Вестхайде В.,Ригер Р.( ред.) Зоология беспозвоночных в двух томах. Том 1: от простейших до моллюсков и артропод
Подождите немного. Документ загружается.


216
Трёхслойные Eumetazoa
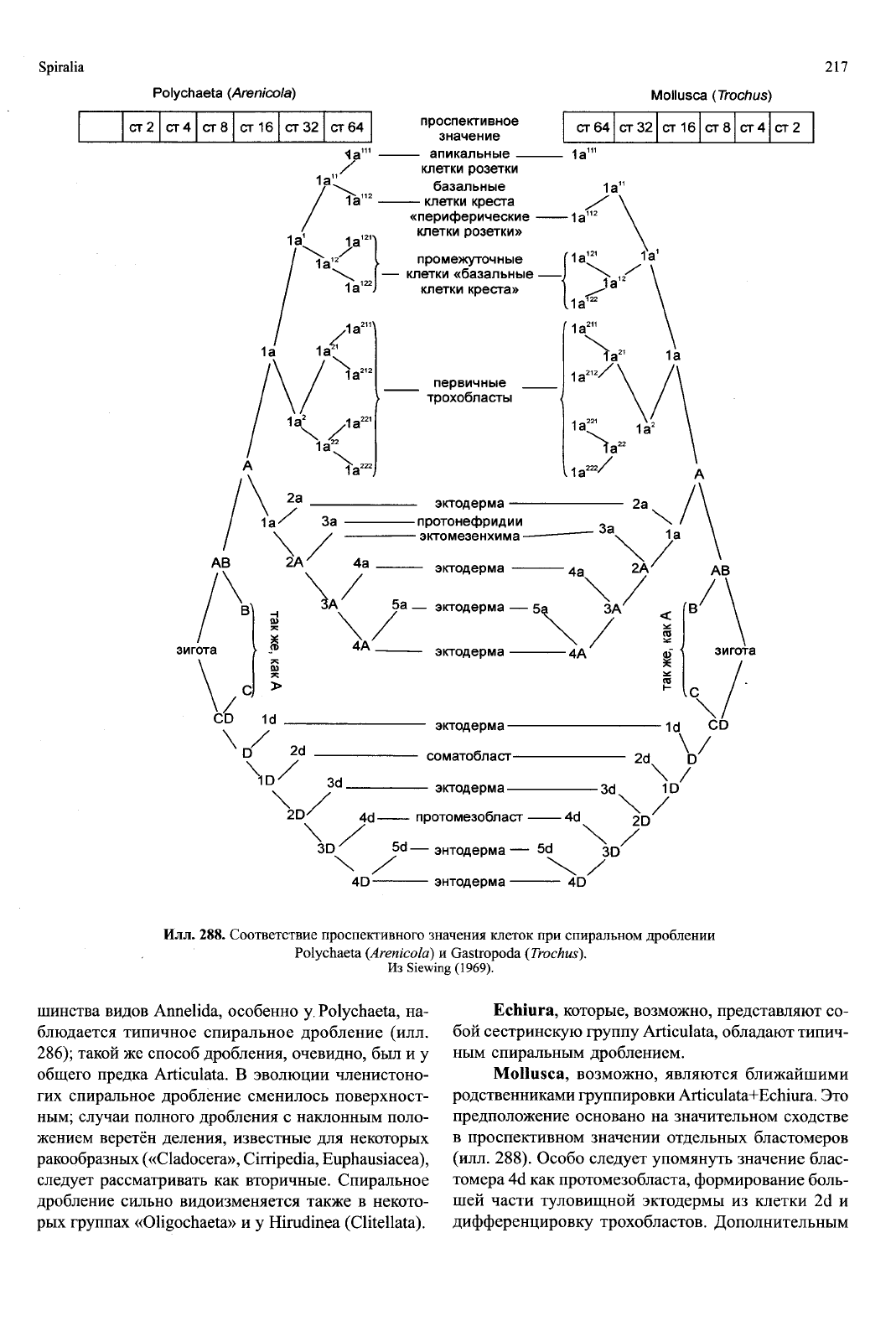
квартет макромеров ещё раз делится экваториально
и несколько неравномерно, в результате чего образу-
ется второй квартет микромеров (клетки 2a-2d), сме-
щённый приблизительно на 45° влево (леотропно)
относительно макромеров, которые теперь обозна-
чаются 2A-2D. Первый квартет микромеров также
экваториально делится на два квартета. Клетки, ле-
жащие ближе к анимальному полюсу, обозначаются
верхним индексом 1 (la
1
—Id
1
), а те бластомеры, ко-
торые лежат ближе к вегетативному полюсу, — ин-
дексом 2 (la
2
-Id
2
) (илл. 287Б). Последние представ-
ляют собой первичные трохобласты, из которых
впоследствии развиваются ресничные клетки про-
тотроха. Таким образом, в результате 4-го деления
возникают 16 бластомеров.
Два последующих деления (дексиотропное и
леотропное) приводят к образованию ещё двух квар-
тетов микромеров, а именно 3a-3d после 5-го деле-
ния и 4a-4d после 6-го деления. На этой стадии син-
хронность дробления утрачивается: вначале делятся
клетки трохобласта, а позже всех — бластомер 3D,
который разделяется на 4D и 4d. После 6-го деления
дробления зародыш состоит из 64 клеток, дальней-
шая судьба которых строго детерминирована (илл.
288).
Очень важной в морфогенетическом отноше-
нии является клетка 4d, или протомезобласт, из ко-
торого возникает почти вся мезодерма. Бластомер 4d
делится равномерно на правую и левую первичные
мезодермальные клетки (4d' и 4d
2
), которые в ре-
зультате интенсивных делений формируют парные
мезодермальные полоски. Большое значение имеет
также клетка 2d, из которой в основном формирует-
ся эктодерма личинки. В состав ресничных колец мо-
гут входить, помимо первичных трохобластов, так-
же вторичные и дополнительные трохобласты (илл.
287Е).
Некоторые из них вторично утрачивают рес-
нички и превращаются в опорные клетки прототроха.
У Patella и многих других моллюсков все че-
тыре макромера имеют одинаковые размеры и делят-
ся синхронно, то есть отдельные квадранты эмбрио-
на морфологически неразличимы (гомоквадрантное
дробление). Эксперименты показали, что роль каж-
дого квадранта на данной стадии ещё не определе-
на, то есть квадранты являются эквипотентными.
Определение судьбы макромеров происходит толь-
ко после отделения 3-го квартета микромеров и осу-
ществляется благодаря взаимодействию клеток: тот
макромер, который первым вступает в контакт с мик-
ромерами, детерминируется как 3D. У других видов
моллюсков и большинства полихет уже первых два
деления являются неравномерными (гетероквадран-
тный эмбрион). Самый крупный макромер почти все-
гда дифференцируется как D. Детерминация макро-
меров определяется распределением цитоплазмати-
ческих факторов: те из них, которые имеют отноше-
ние к формированию бластомеров 2d и 4d, попада-
ют преимущественно в клетку D.
