проникают щу-
пальца, которые доставляют ко рту детрит и мелкие
органические частички. Время от времени образую-
щийся ток воды выносит из трубки песок и другие
ненужные частички. Удалённые частицы откладыва-
ются по краю заднего цилиндра. Таким образом, пе-
ред передним концом образуется подобие кормовой
камеры. Эта камера постоянно увеличивается благо-
даря работе щупалец, что продолжается до тех пор,
пока не обваливается проход, благодаря чему посту-
пает новая пища к щупальцам. Перистальтические
движения создают ток воды, необходимый для ды-
хания, который начинается у узкого отверстия труб-
ки (длинные стрелки!) и проходит через весь канал.
А — по Fretter и Graham (1966); Б — по Wilcke (1952).
нотоподия
стенка
трубки
щупальцевая
мембрана
щупальца
выводное
отверстие
Б
Обрушение
осадка
вниз
пищевая
камера
Илл. 512. Сидячие фильтрующие полихеты.
А, Б — Hydroides norvegica (Serpulidae). А — венчик щупалец, состоя-
щий из двух частей, каждая из которых включает в себя большое число
упорядоченно расположенных щупалец (лучей); известковая трубка. Б —
лучи с двумя рядами ресничных придатков (пиннул). В — Sabella peni-
cillus (Sabellidae), часть луча. Схема тока воды и транспорта частичек.
Большие стрелки — течение воды через пиннулы; маленькие стрелки —
направление транспортировки частичек по ресничкам пиннул и по же-
лобку луча. Разные диаметры желобка на поперечном срезе дают воз-
можность сортировать частицы на 3 фракции: Крупные частицы (1) уст-
раняются с потоком воды из воронки щупалец; частицы среднего разме-
ра (2) остаются в кармашке в переднем конце тела и далее используются
как строительный материал; и только тонкий фильрат (3) доставляется
ко рту. Венчик щупалец служит также для газообмена. А, Б — оригиналы
W. Westheide, Оснабрюк; В — по Nicol (1979).
кровеносный
сосуд
опорный
тяж
http://jurassic.ru/
Annelida
385
Илл.
513.
Sabellaria sp. (Sabellariidae).
A — общий вид сбоку (длина 2-5 см), тело
разделено на три отдела: (1) торакс: два сег-
мента (с оперкулюмом) и три параторакаль-
ных сегмента с веслообразными щетинками
для движения в трубке. Оба мускулистых
оперкулярных выступа несут на конце по по-
лукругу густо расположенных, мощных ще-
тинок опахала, закупоривающих вход в труб-
ку как пробкой от врагов и от возможного
высыхания.
Опахала гомологичны нотоподи-
ям двух первых сегментов. Эти два сегмента
распознаются благодаря наличию на них не-
вроподиальных волосовидных щетинок. Про-
стомиум не выделен. (2) Абдомен: около 30
сегментов,
из которых 15-20 несут жабры. (3)
Хвостовой отдел: имеет форму трубки загнут
наверх,
анальное отверстие на конце. Зона ро-
ста расположена между абдоменом и хвосто-
вым отделом.
Б — передний конец (вентрально) животного
во время строительства трубки. Околоротовые
придатки частично удалены. Частицы выби-
раются из воды ротовыми придатками, по рес-
ничным желобкам транспортируются к рото-
вому отверстию и здесь сортируются на: (1)
частицы,
идущие в питание, далее поступаю-
щие в пищеварительный тракт, (2) непригод-
ные частицы, которые выбрасываются, и (3)
частицы,
используемые для строительства
(песчинки и раковины моллюсков — облом-
ки определённого размера); они собираются
к строительному органу, который достраива-
ет трубку. Орган губообразный, каждая час-
тичка грунта обмазывается цементом и зак-
репляется на строящийся край трубки.
По различным авторам.
Размножение и развитие
Размножение у Polychaeta может проходить раз-
личными способами. Как правило, полихеты разно-
полые. Гермафродитизм сформировался независимо,
как минимум, в 18 семействах.
У
Ophryotrocha puerilis (Dorvilleidae)
последователь-
ный гермафродитизм.
Сначала особь проходит мужскую
фазу, и далее с увеличением длины тела и ростом числа
Илл.
514. «Песчаные кораллы» — рифы, построенные
полихетами Phragmatopoma caudata (Sabellariidae).
В приливной зоне восточного берега Флориды. Оригинал W. West-
heide,
Оснабрюк.
сегментов особь становится женской и продуцирует толь-
ко ооциты. При недостаточном количестве пищи или уте-
ре (ампутации) задних сегментов особь начинает снова
продуцировать спермин. Женские особи держатся парами,
так что одна из них превращается
в
функциональную муж-
скую особь («эффект «парных» особей»).
—Синхронный
гермафродитизм встречается у всех видов Microphthalmus
(Hesionidae) — мужской копулятивный орган и спермин
всегда находятся в передней половине тела, женские гаме-
ты — в задней половине тела.
По типу размножения полихет можно разделить
на виды, которые (1) в течение жизненного цикла
только однажды в определённое время достигают по-
ловой зрелости (например, Nereididae), (2) много раз
в определённый отрезок времени формируют поло-
вые клетки и (3) продуцируют гаметы длительное
время (например, мелкие полихеты, мейофауна).
В примитивном состоянии дифференцировка
половых клеток проходила почти во всех метамерах.
Среди полихет встречаются как виды, у которых го-
нады совершенно ничем не ограничены, так и те, у
которых имеются компактные яичники и семенни-
ки,
выстланные целотелием. Сам гаметогенез также
сильно разнится: дифференцировка сперматозоидов
проходит в целоме, часто на периферии больших бе-
зядерных шарообразных скоплений цитоплазмы (ци-
http://jurassic.ru/