Вершигора А.Ю., Пастер Є.У., Колибо Д.В. Імунологія
Подождите немного. Документ загружается.


51
стимуляції кролів антигеном сальмонел плазмоцити виявляються в реґіонарних лімфовузлах на
четверту добу, їх кількість збільшується вдвічі на шосту — восьму добу, а на чотирнадцяту —
зменшується до вихідного рівня. Під час вторинної відповіді вони з'являються вже через дві доби, а
в період між другою і четвертою добою їх кількість збільшується вдвічі кожні 12 год.
У динаміці утворення антитіл виділяють чотири фази:
1) фаза спокою (лаг-фаза, індуктивна фаза) — з моменту надходження антигену в організм до появи
антитіл у сироватці крові;
2) фаза наростання титру антитіл (лог-фаза, продуктивна фаза) — від появи антитіл до моменту
досягнення їх максимальної кількості;
3) фаза стабілізації, в якій титр антитіл залишається незмінним;
4) фаза зниження рівня антитіл унаслідок виведення їх (у комплексах з антигеном) або завдяки
катаболізуванню.
Тривалість кожної фази і рівень антитіл залежить від природи антигену та особливостей організму
(за умови використання однакових доз та способу введення антигену).
Повторна імунізація через кілька тижнів або місяців змінює динаміку імунної відповіді. Латентний
період і період наростання титру антитіл
стають коротшими, а фази стабілізації й
зниження титру — тривалішими. Титр
антитіл значно вищий (у 10 і більше разів),
досягає максимуму швидше і зберігається
на високому рівні довше. Крім того,
підвищується афінність антитіл —
спорідненість до антигену. В складі антитіл
переважають IgG, тоді як за первинної
імунної відповіді значну частку становлять
IgM, синтез яких передує появі IgG (мал.
86).
Підсилена гуморальна вторинна імунна
відповідь на Т-залежні антигени свідчить
про утворення в організмі не лише В-клітин
пам'яті, а й Т-клітин пам'яті з хелперною
активністю.
Прискорений розвиток вторинних
клітинних реакцій, таких як відторгнення
вторинного трансплантата, свідчить також
про утворення Т-клітин пам'яті з
цитотоксичною функцією та їхню роль у
формуванні вторинної імунної відповіді
клітинного типу. Отже, імунна відповідь
клітинного і гуморального типу
супроводжується утворенням ефекторних
клітин та клітин пам'яті. Основою цих
процесів є проліферація і диференціювання
антигенспецифічних клонів. Клітини
пам'яті можуть бути окремою лінією диференціювання активованих антигеном клітин, що
з'являються паралельно з утворенням ефекторів, але в меншій кількості, бо клітини пам'яті — не
повністю зрілі ефектори, диференціювання яких припинилося у зв'язку зі зменшенням кількості
антигену в організмі. Очевидно, що обидва механізми контролюють утворення різних субпопуляцій
клітин пам'яті. Розрізняють клітини центральної пам'яті, або центральні клітини пам'яті, які
рециркулюють у лімфі та крові між вторинними лімфоїдними органами, і клітини периферичної
пам'яті, або ефекторні клітини пам'яті, здатні активуватися на АПК у тканинах. Клітини центральної
пам'яті відрізняються від клітин периферичної пам'яті експресією хемокінового рецептора CCR7,
який зумовлює їхній рух до вторинних лімфоїдних органів.
Вважають, що ефекторні Т-клітини пам'яті після зустрічі з антигеном (у периферичних тканинах) у
відповідь на активацію швидко продукують цитокіни або виділяють цитотоксичні гранули,
опосередковуючи запальні або цитотоксичні реакції. Центральні Т-клітини пам'яті опосередковують
вторинні імунні відповіді до повторно введених (чи тих, що проникли природним шляхом)
антигенів.
Хемокінові рецептори, характерні для наївних Т-клітин, на Т-клітинах периферичної пам'яті
містяться у значно вищій кількості, ніж на ефекторних Т-клітинах, що підтверджує припущення про
те, що Т-клітини пам'яті — попередники ефекторних клітин, які уникли загибелі.
Для диференціювання клітин пам'яті на ефектори потрібна менша кількість сигналів, ніж для
наївних клітин, що і зумовлює прискорений розвиток імунної відповіді після наступного
потрапляння антигену в організм.
Ще одним поясненням існування тривалого імунітету після імунізації є уявлення про персистенцію
антигену на ФДК і ІДК у лімфоїдних структурах упродовж кількох років. Така персистенція
антигену потрібна, з одного боку, для підтримування життєдіяльності певних типів клітин пам'яті, а
з іншого — дає змогу постійно підтримувати процес активації, що супроводжується проліферацією
та диференціюванням, у відносно невеликої кількості наївних Т- і В-клітин.
Завдяки існуванню імунної пам'яті іноді можна спостерігати явище конкуренції антигенів, які були
введені послідовно. Це створює певні прогалини в лінії імунного захисту. Відомо, що за вторинної
імунної відповіді до реакції переважно залучаються клітини пам'яті, які усувають від участі в цьому
процесі наївні клітини, скажімо конкуренцією за сайти зв'язування на АПК. Це може призводити до
несподіваних наслідків. Наприклад, людина, яка щойно перенесла інфекцію грипу, зумовлену
вірусом одного штаму, може легко заразитися аналогічним вірусом іншого штаму. Причому в разі
зараження новим штамом буде зумовлена імунна відповідь переважно на ті антигенні детермінанти,
які є спільними з детермінантами вірусу першого штаму. Особливі, характеристичні епітопи
другого штаму розпізнаватимуться гірше, оскільки наївні клітини майже не будуть залучені до
імунної відповіді. Отже, розвиток імунної відповіді та створення тривалого імунітету до вірусу
другого штаму не таке ефективне, як до вірусу першого штаму.
11.6. ЕФЕКТОРНІ МЕХАНІЗМИ ІМУНІТЕТУ.
Антигенспецифічні ефекторні фактори специфічної імунної відповіді (антитіла та Т-лімфоцити-
можуть самостійно реалізувати захисні фуекції нейтралізацією шкідливої дії патогенів чи їхніх
продуктів (токсинів) та знищенням власних клітин організму, що змінили антигенні властивості
внаслідок інфекції або злоякісної трансформації. Антитіла та продуковані Т-лімфоцитами цитокіни
здатні також підсилювати ефекторні механізми природного імунітету — фагоцитоз,
комплементзалежний цитоліз, клітиноопосередкований цитоліз. Завдяки високій специфічності
щодо антигену зазначені фактори зумовлюють вибірковість дії та підвищують ефективність
ефекторних механізмів природного імунітету. Взаємодія ефекторних механізмів адаптивного та
природного імунітету значно посилює захист організму і забезпечує швидшу елімінацію
чужорідних антигенів чи їхніх носіїв.
Нейтралізація як опосередкований антитілами ефекторний механізм. Нейтралізація антигенів
за допомогою антитіл — важливий самостійний ефекторний механізм та початковий етап інших
ефекторних реакцій. Як самостійний ефекторний механізм нейтралізація реалізується під час
взаємодії антитіл з ізольованими антигенними молекулами, наприклад бактеріальними токсинами.
При цьому ефективними є антитіла, спрямовані як до функціональної групи в молекулі токсину, що
відповідає за прояв ним біологічної дії (токсичного ефекту), так і до акцепторної (рецепторної)
групи, що забезпечує його фіксацію на клітині-мішені.
Нейтралізувальна функція антитіл може виявлятися і в їхній здатності блокувати компоненти
мікробної клітини, що відповідають за прикріплення її до клітин хазяїна. Ця функція здебульшого
властива антитілам, які не спроможні залучати в реакцію інші ефекторні фактори (комплемент,
фагоцити, НК-клітини) або ця здатність у них виражена слабко, що може бути зумовлено
властивостями самих антитіл (наприклад, відсутністю здатності зв'язувати комплемент) чи
ефекторних клітин (відсутністю рецепторів до відповідних антитіл). Так, локалізовані в секретах
слизових оболонок антитіла ізотипу sІgA блокують компоненти клітин патогенних мікробів
(адгезини), відповідальні за прилипання (адгезію) до епітелію, запобігаючи таким чином розвитку
інфекцій, що асоціюються зі слизовими оболонками.
Ефекторні механізми, основані на фагоцитозі. Фагоцитоз можна вважати найважливішим
фактором природного імунітету, якому належить провідна роль в антимікробному, зокрема
антибактеріальному, захисті. Основою його є розпізнавання фагоцитами за допомогою примітивних
(неспецифічних) механізмів патогенних бактерій, поглинання їх і розщеплення за участю
бактерицидних факторів (та гідролітичних ферментів). Однак патогенні бактерії використовують
найрізноманітніші способи для того, щоб протидіяти розпізнаванню і поглинанню їх фагоцитами
або протистояти дії бактерицидних субстанцій усередині фагоцита (докладніше ці способи
викладено у розд. 15).
Фактори адаптивного імунітету підвищують ефективність фагоцитарної реакції привнесенням
елемента специфічності для розпізнавання об'єктів фагоцитозу або підсиленням бактерицидних
механізмів. Підвищення ефективності розпізнавання досягається за допомогою опсонізації об'єктів
фагоцитозу (бактерій та їхніх токсинів) антитілами субкласів IgG1 та IgG3 та компонентом
комплементу С3b (що фіксується на комплексах антиген — антитіло). Такі опсонізовані об'єкти
розпізнаються завдяки наявності на фагоцитах Fcγ-рецепторів (FcγR1 — на макрофагах, FcγRII —
на нейтрофілах), які зв'язуються з антитілами, та рецепторів для комплементу (CR), що розпізнають
переважно С3b.
Основою стимулювальної дії цитокінів на фагоцитоз є підсилення бактерицидних властивостей
фагоцитів. Продуковані Тх1-клітинами цитокіни, насамперед ІФН-γ зумовлюють активацію
макрофагів, яка супроводжується посиленням утворення різних факторів бактерицидності, що
сприяє вбивству та розщепленню внутрішньоклітинних патогенів.
Антитіла, специфічні до поверхневих антигенів клітин-мішеней, зв'язуючись із ними, утворюють
імунні комплекси, які фіксують комплемент і запускають його активацію за класичним шляхом,
різко підвищуючи ефективність цитолізу (порівняно із зумовленим активацією комплементу без
участі антитіл). Цей механізм цитолізу відіграє основну роль у лізисі еритроцитів під час
переливання несумісної крові та під час аутоімунної патології. Однак значення його в
антимікробному імунітеті набагато менше, ніж це вважалося раніше, й обмежується, як нині
доведено, лише інфекціями, що спричинені збудниками роду нейсерія.
Ефекторні механізми, основані на клітиноопосередкованому цитолізі. Описаний вище механізм
цитоксичності за участю Т-клітин є формою імунного цитолізу, пов'язаною зі специфічним
розпізнаванням кілерними клітинами клітин-мішеней. Відомі й інші форми клітино-
опосередкованого цитолізу, що ґрунтуються на функціонуванні клітин природного імунітету,
ефекторні функції яких значно підсилюються факторами імунної відповіді – антитілами та
цитокінами.
Одним із варіантів природного цитолізу, механізми якого «вдосконалюються» антитілами і
цитокінами, є НК-цитотоксичність. НК-клітини використовуї зовсім інший механізм розпізнавання
клітин-мішеней, ніж цитотоксичні Т-лімфоцити (див. розд. 2). Реалізація обох видів
цитотоксичності залежить від експресії на клітинах-мішенях молекул МНС І, однак розпізнавання
цих молекул ЦТЛ та НК-клітинами індукує цілком протилежні сигнали — відповідно «дозвіл» та
«заборону» на вбивство. Цитотоксична активність НК-клітин суворо контролюється інгібіторними
рецепторами (КІР), що реалізують інгібіторну функцію через розпізнавання аутологічних молекул
МНС І.
НК-клітини ідентифікують клітини-мішені, розпізнаючи своїми рецепторами конститутивно
експресовані на їхній поверхні ліганди, наприклад залишки манози на молекулах глікопротеїнів та
гліколіпідів, і вбивають їх лише за умови втрати ними (внаслідок інфікування чи трансформації)
експресії молекул МНС І (природна НК-цитотоксичність).
НК-цитотоксичність може модифікуватися антитілами підкласів IgG1 та IgG3 завдяки наявності на
НК-клітинах Fc-peцепторів типу FcγRIII (CD16), за допомогою яких кілерні клітини зв'язуються з
антитілами, що опсонізували клітину-мішень, активуються і зумовлюють цитоліз
(антитілозалежна клітинна цитотоксичність — АЗКЦ). Залучення Fc-рецепторів до процесу
розпізнавання значно підвищує ефективність цитолізу.
