Вестхайде В.,Ригер Р.( ред.) Зоология беспозвоночных в двух томах. Том 1: от простейших до моллюсков и артропод
Подождите немного. Документ загружается.

206
Трёхслойные Eumetazoa
nodermata). Таким образом, целомические животные
в своем развитии (на стадии личинки) проходят че-
рез псевдоцеломическую или ацеломическую ста-
дию:
вначале у них появляется первичная полость
тела, а затем уже в ней формируются зачатки целб-
ма.
Гистологически три варианта строения отлича-
ются следующими признаками:
1.
У ацеломических форм, как и следует из
названия, отсутствует какая-либо единая заполнен-
ная жидкостью полость, кроме кишечника. Про-
странство между базальными пластинками эпидер-
миса, кишки и внутренних органов заполнено соеди-
нительной тканью и развивающейся из неё мускула-
турой (см. ниже) (илл. 274А, 275А,Б). В эмбриональ-
ном развитии эта соединительная ткань возникает из
энтомезодермальной клеточной массы, к которой у
Spiralia добавляется ещё и эктомезенхима (илл.
267Б,В). В толще соединительной ткани могут воз-
никать заполненные жидкостью промежутки.
2.
У псевдоцеломических форм между эпидер-
мисом и кишкой имеется заполненная жидкостью
полость, которая, однако, не ограничена собственным
эпителием (илл. 274Б, 275Б, 299). Эта полость либо
представляет собой первичную полость тела (неко-
торые Nemathelminthes), либо возникает путём объе-
динения первичной и вторичной полостей в ходе раз-
вития (членистоногие). В последнем случае её обо-
значают также как миксоцель или гемоцель. Таким
образом, псевдоцеломическая организация может
возникать принципиально разными путями. Однако
у взрослого животного «истинный» псевдоцель
нельзя отличить от миксоцеля по гистологическим
признакам: в обоих случаях имеется заполненное
жидкостью пространство, ограниченное базальны-
ми пластинками эпидермиса, кишки и различных
органов (метанефридиев, гонад и половых протоков).
У большинства организмов, обладающих гемо- или
миксоцелем, всё же сохраняются рудименты целома
(например, нефридиальные мешочки у членистоно-
гих).
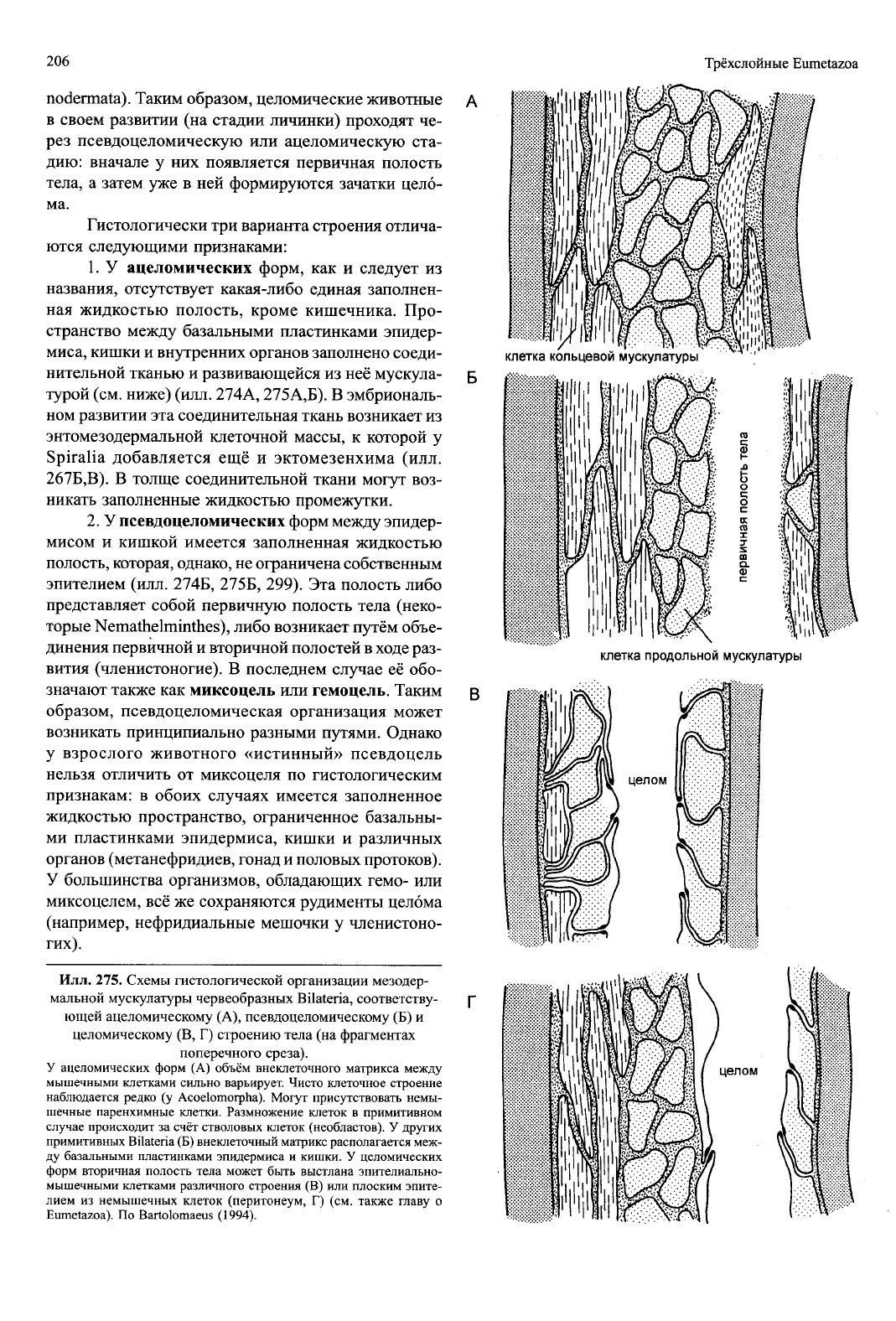
Илл.
275. Схемы гистологической организации мезодер-
мальной мускулатуры червеобразных Bilateria, соответству-
ющей ацеломическому (А), псевдоцеломическому (Б) и
целомическому (В, Г) строению тела (на фрагментах
поперечного среза).
