Вестхайде В.,Ригер Р.( ред.) Зоология беспозвоночных в двух томах. Том 1: от простейших до моллюсков и артропод
Подождите немного. Документ загружается.

196
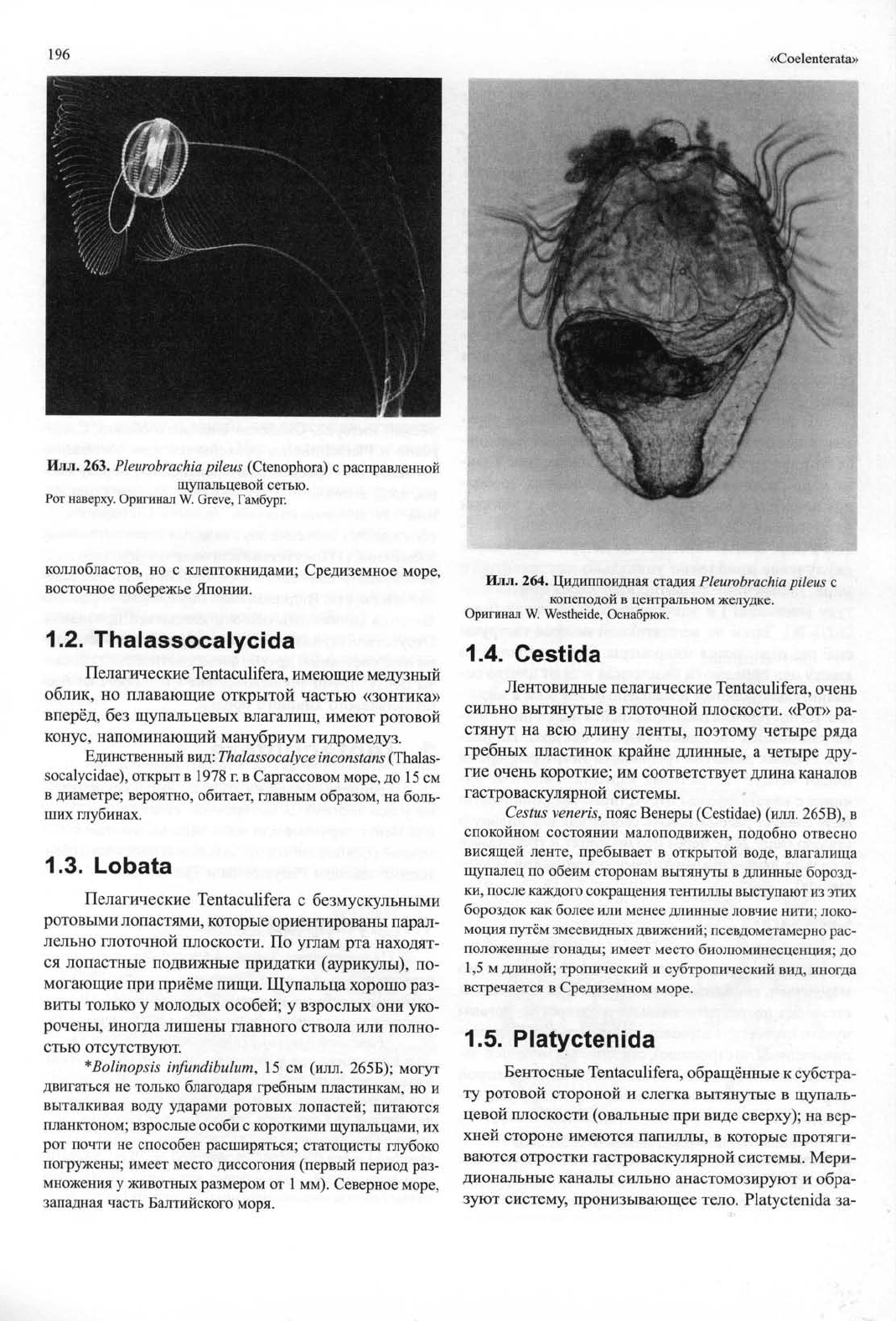
Илл.
263.
Pleurobrachia pileus (Ctenophora) с
расправленной
щупальцевой сетью.
Рот наверху. Оригинал W. Greve, Гамбург.
коллобластов, но с клептокнидами; Средиземное море,
восточное побережье Японии.
1.2.
Thalassocalycida
Пелагические Tentaculifera, имеющие медузный
облик, но плавающие открытой частью «зонтика»
вперёд, без щупальцевых влагалищ, имеют ротовой
конус, напоминающий манубриум гидромедуз.
Единственный
вид:
Thalassocalyce inconstans (Thalas-
socalycidae), открыт
в
1978 г. в Саргассовом море, до 15 см
в диаметре; вероятно, обитает, главным образом, на боль-
ших глубинах.
1.3.
Lobata
Пелагические Tentaculifera с безмускульными
ротовыми лопастями, которые ориентированы парал-
лельно глоточной плоскости. По углам рта находят-
ся лопастные подвижные придатки (аурикулы), по-
могающие при приёме пищи. Щупальца хорошо раз-
виты только у молодых особей; у взрослых они уко-
рочены, иногда лишены главного ствола или полно-
стью отсутствуют.
*Bolinopsis infundibulum, 15 см (илл. 265Б); могут
двигаться не только благодаря гребным пластинкам, но и
выталкивая воду ударами ротовых лопастей; питаются
планктоном; взрослые особи с короткими щупальцами, их
рот почти не способен расширяться; статоцисты глубоко
погружены; имеет место диссогония (первый период раз-
множения у животных размером от
1
мм). Северное море,
западная часть Балтийского моря.
«Coelenterata»
Илл.
264.
Цидиппоидная стадия Pleurobrachia pileus с
копеподой в центральном желудке.
Оригинал W. Westheide, Оснабрюк.
1.4.
Cestida
Лентовидные пелагические Tentaculifera, очень
сильно вытянутые в глоточной плоскости. «Рот» ра-
стянут на всю длину ленты, поэтому четыре ряда
гребных пластинок крайне длинные, а четыре дру-
гие очень короткие; им соответствует длина каналов
гастроваскулярной системы.
Cestus veneris, пояс Венеры (Cestidae) (илл. 265В), в
спокойном состоянии малоподвижен, подобно отвесно
висящей ленте, пребывает в открытой воде, влагалища
щупалец по обеим сторонам вытянуты в длинные борозд-
ки,
после каждого сокращения тентиллы выступают из этих
бороздок как более или менее длинные ловчие нити; локо-
моция путём змеевидных движений; псевдометамерно рас-
положенные гонады; имеет место биолюминесценция; до
1,5 м длиной; тропический и субтропический вид, иногда
встречается в Средиземном море.
1.5.
Platyctenida
Бентосные Tentaculifera, обращенные к субстра-
ту ротовой стороной и слегка вытянутые в щупаль-
цевой плоскости (овальные при виде сверху); на вер-
хней стороне имеются папиллы, в которые протяги-
ваются отростки гастроваскулярной системы. Мери-
диональные каналы сильно анастомозируют и обра-
зуют систему, пронизывающее тело. Platyctenida за-
http://jurassic.ru/

«Coelenterata»
197
Илл.
265.
Ctenophora. Общий вид.
А — Beroe sp; Б — Bolinopsis sp., моло-
дая особь. В — Cestus veneris. Г —
Coeloplana sp. По данным разным авто-
рам.
ботятся о потомстве, прикрывая подошвой яйца в
слизистых оболочках и защищая их.
Ctenoplana kowalevskii (Ctenoplanidae), может мед-
ленно плавать благодаря ундулирующим движениям краёв
тела, до 8 мм длиной; обитатель морского шельфа, Новая
Гвинея. — Виды Coeloplana часто живут на кораллах и
препятствуют разрастанию полипов (илл. 265Д). Coelopla-
na bocki (Coeloplanidae) размером 6 см встречается у вос-
точного берега Японии (о теории целопланы см. выше).