Слід зазначити, що активація НК-клітин як через зв'язування ліганду для Fc-рецептора, так і через
взаємодію з численними лігандами під час здійснення природної цитотоксичності залежить від
тирозинової протеїнкінази. Однак активаційні сигнали, що надходять цими двома шляхами, різні. У
разі АЗКЦ активація НК-клітин залежить від фосфоінозитид-3-кінази, а в разі природної
цитотоксичності — від протеїнкінази С. Незважаючи на відмінності в розпізнаванні клітин-мішеней
та у трансдукції активаційних сигналів, обидва види цитотоксичності здійснюються НК-клітинами з
використанням одного механізму — перфоринзалежного цитолізу.
Цитотоксична активність НК-клітин значно підвищується також під дією цитокінів, насамперед ІЛ-
2. Активовані ІЛ-2 НК-клітини, що становлять більшість лімфокінактивованих клітин — ЛАК,
проліферують, значно підвищуючи свій цитотоксичний потенціал, і набувають здатності зумовити
лізис багатьох клітин-мішеней, в тому числі щойно виділених пухлинних клітин деяких типів.
Оскільки щойно виділені пухлинні клітини досить часто експресують молекули МНС І і не
вбиваються неактивованими НК-клітинами через інгібування кілінгу КІР-рецепторами, то
припускають, що ефект ІЛ-2 може зумовлюватися зняттям КІР-інгібування. Отримання ЛАК-клітин
унаслідок активації in vitro інтерлейкіном 2 лімфоцитів хворого з наступним введенням їх в
організм, як сподівалися буде ефективним способом протипухлинної терапії, однак клінічні
випробування поки що бажаних результатів не дали.
НК-клітини і ЦТЛ відіграють важливу роль у забезпеченні противірусного та протипухлинного
імунітету знищенням інфікованих вірусами та злоякісно трансформованих клітин, функціонуючи як
дві складові, що за певних умов доповнюють одна одну.
Ще одним прикладом ефективної сумісної дії факторів набутого і природного імунітету є
підвищення цитотоксичності еозинофілів, механізм якої пов'язаний з екзоцитозом гранул і
звільненням активних субстанцій (позаклітинний цитоліз). Озброєні антитілами та активовані ІЛ-5,
продукованим Тх2, еозинофіли відіграють важливу роль при паразитарних інфекціях, наприклад
шистосомозі та ін. Зв'язуючись за допомогою FcεRII або FcαRII з опсонізованими IgE або IgA
гельмінтами, еозинофіли починають синтезувати та секретувати токсини (ЕСР, МВР), які уражують
патогени. Залученню еозинофілів до місця локалізації гельмінта сприяють медіатори запалення, які
звільняються внаслідок дегрануляції базофілів і мастоцитів, активованих зв'язуванням фіксованим
на їхній поверхні (на FcεRI) IgE паразитарного антигену.
Таким чином, ефекторні клітини і молекули адаптивної ланки імунної відповіді здійснюють
імунний захист як самостійно, так і в поєднанні з ефекторними факторами природної ланки, завдяки
чому значно розширюється арсенал ефекторних реакцій імунітету й підвищується їхня
інтенсивність.
ВИСНОВКИ.
Імунна відповідь — реакція імунної системи на чужорідні агенти, спрямована на видалення їх з
організму. Вона включає дві ланки — природну та адаптивну, що формуються послідовно і
забезпечуються відповідно неспецифічними та специфічними до антигену факторами. Адаптивна
відповідь є наслідком низки метаболічних процесів, що здійснюються в АПК і лімфоцитах,
включаючи ідентифікацію антигенних молекул АПК, процесинг і переведення їх у доступну для
розпізнавання Т-лімфоцитами форму, активацію антигенспецифічних клонів лімфоцитів і синтез
ефекторів (антитіл та клітин) із чіткою реактивністю щодо цього антигену.
Тип адаптивної імунної відповіді залежить переважно від цитокінів, що продукуються АПК та
клітинами мікрооточення і спрямовують диференціювання СD4Т-клітин в Тх1 чи Тх2, які
запускають відповідно розвиток клітинної чи гуморальної імунної реакції. Основою гуморальної
імунної відповіді є активація В-лімфоцитів і синтез антитіл. У процесі відповіді відбувається
переключення синтезу антитіл з IgM на ефективніші в імунному захисті IgG та IgA (у слизових
оболонках), підвищується афінність антитіл — спорідненість їх до антигену. Антитіла реалізують
захисну функцію різними способами: блокують здатність патогенів прикріплюватися до клітин
хазяїна, нейтралізують токсичні властивості їхніх продуктів, утворюють імунні комплекси,
сприяючи швидшому виведенню антигенів з організму. Клітинна імунна відповідь грунтується на
функціонуванні Т-кілерів (цитотоксична реакція Т-лімфоцитів) та на діяльності активованих Тх1-
клітинами макрофагів (реакція гіперчутливості сповільненого типу). Т-кілери реалізують захисну
функцію, вбиваючи інфіковані патогенами або трансформовані клітини-мішені, а активовані Тх1
макрофаги – знищуючи внутрішньоклітинні патогени, що паразитують у них.
Синтезовані лімфоцитами антитіла і цитокіни підвищують ефективність механізмів природного
захисту. Антитіла активують комплемент, сприяючи лізису корпус кулярних антигенів (і комплексів
антиген – антитіло), опсонізують клітини-мішені фагоцитів і природних кілерів або «озброюють»
самі ефекторні клітини, зумовлюючи специфічність і прицільність їхньої дії, а продуковані Т-
лімфоцитами цитокіни підвищують активність фагоцитів та НК-клітин.
Розвиток імунної відповіді регулюється різними механізмами, які спрямовані переважно на її
поступове обмеження паралельно з виведенням антигену. У результаті імунної відповіді
утворюються Т- і В-клітини пам'яті які заміщують виснажені клони лімфоцитів та забезпечують
розвиток прискореної ефективної реакції на повторне проникнення в організм того самого антигену.
Контрольні запитання.
1. Які істотні відмінності реакції системи неспецифічного захисту від специфічних імунних реакцій?
2. Які головні шляхи появи антигенів в організмі та способи транспортування їх у лімфоїдні органи?
3. Яку функцію виконують М-клітини в організмі?
4. Чому індукування специфічних імунних реакцій може відбуватися лише в спеціалізованій
лімфоїдній тканині та лімфоїдних органах? У чому полягає біологічний сенс уловлювання клонів Т-
клітин у лімфатичному вузлі?
5. Яку функцію виконують дендритні клітини та як відбувається ініціювання специфічних імунних
реакцій?
6. Які процеси відбуваються з активованими В-клітинами в зародкових центрах?
7. Які головні ефекторні реакції клітинного імунітету?
8. Які ефекторні реакції зумовлюються антитілами?
9. Які механізми задіяні в регуляції імунної відповіді?
10. Чим зумовлена більша ефективність вторинної імунної відповіді порівняно з первинною і в чому
вона виявляється?
РОЗДІЛ 12. РОЗВИТОК ЛІМФОЦИТІВ.
Ефективне функціонування імунної системи потребує постійного поповнення пулу периферичних
лімфоцитів для заміщення клітин, що загинули під час виконання своїх функцій в імунних реакціях,
а також старих клітин, які так і не зустрілися зі своїм антигеном. Ці досить значні втрати клітин
компенсуються завдяки постійному, незалежному від антигенних стимулів утворенню лімфоцитів у
первинних лімфоїдних органах — тимусі (Т-лімфоцитів) і кістковому мозку (В-лімфоцитів).
Завдяки процесам лімфопоезу, що здійснюються в цих органах, підтримується не лише загальна
кількість Т- і В-лімфоцитів, а й різноманіття їх антигенспецифічних рецепторів. Клони лімфоцитів,
що формують цей репертуар, здатні розвивати реакції практично на всі можливі антигени, з якими
може зустрітися організм упродовж життя, виявляючи одночасно толерантність до аутоантигенів.
Це досягається у процесі відбору, якого зазнають Т- і В-лімфоцити в центральних лімфоїдних
органах і під час якого відбираються для подальшого дозрівання клітини, здатні розвивати реакції
на чужорідні антигени, та елімінуються аутоагресивні клітини — потенційно реактивні до власних
структур.
Джерелом лімфоцитів, як і всіх інших клітин імунної системи, є недиференційовані плюрипотентні
гемопоетичні стовбурові клітини, що локалізуються у кістковому мозку. Стовбурові кровотворні
клітини (СКК) є своєрідним «посівним матеріалом» для центральних органів імунної системи.
Здатність СКК до самовідновлення своєї популяції (self-renewal) і диференціювання в усі типи
клітин крові є практично невичерпною, що дає змогу компенсувати постійні величезні втрати їх
нащадків на периферії. Спочатку коротко зупинимося на характеристиці СКК, а потім розглянемо
процес диференціювання їх на лімфоцити.
12.1. СТОВБУРОВІ КРОВОТВОРНІ КЛІТИНИ.
У всіх ссавців у період ембріонального розвитку СКК спочатку утворюються в стінці жовткового
мішка, потім мігрують у печінку, а згодом заселяють кістковий мозок. У плоді людини в
жовтковому мішку СКК формуються на 3-му тижні, де знаходяться до 9-го тижня (так звана перша
генерація СКК), на 5 – 6-му тижні заселяють печінку, де знаходяться до кінця ембріонального
розвитку (друга генерація СКК), а в кістковий мозок потрапляють на 10—11-му тижні (третя
генерація СКК).
У кістковому мозку СКК розміщуються біля пристінної частини кістки в тісному контакті зі
спеціалізованими стромальними клітинами, які створюють необхідне
для їх розвитку
мікрооточення. Крім самих СКК тут знаходяться також їхні похідні, так звані напівстовбурові
клітини з більш обмеженими можливостями — комітовані попередники клітин мієлоїдного
(еритроцитів, гранулоцитів, моноцитів, мегакаріоцитів) та лімфоїдного (Т-, В-лімфоцитів, NK-
клітин) рядів.
Під впливом клітинного мікрооточення та локально продукованих цитокінів напівстовбурові та
стовбурові клітини залучаються до проліферації та диференціювання. Процеси проліферації та
диференціювання суворо регулюються, що забезпечує можливість самопідтримання відносно
невеликої популяції СКК. Завдяки цій унікальній особливості — здатності до самопідтримання —
СКК забезпечують постійне поповнення втрат у кровотворній і лімфоїдній тканині зрілих клітин з
обмеженим життєвим циклом. Слід зазначити, що властивості СКК були вивчені задовго до
ідентифікації їх морфології та фенотипу (див. розд. 1). Це стало можливим завдяки розробці
відповідних методичних підходів до їх дослідження.
Методи аналізу СКК. Вивчення функціональних характеристик СКК розпочалося в 1961 p., коли
канадськими вченими Тілом і Мак-Куллахом був запропонований метод клонування цих клітин у
селезінці смертельно опромінених мишей. Після введення тваринам клітин кісткового мозку на
стромі (мал. 87) спустошеної опроміненням селезінки з'являються у вигляді вузлів колонії клітин
(1—3 колонії на 10
4
введених клітин), які можна спостерігати візуально. Колонії на селезінці
опромінених тварин можна отримати також, якщо екранувати кістковий мозок під час опромінення.
Колонії, що утворюються у разі екранування кісткового мозку тварин, називають ендогенними, а ті,
що утворюються після трансплантації кісткового мозку, — екзогенними.
Селезінкові колонії мають клональне походження, що було доведено за допомогою індукованих
опроміненням хромосомних маркерів. Клітини, що отримали високу дозу (6,5 Гр) і вижили, як
правило, несуть індуковані радіацією аберації — хромосомні маркери. За цими маркерами можна
ідентифікувати вихідні клітини та їх нащадки під час каріологічного аналізу. У всіх клітин однієї
колонії виявляється (якщо виявляється) однаковий хромосомний маркер. Це свідчить про те, що
кожна окрема колонія (складається приблизно з 10
6
незрілих клітин) утворилася внаслідок
розмноження однієї вихідної клітини, тобто є клоном її нащадків. Ці вихідні клітини, що здатні
утворювати колонії в селезінці, називають селезінковими колонієутворювальними одиницями
(КУОс).
З погляду гістології колонії на селезінці неоднорідні й представлені трьома типами:
еритроцитарними, гранулоцитарно-моноцитарними і мегакаріоцитарними. Після ретрансплантації
клітин колонії будь-якого типу новому опроміненому реципієнту знову утворюються колонії всіх
трьох типів з одним і тим самим хромосомним маркером (і завжди з однаковим співвідношенням:
42% еритроцитарних, по 21% гранулоцитарно-моноцитарних та мегакаріоцитарних і 16%
змішаних). Отже, клітини, що утворюють колонії, здатні до самопідтримання (в колонії з однієї
КУОс утворюються інші КУОс) та диференціювання на більш зрілі клітинні форми. При цьому
підтримується лінія КУОс, що підтверджує існування спільного попередника клітин мієлоїдного
ряду.