У ацеломических форм (А) объём внеклеточного матрикса между
мышечными клетками сильно варьирует. Чисто клеточное строение
наблюдается редко (у Acoelomorpha). Могут присутствовать немы-
шечные паренхимные клетки. Размножение клеток в примитивном
случае происходит за счёт стволовых клеток (необластов). У других
примитивных Bilateria (Б) внеклеточный матрикс располагается меж-
ду базальными пластинками эпидермиса и кишки. У целомических
форм вторичная полость тела может быть выстлана эпителиально-
мышечными клетками различного строения (В) или плоским эпите-
лием из немышечных клеток (перитонеум, Г) (см. также главу о
Eumetazoa). По Bartolomaeus (1994).
http://jurassic.ru/
Трёхслойные Eumetazoa
207
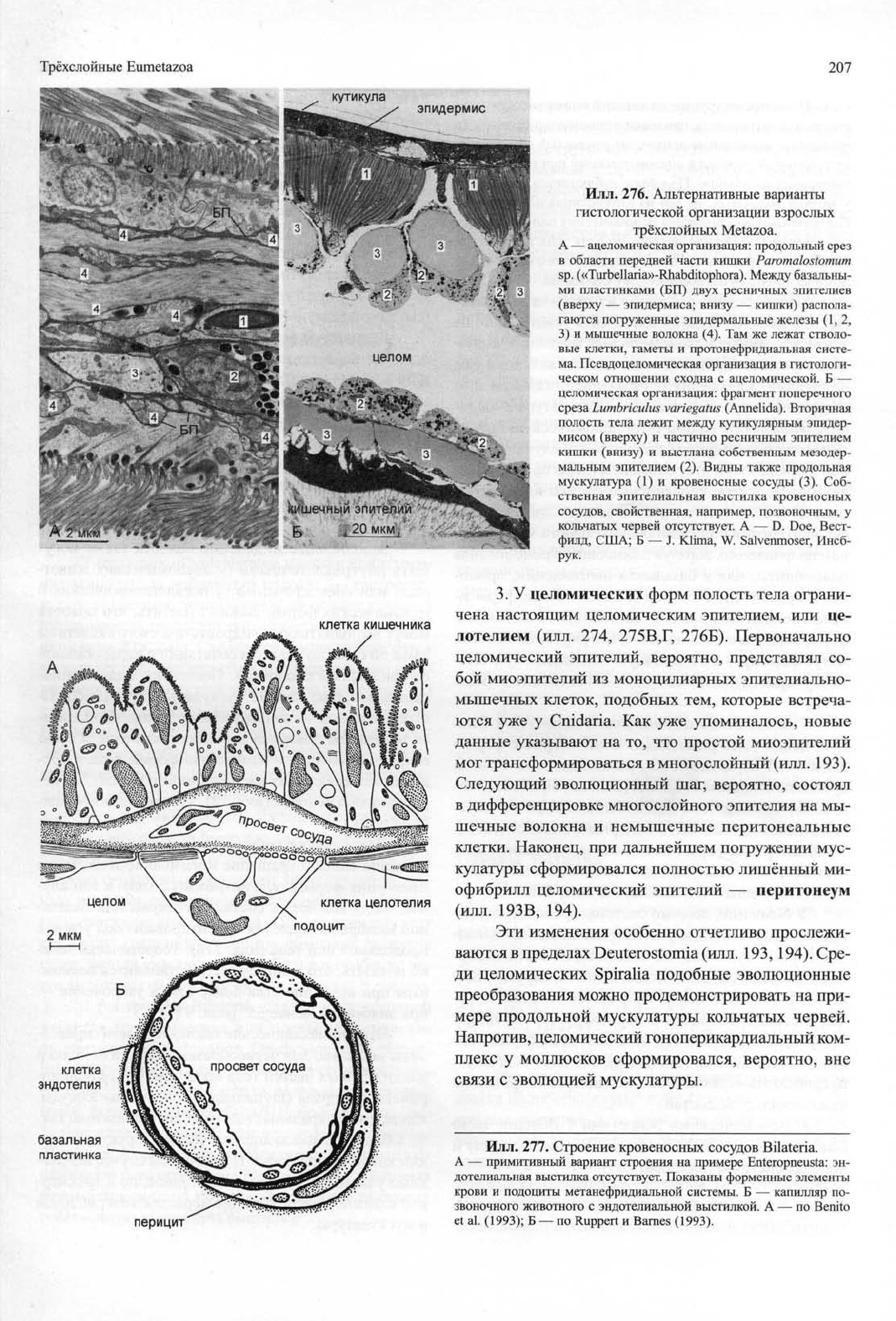
Илл.
276. Альтернативные варианты
гистологической организации взрослых
трёхслойных Metazoa.
А — ацеломическая организация: продольный срез
в области передней части кишки Paromalostomum
sp.
(«Turbellaria»-Rhabditophora). Между базальны-
ми пластинками (БП) двух ресничных эпителиев
(вверху — эпидермиса; внизу — кишки) распола-
гаются погруженные эпидермальные железы (1,2,
3) и мышечные волокна (4). Там же лежат стволо-
вые клетки, гаметы и протонефридиальная систе-
ма. Псевдоцеломическая организация в гистологи-
ческом отношении сходна с ацеломической. Б —
целомическая организация: фрагмент поперечного
среза Lumbriculus variegatus (Annelida). Вторичная
полость тела лежит между кутикулярным эпидер-
мисом (вверху) и частично ресничным эпителием
кишки (внизу) и выстлана собственным мезодер-
мальным эпителием (2). Видны также продольная
мускулатура (1) и кровеносные сосуды (3). Соб-
ственная эпителиальная выстилка кровеносных
сосудов, свойственная, например, позвоночным, у
кольчатых червей отсутствует. А — D. Doe, Вест-
филд,
США; Б — J. Klima, W. Salvenmoser, Инсб-
рук.
клетка кишечника
перицит
3.
У целомических форм полость тела ограни-
чена настоящим целомическим эпителием, или це-
лотелием (илл. 274, 275В,Г, 276Б). Первоначально
целомический эпителий, вероятно, представлял со-
бой миоэпителий из моноцилиарных эпителиально-
мышечных клеток, подобных тем, которые встреча-
ются уже у Cnidaria. Как уже упоминалось, новые
данные указывают на то, что простой миоэпителий
мог трансформироваться в многослойный (илл. 193).
Следующий эволюционный шаг, вероятно, состоял
в дифференцировке многослойного эпителия на мы-
шечные волокна и немышечные перитонеальные
клетки. Наконец, при дальнейшем погружении мус-
кулатуры сформировался полностью лишённый ми-
офибрилл целомический эпителий — перитонеум
(илл. 193В, 194).
Эти изменения особенно отчетливо прослежи-
ваются в пределах Deuterostomia (илл. 193, 194). Сре-
ди целомических Spiralia подобные эволюционные
преобразования можно продемонстрировать на при-
мере продольной мускулатуры кольчатых червей.
Напротив, целомический гоноперикардиальный ком-
плекс у моллюсков сформировался, вероятно, вне
связи с эволюцией мускулатуры.
Илл.
277. Строение кровеносных сосудов Bilateria.
А — примитивный вариант строения на примере Enteropneusta: эн-
дотелиальная выстилка отсутствует. Показаны форменные элементы
крови и подоциты метанефридиальной системы. Б — капилляр по-
звоночного животного с эндотелиальной выстилкой. А — по Benito
et al. (1993); Б — по Ruppert и Barnes (1993).
http://jurassic.ru/

208
Трёхслойные Eumetazoa
Развитие опорного аппарата
В главе, посвященной Eumetazoa, мы уже рас-
сматривали скелетную функцию внеклеточного мат-
рикса, мускулатуры, а также вне- и внутриклеточных
полостей. Поскольку считается, что предки Bilateria
были мелкими червеобразными организмами, следу-
ет более подробно охарактеризовать функциональ-
ный комплекс, состоящий из элементов внеклеточ-
ного матрикса (кутикула и базальные пластинки эпи-
телиев), мускулатуры и гидростатического скелета
(см.