Значительное сходство спирального дробления,
наблюдаемое в различных таксонах, может, разуме-
ется, служить аргументом в пользу их филогенети-
ческой близости. Тем не менее, в каждом конкрет-
ном случае необходим тщательный анализ во избе-
жание возможных ошибок. При этом нужно учиты-
вать следующие соображения:
- С одной стороны, предположение о родствен-
ных отношениях между некоторыми группами, на-
пример между моллюсками и кольчатыми червями
или между немертинами и плоскими червями, ос-
новано только на наличии у них спирального дроб-
ления, в то время как анатомические и ультраструк-
турные исследования не выявляют значительного
сходства.
- С другой стороны, в пределах древних фило-
генетических линий, обладающих спиральным дроб-
лением, могут возникать группы с сильно изменён-
ным характером дробления. Примером таких групп
могут служить Neoophora в пределах Plathelminthes
или Cephalopoda в пределах Mollusca. Отсюда сле-
дует, что могут существовать группы, не имеющие
спирального дробления, которые, тем не менее, сле-
дует относить к Spiralia. Некоторые авторы даже
предполагают, что общий предок всех Bilateria обла-
дал спиральным дроблением.
- Кроме того, имеется много групп, в развитии
которых можно усмотреть «сходство» со спиральным
дроблением. Однако нельзя исключить, что отдель-
ные признаки спирального дробления, такие как на-
клонное положение веретён деления, были приобре-
тены конвергентно.
- Наконец, далеко не все опубликованные опи-
сания можно считать совершенно надёжными. В пос-
леднее время интерес к спиральному дроблению
вновь возрос в связи с изучением вопросов клеточ-
ной дифференцировки. Разработан ряд методов (в
частности, мечение клеток красителями и иммуно-
цитохимическая характеристика отдельных бласто-
меров), которые могут быть использованы для по-
вторного изучения спирального дробления. Можно
ожидать, что будущие исследования дадут обширный
материал для сравнительного анализа.
В настоящее время к Spiralia относят следую-
щие группы:
Annelida и Arthropoda вместе составляют мо-
нофилетическую группу Articulata, которая выделя-
ется на основании таких признаков, как целомичес-
кая сегментация, наличие преанальной зоны роста и
вентральной системы ганглиев в виде брюшной не-
рвной цепочки (или «нервной лестницы»). У боль-
http://jurassic.ru/
Spiralia 217
Polychaeta
(Arenicola)
Mollusca
(Trochus)
ct2 ct 4 CT 8 ct 16
ct 32
cr64
проспективное
ст64
ст32
ст
16
ст
8
ст
4 ст2
значение
апикальные
-
клетки розетки
базальные
клетки
креста
la"'
«периферические
1а"
клетки розетки»
промежуточные
f
1
а]*
клетки «базальные
1 ^> , /
клетки
креста»
' -
а
первичные
трохобласты
эктодерма
протонефридии
эктомезенхима
эктодерма
эктодерма
— 5,
эктодерма
эктодерма
соматобласт
эктодерма
протомезобласт
4d 2D
энтодерма
-
5d
3D
энтодерма
•
4D
Илл.
288. Соответствие проспективного з]
Polychaeta
(Arenicola)
Из Siewi
шинства видов Annelida, особенно
у,
Polychaeta, на-
блюдается типичное спиральное дробление (илл.
286);
такой же способ дробления, очевидно, был и у
общего предка Articulata. В эволюции членистоно-
гих спиральное дробление сменилось поверхност-
ным; случаи полного дробления с наклонным поло-
жением веретён деления, известные для некоторых
ракообразных («Cladocera», Cirripedia, Euphausiacea),
следует рассматривать как вторичные. Спиральное
дробление сильно видоизменяется также в некото-
рых группах «01igochaeta» и у Hirudinea (Clitellata).
чения клеток при спиральном дроблении
Gastropoda
(Trochus).
; (1969).
Echiura, которые, возможно, представляют со-
бой сестринскую группу Articulata, обладают типич-
ным спиральным дроблением.
Mollusca, возможно, являются ближайшими
родственниками группировки Articulata+Echiura. Это
предположение основано на значительном сходстве
в проспективном значении отдельных бластомеров
(илл. 288). Особо следует упомянуть значение блас-
томера 4d как протомезобласта, формирование боль-
шей части туловищной эктодермы из клетки 2d и
дифференцировку трохобластов. Дополнительным
http://jurassic.ru/

218
Трёхслойные Eumetazoa
аргументом служит явное сходство между трохофо-
рой полихет и личинками многих моллюсков. В пре-
делах моллюсков известны многочисленные случаи
модификации спирального дробления. Богатые жел-
тком яйца Cephalopoda претерпевают неполное дис-
коидальное дробление, причем зародышевый диск
исходно обладает билатеральной симметрией; таким
образом, у головоногих моллюсков исчезают все при-
знаки спирального дробления.
Для Sipuncula описано типичное квартетное
спиральное дробление с дифференцировкой мезодер-
мы из бластомера 4d.
Дробление Nemertini можно рассматривать как
спиральное, поскольку клетки располагаются квар-
тетами с характерным смещением. Однако анималь-
ные бластомеры у зародышей немертин обычно круп-
нее,
чем вегетативные, хотя и имеют большую ско-
рость деления. Развитие первичных мезодермальных
клеток из бластомера 4d для немертин не доказано.