Існування в кістковому мозку СКК, здатних диференціюватися у лімфоїдному напрямі, було
показано в дослідах на радіохимерах. Радіохимери — тварини, в організмі яких функціонують
клітини різних генотипів, їх отримують унаслідок трансплантації алогенного кісткового мозку
реципієнтам з пригніченою опроміненням імунною системою. Після трансплантації опроміненим
мишам лінії СВА клітин кісткового мозку з каріологічною міткою (СВА/Т6Т6) разом з лімфоцитами
лімфатичних вузлів, що не мали мітки, нащадки останніх першими заселяють лімфоїдні органи, але
поступово витісняються нащадками клітин донора кісткового мозку.

52
В останні роки існування в кістковому
мозку КУО — попередників лімфоцитів
(як і КУО — попередників клітин різних
типів мієлоїдного ряду) та їх здатність до
самопідтримання підтверджені in vitro
під час культивування клітин кісткового
мозку в спеціальних напіврідких
середовищах (див. розд. 1). Для цього
клітини кісткового мозку тривалий час
(5—10 тижнів) культивують за наявності
стромальних елементів кісткового мозку.
Вважають, що довго жити у такій
культурі можуть лише клітини з високим
проліферативним потенціалом. Потім
такі клітини висівають у напівтверді
середовища з високим вмістом цитокінів,
необхідних для диференціювання на
певні лінії клітин. На таких середовищах
СКК можуть утворювати колонії як
мієлоїдних, так і лімфоїдних клітин
залежно від наявності відповідних
цитокінів.
Для того щоб ідентифікувати фенотип
СКК, запропонували метод вибіркового
вилучення з кісткового мозку
гемопоетичних клітин, що активно
проліферують. Для цього до клітин
кісткового мозку додають
флуоресцентний аналог нуклеотидів — 5-
флуоурацил, який накопичується в
клітинах, що знаходяться у клітинному
циклі. Далі на клітинному сортувальнику
(цитофлуориметрі) можна виділити ці
клітини за флуоресценцією. Така
популяція клітин буде значно збагачена на СКК. Для їх ідентифікації використовують два критерії
— здатність до самовідновлення in vitro і відсутність поверхневих маркерів, характерних для всіх
відомих гемопоетичних ліній (CD3
–
, CD4
–
, CD8
–
, CD19
–
, CD20
–
, CD56
–
, CD11b
–
, CD14
–
, CD15
–
).
Такий фенотип скорочено позначають Lin
–
(lineage
–
). Для того щоб отримати Lin
–
клітини, потрібно
окремо вилучити всі інші субпопуляції клітин за допомогою антитіл до їх поверхневих маркерів.
Нині СКК мишей ідентифікують за експресією рецептора c-Kit до фактора стовбурових клітин та
вибірковою експресією одного з маркерів Thy-1.1 чи Flk2. Тобто СКК миші мають фенотип Lіn
–
c-
Kit
+
Thy-1.1
+
(Flk2
+
).
Для аналізу СКК людини використовують трансплантацію кісткового мозку людини мишам з
тяжкими вадами розвитку лейкоцитарних клітин, які отримують схрещуванням мишей ліній NOD і
SCID. Ця експериментальна модель не є повністю адекватною, тому більшість дослідів зі СКК
людини проводять у культурі in vitro.
Показано, що майже всі СКК людини експресують сіаломуцин CD34, який виявляється також на
клітинах ендотелію, де він є лігандом для L-селектину. Вважають, що CD34 може зумовлювати
взаємодію СКК зі стромальними елементами кісткового мозку. На СКК людини відсутня експресія
маркера CD38. Отже, найімовірніше, що СКК людини мають фенотип Lin
–
CD34
+
CD38
–
.
Міграція СКК. Важливою особливістю СКК є їх здатність виходити з кісткового мозку в кров,
циркулювати в кров'яному руслі та колонізувати інші відділи кровотворної й лімфоїдної систем
(насамперед тимус) і навіть нелімфоїдні периферичні тканини.
Міграція СКК починається ще під час ембріогенезу. Як уже зазначалося, СКК з'являються спочатку
в жовтковому мішку приблизно через 7,5 доби розвитку ембріона миші. Ці клітини дають початок
першим еритроїдним клітинам та, ймовірно, клітинам судин — ендотеліоцитам. Тому їх ще
називають гемангіобластами. Вони не мають здатності захищати смертельно опромінених дорослих
тварин, очевидно, через відсутність у них хемокінових рецепторів і здатності заселяти кістковий
мозок. Через 9 — 10 діб розвитку в ембріональній печінці з'являються СКК, які, ймовірно,
потрапляють сюди з жовткового мішка. Ці клітини вже набувають здатності захищати опромінених
реципієнтів та мігрувати в кістковий мозок. Аналогічні властивості набувають через 10 діб розвитку
СКК жовткового мішка. На 12-ту добу печінка набуває максимальної гемопоетичної активності.
СКК печінки Мас-1
+
відрізняються від СКК кісткового мозку за профілем клітин, на які вони
можуть диференціюватися. Так, тільки в ембріональній печінці утворюються В-клітини
субпопуляції В-1 (CD5
+
) та Vγ3 і Vγ4 Т-клітини. Крім того, з печінки виходять CD3
–
CD4
+
, що
експресують інтегрин а4β7. Вважають, що такі клітини є напівстовбуровими клітинами, які дають
початок антигенпрезентувальним ДК (таким як клітини Лангерганса), НК-клітинам і, можливо,
ФДК. На 16— 17-ту добу розвитку СКК з печінки починають заселяти кістковий мозок. Отже,
завдяки міграції ембріональних СКК забезпечується поетапне заселення різних гемопоетичних
органів.
Певна частина СКК постійно циркулює з кров'ю і в дорослих мишей. Цей показник у мишей
становить 0,3 % загальної кількості (3,3 • 10
3
) СКК в організмі. Концентрація СКК в крові ембріонів
перевищує в кілька разів їхній уміст у дорослих, а максимальний рівень реєструється у пізній
пренатальний і ранній постнатальний період. У процесі старіння мишей інтенсивність міграції СКК
знижується.
У крові людини також спостерігається висока концентрація СКК в пренатальний період, тому
клітини пуповинної крові можна використовувати як гемопоетичні клітини замість кісткового мозку
при трансплантаціях. Очевидно, в перспективі будуть проводити кріоконсервацію пуповинної крові
кожного народженого малюка, що дасть можливість (у разі потреби) проводити аутотрансплантацію
власної гемопоетично активної тканини.
Ендотеліальні клітини кісткового мозку завжди експресують молекули адгезії VCAM-1, інтегрин
VLA-4, L-селектин тощо, що зумовлює хомінг («повернення додому») СКК, що мігрують. Крім
того, СКК експресують хемокіновий рецептор CXCR4 і селективно відповідають на ліганд SDF-1α
що експресується ендотеліоцитами кістко вого мозку.
Переконливі докази здатності СКК ді міграції були отримані (Р. В. Петров P. M. Хаїтов) в
експериментах на з'єднаних у парабіоз (целіарний анастомоз) сингенних мишах (лінії СВА/Н), що
різнилися за хромосомним маркером Т6Т6 Одного із партнерів по парабіозу до операції
опромінювали тотально, а іншому (з маркером) екранували задні кінцівки. У селезінці тотально
опроміненого парабіонта всі ендогенні КУОс несли хромосомний маркер, що свідчить про міграції
їх з екранованого кісткового мозку. Автори встановили, що міграція і рециркуляція СКК на рівні
організму контролюється ендогенними глюкокортикоїдами (гормонами надниркових залоз).
Вважають, що фізіологічні концентрації цих гормонів вірогідно спричинюють стримувальний вплив
на міграцію СКК, а підвищення їх рівня, наприклад унаслідок інтенсивної дії різних стресових
факторів, супроводжується зменшенням кількості СКК в циркуляції.
Міграція й рециркуляція СКК є важливим етапом імуногенезу. Імунізація антигенами і вакцинами
значно інтенсифікує ці процеси. Після введення поліцукридних антигенів кількість СКК, що
рециркулюють, збільшується в десятки разів. Процеси запалення і різні цитокіни та значні втрати
крові стимулюють міграцію та рециркуляцію СКК.
Фізіологічна роль міграції СКК, крім міграції в тимус, залишається невідомою. Одним із можливих
пояснень міграції є потреба в перерозподілі гемопоетичної тканини між комірками кісткового
мозку. Завдяки міграції СКК можуть потрапляти в різні комірки, що підтримує баланс кровотворної
тканини в різних ділянках кісткового мозку. Припускають, що СКК можуть потрапляти в різні
органи нелімфоїдної природи і диференціюватися на клітини цих органів. Так, in vitro показана
можливість диференціювання СКК на нервові клітини, гепатоцити, серцеві й скелетні м'язи тощо.
Таке диференціювання СКК називають трансдиференціюванням. Чи має місце фізіологічне
трансдиференціювання in vivo, поки що залишається предметом дискусій, але, як припускають, таке
диференціювання може відбуватися у разі травм чи ураження периферичних тканин. Можливо, саме
з цим пов'язаний підвищений рівень рециркуляції СКК під час запалення. Відомо, що під час
запалення збільшується вихід з кісткового мозку в кров нейтрофілів, які частково розщеплюють
позаклітинний матрикс на судинах, залишаючи «дірки» для виходу інших клітин. Показано, що
інгібітори еластази нейтрофілів блокують підвищення рівня рециркуляції СКК під час запалення.
Нині інтенсивно вивчається можливість використання СКК для відновлення ушкоджених тканин за
різних патологічних станів, зокрема при інфаркті міокарда.
Отже, завдяки міграції й рециркуляції СКК та заселенню ними лімфоїдних органів, насамперед
тимуса, де здійснюється їх диференціювання, в організмі підтримується гомеостаз гемопоетичної
системи.
Проліферація і диференціювання СКК. Стовбурові клітини характеризуються двома важливими
властивостями — здатністю до самовідновлення своєї популяції внаслідок поділу і здатністю до
диференціювання на різні типи клітин крові. Здатність до самовідновлення зумовлює значну
тривалість життя популяції СКК та її здатність швидко компенсувати можливі втрати різних типів
клітин крові, зокрема лімфоїдні, впродовж усього життя організму. Можливість поповнення
постійних втрат клітин за незначної частки проліферувальних СКК зумовлена існуванням
напівстовбурових клітин (похідних від СКК). Останні, як уже зазначалося, інтенсивно
проліферують, але диференціювання їх на комітовані попередники відбувається повільно, через
багато мітотичних циклів.
У нормі переважна більшість СКК (90 %) перебувають у стадії спокою (G
Q
) або в тривалому періоді
G1. Частка проліферувальних клітин є незначною і становить не більше 10 % загальної кількості (і
лише у деяких ліній мишей близько 20%). В умовах різких ушкоджувальних впливів на кістковий
мозок (наприклад, під час опромінення) проліферація може індукуватися в усій популяції СКК, що
збереглася. Однак навіть за цих умов здатність СКК до самопідтримання повністю не вичерпується
завдяки чіткій регуляції процесів проліферації і диференціювання. Саме регуляція цих процесів
забезпечує самопідтримання популяції СКК і захист їх від виснаження у результаті
диференціювання в умовах величезного попиту на клітини різних типів на периферії.
Регуляція проліферації та диференціювання СКК здійснюється мікрооточенням і забезпечується
тими сигналами, що індукуються контактами СКК зі стромальними клітинами кісткового мозку, та
локальним продукуванням цитокінів. Перші дослідження, спрямовані на з'ясування ролі
мікрооточення в регуляції проферації і диференціювання СКК, були проведені наприкінці 60-х —
на початку 70-х років XX ст. (N. S. Wolf, I. J. Frentin,1968; А. Я. Фриденштейн та ін., 1973; І. Л.
Чертков, 1974).
Характерною особливістю СКК є те, що під час поділу вони утворюють нащадків, які не обов'язково
стають на шлях диференціювання. Саме тому вони можуть зберігати свій проліферативний
потенціал. В умовах стабільного кровотворення відбувається диференційний поділ СКК, коли після
поділу материнської клітини одна дочірня залишається стовбуровою, заміщуючи материнську, а
інша диференціюється в потрібному напрямі (так званий асиметричний поділ). У разі
гемопоетичного стресу відбувається так звана розігнана проліферація, коли після поділу
материнської клітини обидві дочірні залишаються стовбуровими (не залучаються до
диференціювання). Для підтримання проліферації СКК і самооновлення їх популяції велике
значення має продукування ними спеціального фактора росту (фактор росту СКК) та експресія
рецептора до нього (c-Kit).