1.6.
Tjalfiellida
Эти гребневики бентосные, но не ползающие
(в отличие от Platyctenida), а сидячие. Тело вытянуто
в щупальцевой плоскости, а оральная сторона пре-
образована в прикрепительный диск. Верхняя сто-
рона с двумя вторично появившимися отверстиями
на двух выступах, торчащих вверх наподобие дымо-
вых труб, которые связаны со ртом и из которых вы-
ступают щупальца.
Tjaljiella tristoma (Tjalfiellidae), глубоководная фор-
ма (500 м) с заботой о потомстве (яйца развиваются в вы-
водковых сумках); размер 7 мм; живёт на Umbellula (Реп-
natularia), северные моря (Западная Гренландия).
2. Atentaculata
(Beroida)
Небольшая группа бесщупальцевых Ctenopho-
ra, ведущих только пелагический образ жизни. Очень
растяжимая глотка ведет в сложную, сильно развет-
влённую гастроваскулярную систему с множеством
слепо замкнутых каналов. Щупальца и коллобласты
отсутствуют; глотка занимает большую часть объе-
ма тела и несёт «крючковидные щетинки» (вероят-
но,
для удерживания добычи). Растяжимое мышеч-
ное тело напоминает мешок, который при плавании
ртом вперёд заглатывает жертву, достигающую (а
иногда — превосходящую) его по размерам. Пита-
ется преимущественно макропланктоном, особенно
студенистыми организмами (сальпами, гидромедуза-
ми,
щупальцевыми гребневиками).
Виды Вегое (морские огурцы) распространены все-
светно (илл. 265А). — *Beroe cucumis (Beroidae), 16 см
длиной, космополитический вид. — *Beroe gracilis (Вего-
idea),
3,5 см, эндемик юго-восточной части Северного моря.
Примечательны отношения хищник-жертва с другими
гребневиками: В. gracilis охотится на Pleurobrachia pileus,
в то время как пищей для В. cucumis служит Bolinopsis in-
fundibulum.
http://jurassic.ru/
Трёхслойные
Eumetazoa,
или
Bilateria
Образ
жизни, локомоция,
цефализация
и
симметрия
Среди ныне живущих Bilateria преобладают
одиночные подвижные формы, активно добывающие
пищу. Передний отдел тела, играющий особенно важ-
ную роль в процессах локомоции и питания, часто
обособляется в виде головной части, в которой рас-
полагается интегрирующий центр нервной систе-
мы — церебральный ганглий или головной мозг. Это
явление, обозначаемое как цефализация, наблюдает-
ся в разных филогенетических линиях Bilateria.
Колониальная
и/или
клональная организация
среди Bilateria также встречается в нескольких груп-
пах сидячих организмов (например, Kamptozoa, Bryo-
zoa, Pterobranchia, Ascidiacea). Такие формы обычно
питаются мелкими частицами, добывая их путём
фильтрации.
Все представители Bilateria, по крайней мере,
на личиночной стадии, обладают двусторонней (би-
латеральной) симметрией. Это означает, что правая
и левая половины тела (и только они!) зеркально со-
ответствуют друг другу (илл. 266). Единственная
плоскость симметрии (срединная, или сагиттальная
плоскость) проходит сверху вниз через передний и
задний концы тела. Направление движения живот-
ного
в
большинстве случаев (за исключением, напри-
мер,
ресничных личинок и сидячих форм) совпадает
с этой плоскостью. При локомоции тело, как прави-
ло,
обращено к субстрату определённой стороной,
которую обозначают как вентральную, или брюш-
Reinhard
Rieger,
Инсбрук
ную,
она часто несет специализированные локомо-
торные структуры (конечности, ползательную подо-
шву и т.п.). Сторону тела, противоположную вент-
ральной, называют дорсальной, или спинной. Таким
образом, имеются три взаимно перпендикулярные
плоскости. Сагиттальная плоскость разделяет тело
животного на правую и левую части, трансверсаль-
ная или поперечная плоскость (плоскость попереч-
ного среза) — на переднюю и заднюю части, а гори-
зонтальная (фронтальная) плоскость — на спинную
и брюшную части.
В некоторых группах исходная билатеральная
симметрия взрослых животных нарушена. Например,
ось движения каракатиц морфологически соответ-
ствует не передне-задней, а дорсовентральной оси;
взрослые особи иглокожих вторично приобретают
радиальную симметрию.
Первичноротые
и
вторичноротые формы
Интересно, что в развитии Bilateria ротовое от-
верстие возникает различными способами. Уже в
начале XX века было обнаружено (Grobben, 1908),
что бластопор (первичный рот) гаструлы в одних
случаях превращается в дефинитивный рот (прото-
апикальныи
базальный
сагиттальный дорсальный
вентральный
срединная
сагиттальная
плоскость
фронтальная,
или
горизонтальная
плоскость
дорсальный (спинной)
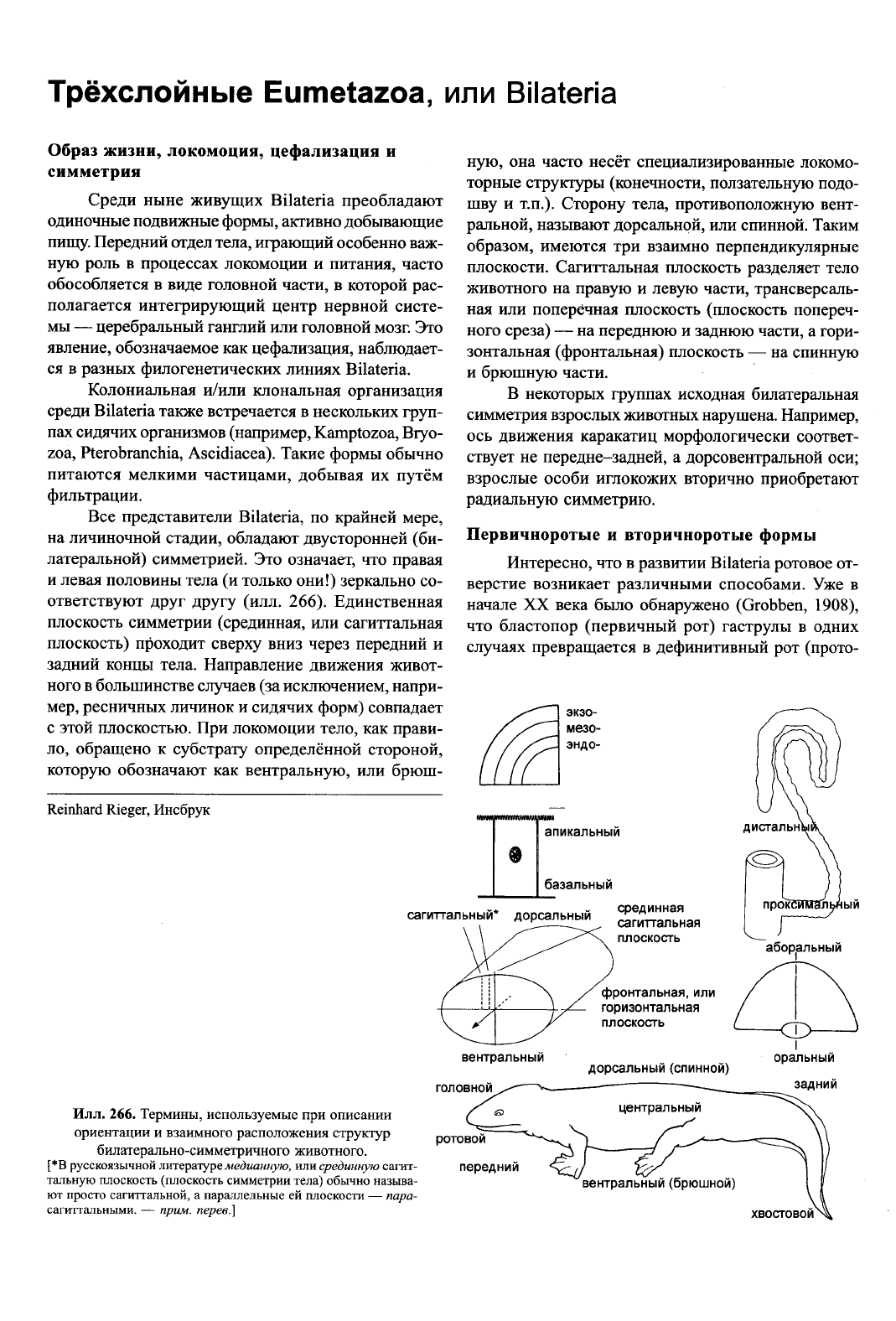
Илл. 266.