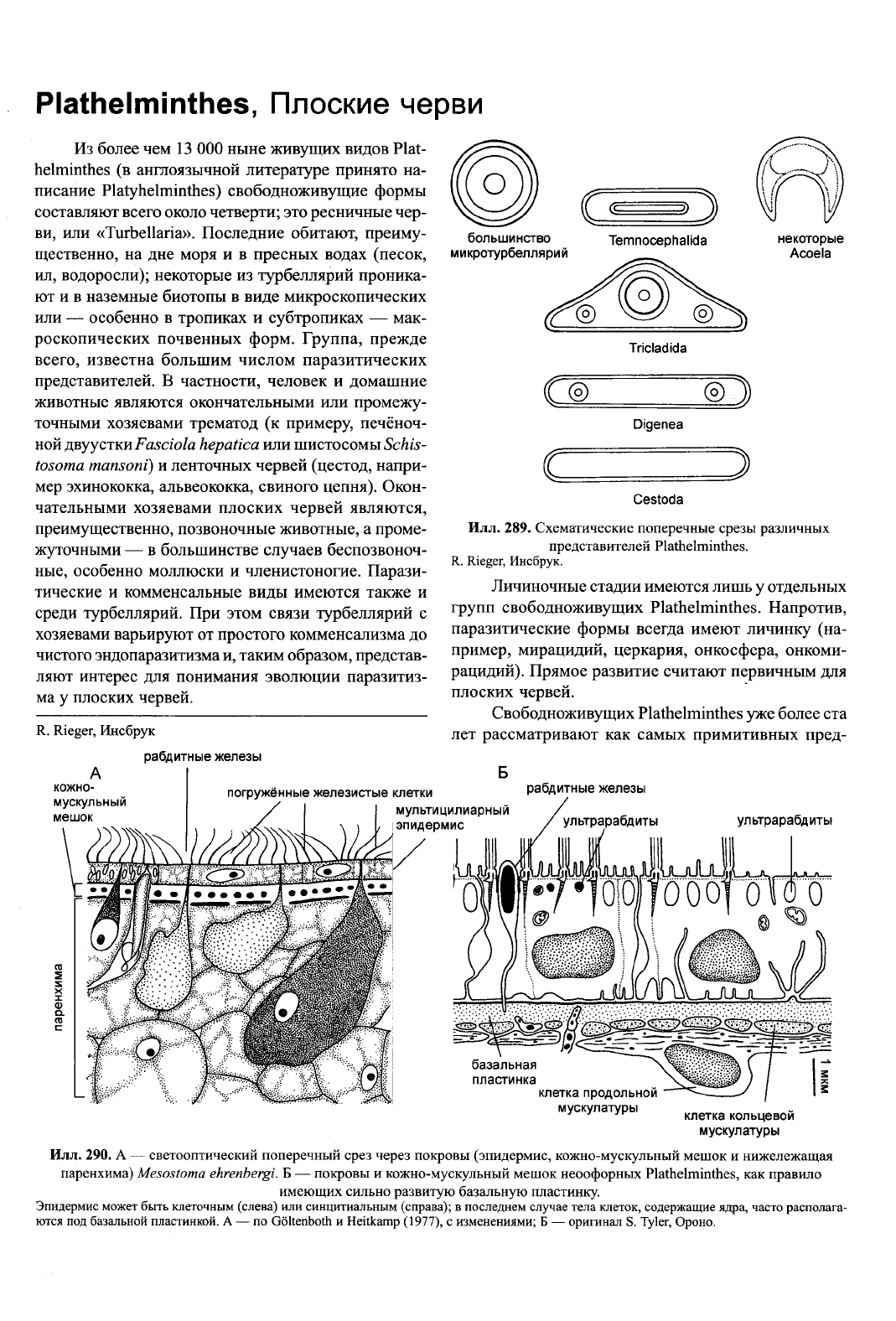
Plathelminthes — ещё одна группа, исходным
для которой считают спиральное дробление. Отчёт-
ливые признаки спирального дробления имеются
только в таксонах с примитивным эндолецитальным
способом формирования яйца («Archoophora»), при-
чём детально изучено только развитие Polycladida.
Имеющиеся сообщения противоречивы. По одним
наблюдениям,' бластомер 4d делится на две первич-
ные мезодермальные клетки, по мнению других ис-
следователей, из 4d образуется не только мезодерма,
но и вся энтодерма зародыша. У Acoela уже второе
деление дробления является экваториальным, поэто-
му формируются не квартеты, а дуэты бластомеров.
Относительно Gnathostomulida в литературе
имеется лишь краткое упоминание о том, что они,
вероятно, обладают спиральным дроблением. Дроб-
ление Kamptozoa также рассматривают как спираль-
ное.
Для некоторых групп животных характерны
«элементы» спирального дробления. В частности, у
Phoronida и Pogonophora образуются квартеты бла-
стомеров, которые, однако, расположены не так,
как при спиральном дроблении. Проспективное зна-
чение бластомеров в этих группах также не соответ-
ствует тому, что наблюдают у кольчатых червей и
моллюсков.
Таким образом, можно заключить, что суще-
ствует несколько групп с типичным спиральным
дроблением (Annelida, Echiura, Mollusca, Sipuncula).
Дробление в некоторых другие группах (Nemertini,
Plathelminthes, Gnathostomulida, Kamptozoa), очевид-
но,
проходит по спиральному типу, но его гомоло-
гию с дроблением кольчатых червей и моллюсков
пока нельзя считать доказанной. Наконец, элементы
спирального дробления, наблюдаемые в некоторых
группах (например, наклонное расположение вере-
тён деления у некоторых Crustacea или квартеты бла-
стомеров у Phoronida и Pogonophora), скорее всего,
возникли в результате конвергенции. Принадлеж-
ность ракообразных и погонофор к Spiralia обосно-
вана не характером дробления, а другими апоморф-
ными признаками.
http://jurassic.ru/
Plathelminthes,
Плоские черви
Из более чем 13
ООО
ныне живущих видов Plat-
helminthes (в англоязычной литературе принято на-
писание Platyhelminthes) свободноживущие формы
составляют всего около четверти; это ресничные чер-
ви,
или «Turbellaria». Последние обитают, преиму-
щественно, на дне моря и в пресных водах (песок,
ил,
водоросли); некоторые из турбеллярий проника-
ют и в наземные биотопы в виде микроскопических
или — особенно в тропиках и субтропиках — мак-
роскопических почвенных форм. Группа, прежде
всего, известна большим числом паразитических
представителей. В частности, человек и домашние
животные являются окончательными или промежу-
точными хозяевами трематод (к примеру, печёноч-
ной двуустки Fasciola hepatica или шистосомы Schis-
tosoma mansoni) и ленточных червей (цестод, напри-
мер эхинококка, альвеококка, свиного цепня). Окон-
чательными хозяевами плоских червей являются,
преимущественно, позвоночные животные, а проме-
жуточными — в большинстве случаев беспозвоноч-
ные,
особенно моллюски и членистоногие. Парази-
тические и комменсальные виды имеются также и
среди турбеллярий. При этом связи турбеллярий с
хозяевами варьируют от простого комменсализма до
чистого эндопаразитизма
и,
таким образом, представ-
ляют интерес для понимания эволюции паразитиз-
ма у плоских червей.
R. Rieger, Инсбрук
рабдитные железы
большинство
м и кроту
рбел
ляр
и й
Temnocephalida
некоторые
Acoela
Tricladida
сж
Digenea
Cestoda
Илл.
289. Схематические поперечные срезы различных
представителей Plathelminthes.
R. Rieger, Инсбрук.
Личиночные стадии имеются лишь у отдельных
групп свободноживущих Plathelminthes. Напротив,
паразитические формы всегда имеют личинку (на-
пример, мирацидий, церкария, онкосфера, онкоми-
рацидий). Прямое развитие считают первичным для
плоских червей.