Для задоволення потреб у кровотворенні СКК використовуються з урахуванням їхнього віку. До
проліферації залучаються насамперед старіші клітини, які зробили більшу кількість поділів, отже,
зберігається молодша їх популяція з вищою здатністю до самопідтримання.
Одночасно з проліферацією здійснюється диференціювання СКК — багатостадійний процес, у ході
якого клітина втрачає здатність до самопідтримання і вибирає один із можливих шляхів розвитку —
комітується. Перетворення СКК на спеціалізованого попередника завершується лише через кілька
десятків поділів. У диференціюванні (як і в проліферації) СКК важлива роль належить їх
клітинному мікрооточенню і факторам, що діють локально, у тих органах, де здійснюється цей
процес.
Молекулярні механізми, що визначають комітування СКК у тому чи іншому напрямі розвитку,
почали інтенсивно вивчати лише в останні роки. Значний прогрес У вивченні цього питання був
досягнутий завдяки розвитку методів специфічного порушення генів у мишей, які дістали назву
«нокауту генів». Розвиток методів генетичного нокауту вимагає спеціальної системи позначення
генотипів змінених тварин. Наявність певного гена в інтактному стані позначають символом «+», а
відсутність — символом «-» після назви відповідного гена. Наприклад, генотип гомозиготних
мишей, дефектних за фактором транскрипції Рах5, позначають Рах5
–
/
–
. Гетерозиготи, у яких
відсутній лише один ген Рах5 з алельної пари, позначають відповідно Рах5
+/–
.
Іншим важливим методом, який дав можливість аналізувати програму розвитку клітин, є метод
аналізу генної експресії на рівні однієї клітини. Завдяки застосуванню цих методів нині з'ясовано
ряд транскрипційних факторів, які є необхідними для диференціювання різних типів гемопоетичних
клітин, хоча причини активації цих факторів у багатьох випадках ще залишаються невідомими (див.
нижче). Усі етапи диференціювання більшості типів клітин мієлоїдного ряду, за винятком
моноцитів, здійснюються безпосередньо в кістковому мозку і завершуються утворенням
функціонально повноцінних клітин, що виходять на периферію, де й виконують притаманні їм
функції (див. розд. 1,2).
Диференціювання спільних попередників лімфодних клітин у напрямі Т- та В-лімфоцитів потребує
різних умов мікрооточення і відбувається в різних лімфоїдних органах. Крім того, навіть
проходження послідовних стадій диференціювання клітин однієї лімфоїдної лінії всередині одного
органа залежить від факторів мікрооточення в різних його компартментах. Диференціювання В-
лімфоцитів відбувається безпосередньо в кістковому мозку. Диференціювання Т-лімфоцитів
здійснюється за умови спрямованої міграції їх попередників з кісткового мозку до тимуса, в різних
компартментах якого і відбуваються різні стадії диференціювання.
Особливістю лімфоїдних клітин-попередників, яка відрізняє їх від клітин-попередників інших ліній
кровотворення, є збереження в них експресії унікального ферменту СКК — теломерази, що
«репарує» (відновлює довжину теломер) хромосоми. Це свідчить про закладені в них великі
проліферативні можливості, які будуть реалізовуватися цими клітинами не лише у лімфопоезі, а й у
процесі розвитку лімфоцитами імунної відповіді.
У життєвому циклі лімфоцитів можна виділити дві фази: дозрівання в центральних лімфоїдних
органах за відсутності антигенного стимулу та постактиваційне диференціювання на периферії
після контакту з чужорідним антигеном. Процес дозрівання супроводжується позитивним і
негативним відбором репертуару рецепторів, зумовленим взаємодією незрілих лімфоцитів з
власними антигенами, і завершується утворенням зрілих наївних лімфоцитів, які несуть рецептори,
реактивні щодо чужорідних і толерантні щодо власних антигенів. Диференціювання, індуковане
активацією наївних лімфоцитів чужорідним антигеном, приводить до збільшення кількості
специфічних до цього антигену лімфоцитів і утворення клонів ефекторних клітин та клітин пам'яті.
Постактиваційне диференціювання Т-і В-клітин було розглянуто у розд. 10 і 11. Тому в цьому
розділі детально розглянемо лише антигеннезалежне диференціювання Т- і В-лімфоцитів зі
стовбурових гемопоетичних клітин і процеси відбору рецепторів певної специфічності, які
відбуваються ще до контактування лімфоцитів з чужорідними антигенами.
12.2. РОЗВИТОК Т-ЛІМФОЦИТІВ У ТИМУСІ.
Розвиток Т-лімфоцитів відбувається в тимусі, куди мігрують їхні кістково-мозкові попередники, і
тому їх назвали тимусзалежними лімфоцитами, або Т- клітинами.
Роль тимуса в розвитку Т-клітин було доведено в експериментах, в яких цей орган вилучали у
нормальних мишей або пересаджували його безтимусним мишам.
12.2.1. Роль тимуса в розвитку Т-клітин.
Т-Клітини, як уже зазначалося, виконують ефекторні функції в реакціях клітинного імунітету і
відіграють важливу роль у гуморальних імунних реакціях на тимусзалежні антигени, активуючи В-
лімфоцити до синтезу антитіл. Саме тому тимусу належить провідна роль у функціонуванні системи
імунітету, в забезпеченні імунної реактивності організму. Однак важливість тимуса для
функціонування імунної системи довго залишалася не з'ясованою і вважалася сумнівною, оскільки
видалення органа у дорослих тварин не спричинює істотного впливу на імунореактивність. Як
виявилося, це пов'язано з існуванням Т-клітин пам'яті і здатних до самопідтримання популяцій Т-
клітин, локалізованих у слизових оболонках та шкірі.
Виняткова важливість тимуса для імунітету і розвитку Т-лімфоцитів була доведена лише на початку
60-х років XX ст. (Міллер та ін., 1961, 1962) в експериментах, в яких, на відміну від усіх попередніх
спроб, тимус був видалений відразу після народження мишей. Така рання (неонатальна) тимектомія
призводить до розвитку в мишей імунодефіцитного стану, що виявляється втратою здатності
розвивати клітиноопосередковані реакції, синтезувати антитіла на тимусзалежні антигени та різким
зниженням кількості лімфоцитів, особливо в тимусзалежних зонах периферичних лімфоїдних
органів і тканин. Пізніше роль тимуса в розвитку Т-лімфоцитів була підтверджена на безтимусних
лініях мишей nude, в яких кількість Т-лімфоцитів становить лише близько 3 % їх рівня у
нормальних мишей. Клінічні спостереження також свідчать, що у дітей з природженими дефектами
(гіпоплазією чи аплазією) тимуса, як, наприклад, при синдромі Ді Джорджі, формується Т-
клітинний імунодефіцит з порушенням дозрівання Т-клітин та зниженням імунореактивності. Отже,
рання тимектомія і природжена відсутність тимуса призводять до розвитку глибокого Т-
імунодефіциту.
Усі явища імунної недостатності повністю ліквідуються після пересаджування неонатально
тимектомованим і безтимусним мишам сингенного тимуса новонароджених тварин. Відновлення
імунної реактивності після пересаджування тимуса спостерігається також у Т-імунодефіцитних
дітей, однак ефект його обмежується терміном функціонування алогенного трансплантата. Після
пересаджування тимуса відбувається міграція Т-лімфоцитів у периферичні лімфоїдні органи та
заселення ними Т-зон з одночасним відновленням імунних функцій. Часткове відновлення імунної
реактивності відбувається і тоді, коли тимус пересаджують у мікродифузійних мембранах, крізь
пори яких клітини не проходять. Це свідчить про те, що тимус здійснює імунорегуляторні функції
за допомогою продукованих гуморальних факторів. Однак спроби замістити функції тимуса за
допомогою екстрагованих із нього гормональних факторів (тимозини, тимопоетини, тимулін) не
дали очікуваних результатів. Це пояснюється тим, що розвиток Т-лімфоцитів залежить не стільки
від гуморальних факторів, продукованих клітинами тимічного мікрооточення, як передусім від
прямих взаємодій з цими клітинами. Імуностимулювальний ефект тимусних препаратів, які
використовуються для терапії вторинних імунодефіцитів, зумовлений переважно їхнім впливом на
функціональне дозрівання клітин, що виселилися з тимуса.
Слід зазначити, що імуновідновлювальний ефект пересаджування тимуса у разі дефектів розвитку
цього органа можливий лише за умови функціональної повноцінності СКК, які дають початок
попередникам Т-лімфоцитів. За тих форм імунодефіцитів, що характеризуються дефектами тимуса
та ураженням СКК, пересаджування лише тимуса не дає відновлювального ефекту. Отже, тимус
забезпечує здійснення генетично детермінованої програми розвитку Т-лімфоцитів із СКК.
Функціонально повноцінні СКК є і в безтимусних мишей.
Безпосередня роль тимуса в диференціюванні СКК на Т-лімфоцити чітко доведена в експериментах,
в яких неонатальна тимектомія у мишей поєднується зі смертельним опроміненням і наступною
трасплантацією кісткового мозку. У таких тварин Т-лімфоцити не утворюються, а імунні функції не
відновлюються. І навпаки, в опромінених мишей, яким тимектомія не проводилася, ці процеси після
трансплантації кісткового мозку відбуваються нормально.
Важливу роль тимічної строми в диференціюванні Т-попередників демонструють результати
експериментів, в яких використовували дві лінії мутантних мишей з різко зниженою кількістю Т-
клітин, зумовленою різними причинами. У мишей nude Т-імунодефіцит пов'язаний з дефектом
розвитку тимічного епітелію, а в мишей scid (severe combined immune deficiency) — з дефектом
реорганізації генів ТкР. Після трансплантації кісткового мозку nude мишам scid розвиток Т-
попередників в організмі реципієнтів відбувається нормально. Власні кістковомозкові Т-
попередники також розвиваються у мишей nude нормально за умови трансплантації їм тимуса
мишей scid.
Тимус забезпечує специфічне мікрооточення, здатне ефективно підтримувати розвиток Т-
лімфоцитів з клітин-попередників, що походять із кісткового мозку.
Тимічне мікрооточення. У створенні тимічного мікрооточення беруть участь спеціалізовані
клітини строми органа в тісній взаємодії з локалізованими в ньому лімфоїдними клітинами.
Стромальні клітини тимуса представлені епітеліоцитами та клітинами мезенхіми.
Головним компонентом тимічного мікрооточення є епітеліальні клітини. Разом з іншими
стромальними клітинами вони формують густу тривимірну сітку, в якій містяться клітини
гемопоетичного походження, що колонізують тимус, — тимоцити, макрофаги та дендритні клітини.
Онтогенетичне походження клітин тимічного епітелію нині ще дискусійне. Невідомо, чи всі вони
походять із однієї стовбурової клітини, яка дає початок кортикальним і медулярним епітеліоцитам,
чи генерація цих типів епітеліальних клітин забезпечується двома пулами стовбурових клітин —
екто- та ендодермального походження. Виявлено субпопуляції тимічних епітеліоцитів, що
одночасно експресують маркери TR4 та TR5 або кератини К8 та К5, які зазвичай експресуються
відповідно виключно на кортикальному та медулярному епітелії. Припускають, що ці клітини, які
коекспресують специфічні для кортикального і медулярного епітелію маркери, можуть бути
проміжною стадією в розвитку епітеліальних клітин тимуса і дають початок кортикальним та
медулярним епітеліоцитам.
Важливу роль у регуляції розвитку тимічного епітелію та його функції відіграють фактори
транскрипції, серед яких найвідоміший Whn, що продукується локусом nude і є важливим для
розвитку тимуса в гризунів. У дорослих мишей nude, у яких Whn немає, є лише рудимент тимуса,
тоді як зародковий тимус у цих мишей (як і у Whn-нокаутних мишей) має відносно нормальну
морфологію до колонізації його лімфоцитами. Припускають, що Whn впливає на розвиток найбільш
ранніх тимічних епітеліальних клітин, а тому за його відсутності утворення нормальної архітектури
тимуса не відбувається. Активну участь у диференціюванні та функції тимічних епітеліоцитів
беруть й інші експресовані ними фактори транскрипції, дефіцит яких призводить до значних
порушень: атимії (Ноха3 та Рах9), зменшення кількості тимоцитів (Рах1).
Епітеліальні клітини, що формують тимічне мікрооточення, морфологічно і функціонально досить
гетерогенні й представлені кількома типами. На розвиток тимоцитів епітеліальні клітини можуть
впливати встановленням прямих міжклітинних контактів та за допомогою секретованих
гуморальних факторів.
Секреторні епітеліальні клітини локалізуються в субкапсулярній та медулярній зонах, продукують
цитокіни (ІЛ-1, -3, -6, -7, КСФ) і гуморальні фактори з властивостями гормонів (тимозини,
тимопоетини, тимулін). Гормони відіграють допоміжну роль у розвитку тимоцитів у тимусі, однак
дуже важливу — в дозріванні Т-клітин після міграції на периферію.