Термины,
используемые при описании
ориентации
и взаимного расположения структур
билатерально-симметричного
животного.
[*В
русскоязычной литературе
медианную,
или срединную
сагит-
тальную
плоскость (плоскость симметрии тела) обычно называ-
ют
просто
сагиттальной,
а параллельные ей плоскости —
пара-
сагитгальными.
—
прим.
перев.]
головной
ротовой
передний
оральный
задний
вентральный
(брюшной)
хвостовой
http://jurassic.ru/
Трёхслойные Eumetazoa
199
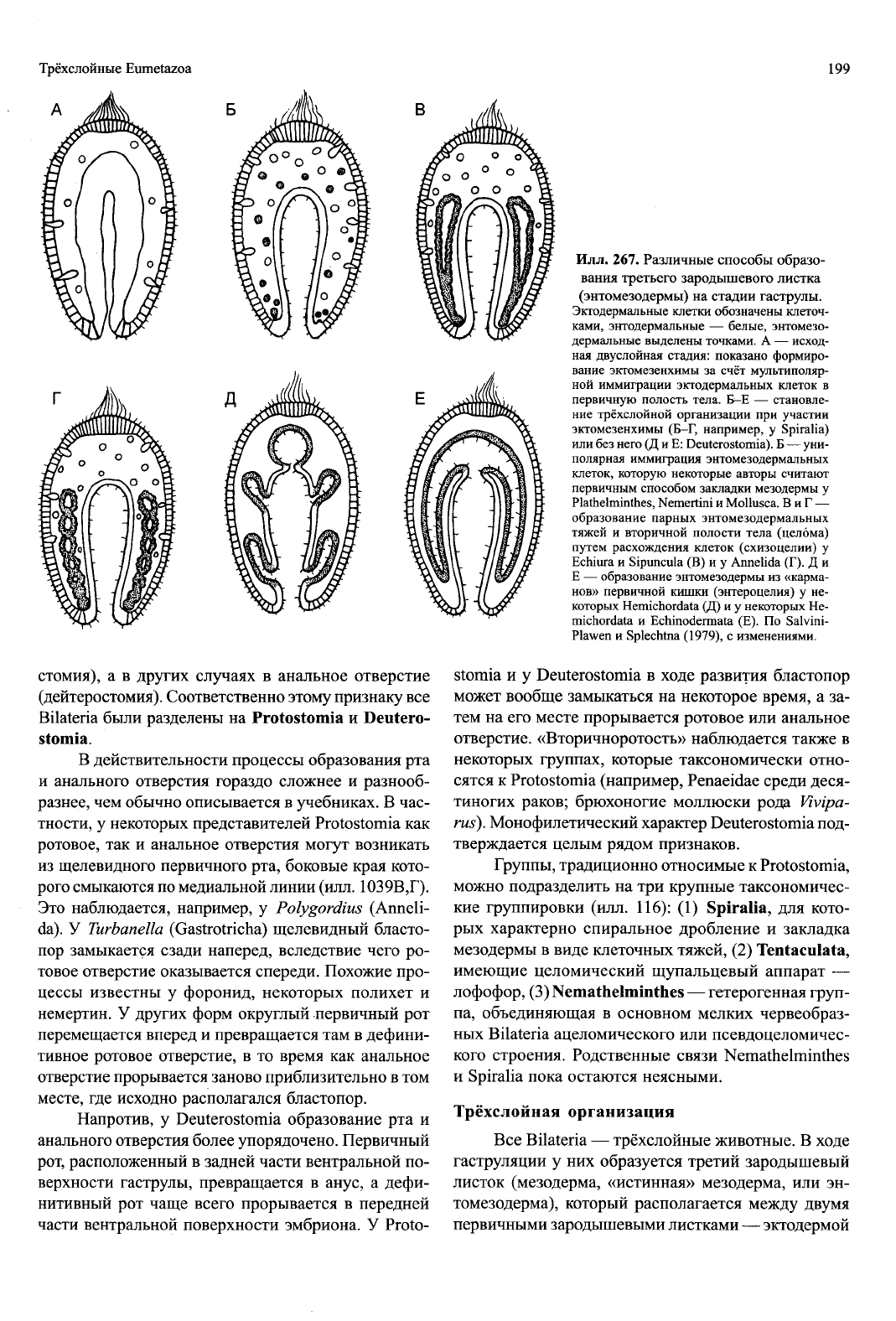
Илл. 267. Различные способы образо-
вания третьего зародышевого листка
(энтомезодермы) на стадии гаструлы.
Эктодермальные клетки обозначены клеточ-
ками, энтодермальные — белые, энтомезо-
дермальные выделены точками. А — исход-
ная двуслойная стадия: показано формиро-
вание эктомезенхимы за счёт мультиполяр-
ной иммиграции эктодермальных клеток в
первичную полость тела. Б-Е — становле-
ние трёхслойной организации при участии
эктомезенхимы (Б-Г, например, у Spiralia)
или без него (Д и Е: Deuterostomia). Б — уни-
полярная иммиграция энтомезодермальных
клеток, которую некоторые авторы считают
первичным способом закладки мезодермы у
Plathelminthes, Nemertini и Mollusca. В и Г —
образование парных энтомезодермальных
тяжей и вторичной полости тела (целома)
путем расхождения клеток (схизоцелии) у
Echiura и Sipuncula (В) и у Annelida (Г). Д и
Е — образование энтомезодермы из «карма-
нов» первичной кишки (энтероцелия) у не-
которых Hemichordata (Д) и у некоторых Не-
michordata и Echinodermata (Е). По Salvini-
Plawen и Splechtna (1979), с изменениями.
стомия), а в других случаях в анальное отверстие
(дейтеростомия). Соответственно этому признаку все
Bilateria были разделены на Protostomia и Deutero-
stomia.
В действительности процессы образования рта
и анального отверстия гораздо сложнее и разнооб-
разнее, чем обычно описывается в учебниках. В час-
тности, у некоторых представителей Protostomia как
ротовое, так и анальное отверстия могут возникать
из щелевидного первичного рта, боковые края кото-
рого смыкаются по медиальной линии (илл. 1039В,Г).
Это наблюдается, например, у Polygordius (Anneli-
da).
У Turbanella (Gastrotricha) щелевидный бласто-
пор замыкается сзади наперед, вследствие чего ро-
товое отверстие оказывается спереди. Похожие про-
цессы известны у форонид, некоторых полихет и
немертин. У других форм округлый первичный рот
перемещается вперед и превращается там в дефини-
тивное ротовое отверстие, в то время как анальное
отверстие прорывается заново приблизительно в том
месте, где исходно располагался бластопор.