Свободноживущих Plathelminthes уже более ста
лет рассматривают как самых примитивных пред-
А
кожно-
мускульный
мешок
погружённые железистые клетки
мультицилиарный
рабдитные железы
/
ультрарабдиты
ультрарабдиты
клетка продольной
мускулатуры
клетка кольцевой
мускулатуры
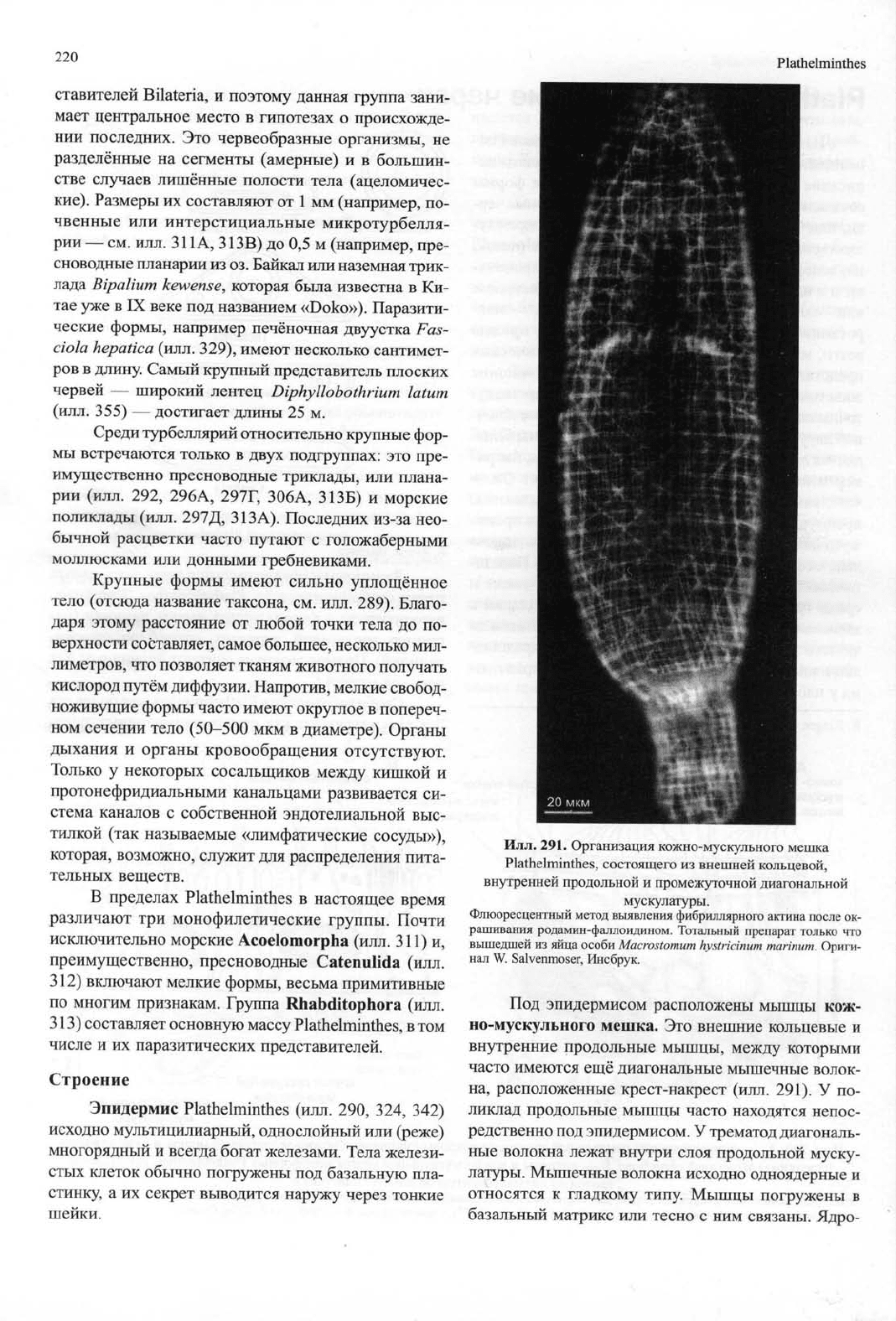
Илл.
290. А — светооптический поперечный срез через покровы (эпидермис, кожно-мускульный мешок и нижележащая
паренхима)
Mesostoma
ehrenbergi.
Б — покровы и кожно-мускульный мешок неоофорных Plathelminthes, как правило
имеющих сильно развитую базальную пластинку.
Эпидермис может быть клеточным (слева) или синцитиальным (справа); в последнем случае тела клеток, содержащие ядра, часто располага-
ются под базальной пластинкой. А — по Goltenboth и Heitkamp (1977), с изменениями; Б — оригинал S. Tyler, Ороно.
http://jurassic.ru/
220
Plathelminthes
Илл.
291. Организация кожно-мускульного мешка
Plathelminthes, состоящего из внешней кольцевой,
внутренней продольной и промежуточной диагональной
мускулатуры.
Флюоресцентный метод выявления фибриллярного актина после ок-
рашивания родамин-фаллоидином. Тотальный препарат только что
вышедшей из яйца особи Macrostomum hystricinum marinum. Ориги-
нал W. Salvenmoser, Инсбрук.
Под эпидермисом расположены мышцы кож-
но-мускульного мешка. Это внешние кольцевые и
внутренние продольные мышцы, между которыми
часто имеются ещё диагональные мышечные волок-
на, расположенные крест-накрест (илл. 291). У по-
ликлад продольные мышцы часто находятся непос-
редственно под эпидермисом. У трематод диагональ-
ные волокна лежат внутри слоя продольной муску-
латуры. Мышечные волокна исходно одноядерные и
относятся к гладкому типу. Мышцы погружены в
базальный матрикс или тесно с ним связаны. Ядро-
ставителей Bilateria, и поэтому данная группа зани-
мает центральное место в гипотезах о происхожде-
нии последних. Это червеобразные организмы, не
разделённые на сегменты (амерные) и в большин-
стве случаев лишённые полости тела (ацеломичес-
кие).
Размеры их составляют от
1
мм (например, по-
чвенные или интерстициальные микротурбелля-
рии — см. илл. 311А, 313В) до 0,5 м (например, пре-
сноводные планарии из
оз.
Байкал или наземная трик-
лада Bipalium kewense, которая была известна в Ки-
тае уже в IX веке под названием «Doko»). Паразити-
ческие формы, например печёночная двуустка Fas-
ciola hepatica (илл. 329), имеют несколько сантимет-
ров в длину. Самый крупный представитель плоских
червей — широкий лентец Diphyllobothrium latum
(илл. 355) — достигает длины 25 м.
Среди турбеллярий относительно крупные фор-
мы встречаются только в двух подгруппах: это пре-
имущественно пресноводные триклады, или плана-
рии (илл. 292, 296А, 297Г, 306А, 313Б) и морские
поликлады (илл. 297Д, 313А). Последних из-за нео-
бычной расцветки часто путают с голожаберными
моллюсками или донными гребневиками.
Крупные формы имеют сильно уплощённое
тело (отсюда название таксона, см. илл. 289). Благо-
даря этому расстояние от любой точки тела до по-
верхности составляет, самое большее, несколько мил-
лиметров, что позволяет тканям животного получать
кислород путём диффузии. Напротив, мелкие свобод-
ноживущие формы часто имеют округлое в попереч-
ном сечении тело (50-500 мкм в диаметре). Органы
дыхания и органы кровообращения отсутствуют.