также главу о Nemathelminthes).
Решающую роль играет специфическое распо-
ложение нерастяжимых волокон в слоях, окружаю-
щих всё тело или отдельные органы животного. Эти
волокна могут быть как внеклеточными (коллаген),
так и внутриклеточными (микрофиламенты цитос-
келета). Системы с различным расположением во-
локон имеют разные механические характеристики
(илл. 278). Чаще всего волокна в соседних слоях рас-
полагаются ортогонально или спирально (см. илл.
200).
Заполненные жидкостью полости также могут
быть внутриклеточными (у ацеломических живот-
ных) или внеклеточными (у псевдоцеломических и
целомических форм). Важно отметить, что полости
могут выполнять роль гидростатического скелета не
сами по себе, но только в сочетании с нерастяжимой
фибриллярной оболочкой. Поскольку жидкости не-
сжимаемы, сокращение мускулатуры (у червеобраз-
ных животных — кольцевой и продольной) вызыва-
ет изменение давления в полости, которое, в свою
очередь, приводит к изменению формы тела в пре-
делах, определяемых расположением гибких, но не-
растяжимых волокон (илл. 121,199; см. также Nemat-
helminthes).
Для червеобразного животного особенно важ-
ны удлинение, сокращение и изгибание тела. Такие
изменения формы тела возможны только в том слу-
чае,
когда волокна в составе кутикулы или базаль-
ной мембраны эпителия ориентированы под углом к
продольной оси тела (илл. 278). Теоретически мож-
но показать, что удлинение тела становится возмож-
ным при величине угла более 55°, а укорочение —
при значениях менее 55° (илл. 971).
Эти биомеханические закономерности справед-
ливы не только для червеобразных организмов, но и
для отдельных частей тела более или менее цилинд-
рической формы (щупальца, кровеносные сосуды,
хорда, амбулакральные ножки). Они приложимы так-
же к более сложным ацеломическим структурам, та-
ким как нога моллюска. В последнем случае необхо-
димо учитывать не только послойное, но и трёхмер-
ное взаимное расположение нерастяжимых волокон
и мускулатуры.
В некоторых группах целотелий может частично или
полностью утрачивать признаки эпителиальной ткани (в
частности, апикальные ленточные контакты), что означа-
ет вторичный переход к ацеломическому или псевдоцело-
мическому состоянию. Подобное наблюдается, например,
у мелких кольчатых червей из интерстициальной фауны.
Преобразование
и
редукция целомических полостей хоро-
шо известны также для пиявок. Во всех этих группах на-
блюдаются переходы от чисто целомического к псевдоце-
ломическому или ацеломическому строению.
Целом развивается либо из мезенхимной мас-
сы (или мезодермальных клеточных тяжей), либо из
эпителизованных мезодермальных мешков. У взрос-
лых животных производными целома являются сам
целотелий, связанная с ним эпителиальная или
погруженная субэпителиальная мускулатура и соеди-
нительная ткань. Кроме того, целомическую приро-
ду имеют мезентерии (в большинстве случаев спин-
ной и брюшной) и диссепименты (перегородки), зак-
лючающие в себе кровеносные сосуды или синусы
(илл. 274, 277). Клетки целомического эпителия мо-
гут выходить из эпителиального пласта в апикаль-
ном направлении, образуя плавающие в полости тела
целомоциты, или в базальном направлении, прони-
кая в кровеносные сосуды и давая начало формен-
ным элементам крови. У высокоорганизованных
форм, таких как позвоночные, мигрирующие клетки
могут формировать вторичную эпителиальную выс-
тилку кровеносных сосудов — эндотелий (илл.
277Б).
Расположение целомических полостей у Bila-
teria может быть различным. Олигомерные формы
имеют 1-3 пары полостей, расположенных одна за
другой по сторонам от кишки (Echiura, Sipuncula, Ten-
taculata, Chaetognatha, Echinodermata), причём пере-
дний целом может быть непарным (Hemichordata).
У полимерных форм с истинной сегментацией име-
ется большое число попарно расположенных цело-
мических мешков (Articulata, Астата, Craniota).
У Nemertini, помимо системы кровеносных со-
судов (которую сейчас рассматривают как производ-
ное парных туловищных целомов), имеется непар-
ная дорсальная целомическая полость, окружающая
хобот. У Mollusca целом сохраняется в области го-
над и сердца (гоноцель и перикард). По характеру
морфогенеза эти полости отличаются от парных це-
ломических мешков (см. выше), что дает основание
предполагать независимое и неоднократное развитие
целомических полостей.
С целомическими полостями у Bilateria часто
бывают связаны выделительная (метанефридии) и
половая системы (илл. 280, 285).
http://jurassic.ru/

Трёхслойные Eumetazoa
209
Илл. 278. Варианты расположения нерастяжимых волокон,
окружающих цилиндрическое тело с жидкостью под
давлением.
А — ортогональное расположение (продольные и кольцевые волокна).
Б — спиральное расположение (под углом к продольной оси тела). Во
втором случае тело может вытягиваться и сокращаться, а также изги-
баться без «излома». Оба варианта расположения волокон допускают
скручивание тела вокруг продольной оси. По Wainwright (1988).
Циркуляторные системы
У взрослых «Coelenterata» имеется единствен-
ная заполненная жидкостью полость — гастровас-
кулярная система, возникающая на основе кишки и
выполняющая транспортную и гидроскелетную фун-
кции (илл. 218, 241). Сходная ситуация наблюдается
у крупных плоских червей. У Bilateria, помимо киш-
ки,
могут возникать ещё три системы полостей: (1)
кровеносная система, (2) псевдоцель и (3) целом.
Кровеносные сосуды и псевдоцель представляют
собой заполненные жидкостью пустоты во внекле-
точном матриксе и служат, прежде всего, для распре-
деления питательных веществ, газообмена и транс-
порта отходов метаболизма. Их значение как гидро-
статического скелета было рассмотрено выше. В от-
личие от первых двух систем, вторичная полость тела
(целом) ограничена настоящей эпителиальной тка-
нью и в этом отношении принципиально сходна с
гастроваскулярной системой. Целом используется
как место хранения продуктов обмена и развития
гамет, иногда участвует в газообмене и особенно ча-
сто выполняет гидростатическую опорную функцию.
В тех группах Bilateria, для которых исходной
является ацеломическая или псевдоцеломическая
организация (Plathelminthes, Gnathostomulida, Nemat-
helminthes и Kamptozoa), либо отсутствуют все три
системы, либо имеется только псевдоцель, который
может быть слабо выраженным (у большинства сво-
бодноживущих нематод) или достаточно обширным
(Rotatoria, Acanthocephala). Исключение составляют
некоторые паразитические Plathelminthes, у которых
развивается система каналов, иногда даже имеющих
собственную клеточную выстилку.