Напротив, у Deuterostomia образование рта и
анального отверстия более упорядочено. Первичный
рот, расположенный в задней части вентральной по-
верхности гаструлы, превращается в анус, а дефи-
нитивный рот чаще всего прорывается в передней
части вентральной поверхности эмбриона. У Proto-
stomia и у Deuterostomia в ходе развития бластопор
может вообще замыкаться на некоторое время, а за-
тем на его месте прорывается ротовое или анальное
отверстие. «Вторичноротость» наблюдается также в
некоторых группах, которые таксономически отно-
сятся к Protostomia (например, Penaeidae среди деся-
тиногих раков; брюхоногие моллюски рода Vivipa-
rus).
Монофилетический характер Deuterostomia под-
тверждается целым рядом признаков.
Группы, традиционно относимые к Protostomia,
можно подразделить на три крупные таксономичес-
кие группировки (илл. 116): (1) Spiralia, для кото-
рых характерно спиральное дробление и закладка
мезодермы в виде клеточных тяжей, (2) Tentaculata,
имеющие целомический щупальцевый аппарат —
лофофор, (3) Nemathelminth.es — гетерогенная груп-
па, объединяющая в основном мелких червеобраз-
ных Bilateria ацеломического или псевдоцеломичес-
кого строения. Родственные связи Nemathelminthes
и Spiralia пока остаются неясными.
Трёхслойная организация
Все Bilateria — трёхслойные животные. В ходе
гаструляции у них образуется третий зародышевый
листок (мезодерма, «истинная» мезодерма, или эн-
томезодерма), который располагается между двумя
первичными зародышевыми листками — эктодермой
http://jurassic.ru/
200
Трёхслойные Eumetazoa
теменной
диск
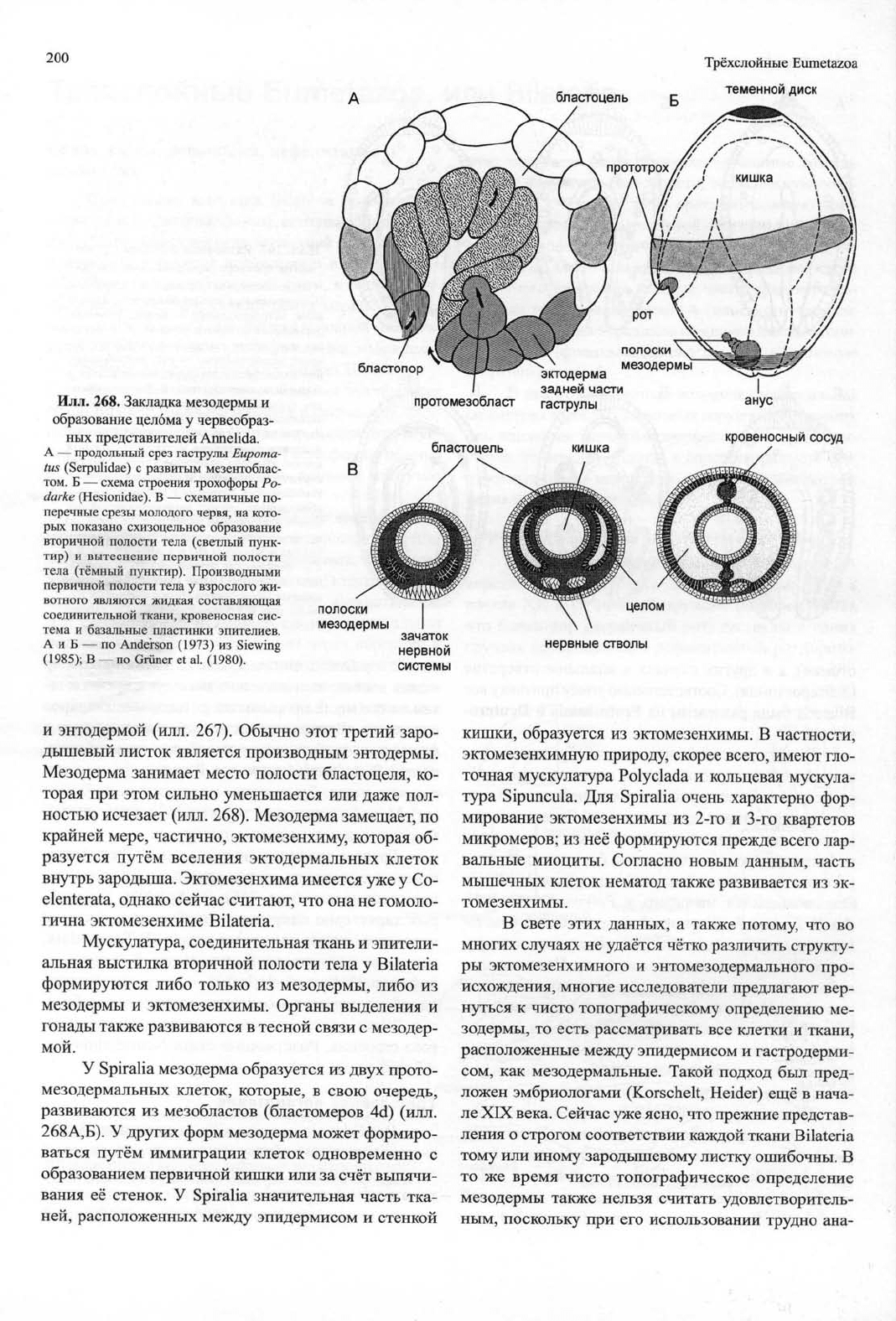
Илл. 268. Закладка мезодермы и
образование целома у червеобраз-
ных представителей Annelida.
А — продольный срез гаструлы Еирота-
tus (Serpulidae) с развитым мезентоблас-
том.
Б — схема строения трохофоры Ро-
darke (Hesionidae). В — схематичные по-
перечные срезы молодого червя, на кото-
рых показано схизоцельное образование
вторичной полости тела (светлый пунк-
тир) и вытеснение первичной полости
тела (тёмный пунктир). Производными
первичной полости тела у взрослого жи-
вотного являются жидкая составляющая
соединительной ткани, кровеносная сис-
тема и базальные пластинки эпителиев.
А и Б — по Anderson (1973) из Siewing
(1985);
В — по Griiner et al. (1980).
бластопор
протомезобласт
бластоцель
эктодерма
задней
части
гаструлы
кишка
анус
кровеносный
сосуд
полоски
мезодермы
целом
нервные стволы
и энтодермой (илл. 267). Обычно этот третий заро-
дышевый листок является производным энтодермы.
Мезодерма занимает место полости бластоцеля, ко-
торая при этом сильно уменьшается или даже пол-
ностью исчезает (илл. 268). Мезодерма замещает, по
крайней мере, частично, эктомезенхиму, которая об-
разуется путём вселения эктодермальных клеток
внутрь зародыша. Эктомезенхима имеется уже у Co-
elenterata, однако сейчас считают, что она не гомоло-
гична эктомезенхиме Bilateria.
Мускулатура, соединительная ткань и эпители-
альная выстилка вторичной полости тела у Bilateria
формируются либо только из мезодермы, либо из
мезодермы и эктомезенхимы. Органы выделения и
гонады также развиваются в тесной связи с мезодер-
мой.
У Spiralia мезодерма образуется из двух прото-
мезодермальных клеток, которые, в свою очередь,
развиваются из мезобластов (бластомеров 4d) (илл.
268А,Б). У других форм мезодерма может формиро-
ваться путём иммиграции клеток одновременно с
образованием первичной кишки или за счёт выпячи-
вания её стенок. У Spiralia значительная часть тка-
ней, расположенных между эпидермисом и стенкой
кишки, образуется из эктомезенхимы. В частности,
эктомезенхимную природу, скорее всего, имеют гло-
точная мускулатура Polyclada и кольцевая мускула-
тура Sipuncula. Для Spiralia очень характерно фор-
мирование эктомезенхимы из 2-го и 3-го квартетов
микромеров; из неё формируются прежде всего лар-
вальные миоциты. Согласно новым данным, часть
мышечных клеток нематод также развивается из эк-
томезенхимы.