Іншим типом тимусних епітелоцитів є клітини-няньки, або клітини-годувальниці, — клітини
великих розмірів з відростками, що локалізуються у зовнішніх шарах кортикальної зони і в своїх
цитоплазматичних заглибинах («кишенях») містять великі агрегати (до кількох десятків) тимоцитів.
Вважають, що ці клітини спричинюють вплив на дозрівання та диференціювання на субпопуляції
незрілих тимоцитів і здійснюють елімінацію загиблих унаслідок апоптозу клітин, а можливо, й самі
індукують цей процес.
Епітеліальні клітини, що утворюють складну тривимірну сітку в кортексі, впливають на розвиток
тимоцитів за допомогою індукування сигналів унаслідок прямих міжклітинних контактів. Взаємодія
тимоцитів з епітеліоцитами, які експресують молекули МНС, є основою позитивного відбору.
Важливими компонентами тимічного мікрооточення є макрофаги та дендритні клітини. Макрофаги
локалізуються в обох зонах тимуса і приблизно половина з них експресують молекули МНС II, а ДК
– переважно зосереджуються в кортикомедулярній ділянці, менше — в медулярній зоні та всі без
винятку несуть МНС II. Серед ДК є клітини мієлоїдного і лімфоїдного походження. ДК і макрофаги
утворюються безпосередньо в тимусі з кістковомозкових попередників.
Експериментальні дані свідчать про вплив медулярного епітелію на розвиток у тимусі певних
субпопуляцій ДК. У мишей, які не експресують транскрипційний фактор RelB, у тимусі відсутні
деякі субпопуляції медулярних епітеліоцитів і немає лімфоїдних СD8
+
ДК, хоча наявні мієлоїдні
СD8
–
ДК. Разом з тим після введення Rel В
–
/
–
кісткового мозку опроміненим особинам дикого типу в
них утворюються ДК обох субпопуляцій. Локалізовані в тимусі ДК та макрофаги (МНС ІІ
+
)
відіграють роль факторів, що підтримують розвиток тимоцитів і зумовлюють елімінацію потенційно
аутореактивних клітин.
Крім тимічного епітелію на розвиток тимоцитів можуть впливати тимусні клітини мезенхімного
походження, що представлені в кортексі. їхній вплив реалізується на стадії незрілих подвійно-
53
негативних тимоцитів (CD4
–
CD8
–
CD44
+
CD25
+
) і, як припускають, здійснюється опосередковано
— сорбцією (і презентацією) факторів росту та цитокінів на міжклітинному матриксі.
Мезенхімальні клітини можуть впливати також на формування раннього тимічного епітелію,
вірогідно стимулюючи ріст і диференціювання епітеліоцитів безпосередньо — за допомогою
продукованих факторів росту фібробластів.
Слід зазначити, що самі тимоцити можуть безпосередньо впливати на формування необхідного для
їх розвитку мікрооточення. Тимоцити спричинюють регуляторний вплив на розвиток як
кортикального, так і медулярного епітелію. Так, у трансгенних мишей з мутаціями гена ε-ланцюга
молекули CD3 пригнічення розвитку тимоцитів на ранній стадії поєднується з глибокими
аномаліями розвитку тимічного кортексу. Розвиток медулярного епітелію може регулюватися
локалізованими в медулі зрілими тимоцитами. Підтвердженням цього є дані про виникнення
дефектів в організації медулярного епітелію під час блокування розвитку тимоцитів та відновлення
структури медулярної речовини після введення Т-клітин, наприклад у мишей scid з тяжким
комплексним імунодефіцитом.
Отже, різні взаємодії і взаємовплив епітеліального та лімфоїдного компартментів тимуса
забезпечують утворення в різних зонах органа специфічних клітинних мікрооточень, які впливають
на різні стадії розвитку Т-лімфоцитів зі спеціалізованих клітин-попередників.
12.2.2. Заселення тимуса Т-попередниками та їх дозрівання.
Заселення зачатка тимуса попередниками Т-лімфоцитів з ембріональної печінки у птахів і ссавців
відбувається у вигляді кількох міграційних актів. У розвинений тимус Т-попередники проникають у
постнатальний період з кісткового мозку постійно. Так, в ембріонів мишей заселення відбувається у
вигляді двох хвиль — через 11 – 13 та 17 – 19 діб ембріогенезу, а в дорослих мишей — починаючи з
7-ї доби після народження.
В ембріонів попередники Т-клітин потрапляють до тимуса через мезенхімну капсулу, оскільки
перша колонізація органа передує васкуляризації. В дорослих організмів клітини-попередники
проникають усередину тимуса крізь стінки великих венул у кортикомедулярній ділянці, долаючи
гематотимічний бар'єр. В утворенні бар'єра беруть участь судинні ендотеліальні клітини, що
частково перекривають одна одну, розміщений за ними шар макрофагів і базальна мембрана
(зовнішня епітеліальна вистілка). Бар'єр забезпечує вибіркове проникнення в тимус СКК
(попередників Т-клітин), запобігаючи проникненню клітин інших типів. Подолання бар'єра Т-
попередниками зумовлено наявністю на їхній поверхні рецепторів хомінгу (молекул CD44,
інтегринів VLA-4 і 6, L-селектину), за допомогою яких вони розпізнають комплементарні структури
на судинному ендотелії та позаклітинному матриксі. Лігандами молекули CD44 є похідні
гіалуронової кислоти на ендотелії та компонент міжклітинного матриксу фібронектин, а лігандами
інтегринів VLA-4 і VLA-6 — білки матриксу фібронектин і ламінін. L-селектин розпізнає кінцеві
залишки D-β-N-ацетилглюкозаміну в складі глікопротеїнів ендотелію. Проходження крізь базальну
мембрану зумовлюється інвазивними властивостями Т-попередників — продукуванням ферментів
гіалуронідази та колагенази.
Т-попередники рекрутуються в тимус за допомогою певних хемотаксичних факторів, що
продукуються клітинами цього органа. Одним із можливих кандидатів, відповідальним за залучення
попередників Т-лімфоцитів з кісткового мозку до тимуса, вважається хемокін CCL25 (ТЕСК). Цей
хемокін експресується тимічними ДК та індукує міграцію in vitro незрілих та зрілих тимоцитів, що
несуть рецептор до нього (CCR9), але не є хемоатрактантом для зрілих периферичних Т-клітин.
Однак, як виявилося, МкАТ до цього хемокіну не перешкоджають колонізації тимуса. Це може
свідчити про те, що ТЕСК не відповідає за міграцію попередників Т-клітин до тимуса або
принаймні не є єдиним хемоатрактантом для них. Є дані, що функцію хемоатрактантів для Т-
попередників виконують такі хемокіни, як XCL1 (лімфотактин) та CCL2 (МСР-1).
Важливими процесами у розвитку Т-лімфоцитів є комітування попередників до диференціювання
на Т-клітини та вибір між αβ та γδ-клітинними лініями. Тривалий час було невідомо, чи комітування
клітин-попередників до розвитку на Т-клітини відбувається ще до проникнення їх у тимус, чи їхній
розвиток детермінується вже всередині органа кортикальним мікрооточенням. Нещодавно отримані
експериментальні дані свідчать на користь другого припущення.
Першими в ембріональному тимусі розвиваються γδТ-клітини з попередників першої хвилі міграції.
αβТ-клітини диференціюються з попередників другої хвилі міграції, що проникли в тимус із
ембріональної печінки. Попередники з кісткового мозку, що після народження заселяють тимус
мишей, переважно диференціюються на αβТ-клітини, які поповнюють периферичний пул Т-
лімфоцитів.
Стадії диференціювання Т-клітин. Після проникнення всередину тимуса клітини-попередники
мігрують до зовнішнього шару кори — в субкапсулярну ділянку. У міру дозрівання клітини
переміщуються у глибший внутрішній шар кори та в медулярну зону. Розвиток попередників на
зрілі Т-клітини відбувається внаслідок багатоетапного диференціювання. На всіх етапах
диференціювання в тимусі клітини називають тимоцитами, а процес розвитку Т-клітин у тимусі —
дозріванням. На сьогодні детально досліджено розвиток αβТ-клітин, хоча деякі важливі моменти,
зокрема механізми їх позитивного відбору, залишаються ще недостатньо з'ясованими.
Дозрівання Т-клітин супроводжується змінами їх фенотипу — появою або зникненням поверхневих
молекул. Певні специфічні комбінації цих молекул є маркерами Т-клітин на різних стадіях
дозрівання і дають змогу диференціювати різні субпопуляції тимоцитів. Серед численних молекул
клітинної поверхні найважливішими є ті, що відображають стан функціонального дозрівання Т-
клітин. Це насамперед Т-рецепторний комплекс (ТкР і CD3) та корецептори CD4 і CD8.
Кістковомозкові клітини-попередники, що вперше колонізують тимус, можуть давати початок як γδ
і αβТ-лімфоцитам, так і лімфоїдним ДК. Після диференціювання і проліферації впродовж тижня
вони вже несуть специфічні для Т-клітинної лінії молекули (наприклад, CD2 у людини, Thy1 у
мишей), але не експресують жодного з трьох функціонально значущих маркерів (ТкР-CD3, CD4,
CD8).
Важливими подіями у розвитку αβТ-клітин є реорганізація й експресія генів α- і β-ланцюгів ТкР та
відбір репертуару незрілих αβТкР. Нагадаємо, що перебудові генів ТкР передує активація генів
рекомбіназ RAG1 і RAG2 та гена ТdТ (термінальної дезоксинуклеотидилтрансферази), що каталізує
синтез олігонуклеотидних вставок у ДНК у місцях з'єднання V-, D-і J-генетичних сегментів.
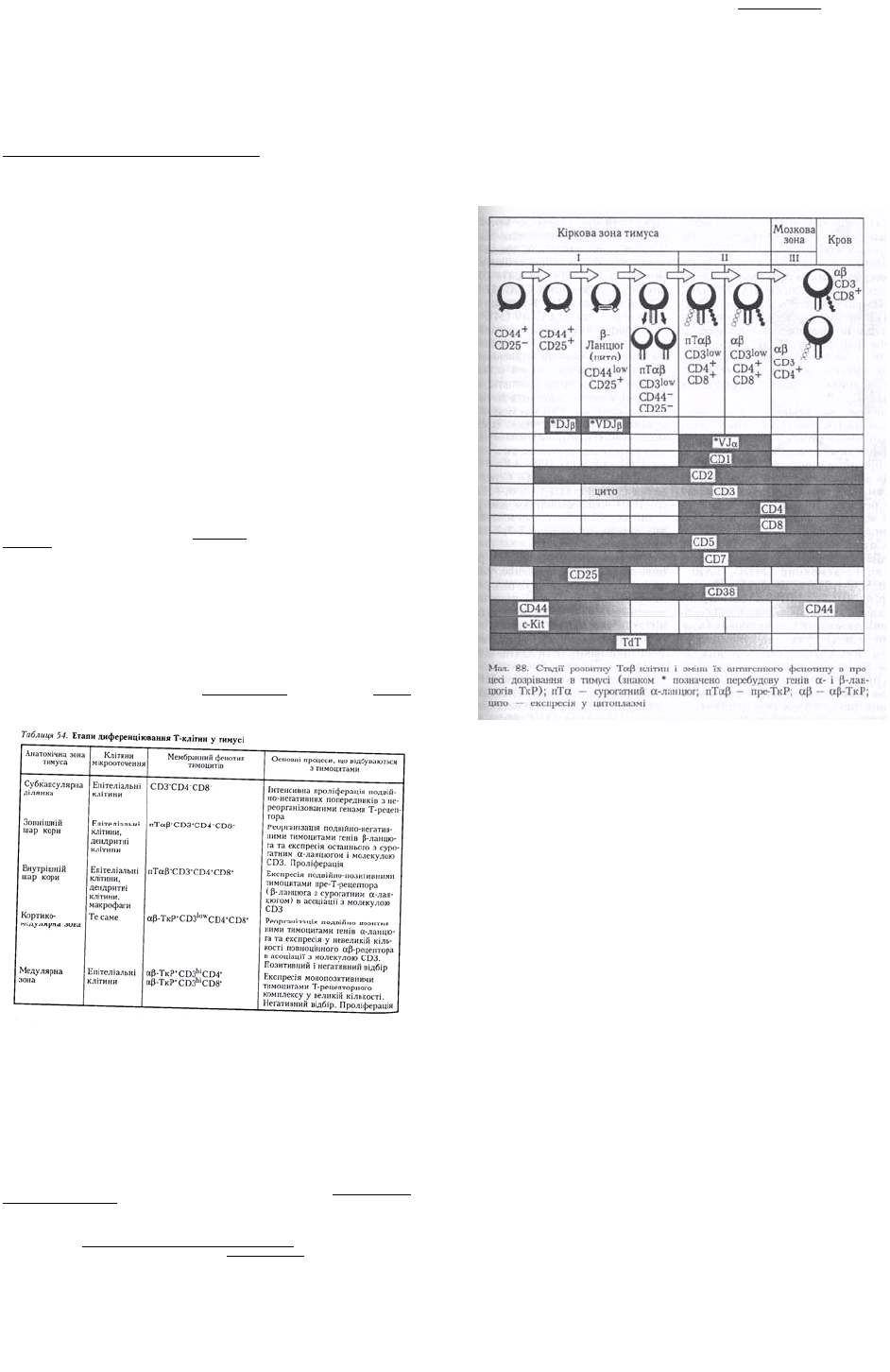
Процес диференціювання αβТ-клітин у тимусі можна поділити залежно від експресії ними
корецепторів CD4 і CD8 на три стадії: І — подвійно-негативних (CD4
–
CD8
–
), ІІ — подвійно-
позитивних (CD4
+
CD8
+
) та ІІІ — монопозитивних (CD4
+
i CD8
+
) тимоцитів. У табл. 54 показано
зміни експресії основних функціональних маркерів (αβарТкР, CD3, CD4, CD8) у процесі
диференціювання Т-клітин у різних анатомічних зонах тимуса і зазначено, на яких етапах
відбуваються процеси позитивного та негативного відбору (див. далі).