Кровеносная система гистологически являет-
ся производным от первичной полости тела, которая
у некоторых групп преобразуется в очень сложную
систему каналов. Отличие кровеносной системы от
псевдоцеля чисто топографическое: псевдоцель пред-
ставляет собой единое внеклеточное пространство
между стенкой тела и кишкой, в то время как крове-
носная система соответствует этому же внеклеточ-
ному пространству, но редуцированному до отдель-
ных лакун, синусов и сосудов. В зависимости от сте-
пени дифференцировки сосудов различают замкну-
тые и незамкнутые кровеносные системы. В замкну-
той кровеносной системе гемолимфа течёт по хоро-
шо оформленным приводящим и отводящим сосу-
дам, связанным капиллярами. В незамкнутой систе-
ме приводящие и отводящие сосуды связаны лакуна-
ми в межклеточном матриксе соединительной ткани.
Кровеносная система и каналы целомического
происхождения могут быть по-разному развиты в
различных группах целомических Bilateria. В част-
ности, у иглокожих имеется обширная система це-
ломических каналов, пронизывающих тело наряду с
кровеносными сосудами. У Euhirudinea целом реду-
цируется до системы каналов, которые у Gnathobdel-
liformes и Pharyngobdelliformes полностью замеща-
ют настоящие кровеносные сосуды. Однако у боль-
шинства целомических животных обе системы хо-
рошо развиты и работают согласованно, особенно
при обеспечении газообмена.
В тех группах (например, членистоногие), где
первичная и вторичная полости тела сливаются на
ранних этапах развития, происходит также частич-
ное объединение полости тела и кровеносной систе-
мы.
Как сказано выше, такое состояние часто обо-
значают как гемоцель или миксоцель.
Органы дыхания
В дыхательных органах Bilateria происходит
обмен 0
2
и С0
2
между внешней средой (водой или
воздухом) и гемолимфой. Диффузия газов осуществ-
ляется через легко проницаемые мембраны. Те же
физические принципы лежат в основе транспорта
газов циркуляторной системой от органов дыхания
к тканям: газы переносятся вместе с током гемолим-
фы,
а газообмен с тканями происходит путём диф-
фузии через мембраны.
Известно, что объём тела животного увеличи-
вается пропорционально кубу (третьей степени) ли-
нейных размеров, в то время как поверхность тела,
через которую происходит диффузия газов, увеличи-
вается медленнее, пропорционально квадрату (вто-
рой степени) линейных размеров. Поэтому увеличе-
ние размеров тела должно сопровождаться появле-
нием дополнительных поверхностей газообмена. У
http://jurassic.ru/

210
Илл.
279. Гистологический продольный срез через жабер-
ную нить с боковыми жаберными лепестками у форели.
Очень тонкий эпидермис обеспечивает лучшие условия газообмена.
W. Salvenmoser, Инсбрук.
крупных представителей Metazoa многократно и не-
зависимо возникают специализированные органы
дыхания. При этом органы, обеспечивающие транс-
порт газов (например, жаберные крышки, лёгочная
мускулатура, воздушные мешки трахейной системы)
могут быть анатомически отделены от структур, в
которых происходит собственно газообмен (напри-
мер,
жаберные лепестки (илл. 279), лёгочные альве-
олы, трахеолы).
Напротив, мелкие Bilateria обладают более вы-
годным с точки зрения газообмена соотношением
поверхности и объёма, что позволяет им обходиться
более простыми органами дыхания. В то же время
при малых размерах тела более острыми становятся
проблема осморегуляции для водных форм и пробле-
ма сохранения влаги — для наземных Bilateria.
У крупных представителей Plathelminthes дистанция
диффузии газов сокращается благодаря уплощению тела и
наличию сильно разветвлённой кишки. Активная локомо-
ция (например, путём изгибания тела) способствует пере-
Трёхслойные Eumetazoa
мешиванию наружной среды
и
внутренних жидкостей тела,
что также благоприятствует газообмену.
Органы выделения
Строение органов выделения и осморегуляции
у Bilateria тесно связано с организацией полости тела
и с размерами животного. Эти органы, выполняю-
щие функции регуляции водного баланса и ионного
состава жидкостей тела, а также выведения различ-
ных отходов метаболизма, необходимы всем Bilateria.
Простейшим способом удаления конечных про-
дуктов метаболизма является, по-видимому, отложе-
ние их в особых клетках (например, «конкремент-
ные гранулы» у бескишечных турбеллярий) или спе-
циализированных тканях (у пауков). Так называемые
дермонефридии, недавно найденные у Paratomella
rubra (Acoelomorpha), возможно, также представля-
ют собой примитивные выделительные клетки.
Имеются два принципиально различных типа
органов выделения: (1) фильтрующие почки, кото-
рые могут быть представлены протонефридиями или
метанефридиями (илл. 280, 282, 283), и (2) секрети-
рующие почки — придатки пищеварительного трак-
та Arachnida и Antennata, так называемые мальпиги-
евы сосуды (илл. 281).
Для работы фильтрующих почек обычно не-
обходим большой объём жидкости. В них вначале
образуется первичная моча, состав которой перед
окончательным выведением из организма изменяет-
ся благодаря реабсорбции и секреции в выводящих
каналах.
В метанефридиальной системе первичная моча об-
разуется путём фильтрации под давлением из кровеносной
системы в полость целума. Фильтром служит внеклеточ-
ный матрикс специализированных клеток целомического
эпителия — подоцитов (илл. 211
А,
280В,Г). Повышенное
давление в кровеносных сосудах создаётся сокращением
их мускулатуры. В протонефридиальной системе моча
фильтруется в выводящий канал протонефридия прямо из
тканевой жидкости (а не из кровеносных сосудов) через
внеклеточный матрикс терминальной или канальной клет-
ки (илл. 280А,Б, 282). Предполагают, что разность давле-
ния,
необходимая для фильтрации, возникает благодаря
работе жгутиков терминальных клеток (циртоцитов). Пос-
ледние могут быть мультицилиарными или моноцилиар-
ными (так наз. соленоциты).
В исходном варианте метанефридиальной системы
структуры, обеспечивающие фильтрацию, топографичес-
ки не связаны с собственно выделительными органами
(илл.