В свете этих данных, а также потому, что во
многих случаях не удаётся чётко различить структу-
ры эктомезенхимного и энтомезодермального про-
исхождения, многие исследователи предлагают вер-
нуться к чисто топографическому определению ме-
зодермы, то есть рассматривать все клетки и ткани,
расположенные между эпидермисом и гастродерми-
сом, как мезодермальные. Такой подход был пред-
ложен эмбриологами (Korschelt, Heider) ещё в нача-
ле XIX века. Сейчас уже ясно, что прежние представ-
ления о строгом соответствии каждой ткани Bilateria
тому или иному зародышевому листку ошибочны. В
то же время чисто топографическое определение
мезодермы также нельзя считать удовлетворитель-
ным, поскольку при его использовании трудно ана-
http://jurassic.ru/

Трёхслойные Eumetazoa
201
лизировать филогенетически значимые различия
в
характере закладки мезодермальных тканей
у
отдель-
ных таксонов Bilateria.
Энтомезодерма может формироваться разными
способами. Эмбриональные мезодермальные клет-
ки могут располагаться поодиночке или группами
по
периферии первичной полости тела (псевдоцеломи-
ческая организация)
или
полностью вытеснять
её
(ацеломическая организация,
см. илл.
267Б).
У це-
ломических животных, имеющих ресничную личин-
ку, встречаются
два
основных способа развития
ме-
зодермы.
(1)
Эмбриональные клетки мезодермаль-
ного зачатка дают начало парным клеточным тяжам,
расположенным
по
бокам
от
первичной кишки,
—
так
наз.
мезодермальным полоскам
(илл.
267В,Г,
268В).
Вторичная полость тела (см. ниже) возникает
путём расслоения этих полосок. Такой вариант раз-
вития, обозначаемый
как
схизоцелия, особенно
ха-
рактерен для моллюсков и членистоногих (илл. 267Г,
268В).
У
моллюсков мезодермальные полоски быст-
ро превращаются
в
мезенхиматозную ткань.
(2)
Ме-
зодерма образуется путём выпячивания «карма-
нов» первичной кишки (энтероцелия). При этом об-
разуются эпителиальные мешки, полость которых
и
представляет собой вторичную полость тела. Такой
способ часто встречается
у
вторичноротых
(илл.
267Д,Е, 1060, 1096). Существуют разнообразные мо-
дификации
и
комбинации этих двух способов, осо-
бенно
у
организмов
с
прямым развитием
(в
том чис-
ле
у
позвоночных). Однако даже
для
форм, имею-
щих ресничную личинку, многие детали развития
мезодермы остаются неясными.
Исходные варианты строения нервной
системы
Как было сказано ранее, для эволюции Bilateria
очень характерно обособление центральной нервной
системы
и
развитие головного мозга. Мозг возника-
ет
в
результате концентрации нервных клеток
на
пе-
реднем конце тела. Одновременно
с
этим исходно
равномерная периферическая сеть нейронов (нерв-
сенсорные клетки
железистые
клетки
спинные
гигантские
волокна
нейрон
базиэпидермальный
нервный плексус
Илл.
269.
Базиэпителиальная нервная система
Saccoglossus
(Hemichordata).
По Bullock
и
Horridge (1965).
ный плексус) преобразуется
в
систему продольных
нервных стволов, соединённых поперечными тяжа-
ми.
Вне
Bilateria подобная концентрация нервных
элементов известна только
у
отдельных представи-
телей Metazoa.
В исходном
для
Bilateria варианте
вся
нервная
система, включая головной мозг, располагается
ба-
зиэпителиально. Периферическая нервная система,
по-видимому, развивается
из
нескольких нервных
сетей (сенсорной
и
моторной, бази-
и
субэпители-
альной). Уже
у
примитивных форм
в
пределах Bila-
teria может возникать частичное пространственное
разделение
тел
нейронов
и
плотных сплетений
от-
ростков. Такие базиэпителиальные нервные систе-
мы
со
слабо выраженным мозгом
и
нерегулярным
периферическим плексусом встречаются
у
различ-
ных личинок,
а
также
у
некоторых взрослых бески-
шечных турбеллярий, Tentaculata, Pterobranchia иХе-
noturbella
bocki
(Enteropneusta, илл. 1069).
В дальнейшей эволюции нервной системы мож-
но различить
два
основных направления:
оболочка
из
соединительной
ткани
яв^"
тела
нейронов
кровеносный
сосуд
Илл.
270.
Поперечный срез погружённого
вентрального нервного ствола Lumbricus
(Annelida)
с
боковыми нервами.
Исходно нервная система напоминала верёвочную
лестницу; в дальнейшем
её
продольные коннекти-
вы практически слились. Погружение нервной
си-
стемы происходило
без
впячивания покровов.
W. Westheide, Оснабрюк.
http://jurassic.ru/
202
Трёхслойные Eumetazoa
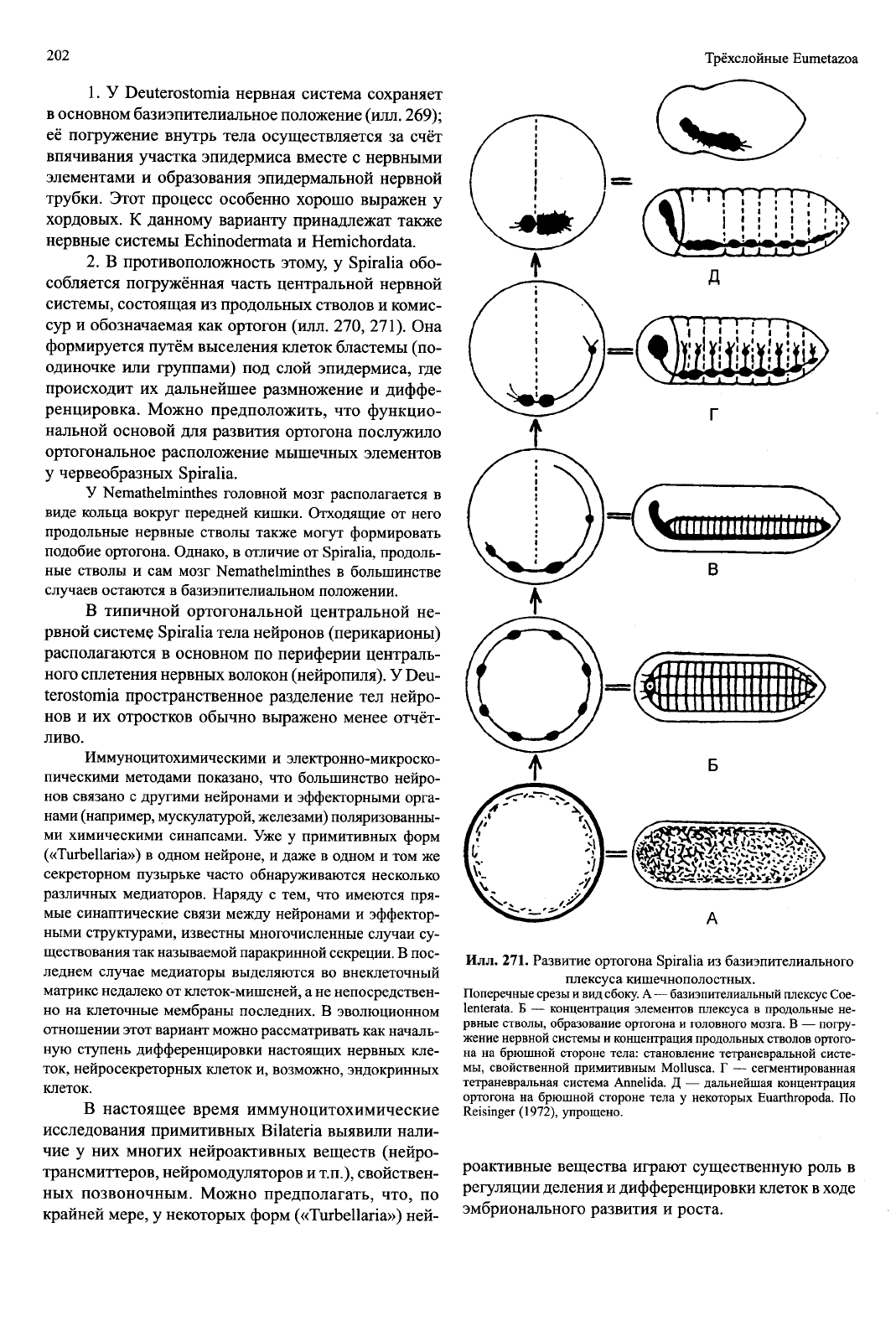
Илл. 271. Развитие ортогона Spiralia из базиэпителиального
плексуса кишечнополостных.