Подвійно-негативні тимоцити. Стадію подвійно-негативних тимоцитів зазвичай поділяють на
етапи залежно від експресії молекули міжклітинної адгезії CD44 та молекули CD25 — α-ланцюга
рецептора ІЛ-2. На ранньому етапі ці клітини експресують CD44, але не несуть CD25. Гени обох
ланцюгів ТкР у цих клітин, що мають фенотип CD44
+
CD25
–
, зберігають зародкову конфігурацію. На
наступному етапі дозрівання клітини починають експресувати CD25, набуваючи фенотип
CD44
+
CD25
+
, і розпочинають реорганізацію генів, що кодують β-ланцюг ТкР. Спочатку
відбувається D-J-реаранжування генів β-ланцюга, причому в обох хромосомах. Пізніше ці клітини
зменшують експресію CD44 за збереження рівня експресії CD25. На цьому етапі клітини фенотипу
CD44
low
CD25
+
затримуються до повного завершення реорганізації генів β-ланцюга (відбувається V-
DJ-рекомбінація, але тільки в одній хромосомі) та експресують цитоплазматичну форму CD3.
Експресований β-ланцюг з'єднується з сурогатним α-ланцюгом пТα — пре-Т-сеll α (gр33), внаслідок
чого формується пре-ТкР (пТαβ), який експресується в асоціації з CD3 на поверхні клітини у
невеликій кількості.
Клітина припиняє експресію CD25 (набуває фенотипу CD44
–
CD25
–
) та реорганізацію генів β-
ланцюга, виходить зі стану спокою в клітинний цикл і проліферує. Показано, що в індукуванні
утворення CD44
–
CD25
–
тимоцитів головну роль відіграє транскрипційний фактор GATA-3, а в
регуляції проліферації тимоцитів після успішного утворення пре-ТкР бере участь молекула Notch-З.
Внаслідок проліферації накопичується велика кількість клітин з певним β-ланцюгом, але з ще не
реорганізованими генами α-ланцюга. Пізні подвійно-негативні тимоцити, що активно
проліферують, мають морфологію бластних клітин і тому їх іноді називають великими подвійно-
негативними лімфоцитами. На цій стадії вони залишаються впродовж шести поділів. Під час
проліферації клітини розпочинають експресувати молекули CD4 та CD8 і переходять до наступної
стадії розвитку.
Подвійно-позитивні тимоцити. Після припинення проліферації клітини реорганізовують гени α-
ланцюга ТкР та експресують повноцінний αβ-Т-клітинний рецептор в асоціації з комплексом CD3 і
набувають статусу малих подвійно-позитивних (CD4
+
CD8
+
) тимоцитів. Щільність експресії ТкР на
їхній поверхні низька. На цій стадії відбувається позитивний відбір — скринінг (перевірка)
тимоцитів на здатність їхніх рецепторів розпізнавати власні молекули МНС. Якщо зв'язування ТкР з
комплексами власних МНС — пептид не відбудеться, клітини гинуть. Середня тривалість життя цих
СD4
+
СD8
+
-тимоцитів – 3-4 доби. Тимоцити, рецептори яких уже провзаємодіяли з комплексами
власні МНС — пептид, виживають і продовжують диференціювання. Пізні подвійно-позитивні
тимоцити, що пройшли позитивний відбір, починають експресувати велку кількість комплексів ТкР
— CD3. Вони зазнають подальшого позитивного відбору на здатність розпізнавати МНС І або МНС
II і визначають свою майбутню спеціалізацію.
Монопозитивні тимоцити. Згодом подвійно-позитивні тимоцити припиняють експресію одного з
корецепторів і стають монопозитивними CD4
+
або СD8
+
тимоцитами. СD4Т-клітини у
функціональному відношенні є попередниками хелперів, а CD8T-клітини — попередниками
цитотоксичних Т-лімфоцитів.
Під час розвитку тимоцити зазнають не лише позитивного, а й негативного відбору — скринінгу на
аутореактивність, який відбувається як під час монопозитивної, так і подвійно-позитивної стадій
розвитку. Аутореактивні клітини елімінуються.
Клітини, що мігрують у медулярну зону, затримуються тут на досить тривалий проміжок часу —
два тижні з трьох, упродовж яких триває у мишей процес дозрівання в тимусі Т-клітин з їхніх
попередників. У медулярній зоні, очевидно, триває диференціювання монопозитивних CD4 і CD8 Т-
клітин, яке супроводжується зміною профілю експресії багатьох поверхневих молекул (CD24, Qa2,
CD62L, CD69, 3G11). Однак не з'ясовано, чи диференціювання на стадії монопозитивних тимоцитів
здійснюється автономно, чи під впливом певних стромальних (але не епітеліальних) клітин. На
цьому етапі відбувається незалежна від взаємодії з МНС проліферація зрілих Т-клітин (щонайменше
шість мітотичних циклів), яка принаймні частково індукується ІЛ-7. Припускають, що друга хвиля
внутрішньотимічної проліферації зрілих Т-клітин перед їхньою міграцією з тимуса потрібна для
збільшення кількості відібраного peпертуару Т-клітин.
Отже, тимоцити на різних стадіях диференціювання різняться за експресією основних
функціональних маркерів. Тимоцити першої стадії (ранні) не несуть корецепторів CD4 і CD8,
реорганізовують гени β-ланцюга ТкР та експресують пре-ТкР (комплекс β-ланцюга з сурогатним α-
ланцюгом) в асоціації з молекулою CD3. Тимоцити другої стадії (проміжні) коекспресують
корецептори CD4 і CD8, реорганізовують гени α-ланцюга ТкР та експресують невелику кількість
комплексів αβ-ТкР з CD3. Тимоцити третьої стадії (зрілі) несуть лише один із корецепторів (CD4
або CD8) і велику кількість комплексів αβ-ТкР з CD3 (табл. 54).
Крім зазначених маркерів Т-клітини несуть інші поверхневі молекули, експресія яких може
змінюватися на різних стадіях диференціювання або залишатися постійною впродовж дозрівання в
тимусі чи навіть після міграції з нього. Експресію маркерів на різних стадіях дозрівання Т-клітин у
людини наведено на мал. 88.
Одним із перших Т-клітинних маркерів, що експресуються на клітинах-попередниках ще до
проникнення їх у тимус, є молекула CD7 (експресується на лімфоїдних стовбурових клітинах і на
зрілих Т-клітинах). Однак за певних умов вони можуть диференціюватися також на В-лімфоцити,
НК-клітини, ДК. Ця здатність зберігається у них ще впродовж короткого періоду після проникнення
в тимус. З появою на поверхні тимоцитів молекули CD25, яка збігається в часі з експресією маркера
Т-клітин — молекули CD2, диференціювальні потенції Т-попередників звужуються. На цьому етапі
вони зберігають здатність диференціюватися, крім Т-клітин, також на ДК, однак втрачають її
одночасно з втратою експресії молекули CD44. До речі, молекула CD44, що виконує роль рецептора
хомінгу під час проникнення клітин-попередників у тимус, зберігається лише на найбільш юних
тимоцитах і знову експресується у невисокому рівні на зрілих тимоцитах перед міграцією їх на
периферію. Вона експресується також Т-клітинами пам'яті та ефекторними Т-клітинами і відіграє
певну роль у їхній міграції.
Іншим маркером Т-попередників є фермент TdT, активність якого найбільша у СК4
–
СК8
–
-
тимоцитах, зменшується у СК4
+
СК8
+
-тимоцитах і зникає у зрілих монопозитивних Т-клітинах. TdT
відіграє важливу роль у створенні репертуару ТкР.
Нині залишається ще не з'ясованим функціональне значення експресії молекули СК25 (α-ланцюга
рецептора до ІЛ-2). Ця молекула експресується лише на ранніх СК4
–
СК8
–
-тимоцитах, що
характеризуються високою проліферативною активністю (зникає після перебудови β-ланцюга ТкР).
Однак після руйнування гена ІЛ-2 внаслідок нокауту розвиток Т-клітин у мишей відбувається
нормально. Можливо, ІЛ-2 поряд з іншими цитокінами (ІЛ-1,-6, ГМ-КСФ, ФНП-α) виконує роль
костимулятора проліферації тимоцитів (зокрема, на етапі відбору тимоцитів, що пройшли
реаранжування генів β-ланцюга). Певну роль у проліферативних процесах на ранній стадії,
очевидно, відіграє і фактор стовбурових клітин, рецептор для якого (СК117) експресується на СКК,
Т-попередниках і на ранніх CD4
–
CD8
–
-тимоцитах. Після інактивації гена цього фактора кількість
тимоцитів зменшується вдвічі.
Невідома також роль молекули CD1, яка експресується у п'ятьох ізоформах (a, b, c, d, е) тимоцитами
тільки на стадії CD4
+
CD8
+
, коли відбувається перебудова генів α-ланцюга ТкР і позитивний відбір
тимоцитів.
Маркерами проліферації тимоцитів є рецептор трансферину (CD71) та молекула CD38, які
експресуються на кортикальних тимоцитах і лише на незначній частині медулярних та зрілих
периферичних Т-клітин.
Зазначимо, що більшість етапів диференціювання Т-клітин відбувається в кортикальній зоні, в
різних її ділянках, у міру просування тимоцитів до медулярної зони, де цей процес завершується.
Міграція тимоцитів усередині тимуса є спрямованим процесом, що чітко регулюється. Важливу
роль у регуляції міграції тимоцитів відіграють хемокіни, що виробляються самими тимоцитами та
тимічними стромальними клітинами.
Відомо, що в тимусі експресується не менш як 12 хемокінів (MDC, ТАКС, 6 CKine, SDF-1, ТЕСК,
ІР-10,1-ТАС, еотаксин, Lptn, МІР-1α, МІР-3α, МІР-3β) і, як вважають, специфічна локалізація
субпопуляцій тимоцитів у різних зонах (кортексі чи медулі) може бути зумовлена їх диференційною
чутливістю до різних хемокінів. У процесі розвитку тимоцити змінюють профіль експресії
хемокінових рецепторів і набувають чутливості до інших хемокінів, що зумовлює їх міграцію з
одних ділянок тимуса до інших. Наприклад, експресія рецептора CXCR4 та хемотактична відповідь
на його ліганд CXCL12 (SDF-1) in vitro спостерігаються на всіх стадіях розвитку тимоцитів. На такі
хемокіни, як CCL22 (MDC) та CCL17 (ТАRС) — ліганди рецептора CCR4, — відповідають тільки
пізньокортикальні та ранньомедулярні тимоцити. Зрілі медулярні тимоцити експресують рецептор
CCR6 і відповідають на його ліганд — хемокін CCL25 (ТЕСК). Перед міграцією з тимуса тимоцити
втрачають експресію CCR6, а з нею і чутливість до ТЕСК, та набувають чутливості до хемокінів, які
залучають їх до вторинних лімфоїдних органів (див. нижче). Є також дані, що локалізовані в
кортексі субпопуляції тимоцитів CD4
–
CD8
–
та CD4
+
CD8
+
чутливіші до дії хемокіну CXCL12 (SDF-1)
а зрілі CD4 та CD8 Т-клітини мозкової речовини – до дії CCL19 (МІР-3β).