280В,Г). Это означает, что первичная моча выделят-
ся непосредственно в целом, смешиваясь с целомической
жидкостью. Затем содержимое целбма поступает через
ресничную воронку
в
выводящий канал метанефридия, где
осуществляются реабсорбция и секреция, т.е. формирует-
ся вторичная моча. Каналы метанефридиев обычно окру-
жены кровеносными сосудами.
http://jurassic.ru/
Трёхслойные Eumetazoa
211
реабсорбция
и секреция
фильтрация
первичной
мочи
в
целом
кровь
гаметы
О
пластинка
нефропор
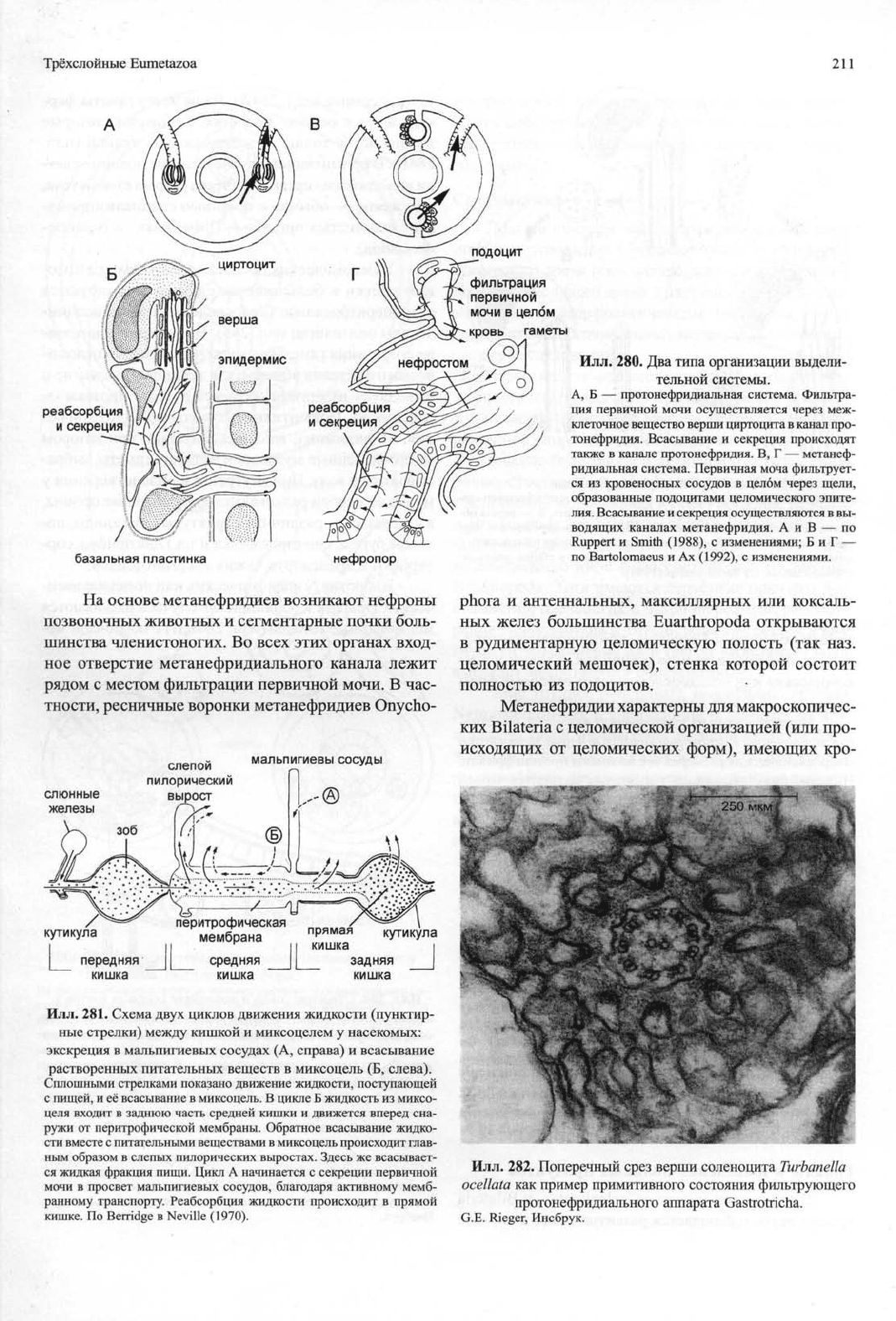
Илл.
280. Два типа организации выдели-
тельной системы.
А, Б — протонефридиальная система. Фильтра-
ция первичной мочи осуществляется через меж-
клеточное вещество верши циртоцита в канал про-
тонефридия. Всасывание и секреция происходят
также в канале протонефридия. В, Г — метанеф-
ридиальная система. Первичная моча фильтрует-
ся из кровеносных сосудов в целом через щели,
образованные подоцитами целомического эпите-
лия.
Всасывание и секреция осуществляются в вы-
водящих каналах метанефридия. А и В — по
Ruppert и Smith (1988), с изменениями; Б и Г —
по Bartolomaeus и Ах (1992), с изменениями.
На основе метанефридиев возникают нефроны
позвоночных животных и сегментарные почки боль-
шинства членистоногих. Во всех этих органах вход-
ное отверстие метанефридиального канала лежит
рядом с местом фильтрации первичной мочи. В час-
тности, ресничные воронки метанефридиев Onycho-
слепои
пилорический
вырост
игиевы
сосуды
/~**==^
перитрофическая
кутикула
мембрана
передняя
||
средняя
|
кишка кишка
прямая
кишка
кутикула
задняя
кишка
Илл.
281. Схема двух циклов движения жидкости (пунктир-
ные стрелки) между кишкой и миксоцелем у насекомых:
экскреция в мальпигиевых сосудах (А, справа) и всасывание
растворенных питательных веществ в миксоцель (Б, слева).
Сплошными стрелками показано движение жидкости, поступающей
с пищей, и её всасывание в миксоцель. В цикле Б жидкость из миксо-
целя входит в заднюю часть средней кишки и движется вперед сна-
ружи от перитрофической мембраны. Обратное всасывание жидко-
сти вместе с питательными веществами в миксоцель происходит глав-
ным образом в слепых пилорических выростах. Здесь же всасывает-
ся жидкая фракция пищи. Цикл А начинается с секреции первичной
мочи в просвет мальпигиевых сосудов, благодаря активному мемб-
ранному транспорту. Реабсорбция жидкости происходит в прямой
кишке. По Berridge в Neville (1970).
phora и антеннальных, максиллярных или коксаль-
ных желез большинства Euarthropoda открываются
в рудиментарную целомическую полость (так наз.
целомический мешочек), стенка которой состоит
полностью из подоцитов.
Метанефридии характерны для макроскопичес-
ких Bilateria с целомической организацией (или про-
исходящих от целомических форм), имеющих кро-
Илл.
282. Поперечный срез верши соленоцита
Turbanella
ocellata
как пример примитивного состояния фильтрующего
протонефридиального аппарата Gastrotricha.
G.E. Rieger, Инсбрук.
http://jurassic.ru/

212
Трёхслойные Eumetazoa
Илл.
283. Эволюция циртоцитов.
А — примитивное состояние: фильтрующий аппарат образован ще-
лями в цилиндрическом придатке самого циртоцита. Б — верша об-
разована переплетением пальцевидных выростов циртоцита и пер-
вой канальной клетки. В — моноцилиарный циртоцит (соленоцит); у
взрослых кольчатых червей соленоциты впадают в общие нефриди-
альные каналы. Из Bartolomaeus (1994).