Поперечные срезы и вид сбоку. А — базиэпителиальный плексус Coe-
lenterata. Б — концентрация элементов плексуса в продольные не-
рвные стволы, образование ортогона и головного мозга. В — погру-
жение нервной системы и концентрация продольных стволов ортого-
на на брюшной стороне тела: становление тетраневральной систе-
мы,
свойственной примитивным Mollusca. Г — сегментированная
тетраневральная система Annelida. Д — дальнейшая концентрация
ортогона на брюшной стороне тела у некоторых Euarthropoda. По
Reisinger (1972), упрощено.
роактивные вещества играют существенную роль в
регуляции деления и дифференцировки клеток
в
ходе
эмбрионального развития и роста.
1.
У Deuterostomia нервная система сохраняет
в основном базиэпителиальное положение (илл. 269);
её погружение внутрь тела осуществляется за счёт
впячивания участка эпидермиса вместе с нервными
элементами и образования эпидермальной нервной
трубки. Этот процесс особенно хорошо выражен у
хордовых. К данному варианту принадлежат также
нервные системы Echinodermata и Hemichordata.
2.
В противоположность этому, у Spiralia обо-
собляется погружённая часть центральной нервной
системы, состоящая из продольных стволов и комис-
сур и обозначаемая как ортогон (илл. 270, 271). Она
формируется путём выселения клеток бластемы (по-
одиночке или группами) под слой эпидермиса, где
происходит их дальнейшее размножение и диффе-
ренцировка. Можно предположить, что функцио-
нальной основой для развития ортогона послужило
ортогональное расположение мышечных элементов
у червеобразных Spiralia.
У Nemathelminthes головной мозг располагается в
виде кольца вокруг передней кишки. Отходящие от него
продольные нервные стволы также могут формировать
подобие ортогона. Однако, в отличие от Spiralia, продоль-
ные стволы и сам мозг Nemathelminthes в большинстве
случаев остаются в базиэпителиальном положении.
В типичной ортогональной центральной не-
рвной системе Spiralia тела нейронов (перикарионы)
располагаются в основном по периферии централь-
ного сплетения нервных волокон (нейропиля). У Deu-
terostomia пространственное разделение тел нейро-
нов и их отростков обычно выражено менее отчёт-
ливо.
Иммуноцитохимическими и электронно-микроско-
пическими методами показано, что большинство нейро-
нов связано с другими нейронами и эффекторными орга-
нами (например, мускулатурой, железами) поляризованны-
ми химическими синапсами. Уже у примитивных форм
(«Turbellaria») в одном нейроне, и даже в одном и том же
секреторном пузырьке часто обнаруживаются несколько
различных медиаторов. Наряду с тем, что имеются пря-
мые синаптические связи между нейронами и эффектор-
ными структурами, известны многочисленные случаи су-
ществования так называемой паракринной
секреции.
В
пос-
леднем случае медиаторы выделяются во внеклеточный
матрикс недалеко от клеток-мишеней, а не непосредствен-
но на клеточные мембраны последних. В эволюционном
отношении этот вариант можно рассматривать как началь-
ную ступень дифференцировки настоящих нервных кле-
ток, нейросекреторных клеток и, возможно, эндокринных
клеток.
В настоящее время иммуноцитохимические
исследования примитивных Bilateria выявили нали-
чие у них многих нейроактивных веществ (нейро-
трансмиттеров, нейромодуляторов и т.п.), свойствен-
ных позвоночным. Можно предполагать, что, по
крайней мере, у некоторых форм («Turbellaria») ней-
http://jurassic.ru/
Трёхслойные Eumetazoa
пелагическая взрослая форма
(нерест)
бентосные ювенильные стадии
Илл.
272.
Двухфазный жизненный цикл на примере черве-
образных представителей Spiralia и прогенетическое
происхождение новых таксонов от ювенильных форм,
временно обитающих в толще грунта.
По Westheide(1987).
203
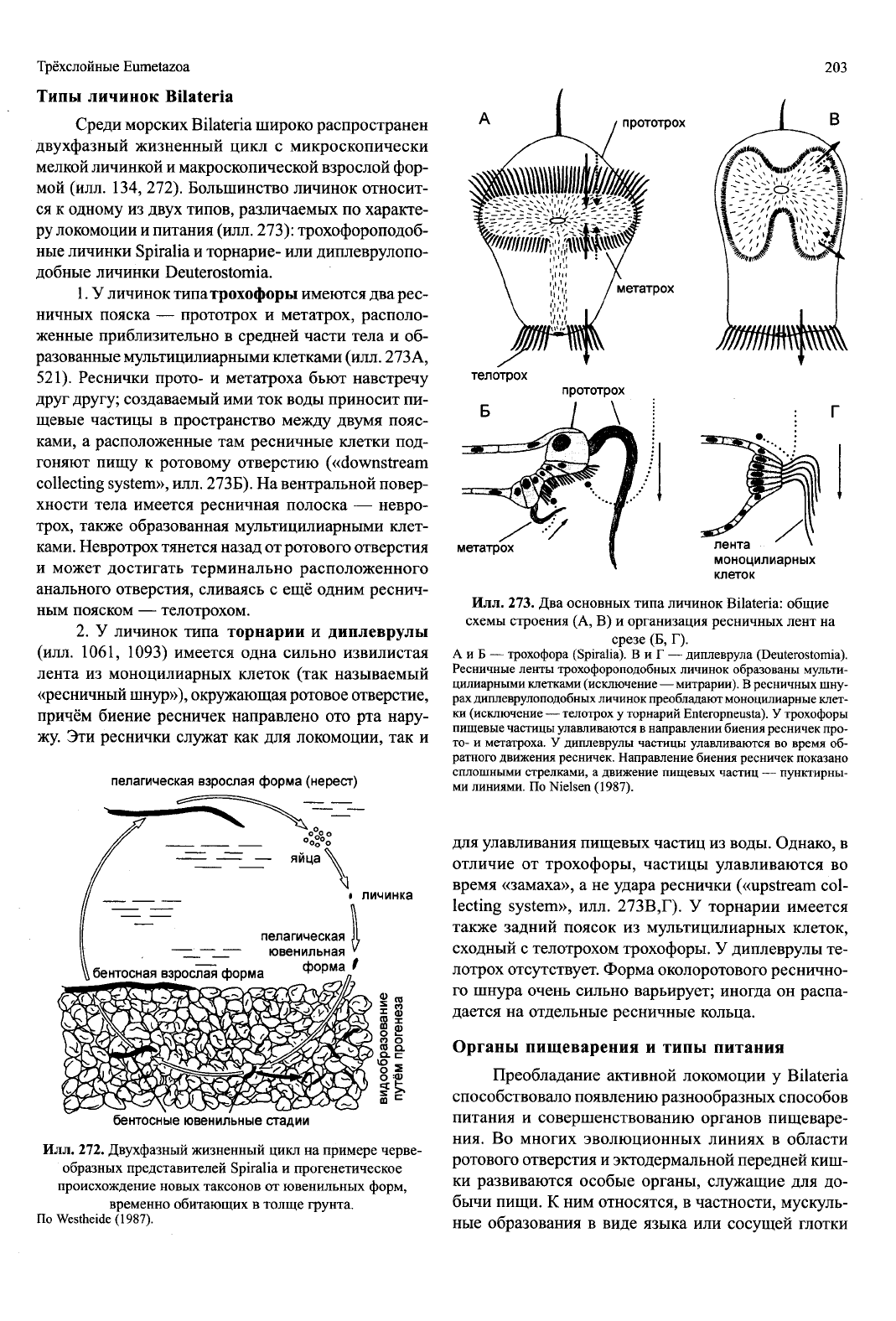
телотрох
прототрох
клеток
Илл.