Только у некоторых сосальщиков между кишкой и
протонефридиальными канальцами развивается си-
стема каналов с собственной эндотелиальной выс-
тилкой (так называемые «лимфатические сосуды»),
которая, возможно, служит для распределения пита-
тельных веществ.
В пределах Plathelminthes в настоящее время
различают три монофилетические группы. Почти
исключительно морские Acoelomorpha (илл. 311) и,
преимущественно, пресноводные Catenulida (илл.
312) включают мелкие формы, весьма примитивные
по многим признакам. Группа Rhabditophora (илл.
313) составляет основную массу Plathelminthes, в том
числе и их паразитических представителей.
Строение
Эпидермис Plathelminthes (илл. 290, 324, 342)
исходно мультицилиарный, однослойный или (реже)
многорядный и всегда богат железами. Тела желези-
стых клеток обычно погружены под базальную пла-
стинку, а их секрет выводится наружу через тонкие
шейки.
http://jurassic.ru/
эпидермис
с
рабдитами
дорсолатеральные
комиссуры
спинной
продольный
нервный
ствол
кольцевая
мускулатура
кишка
фолликул
семенника
фолликул
желточника
продольная
мускулатура
брюшная
комиссура
брюшной
продольный
нервный ствол
мускулатура
паренхимы
боковое
утолщение
эпидермиса
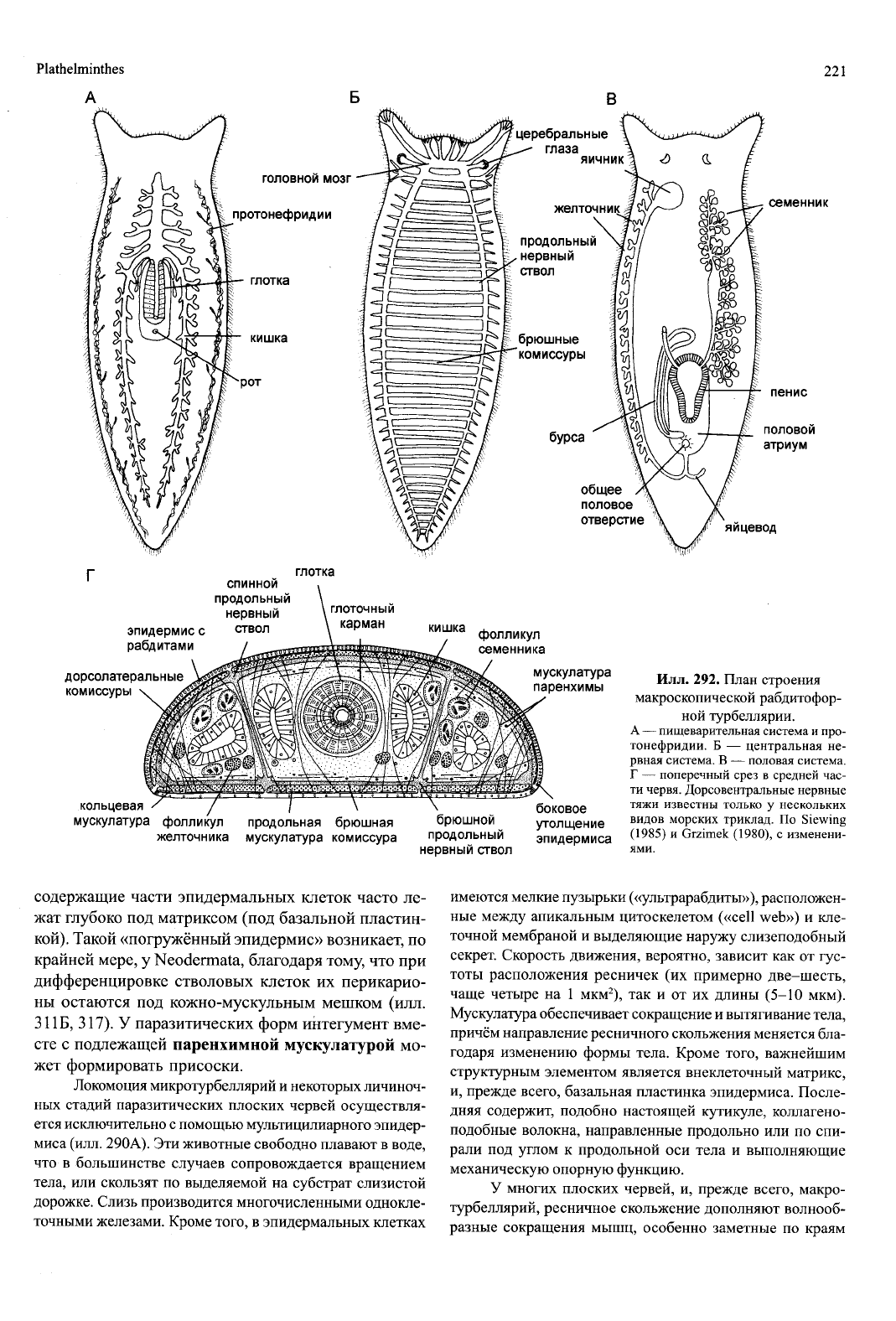
Илл.
292.
План строения
макроскопической рабдитофор-
ной турбеллярий.
А
—
пищеварительная система
и
про-
тонефридии.
Б —
центральная
не-
рвная система.
В —
половая система.
Г
—
поперечный срез
в
средней час-
ти червя. Дорсовентральные нервные
тяжи известны только
у
нескольких
видов морских триклад.
По
Slewing
(1985)
и
Grzimek (1980),
с
изменени-
содержащие части эпидермальных клеток часто ле-
жат глубоко под матриксом (под базальной пластин-
кой).
Такой «погружённый эпидермис» возникает, по
крайней мере, у Neodermata, благодаря тому, что при
дифференцировке стволовых клеток их перикарио-
ны остаются под кожно-мускульным мешком (илл.
311Б,
317). У паразитических форм интегумент вме-
сте с подлежащей паренхимной мускулатурой мо-
жет формировать присоски.
Локомоция микротурбеллярий
и
некоторых личиноч-
ных стадий паразитических плоских червей осуществля-
ется исключительно с помощью мультицилиарного эпидер-
миса (илл. 290А). Эти животные свободно плавают
в
воде,
что
в
большинстве случаев сопровождается вращением
тела,
или
скользят
по
выделяемой
на
субстрат слизистой
дорожке. Слизь производится многочисленными однокле-
точными железами. Кроме того, в эпидермальных клетках
имеются мелкие пузырьки («ультрарабдиты»), расположен-
ные между апикальным цитоскелетом («cell web»)
и
кле-
точной мембраной
и
выделяющие наружу слизеподобный
секрет. Скорость движения, вероятно, зависит как
от
гус-
тоты расположения ресничек
(их
примерно две-шесть,
чаще четыре
на 1
мкм
2
),
так и от их
длины
(5-10 мкм).