веносную систему. Напротив, протонефридии встре-
чаются преимущественно у личинок, а также у аце-
ломических или псевдоцеломических взрослых форм
размером не более нескольких миллиметров.
Исключениями из этого правила
являются,
в частно-
сти,
приапулиды
и
некоторые кольчатые
черви,
которые при
макроскопических размерах
всё же
имеют протонефридии.
Наличие протонефридиев у взрослых кольчатых червей
практически всегда сочетается
с
отсутствием кровеносной
системы.
Протонефридии считаются наиболее примитив-
ными органами выделения (илл. 283). Однако они
могут возникать и вторично, что было показано не-
давними исследованиями мелких кольчатых червей
и карликовых самцов Bonellia. Для кольчатых чер-
вей известны также случаи, когда циртоциты личи-
нок непосредственно превращаются в метанефриди-
альные подоциты взрослых особей. Эти факты по-
зволяют предположить, что метанефридии взрослых
целомических Spiralia и протонефридии их личинок
происходят из одного и того же зачатка.
Секретирующие почки — это мальпигиевы
сосуды, развитые, например, у насекомых (илл. 857).
Их функционирование основано на перемещении
жидкости в кишке, как показано на илл. 281.
Гонады и выведение половых продуктов
В отличие от многих «Coelenterata», у Bilateria
крайне редко наблюдается развитие гамет в составе
гастродермиса (илл. 284А). Чаще всего гаметы фор-
мируются в особых полостях — гонадах, которые
возникают в толще мезодермальных тканей (илл.
284Б).
В примитивном случае женские половые клет-
ки выводятся из организма через разрыв стенки тела,
а мужские — обычно с помощью специализирован-
ных железистых органов («Turbellaria» — Nemerto-
dermatida).
У целомических представителей Bilateria поло-
вые клетки в большинстве случаев формируются
ретроперитонеально (непосредственно под целоми-
ческим эпителием, илл. 285). На определённом эта-
пе созревания гаметы выходят через разрыв целоми-
ческого эпителия в целом, где завершают развитие и
выводятся из организма через особые протоки —
гонодукты. Примитивным способом оплодотворе-
ния, по-видимому, является внешний, при котором
многочисленные мужские и женские гаметы выбра-
сываются в воду. При внутреннем оплодотворении у
мужских особей развиваются копулятивные органы,
а у женских—различные структуры (влагалища, по-
ловые бурсы, семяприемники и т.д.) для приёма, сор-
тировки и транспорта чужих сперматозоидов.
Напротив, у ацеломических или псевдоцеломи-
ческих Bilateria в большинстве случаев развиваются
мешковидные гонады (илл. 284Б,В) с оболочкой ме-
Ш Ш.
Илл.
284. Строение гонад и выведение половых клеток у
ацеломических форм. На схеме изображена женская половая
система; мужская половая система устроена принципиально
так же.
А — гаметы развиваются в базальной зоне гастродермиса (некото-
рые Nemertodermatida, Plathelminthes) или в мезодермальной соеди-
нительной ткани (напр., у Acoela и некоторых Prolecithophora). Жен-
ские половые клетки в примитивном случае выводятся через разрыв
стенки тела; возможно также выведение через кишку. Б — гаметы
развиваются в мешковидных гонадах, стенки которых формируются
в ходе постэмбрионального развития из мезодермальных клеток (ниж-
ний ряд рисунков). С гонадами могут быть связаны выводные прото-
ки эктодермального происхождения. Половое отверстие чаще всего
непарное, расположено медианно на брюшной стороне тела. R. Rieger,
Инсбрук.
http://jurassic.ru/
Трёхслойные Eumetazoa
213
вывод
через
разрыв
стенки
тела
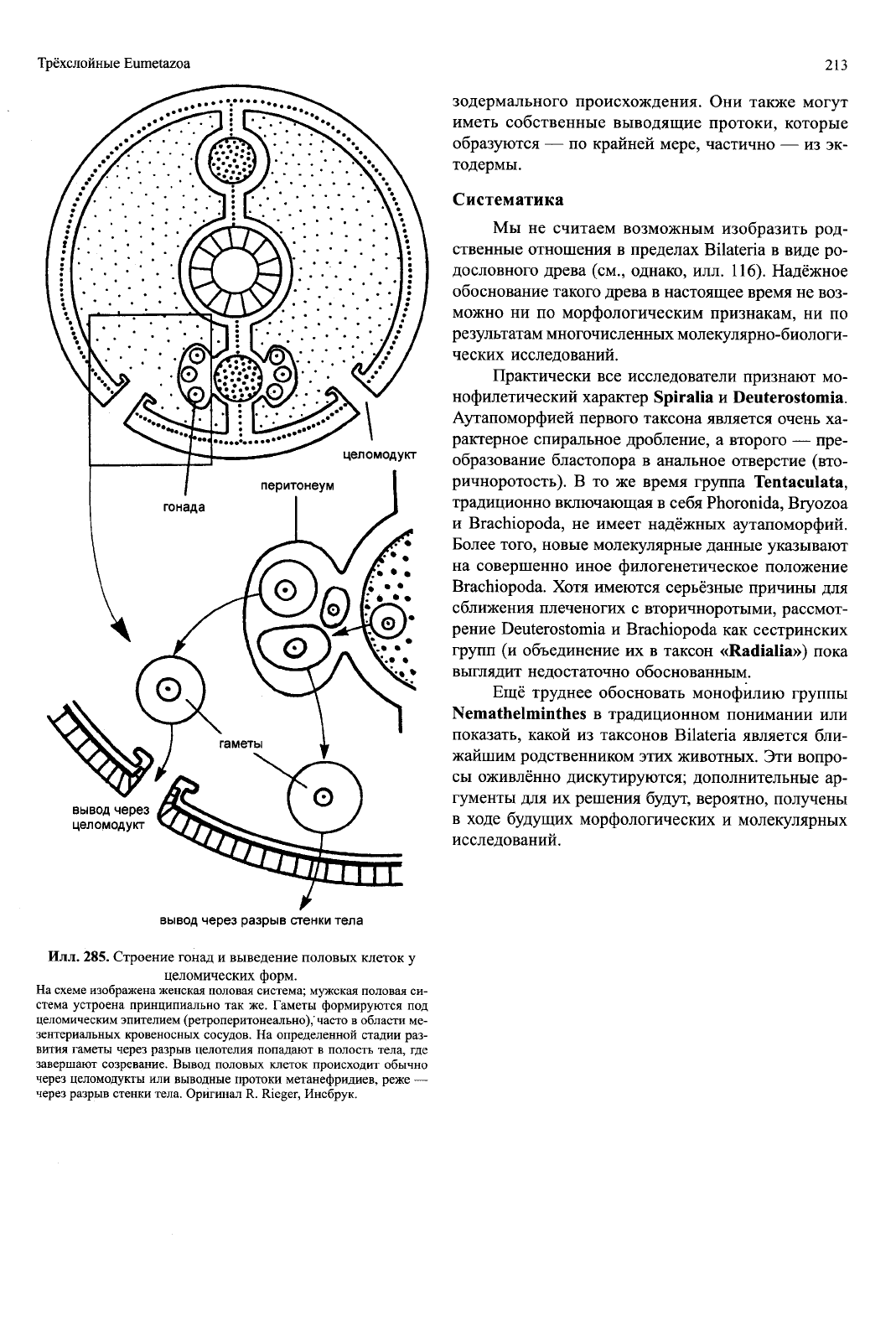
Илл.