273.
Два основных типа личинок Bilateria: общие
схемы строения (А, В) и организация ресничных лент на
срезе (Б, Г).
А и Б — трохофора (Spiralia). В и Г — диплеврула (Deuterostomia).
Ресничные ленты трохофороподобных личинок образованы мульти-
цилиарными клетками (исключение — митрарии). В ресничных шну-
рах диплеврулоподобных личинок преобладают моноцилиарные клет-
ки (исключение — телотрох у торнарий Enteropneusta). У трохофоры
пищевые частицы улавливаются в направлении биения ресничек про-
то-
и метатроха. У диплеврулы частицы улавливаются во время об-
ратного движения ресничек. Направление биения ресничек показано
сплошными стрелками, а движение пищевых частиц — пунктирны-
ми линиями. По Nielsen (1987).
для улавливания пищевых частиц из воды. Однако, в
отличие от трохофоры, частицы улавливаются во
время «замаха», а не удара реснички («upstream col-
lecting system», илл. 273В,Г). У торнарии имеется
также задний поясок из мультицилиарных клеток,
сходный с телотрохом трохофоры. У диплеврулы те-
лотрох отсутствует. Форма околоротового реснично-
го шнура очень сильно варьирует; иногда он распа-
дается на отдельные ресничные кольца.
Органы пищеварения и типы питания
Преобладание активной локомоции у Bilateria
способствовало появлению разнообразных способов
питания и совершенствованию органов пищеваре-
ния. Во многих эволюционных линиях в области
ротового отверстия и эктодермальной передней киш-
ки развиваются особые органы, служащие для до-
бычи пищи. К ним относятся, в частности, мускуль-
ные образования в виде языка или сосущей глотки
Типы личинок Bilateria
Среди морских Bilateria широко распространен
двухфазный жизненный цикл с микроскопически
мелкой личинкой и макроскопической взрослой фор-
мой (илл. 134, 272). Большинство личинок относит-
ся к одному из двух типов, различаемых по характе-
ру локомоции и питания (илл. 273): трохофороподоб-
ные личинки Spiralia и торнарие- или диплеврулопо-
добные личинки Deuterostomia.
1.
У личинок типатрохофоры имеются два рес-
ничных пояска — прототрох и метатрох, располо-
женные приблизительно в средней части тела и об-
разованные мультицилиарными клетками (илл. 273А,
521).
Реснички прото- и метатроха бьют навстречу
друг другу; создаваемый ими ток воды приносит пи-
щевые частицы в пространство между двумя пояс-
ками, а расположенные там ресничные клетки под-
гоняют пищу к ротовому отверстию («downstream
collecting system», илл. 273Б). На вентральной повер-
хности тела имеется ресничная полоска — невро-
трох, также образованная мультицилиарными клет-
ками. Невротрох тянется назад от ротового отверстия
и может достигать терминально расположенного
анального отверстия, сливаясь с ещё одним реснич-
ным пояском — телотрохом.
2.
У личинок типа торнарии и диплеврулы
(илл. 1061, 1093) имеется одна сильно извилистая
лента из моноцилиарных клеток (так называемый
«ресничный шнур»), окружающая ротовое отверстие,
причём биение ресничек направлено ото рта нару-
жу. Эти реснички служат как для локомоции, так и
http://jurassic.ru/

204
Трёхслойные Eumetazoa
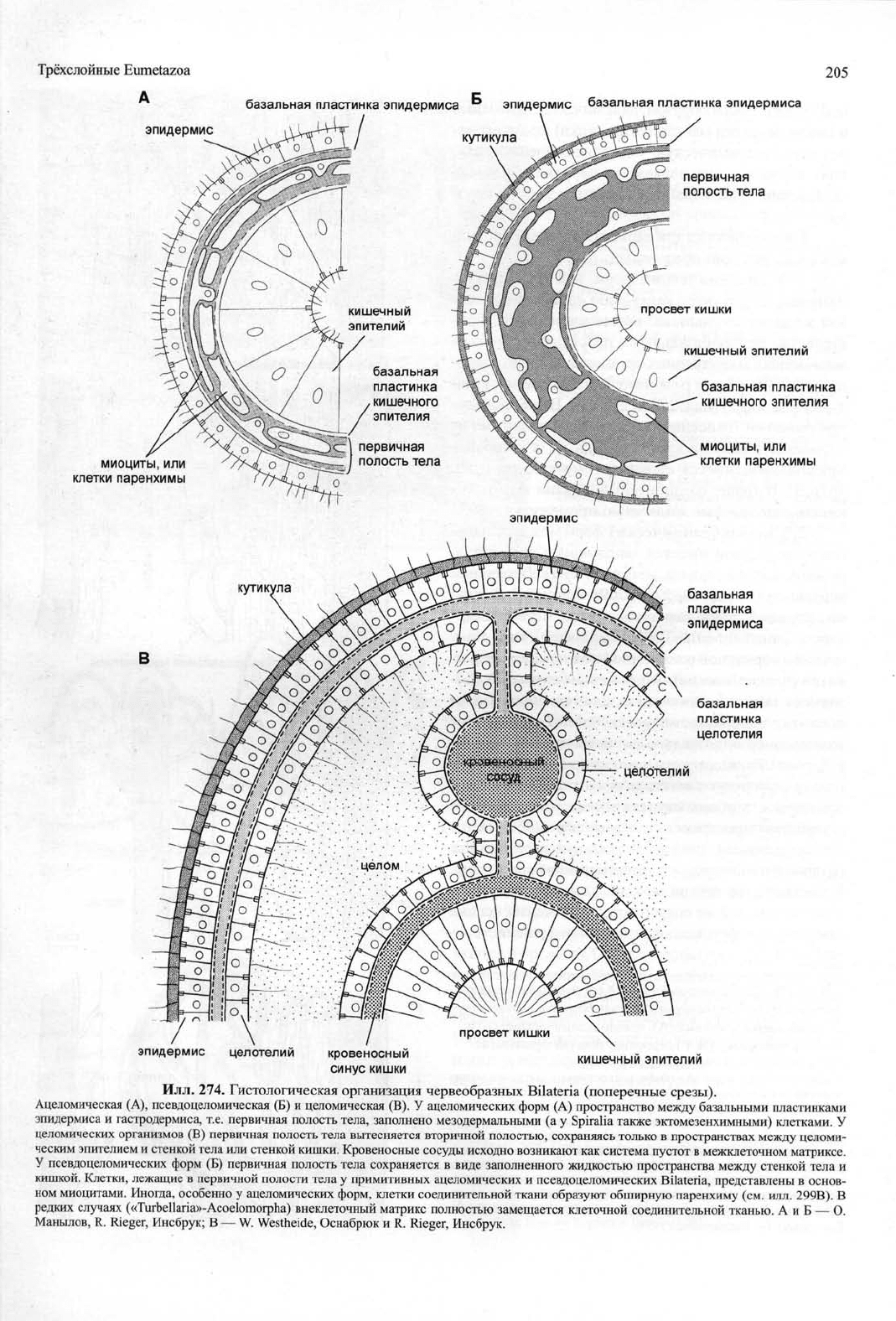
Основные типы полости тела
В теле Bilateria различают две системы полос-
тей,
расположенных между кишкой и эпидермисом:
(1) первичная полость тела (псевдоцель), соответ-
ствующая эмбриональному бластоцелю и дающая
начало кровеносным сосудам, и (2) вторичная по-
лость тела, или целом (илл. 274).