У процесі внутрішньотимічної міграції тимоцити взаємодіють з клітинами строми, які впливають на
їх дозрівання. Цей вплив здійснюється через встановлення прямих міжклітинних контактів, які
забезпечують адгезію тимоцитів до епітеліальних та допоміжних клітин тимуса, і продукування
цитокінів. Основою адгезії є взаємодії комплементарних молекул, що експресуються на тимоцитах і
клітинах строми: CD2 з LFA-3 (CD58), LFA-1 (CD11a/CD18) з ІСАМ-1 (CD54). Серед секретованих
цитокінів (ІЛ-1, -3, -6, -7, ГМ-КСФ, ФНП-β) найважливішим для розвитку Т-клітин є ІЛ-7. У людей і
мишей, дефектних за рецептором для ІЛ-7, Т-клітини не розвиваються.
Взаємодії клітин за участю рецепторних, адгезивних і, можливо, унікальних костимуляторних
молекул відіграють важливу роль у реалізації позитивного та негативного відбору Т-клітин за
специфічністю їх ТкР під час розвитку в тимусі.
Відбір репертуару Т-клітин. Зрілі Т-клітини мають бути здатні відповідати на чужорідні антигенні
пептиди в асоціації з власними молекулами МНС (бути МНС-рестриктованими) і не здатні
реагувати на власні антигенні пептиди в комплексі з власними МНС (бути аутотолерантними).
Репертуар МНС-рестриктованих і аутотолерантних Т-клітин формується під час дозрівання їх у
тимусі в процесі подвійного відбору. Перевірка Т-клітин на МНС-рестрикцію здійснюється
позитивним відбором, а перевірка на аутотолерантність — у процесі негативного відбору. МНС-
рестриктовані Т-клітини підтримуються позитивним відбором, а аутореактивні елімінуються
внаслідок апоптозу в процесі негативного відбору. Факторами відбору в обох випадках є комплекси
власних пептидів з МНС, які презентуються тимоцитам як класичними (макрофагами, ДК), так і
некласичними (епітеліальними і мезенхімальними клітинами) АПК. Механізми, за допомогою яких
здійснюються процеси відбору, є предметом інтенсивних експериментальних досліджень (деякі з
них буде розглянуто нижче).
Позитивний відбір Т-клітин. Явище позитивного відбору було відкрито на радіаційних
кістковомозкових химерах, яких отримували введенням клітин кісткового мозку F1-гібридів
генотипу МНС
ахb
смертельно опроміненим мишам батьківського генотипу, наприклад МНС
а
. У цих

54
химер Т-клітини, що розвинулися в тимусі, та всі АПК гемопоетичного походження як у тимусі, так
і на периферії мають генотип МНС
ахb
.
Якщо проімунізувати химер антигеном, то його пептиди будуть презентуватися АПК в асоціації як з
МНС
а
, так із МНС
b
. Однак Т-клітини
химер відповідають in vitro переважно на
антиген, презентований молекулами
МНС
а
. Це свідчить про те, що МНС-
рестрикцію ТкР Т-клітин визначають
стійкіші щодо опромінення клітини
тимічного оточення, в якому вони
дозрівають.
У наступних експериментах на
тимектомованих F1-гібридних мишах,
захищених тимусом батьківського
гаплотипу, було підтверджено, що за
формування МНС-рестрикції і позитивний
відбір Т-клітин відповідальна
радіорезистентна строма тимуса. Схему
експерименту наведено на мал. 89.
Гібридів MHC
axb
F1 після тимектомії та
імплантації тимуса МНС
а
або МНС
b
опромінювали для елімінації власних Т-
клітин і вводили сингенний кістковий
мозок (МНС
ахb
) як джерело СКК.
Проімунізувавши мишей антигеном,
вивчали in vitro проліферативну реакцію
Т-клітин лімфовузлів гібридів на антиген,
презентований АПК батьківських
гаплотипів і для контролю — гібридного
гаплотипу (мал. 89). Т-клітини гібридів з
пересадженим тимусом гаплотипу МНС
а
розпізнавали антиген і проліферували
лише в комплексі з молекулами МНС
а
, але
не МНС
b
, а Т-клітини з тимусом гаплотипу
МНС
b
— лише в асоціації з МНС
b
. Т-
клітини гібридів із сингенним тимусом
були здатні розвивати реакцію на антиген,
що був презентований як на МНС
а
-, так і
на МНС
b
- клітинах. Отже, МНС-
рестрикція визначається МНС-фенотипом
не самих Т-клітин, а фенотипом тимуса, в
якому вони дозрівали. Попереднє
оброблення тимуса дезоксигуанідином, що
руйнує дендритні клітини та макрофаги,
не впливало на виникнення МНС-
рестрикції:
Виникнення МНС-рестрикції відбувається за участю
радіорезистентних стромальних клітин тимуса, якими
є, як з'ясувалося пізніше, кортикальні епітеліоцити
(див. далі).
Велике значення для вивчення процесу позитивного
відбору (та ролі власних молекул МНС у дозріванні
тимоцитів) має використання трансгенних мишей з внесеними повністю реорганізованими генами
ТкР, що кодують рецептор відомої специфічності. У таких мишей переважна більшість Т-клітин
експресують введені гени ТкР, а реорганізація власних генів ТкР інгібується. Отже, отримуючи
трансгенних мишей з наперед відомим ТкР, рестриктованим за певними молекулами МНС, можна
простежити роль саме цих молекул МНС у дозріванні тимоцитів. Показано, що у трансгенних за
ТкР мишей Т-клітини дозрівають лише тоді, коли їхні рецептори рестриктовані за МНС саме цього
гаплотипу. Т-клітини, що не можуть розпізнавати молекули власних МНС на епітелії тимуса, гинуть
на стадії подвійно-позитивних тимоцитів.
У процесі позитивного відбору визначається експресія корецепторів CD4 та CD8 на зрілих Т-
клітинах, яка чітко відповідає специфічності ТкР до власних МНС. Якщо подвійно-позитивний
тимоцит (CD4
+
CD8
+
) розпізнає МНС І, то він перетворюється на монопозитивний СD8
+
-тимоцит, а
якщо МНС II — то на СD4
+
-тимоцит. У мишей, трансгенних за ТкР, рестриктованим за власними
молекулами МНС І, майже всі зрілі Т-клітини експресують цей рецептор і корецептор CD8, а у
мишей, трансгенних за рестриктованим за МНС II рецептором, — корецептор CD4. Залежність
експресії молекул CD4 та CD8 на зрілих Т-клітинах від МНС І- чи МНС ІІ-рестрикції ТкР чітко
простежується в імунодефіцитних людей з синдромом оголених лімфоцитів (bare lymphocyte
syndrome), для якого характерна відсутність експресії молекул МНС на лімфоцитах та епітеліоцитах
тимуса. Особи, в яких втрачена експресія молекул МНС І або МНС II, мають відповідно лише CD4
або CD8 Т-клітини. Така сама залежність спостерігається також у мишей, в яких експресія МНС І чи
МНС II блокована генним нокаутом. Отже, в процесі позитивного відбору Т-клітин у тимусі
відбувається координування експресії рецептора та корецептора у суворій відповідності до молекул
МНС. Дозвіл на виживання і подальший розвиток отримують ті тимоцити, які експресують
корецептор, здатний зв'язувати ту саму молекулу МНС, що й рецептор до антигену. Одночасно з
розгалуженням розвитку тимоцитів у напрямі СD4
+
- чи СD8
+
-клітин відбувається їхнє
програмування до виконання подальших функцій. CD4 Т-клітини майже всі програмуються для
секретування цитокінів, a CD8 Т-клітини — для здійснення цитотоксичної функції.
Механізми, за якими здійснюється вибір експресії корецептора і напряму диференціювання
тимоцитів у CD4 і CD8 Т-клітини, на сьогодні недостатньо вивчені. Важливу роль у цьому процесі
відіграють рецептори родини Notch 1, які експресуються на різних стадіях диференціювання (від
детермінування αβ/γδ-ТкР до позитивного відбору CD4
+
CD8
+
) тимоцитів. Підвищення експресії
рецептора Notch-1 у тимоцитах комітує їхній розвиток у напрямі CD8-клітин, що може свідчити про
роль сигналу, генерованого взаємодією цієї молекули з її лігандом у тимусі, в інгібуванні
диференціювання тимоцитів на зрілі СD4Т-клітини.
Позитивний відбір тимоцитів відбувається у глибоких шарах тимічного кортексу і визначається
кортикальними епітеліальними клітинами. Докази цього були отримані в експериментах на мишах,
трансгенних за молекулами МНС І чи МНС II, які експресувалися лише на кортикальних
епітеліоцитах. Після введення мишам, в яких власні гени МНС були зруйновані, векторів з генами
відсутніх МНС та експресії цих векторів у кортикальних епітеліоцитах розвиток рестриктованих за
відповідними молекулами МНС Т-клітин відбувається нормально. При цьому нормалізація розвитку
Т-клітин спостерігається лише за умови взаємодії молекул МНС на поверхні кортикального
епітеліоцита з відповідним корецептором (МНС І — CD8, МНС II – CD4) тимоцита.
Отже, в процесі позитивного відбору здійснюється перевірка тимоцитів на правильність побудови
Т-рецептора і його здатність впізнавати антигенний пептид у комплексі з МНС. Факторами відбору
є експресовані на кортикальних епітеліоцитах власні пептиди в комплексі з власними молекулами
МНС, а об'єктом відбору — СВ4
+
СВ8
+
-тимоцити, що розпізнають ці комплекси.
У результаті позитивного відбору формується популяція Т-клітин, рецептори яких мають ту чи іншу
спорідненість до власних молекул МНС і здатні розпізнавати в асоціації з ними широкий спектр
пептидів (власних і чужорідних). Ці тимоцити проходять подальшу перевірку на аутореактивність у
процесі негативного відбору.
Негативний відбір Т-клітин. Серед позитивно відібраних у тимусі тимоцитів є специфічні до
досить поширених власних антигенів. Після дозрівання в тимусі та виходу на периферію вони
можуть завдати шкоди організму, ушкоджуючи нормальні клітини, що експресують аутоантигени.
Для запобігання цьому потенційно аутореактивні Т-клітини елімінуються в тимусі внаслідок
інтенсивного негативного відбору. Основою відбору є відповідь тимоцита під час розвитку на
комплекси власних пептидів з власними МНС. Елімінуються ті тимоцити, які, впізнаючи ці
комплекси, виявляють певну реактивність щодо них.
Негативний відбір тимоцитів здійснюють тимічні клітини кількох типів, серед яких
найефективнішими є професійні АПК — дендритні клітини та макрофаги. Миші-химери, отримані
трансплантацією кісткового мозку гібридів МНС
ахb
мишам батьківської лінії МНС
а
, не відторгають
пересаджені клаптики шкіри мишей генотипів як МНС
а
, так і МНС
b
. У цих мишей підтримується
толерантність не тільки до власних пептидів у комплексі з власними МНС
а
, а й до комплексів
пептидів з МНС
b
. Толерантність сформувалася в результаті негативного відбору (елімінації)
тимоцитів цими комплексами, презентованими тимічними клітинами. В тимусі химер презентувати
комплекси МНС
b
з пептидами здатні тільки ДК і макрофаги, що походять із пересадженого
кісткового мозку. Вони здебільшого й опосередковують негативний відбір тимоцитів.
Крім ДК і макрофагів у здійсненні негативного відбору тимоцитів певну роль відіграють також
епітеліальні клітини медулярної зони тимуса. Це доведено в експериментах на мишах, трансгенних
за такими білками, як панкреатичний гормон соматостатин та печінковий С-реактивний білок, які
експресуються, як з'ясувалося в дослідах, й невеликою часткою медулярних епітеліоцитів. Як
вважають, на відміну від ДК, що здійснюють делецію аутореактивних Т-клітин, медулярні
епітеліоцити можуть індукувати аутотолерантність за допомогою ще й інших механізмів.
Процес негативного відбору Т-клітин безпосередньо у тимусі вдалося простежити у мишей, які
експресують ендогенні суперантигени (Mls — мінорні лімфоцитстимулювальні антигени), що
кодуються інтегрованим у хромосому геномом мишачого вірусу пухлини молочної залози (MMTV).
На відміну від звичайних антигенів, які зв'язуються з активним центром специфічного Т-рецептора,
Mls-білки (як й інші суперантигени) зв'язують з високою афінністю молекули МНС та Vβ-домени
певних типів ТкР незалежно від антигенної специфічності останніх. Експресовані в тимусі Mls-
білки зв'язуються з великою кількістю тимоцитів з ТкР відповідного типу, індукуючи у них апоптоз,
і сприяють виникненню у мишей специфічної толерантності. Показано, що у тимічному кортексі
уражених вірусом MMTV мишей міститься велика кількість СD4
+
СD8
+
-тимоцитів, що експресують
рецептори, здатні зв'язатися з Mls-білком, тоді як у медулярній зоні (та на периферії) вони відсутні.