285. Строение гонад и выведение половых клеток у
целомических форм.
На схеме изображена женская половая система; мужская половая си-
стема устроена принципиально так же. Гаметы формируются под
целомическим эпителием (ретроперитонеально),' часто в области ме-
зентериальных кровеносных сосудов. На определенной стадии раз-
вития гаметы через разрыв целотелия попадают в полость тела, где
завершают созревание. Вывод половых клеток происходит обычно
через целомодукты или выводные протоки метанефридиев, реже —
через разрыв стенки тела. Оригинал R. Rieger, Инсбрук.
зодермального происхождения. Они также могут
иметь собственные выводящие протоки, которые
образуются — по крайней мере, частично — из эк-
тодермы.
Систематика
Мы не считаем возможным изобразить род-
ственные отношения в пределах Bilateria в виде ро-
дословного древа (см., однако, илл. 116). Надёжное
обоснование такого древа в настоящее время не воз-
можно ни по морфологическим признакам, ни по
результатам многочисленных молекулярно-биологи-
ческих исследований.
Практически все исследователи признают мо-
нофилетический характер Spiralia и Deuterostomia.
Аутапоморфией первого таксона является очень ха-
рактерное спиральное дробление, а второго — пре-
образование бластопора в анальное отверстие (вто-
ричноротость). В то же время группа Tentaculata,
традиционно включающая в себя Phoronida, Bryozoa
и Brachiopoda, не имеет надёжных аутапоморфий.
Более того, новые молекулярные данные указывают
на совершенно иное филогенетическое положение
Brachiopoda. Хотя имеются серьёзные причины для
сближения плеченогих с вторичноротыми, рассмот-
рение Deuterostomia и Brachiopoda как сестринских
групп (и объединение их в таксон «Radialia») пока
выглядит недостаточно обоснованным.
Ещё труднее обосновать монофилию группы
Nemathelminthes в традиционном понимании или
показать, какой из таксонов Bilateria является бли-
жайшим родственником этих животных. Эти вопро-
сы оживлённо дискутируются; дополнительные ар-
гументы для их решения будут, вероятно, получены
в ходе будущих морфологических и молекулярных
исследований.
http://jurassic.ru/
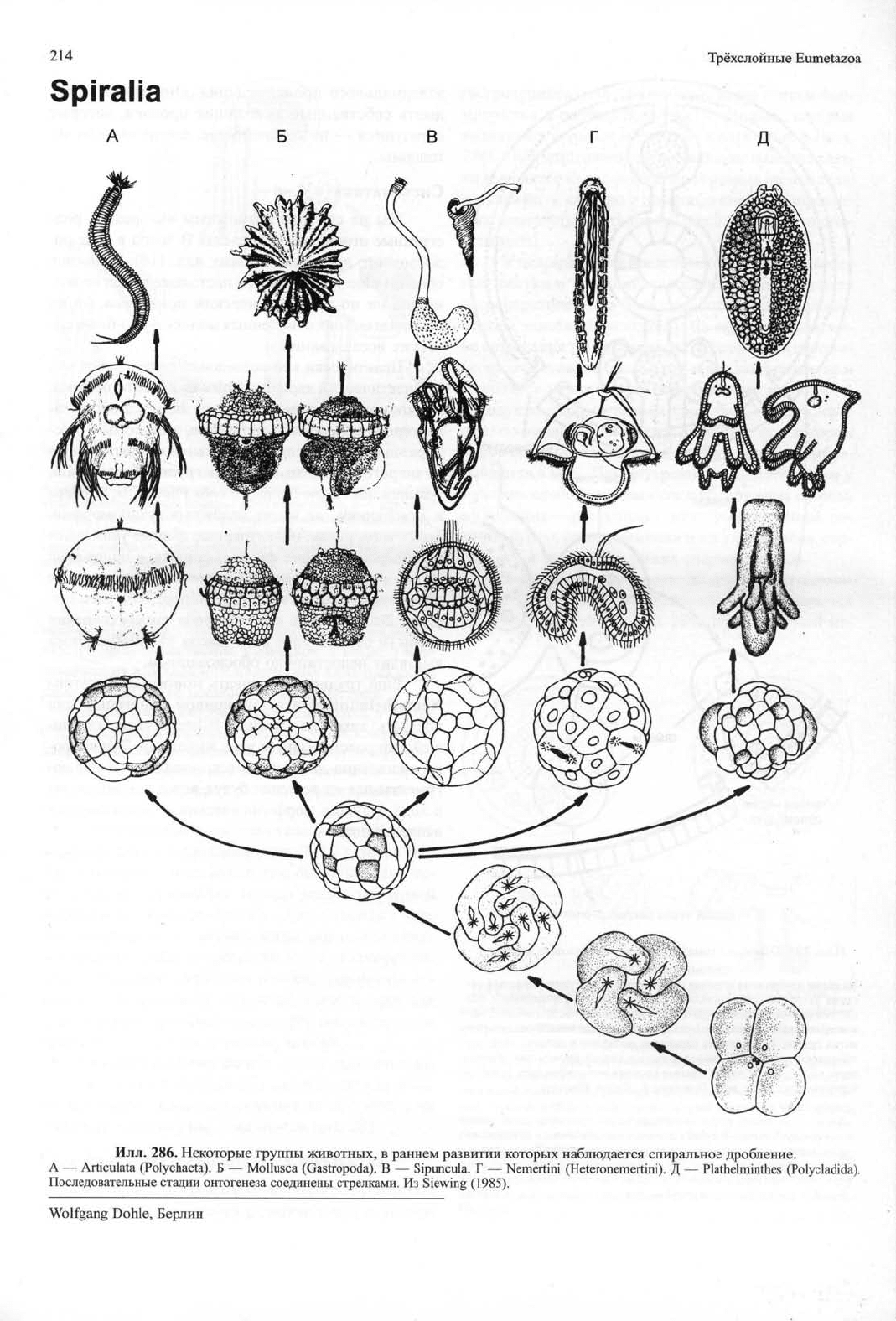
214 Трёхслойные Eumetazoa
Spiralia
А
Б В Г Д
Илл.
286. Некоторые группы животных, в раннем развитии которых наблюдается спиральное дробление.
А — Articulata (Polychaeta). Б — Mollusca (Gastropoda). В — Sipuncula. Г — Nemertini (Heteronemertini). Д — Plathelminthes (Polycladida).