Мускулатура обеспечивает сокращение и вытягивание тела,
причём направление ресничного скольжения меняется бла-
годаря изменению формы тела. Кроме того, важнейшим
структурным элементом является внеклеточный матрикс,
и, прежде всего, базальная пластинка эпидермиса. После-
дняя содержит, подобно настоящей кутикуле, коллагено-
подобные волокна, направленные продольно
или по
спи-
рали
под
углом
к
продольной
оси
тела
и
выполняющие
механическую опорную функцию.
У многих плоских червей,
и,
прежде всего, макро-
турбеллярий, ресничное скольжение дополняют волнооб-
разные сокращения мышц, особенно заметные
по
краям
http://jurassic.ru/

222
Plathelminthes
Илл.
293. Строение нервной системы Plathelminthes.
А — периферический сенсорный плексус Macrostomum pusillum как пример примитивного состояния, сходного с тем, которое наблюдается у
«Coelenterata». Б — нервная система Proseriata, построенная по типу ортогона. В — фрагмент ортогона на брюшной стороне тела Bothriomolus
balticus. А, В — иммуноцитохимическое окрашивание серотонинэргических нейронов. А — оригинал W. Salvenmoser и P. Ladurner, Инсбрук;
Б — по Reisinger (1972); В — по Joffe и Reuter (1993).
тела
(илл.
313А). Паразитические формы часто передвига-
ются только при помощи мышц. Личинки трематод (цер-
карии) плавают благодаря работе мускулистого хвоста;
данный способ локомоции не встречается у других пред-
ставителей Plathelminthes.
Увеличение числа эпидермальных клеток
(включая и железистые) происходит, как и во всех
других тканях, за счёт недифференцированных, так
называемых стволовых клеток (необластов). Необла-
сты почти всегда располагаются в паренхиме и отту-
да мигрируют в эпидермис. Данные клетки хорошо
известны во всех группах, кроме Acoelomorpha. Нео-
бласты являются также основным источником фор-
мирования новых покровов (неодермиса) после сбра-
сывания личиночного эпидермиса в развитии пара-
зитических Plathelminthes, которые по этой причине
обозначаются как Neodermata.
Интересно, что в пределах Plathelminthes край-
не редко встречаются настоящие твёрдые кутикуляр-
ные образования (например, у Temnocephalida). Твёр-
дые склеротизированные элементы часто присутству-
ют в составе эпидермиса и органов размножения у
некоторых червей. Однако они представляют собой
либо уплотнения базальной пластинки (ср. крючья
хобота скребней), либо внутриклеточные образова-
ния («псевдокутикула»; ср. панцири некоторых ко-
ловраток).
Нервная система плоских червей (за исклю-
чением некоторых Acoelomorpha) состоит из голов-
ного мозга, погружённого под кожно-мускульный
мешок (илл. 292Б, 293Б, 294), от которого отходит
некоторое число одетых оболочкой продольных
стволов. Последние часто связаны поперечными ко-
миссурами, образуя характерную прямоугольную ре-
шетку — ортогон (илл. 293Б,В). Ортогон складыва-
ется из локальных сгущений нервного плексуса, рас-
положенного внутри мышечных слоев или под ними.
Помимо ортогона, может присутствовать эпидер-
мальный и субэпидермальный плексус — сетчатое
или войлокообразное переплетение отростков ней-
ронов (илл. 293А). Эти нейроны образуют перифе-
рическую нервную систему, которая связана волок-
нами с головным мозгом и продольными стволами
ортогона (центральной нервной системой).
Большинство Plathelminthes, включая, как ни
удивительно, и паразитические формы, имеют бога-
то развитые органы чувств и ресничные рецепто-
ры (воротничковые рецепторы, сенсорные реснич-
ки) самой разнообразной ультраструктуры (илл. 196).
Широко распространены несколько типов глаз, уст-
роенных по схеме «пигментных бокалов» (илл.
296),
и статоцисты (илл. 311А).
Пищеварительная система в некоторых пара-
зитических группах полностью редуцирована (напри-
мер,
Cestoda, Fecampiida); в этих случаях питатель-
ные вещества поглощает специализированная повер-
хность тела, т.е. это осуществляется «парентераль-
но».
Особым способом получения питательных ве-
ществ отличаются виды, имеющие одноклеточных
симбионтов. В обычном же случае пища попадает в
кишку через мускулистую и богатую железами экто-
дермальную глотку. Анальное отверстие почти все-
гда отсутствует. Характер разветвления кишки и раз-
личные морфологические типы глотки (простое ро-
http://jurassic.ru/

Plathelminthes
223
ганглиозные
клетки
вне
капсулы
головного
мозга
Илл.
294. А — реконструкция головного
мозга, брюшных (толстые) и спинных
(тонкие) продольных нервных стволов
Notoplana. Размер головного мозга
несколько мм. Б — поперечный срез
головного мозга Kythorhynchus sp.,
демонстрирующий периферичесое
расположение клеточных тел нейронов.
А — по Chien и Koopowitz (1972); Б — ориги-
нал G. Rieger, Инсбрук.
товое отверстие, pharynx simplex, pharynx plicatus,
pharynx bulbosus, илл. 298) составляют важные ди-
агностические признаки. Pharynx bulbosus встреча-
ется также у паразитических форм, поскольку дале-
ко не все они лишены ротового отверстия и кишки.
Имеется как внеклеточное, так и внутриклеточное
пищеварение, в связи с чем в стенках кишки присут-
ствуют и фагоцитирующие клетки, и экзокринные
железы (илл. 189Г). Кишка, подобно гастроваскуляр-
ной системе кишечнополостных, служит также для
распределения питательных веществ (илл. 297). Осо-
бым строением пищеварительного аппарата облада-
ют бескишечные турбеллярий (илл. 298А).
Большинство свободноживущих форм — хищ-
ники. Многие микротурбеллярии и некоторые поли-
клады питаются водорослями (прежде всего диато-
мовыми).
Некоторые виды, имеющие симбионтов из числа зе-
лёных водорослей, диатомей или динофлагеллят, во взрос-
лом состоянии
не
питаются.
Однако для большинства форм
А
VI/
л
V
л
д
Илл.
295. Реакции планарий на струю воды из пипетки.
Раздражение заднего конца (А) или середины тела (Б) не вызывает
никакой реакции; (В) раздражение аурикул («ушей»), (Г) реакция на
(В);
(Д) раздражение сзади по продольной оси животного, (Е) реак-
ция на (Д), после того как струя достигает «ушей». По Doflein (1925)
из Bresslau (1927-33).
с автотрофными симбионтами, вероятно, характерен сме-
шанный
тип
питания.