Жидкое или гелеобразное содержимое первич-
ной полости тела (вместе с находящимися в нем от-
дельными клетками) в примитивном случае ограни-
чено базальными пластинками эпидермиса и гаст-
родермиса; иными словами, эпителиальные клетки,
ограничивающие первичную полость, обращены к
ней базальной частью. Напротив, вторичная по-
лость тела (целомическая жидкость и плавающие в
ней клетки-целомоциты) располагается с апикаль-
ной стороны ограничивающего её целомического
эпителия (илл. 274). Поскольку апикальная и базо-
латеральная части мембраны эпителиальных клеток
имеют различный состав мембранных белков и ион-
ных насосов, указанное топологическое различие
имеет определенные функциональные последствия.
В зависимости от отсутствия или наличия вто-
ричной полости, различают три типа организации
тела Bilateria: ацеломический, псевдоцеломичес-
кий (включая такие варианты, как миксоцель или ге-
моцель — см. ниже) и целомический.
Псевдоцеломический и ацеломический варианты
могут быть гистологически сопоставлены со строени-
ем эмбрионального бластоцеля (илл. 275А,Б, 276А).
Различие между ними состоит в наличии или отсут-
ствии достаточно обширного заполненного жидко-
стью пространства в составе внеклеточного матрик-
са между кишкой и стенкой тела. По расположению
и гистологической организации этот матрикс соот-
ветствует мезоглее «Coelenterata» и мезохилу Porifera.
Целомическая организация, напротив, подразумева-
ет наличие вторичной полости тела мезодермальной
природы — целома (илл. 274, 275В,Г, 276Б).
Вторичная полость тела исходно ограничена
мезодермальным (частично также эктомезенхимным)
эпителием (илл. 274, 275В,Г). В ходе развития она
вытесняет бластоцель (илл. 268В), от которого у при-
митивных целомических форм остаются только кро-
веносные сосуды и тканевая жидкость межклеточ-
ного матрикса, заключенного между эпителиальны-
ми пластами эпидермиса, гастродермиса и целоми-
ческой выстилки (целотелия) (илл. 274В, 276Б).
Личинки Bilateria обладают псевдоцеломичес-
кой или ацеломической организацией. В большин-
стве случаев у них имеются только зачатки мезодер-
мы,
хотя целомические полости в этих зачатках уже
могут быть дифференцированы (например, у поздних
личинок Polychaeta, Phoronida, Hemichordata, Echi-
со стилетами или без таковых (у многих Nemathel-
minthes, Annelida и высших Plathelminthes), твёрдые
ротовые части (радула Mollusca, мастаке Rotatoria,
челюсти Annelida или Gnathostomulida), а также че-
люсти высших позвоночных.
Многие водные Bilateria, подобно многим «Сое-
lenterata», питаются мелкими взвешенными в воде
частицами. Добыча пищи у них сводится к отсорти-
ровыванию съедобных частиц от частиц, непригод-
ных для питания. Bilateria используют различные ме-
ханизмы извлечения пищевых частиц из воды, кото-
рые могут быть основаны на работе ресничек, выде-
лении слизи или сокращении мышц. Реснички, со-
здающие ток воды, обычно располагаются на боко-
вой поверхности щупалец (например, Tentaculata, не-
которые сидячие полихеты) или жабр (например, дву-
створчатые моллюски, асцидии). При использовании
мускулатуры для прокачки воды всегда развиваются
фильтрующие аппараты (например, видоизмененные
конечности у Cirripedia, жаберные выросты у рыб).
Животные, которые сами создают ток воды,
обозначаются как активные фильтраторы (например,
Polychaeta, многие Crustacea). Напротив, пассивные
фильтраторы (многие сидячие полихеты) подставля-
ют свои щупальца течению. Пассивный и активный
способы питания часто сочетаются (например, у мно-
гих Cirripedia). Другие животные добывают пище-
вые частицы не из воды, а из рыхлого грунта (ила
или песка); их называют детритофагами.
За немногими исключениями (Plathelminthes,
Gnathostomulida), пищеварительный тракт Bilateria
имеет не только ротовое — как у «Coelenterata» —
но и анальное отверстие. Наличие ануса позволяет,
во-первых, осуществлять транспорт пищи по кишке
только в одном направлении, а во-вторых, разделить
процессы поглощения пищи и выделения неперева-
ренных остатков. У многих животных наблюдается
функциональная специализация отдельных участков
сквозного кишечника.
Отсутствие анального отверстия у Plathelminthes и
Gnathostomulida обычно рассматривают как примитивный
признак,
а
анальные
поры,
возникающие
у
отдельных пред-
ставителей Plathelminthes,
— как
вторичное приобретение.
Следует заметить, что у многих (возможно, у всех) гнато-
стомулид задний конец кишки находится
в
тесном контак-
те с дорсальным эпидермисом; вероятно, в этой области
располагается временное анальное
отверстие.
В
некоторых
группах Bilateria (например, у различных представителей
Echinodermata) анус вторично исчезает.
Для многих таксонов характерно развитие вспо-
могательных железистых органов, обеспечивающих
внеклеточное (например, поджелудочная железа и
печень позвоночных) или внутриклеточное (напри-
мер,
печень моллюсков) пищеварение.
http://jurassic.ru/
Трёхслойные Eumetazoa
205
базальная пластинка эпидермиса " эпидермис базальная пластинка эпидермиса
пидермис
просвет кишки
миоциты,
или
клетки паренхимы
- первичная
полость тела
кишечный эпителии
базальная пластинка
кишечного эпителия
миоциты,
или
клетки паренхимы
базальная
пластинка
эпидермиса
базальная
пластинка
целотелия
эпидермис целотелии
кровеносный
синус кишки
кишечный эпителии
Илл.
274. Гистологическая организация червеобразных Bilateria (поперечные срезы).
Ацеломическая (А), псевдоцеломическая (Б) и целомическая (В). У ацеломических форм (А) пространство между базальными пластинками
эпидермиса и гастродермиса, т.е. первичная полость тела, заполнено мезодермальными (а у Spiralia также эктомезенхимными) клетками. У
целомических организмов (В) первичная полость тела вытесняется вторичной полостью, сохраняясь только в пространствах между целоми-
ческим эпителием и стенкой тела или стенкой кишки. Кровеносные сосуды исходно возникают как система пустот в межклеточном матриксе.
У псевдоцеломических форм (Б) первичная полость тела сохраняется в виде заполненного жидкостью пространства между стенкой тела и
кишкой. Клетки, лежащие в первичной полости тела у примитивных ацеломических и псевдоцеломических Bilateria, представлены в основ-
ном миоцитами. Иногда, особенно у ацеломических форм, клетки соединительной ткани образуют обширную паренхиму (см. илл. 299В). В
редких случаях («Turbellaria»-Acoelomorpha) внеклеточный матрикс полностью замещается клеточной соединительной тканью. А и Б — О.
Манылов, R. Rieger, Инсбрук; В — W. Westheide, Оснабрюк и R. Rieger, Инсбрук.
http://jurassic.ru/