Последовательные стадии онтогенеза соединены стрелками. Из Siewing (1985).
Wolfgang Dohle, Берлин
http://jurassic.ru/
Spiralia
215
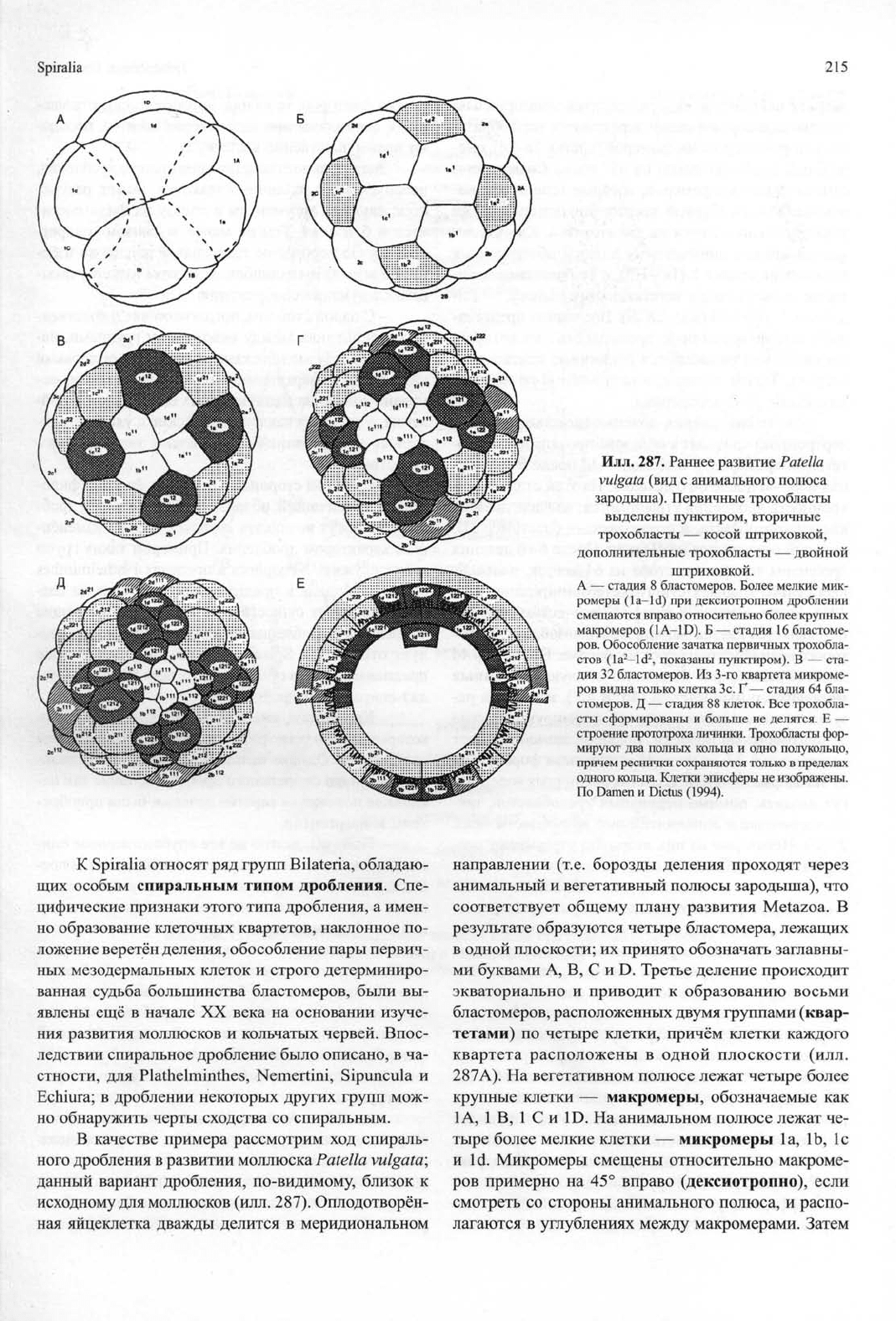
Илл.
287. Раннее развитие Patella
vulgata (вид с анимального полюса
зародыша). Первичные трохобласты
выделены пунктиром, вторичные
трохобласты — косой штриховкой,
дополнительные трохобласты — двойной
штриховкой.
А — стадия 8 бластомеров. Более мелкие мик-
ромеры (la-Id) при дексиотропном дроблении
смещаются вправо относительно более крупных
макромеров (1A-1D). Б — стадия 16 бластоме-
ров.
Обособление зачатка первичных трохобла-
стов (la
2
—Id
2
, показаны пунктиром). В — ста-
дия 32 бластомеров. Из 3-го квартета микроме-
ров видна только клетка Зс. Г— стадия 64 бла-
стомеров. Д — стадия 88 клеток. Все трохобла-
сты сформированы и больше не делятся. Е —
строение прототроха личинки. Трохобласты фор-
мируют два полных кольца и одно полукольцо,
причем реснички сохраняются только в пределах
одного кольца. Клетки эписферы не изображены.
По Damen и Dictus (1994).
К Spiralia относят ряд групп Bilateria, обладаю-
щих особым спиральным типом дробления. Спе-
цифические признаки этого типа дробления, а имен-
но образование клеточных квартетов, наклонное по-
ложение веретён деления, обособление пары первич-
ных мезодермальных клеток и строго детерминиро-
ванная судьба большинства бластомеров, были вы-
явлены ещё в начале XX века на основании изуче-
ния развития моллюсков и кольчатых червей. Впос-
ледствии спиральное дробление было описано, в ча-
стности, для Plathelminthes, Nemertini, Sipuncula и
Echiura; в дроблении некоторых других групп мож-
но обнаружить черты сходства со спиральным.
В качестве примера рассмотрим ход спираль-
ного дробления в развитии моллюска
Patella vulgata;
данный вариант дробления, по-видимому, близок к
исходному для моллюсков (илл. 287). Оплодотворён-
ная яйцеклетка дважды делится в меридиональном
направлении (т.е. борозды деления проходят через
анимальный и вегетативный полюсы зародыша), что
соответствует общему плану развития Metazoa. В
результате образуются четыре бластомера, лежащих
в одной плоскости; их принято обозначать заглавны-
ми буквами А, В, С и D. Третье деление происходит
экваториально и приводит к образованию восьми
бластомеров, расположенных двумя группами (квар-
тетами) по четыре клетки, причём клетки каждого
квартета расположены в одной плоскости (илл.
287А).
На вегетативном полюсе лежат четыре более
крупные клетки — макромеры, обозначаемые как
1А, 1 В, 1 С и 1D. На анимальном полюсе лежат че-
тыре более мелкие клетки — микромеры la, lb, 1с
и Id. Микромеры смещены относительно макроме-
ров примерно на 45° вправо (дексиотропно), если
смотреть со стороны анимального полюса, и распо-
лагаются в углублениях между макромерами. Затем
http://jurassic.ru/