Широко распространена способность
переживать длительные периоды без пищи, что сопровож-
дается уменьшением размеров тела (явление «обратного
роста», или «degrowth»).
С гистологической точки зрения пространство
между кишкой и покровами можно рассматривать как
первичную полость тела. У микротурбеллярии, сре-
ди которых, вероятно, находятся и самые примитив-
ные представители Plathelminthes, полость тела мо-
жет быть практически вытеснена кишкой (илл.
299А),
но может присутствовать даже в виде более
или менее обширного псевдоцеля (некоторые Cate-
nulida, илл. 299Б).
Почти все Plathelminthes — ацеломические
формы. Это означает, что пространство между ба-
зальными пластинками эпидермиса и гастродерми-
са заполнено паренхимой, в которой лежат органы
выделения и осморегуляции, а также половая систе-
ма (илл. 299В). Соединительная ткань состоит в боль-
шинстве случаев из межклеточного матрикса и кле-
ток различных типов (необласты, мышечные клет-
ки,
клетки паренхимы). В пространстве между эпи-
дермисом и гастродермисом могут также лежать по-
гружённые тела клеток эпидермиса (либо неодер-
миса).
Цитоплазма всех этих клеток, за исключением
регенеративных необластов (илл. 299А), играет роль
внутриклеточного гидростатического скелета. Вме-
сте с кишкой и межклеточным матриксом (в частно-
сти,
базальной пластинкой эпидермиса) клетки дей-
ствуют как антагонисты мускулатуры (кожно-мус-
кульного мешка и паренхимных мышц), что и обес-
печивает поддержание и изменение видоспецифич-
ной формы тела.
http://jurassic.ru/
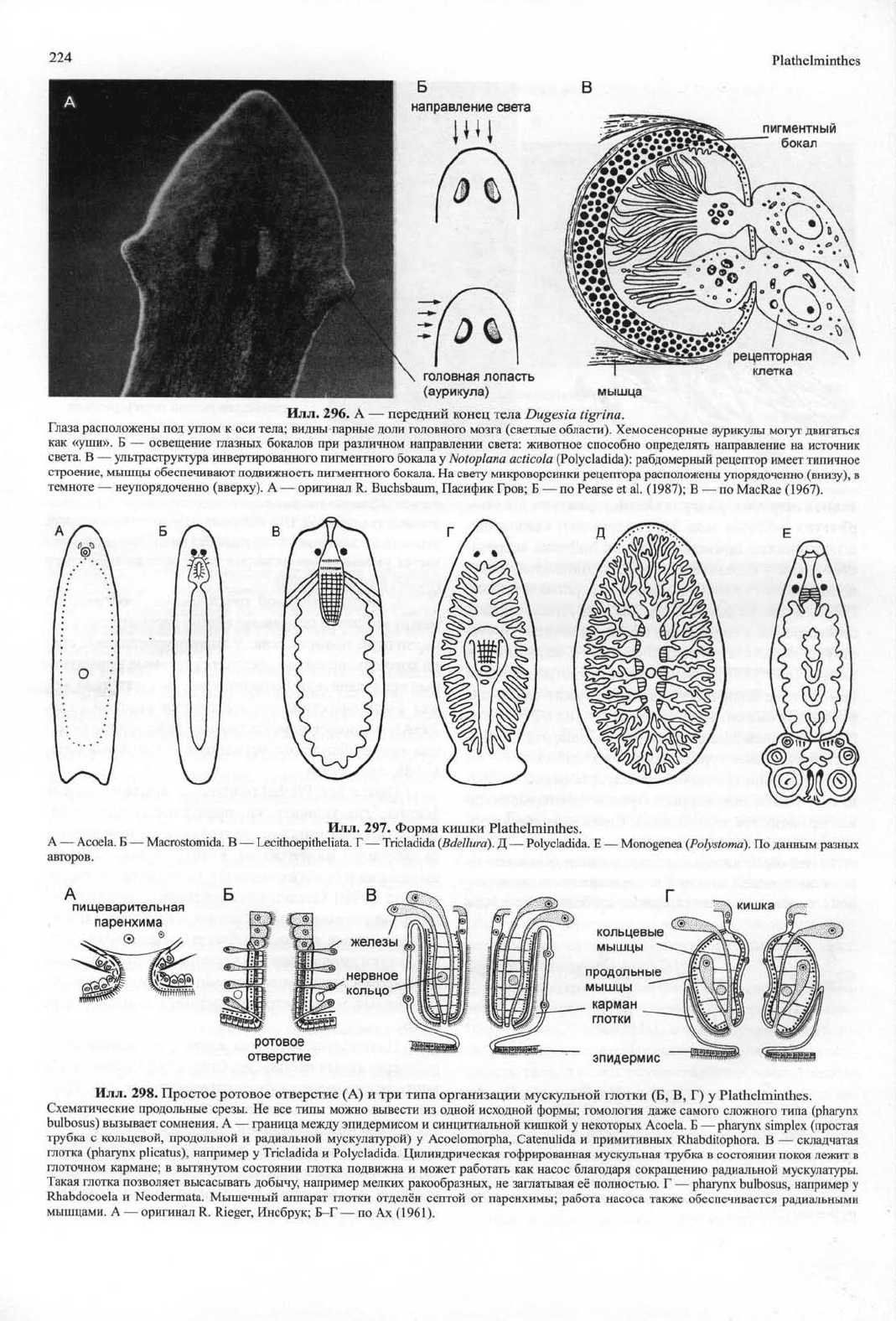
224
Plathelminthes
Илл.
296. A — передний конец тела Dugesia tigrina.
Глаза расположены под углом к оси тела; видны парные доли головного мозга (светлые области). Хемосенсорные аурикулы могут двигаться
как «уши». Б — освещение глазных бокалов при различном направлении света: животное способно определять направление на источник
света. В — ультраструктура инвертированного пигментного бокала у Notoplana acticola (Polycladida): рабдомерный рецептор имеет типичное
строение, мышцы обеспечивают подвижность пигментного бокала. На свету микроворсинки рецептора расположены упорядочение (внизу), в
темноте — неупорядоченно (вверху). А — оригинал R. Buchsbaum, Пасифик Гров; Б — по Pearse et al. (1987); В — по MacRae (1967).
Илл.
297. Форма кишки Plathelminthes.
А — Acoela. Б — Macrostomida. В — Lecithoepitheliata. Г — Tricladida (Bdellura). Д — Polycladida. Е — Monogenea (Polystomd). По данным разных
авторов.
Илл.
298. Простое ротовое отверстие (А) и три типа организации мускульной глотки (Б, В, Г) у Plathelminthes.
Схематические продольные срезы. Не все типы можно вывести из одной исходной формы; гомология даже самого сложного типа (pharynx
bulbosus) вызывает сомнения. А — граница между эпидермисом и синцитиальной кишкой у некоторых Acoela. Б — pharynx simplex (простая
трубка с кольцевой, продольной и радиальной мускулатурой) у Acoelomorpha, Catenulida и примитивных Rhabditophora. В — складчатая
глотка (pharynx plicatus), например у Tricladida и Polycladida. Цилиндрическая гофрированная мускульная трубка в состоянии покоя лежит в
глоточном кармане; в вытянутом состоянии глотка подвижна и может работать как насос благодаря сокращению радиальной мускулатуры.
Такая глотка позволяет высасывать добычу, например мелких ракообразных, не заглатывая её полностью. Г — pharynx bulbosus, например у
Rhabdocoela и Neodermata. Мышечный аппарат глотки отделён септой от паренхимы; работа насоса также обеспечивается радиальными
мышцами. А — оригинал R. Rieger, Инсбрук; Б-Г — по Ах (1961).
http://jurassic.ru/
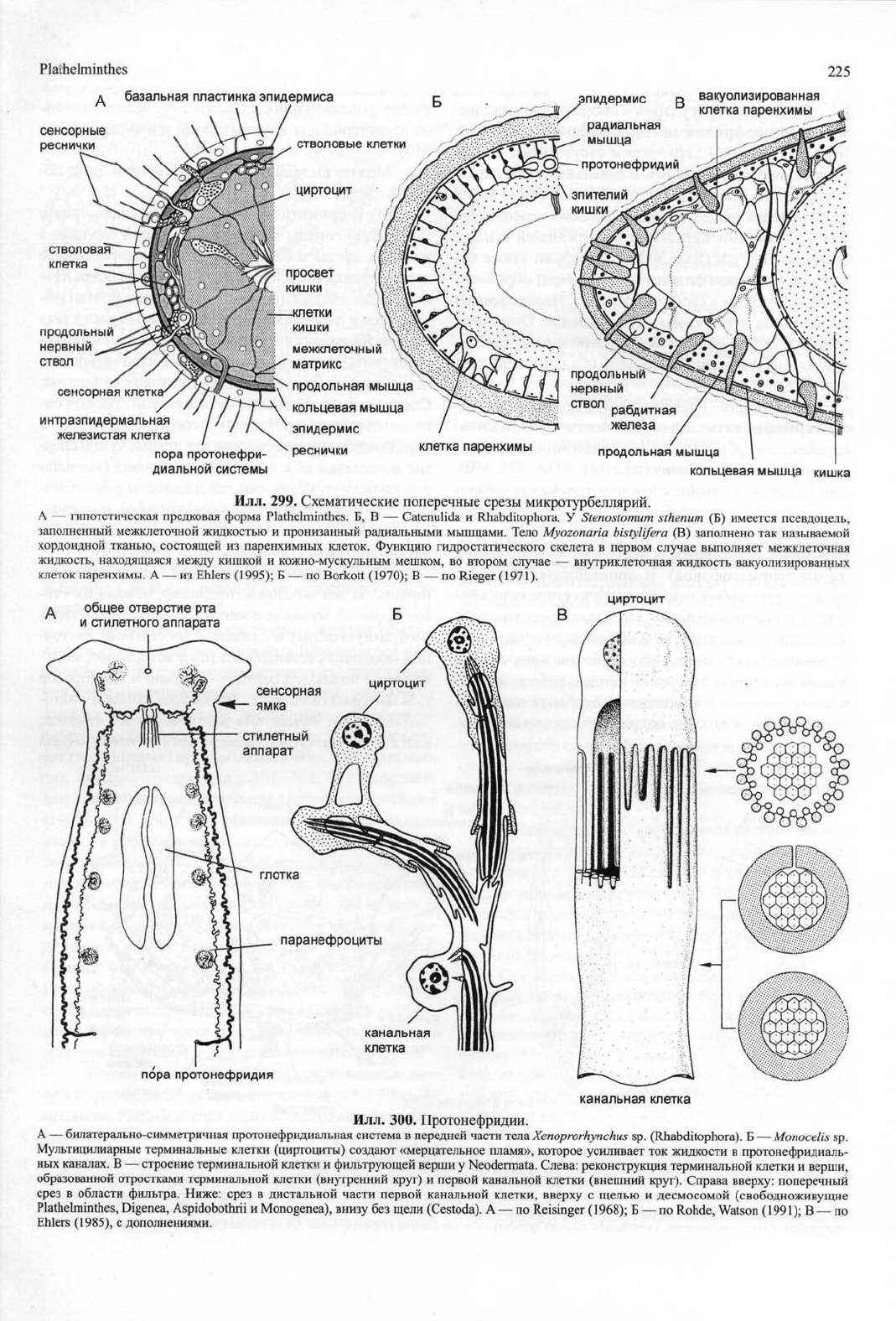
Plaihelminthes
225
базальная пластинка эпидермиса
сенсорные
реснички
пидермис
вакуолизированная
стволовые
клетки
циртоцит
продольный
нервный
ствол
сенсорная клетк
интраэпидермальная
железистая
клетка
пора протонефри-
диальной системы
межклеточный
матрикс
продольная мышца
кольцевая мышца
эпидермис
реснички
клетка паренхимы
продольная мышца
кольцевая мышца кишка
Илл.
299. Схематические поперечные срезы микротурбеллярии.
А — гипотетическая предковая форма Plathelminthes. Б, В — Catenulida и Rhabditophora. У Stenostomum sthenum (Б) имеется псевдоцель,
заполненный межклеточной жидкостью и пронизанный радиальными мышцами. Тело Myozonaria bistylifera (В) заполнено так называемой
хордоидной тканью, состоящей из паренхимных клеток. Функцию гидростатического скелета в первом случае выполняет межклеточная
жидкость, находящаяся между кишкой и кожно-мускульным мешком, во втором случае — внутриклеточная жидкость вакуолизированных
клеток паренхимы. А — из Ehlers (1995); Б — по Borkott (1970); В — по Rieger (1971).
общее
отверстие
рта
и стилетного
аппарата
циртоцит
пора протонефридия
канальная клетка
Илл.
300. Протонефридии.
А — билатерально-симметричная протонефридиальная система в передней части тела Xenoprorhynchus sp. (Rhabditophora). Б — Monocelis sp.
Мультицилиарные терминальные клетки (циртоциты) создают «мерцательное пламя», которое усиливает ток жидкости в протонефридиаль-
ных каналах. В — строение терминальной клетки и фильтрующей верши у Neodermata. Слева: реконструкция терминальной клетки и верши,
образованной отростками терминальной клетки (внутренний круг) и первой канальной клетки (внешний круг). Справа вверху: поперечный
срез в области фильтра. Ниже: срез в дистальной части первой канальной клетки, вверху с щелью и десмосомой (свободноживущие
Plathelminthes, Digenea, Aspidobothrii и Monogenea), внизу без щели (Cestoda). А — по Reisinger (1968); Б — по Rohde, Watson (1991); В — по
Ehlers (1985), с дополнениями.
http://jurassic.ru/