Зважаючи на це, було зроблено припущення, що клональна делеція специфічних до Mls лімфоцитів
відбувається вже на пізніх стадіях розвитку тимоцитів під час міграції з кіркової до медулярної зони
в ділянці їх з'єднання, де зосереджена велика кількість ДК.
Отже, зв'язування ТкР одного і того самого комплексу власного МНС з власним пептидом індукує
різні процеси: подальший розвиток тимоцитів за позитивного відбору та загибель їх за негативного
відбору. Як це відбувається сьогодні, ще невідомо, але для пояснення механізмів цих процесів
запропоновано дві гіпотези — гіпотезу авідності та гіпотезу диференційної сигналізації.
Згідно з гіпотезою авідності, виживання чи загибель тимоцита залежить від сили сигналу, що
передається від рецептора, який розпізнав антиген, усередину клітини. Слабкий або частковий
(неповний) сигнал індукує виживання клітини (позитивний відбір), а сильний сигнал — її загибель
(негативний відбір). Сила сигналу залежить від афінності рецептора та щільності розпізнаних
комплексів пептид—МНС на поверхні кортикальних епітеліоцитів тимуса. Тимоцити, що зв'язують
комплекси власний пептид —МНС з високою афінністю, елімінуються, але такий механізм діє лише
щодо незрілих клітин. Зв'язування з високою афінністю цих комплексів зрілими Т-клітинами
призводить до їх активації.
Загибель тимоцитів у разі негативного відбору може бути зумовлена також відсутністю додаткових
(костимуляторних чи інших) сигналів. Можливо, при цьому важливу роль відіграє природа сигналу,
як це передбачає гіпотеза диференційної сигналізації. За цією гіпотезою, сигнали позитивного і
негативного відбору тимоцитів різняться не кількісно, а якісно. На користь цього припущення
свідчать результати досліджень, в яких сигнали від зв'язаних з антигеном рецепторів імітували за
допомогою пептидів-антагоністів та перехреснозшивних реагентів.
Пептиди-антагоністи є пептидами, що здатні заміняти собою оригінальний антигенний пептид у
комплексах МНС —пептид. ТкР взаємодіє з комплексом пептид-антагоніст — МНС під іншим
кутом, ніж з оригінальним комплексом МНС — пептид, унаслідок чого корецептор Т-клітин
усувається від участі у проведенні сигналу, оскільки «не може доставити» зв'язану з ним кіназу Lck
до цитоплазматичних ділянок CD3. Зріла Т-клітина не отримує повного сигналу і не відповідає на
цей антиген. Отже, пептиди-антагоністи можуть бути ефективною моделлю для вивчення ролі
корецепторів у процесах селекції тимоцитів. Перехреснозшивні реагенти, або крос-лінкери, — це
реагенти, що здатні ковалентно зшивати наближені один до одного рецептори або корецептори Т-
клітин.
В експериментах використовували дві трансгенні культури тканини тимуса. Одна культура
експресувала молекули МНС, які утворювали комплекси з пептидами-антагоністами та які були
здатні взаємодіяти лише з певним трансгенним ТкР. Інша культура була дефектна за молекулами
МНС обох класів та експресувала ТкР однієї специфічності. Було досліджено, відбір яких тимоцитів
(CD4 чи CD8) відбувається в разі надходження до клітини різних сигналів, а саме, під час зшивання
крос-лінкерами лише ТкР, ТкР з корецепторами та під час утворення потрійного комплексу ТкР —
пептид-антагоніст — МНС.
Застосування такого методичного підходу дало змогу встановити, що позитивний відбір CD4- і
CD8-тимоцитів відбувається за різних умов. Зшивання одних лише ТкР крос-лінкерами або
зв'язування ТкР комплексами пептид-антагоніст — МНС зумовлює позитивний відбір CD8 Т-
клітин, але не CD4 Т-клітин. Для розвитку CD4 Т-клітин і негативного відбору CD8 Т-клітин
потрібні сигнали, які генеруються зшиванням ТкР з корецепторами (CD4, CD8) і отже, є подібними
до сигналів, що індукуються в нормі під час активації. З урахуванням цього припускають, що
позитивний відбір CD8 Т-клітин може індукуватися зв'язуванням ТкР тимоцитів з комплексами
ендогенний пептид — МНС на кортикальних епітеліоцитах, за якого генерується слабкий або
частковий сигнал. Позитивний відбір CD4 Т-клітин може індукуватися лише сильним сигналом,
рівнозначним активаційному, який генерується зв'язуванням Т-рецептора і будь-якого з двох
корецепторів. Існує також думка, що кортикальні епітеліоцити можуть експресувати унікальні, поки
що невідомі молекули, які передають костимуляторні або допоміжні сигнали під час позитивного
відбору тимоцитів і відрізняються від молекул, задіяних під час активації зрілих Т-клітин.
Основою негативного відбору за механізмами апоптозу, як вважають, можуть бути взаємодії
молекул CD30, що експресуються на активованих аутоантигенами тимоцитах фенотипу
CD4+CD8+CD45RO+ CD30
+
IL-4R
+
, з їхніми лігандами CD30L, які містяться у великій кількості на
ДК та епітеліоцитах мозкової зони.
Слід зазначити, що уявлення про елімінацію аутореактивних Т-клітин у тимусі не пояснює, як
формується толерантність імунної системи до власних антигенів, що наявні лише в периферичних
тканинах. Відомо, що на периферії усунення аутореактивних клітин від імунної відповіді
реалізується переважно за рахунок інших механізмів (див. розд. 13), а не апоптозу, який є основним
механізмом реалізації негативного відбору тимоцитів у тимусі.
Отже, розвиток Т-лімфоцитів з попередників і пов'язані з ним процеси відбору супроводжуються
інтенсивною проліферацією та масовою загибеллю тимоцитів. Кожної доби у тимусі молодих
дорослих мишей утворюється приблизно 5·10
7
нових клітин, а виходить із тимуса лише 1·10
6
- 2·10
6
зрілих Т-клітин. Oднак кількість клітин у тимусі залишається постійною і становить 1·10
8
- 2·10
8
.
Більшість тимоцитів, що розвиваються в тимусі (близько 98 %), гинуть унаслідок апоптозу. Тому
тимус прирівнюють до «інтенсивно працюючої фабрики смертників». Проліферативний потенціал
схильність до апоптозу змінюються в процесі розвитку Т-лімфоцитів, і тимоцити на різних стадіях
дозрівання різняться за цими властивостями. Найбільшою проліферативною активністю
характеризуються ранні подвійно-негативні (CD4
–
CD8
–
) тимоцити. Завдяки цьому забезпечується
постачання необхідної кількості CD4
+
CD8+-тимоцитів — «матеріалу» для ефективного відбору, в
процесі якого більшість цих клітин «вибраковується».
Схильність до апоптозу, як відомо, залежить від співвідношення експресії в клітині факторів, що
сприяють та запобігають цьому процесу, зокрема молекули Fas (CD95), яка є провідником апоптозу,
та молекули Всl-2, що захищає від апоптозу. Більшість (80-90 %) CD4+CD8+-тимоцитів, що
зазнають позитивного відбору, несуть молекулу Fas і лише незначна частка їх (5-10 %) містить
молекулу Всl-2. Серед їх попередників — CD4
–
CD8
–
-тимоцитів лише 10—15 % клітин експресують
Fas і 25 —40 % — молекулу Всl-2, а їхні нащадки монопозитивні CD4
+
CD8
–
і СD4
–
СD8
+
-тимоцити
всі без винятку несуть Всl-2. При цьому значна частина CD4+CD8
–
(48-65 %) і СD4
–
СD8
+
(40 – 68%)
тимоцитів експресує молекулу Fas, з якою пов'язана регуляція їх проліферації та функціонування.
Серед трьох субпопуляцій тимоцитів CD4
+
CD8
+
є найчутливішими до дії індукторів апоптозу. Саме
тому на дію багатьох стимулювальних впливів вони відповідають не активацією, а розвитком
апоптозу Тому, як вважають, сильні сигнали, що надходять від ДК (індуковані зв'язуванням
комплексів МНС — пептид), які зазвичай зумовлюють активацію зрілих Т-клітин, у незрілих
тимоцитів індукують апоптоз. Цим пояснюється і висока чутливість кортикальних СD4+СD8+-
тимоцитів до випромінювання та кортикостероїдів. Фармакологічні дози цих гормонів
спричинюють в експерименті майже повне виснаження (99 %) популяції тимоцитів. В індукуванні
апоптозу тимоцитів під час відбору важливу роль відіграє внутрішньотимічний гормон
прегненолон, продуцентами якого є епітеліальні клітини тимуса. Медулярні тимоцити нечутливі до
дії кортикостероїдів, що зумовлюється як підвищенням у них експресії гена Всl-2, так і насамперед
наявністю ферменту 20α-гідроксилстероїддегідрогенази.
Однією з причин загибелі тимоцитів може бути також накопичення в них токсичних метаболітів
пуринового обміну — ГТФ та дезоксиГТФ. Це зумовлено підвищеною активністю в тимоцитах
порівняно зі зрілими Т-клітинами екто-5'-нуклеотидази та аденозиндезамінази і зниженою
активністю пуриннуклеотидилфосфорилази — ферментів, що каталізують послідовні фази
перетворення пуринових нуклеотидів.
Згідно з сучасними уявленнями, розвиток Т-клітин відбувається у дві стадії відбору, які
здійснюються за поки що до кінця не з'ясованими механізмами різними клітинами тимічного
мікрооточення внаслідок контактних взаємодій з тимоцитами та, можливо, за допомогою
секретованих цитокінів. Основою відбору є перевірка антигенрозпізнавальних рецепторів
тимоцитів, що розвиваються, на ауто-МНС-рестрикцію та аутотолерантність. Підраховано, що
приблизно 2% з утворених у тимусі тимоцитів переживають подвійний скринінг, дозрівають до
монопозитивних Т-клітин і виходять на периферію. Відібрані клітини залучаються до проліферації
для утворення клонів антигенспецифічних Т-лімфоцитів. Специфічністю відібраних Т-клітин і
визначається клональний репертуар популяції зрілих Т-лімфоцитів, які після виселення на
периферію забезпечують захист від чужорідних антигенів.
12.3. РОЗВИТОК В-ЛІМФОЦИТІВ.
12.3.1. Розвиток В2-лімфоцитів у кістковому мозку.
В-лімфоцити, так само, як і Т-лімфоцити, проходять складний процес розвитку, що
супроводжується формуванням широкого репертуару специфічностей їхніх рецепторів. Оскільки
диференціювання В-клітин вимагає перебудови генів, що кодують антигенспецифічні рецептори,
більшість етапів розвитку В-клітин аналогічні до етапів розвитку Т-клітин.
На відміну від розвитку Т-лімфоцитів незалежне від антигену диференціювання В-лімфоцитів у
ссавців майже повністю відбувається у кістковому мозку. У птахів є спеціалізований орган для
розвитку В-клітин — сумка Фабриціуса, яка заселяється стовбуровими клітинами ще під час
ембріонального розвитку птахів і впродовж життя генерує нові В-лімфоцити. В-лімфопоез у ссавців
залежить від локального мікрооточення, яке формується у кістковому мозку.
Мікрооточення в кістковому мозку. Кровотворний кістковий мозок розміщений у комірках
трубчастих кісток (див. розд. 1). З внутрішнього боку кісткова тканина вкрита ендостом —
спеціалізованою сполучною тканиною. Біля ендосту знаходяться власне СКК, які у міру
диференціювання переміщуються від стінок комірки до середини. В середині комірки проходять
судини, крізь стінки яких зрілі клітини надходять у кров. Навколо синусоїдів розміщені ретикулярна
тканина та острівці жирової тканини, а також місцеві макрофаги. Ретикулярні клітини утворюють
волокна, встановлюють адгезивні контакти з СКК, синтезують ростові фактори і хемокіни, які
регулюють утворення та міграцію клітин різних типів. Зокрема, для комітування СКК у бік
лімфоїдного ряду головну роль відіграє ІЛ-7, що синтезується ретикулоцитами. Завдяки здатності
утворювати безпосередні контакти з СКК ретикулярні клітини надають певні костимуляторні
сигнали, необхідні для комітування тих чи інших клітин та їх міграції у міру дозрівання. Завдяки
жорсткій структурі кісткова тканина обмежує об'єм кісткового мозку, а жирова — створює
необхідний тиск для підтримання функціонування капілярів і виходу зрілих клітин у циркуляцію.
Судини кісткового мозку адаптовані до забезпечення переходу нових клітин крові у судинне русло.
Процес міграції вибірковий, оскільки до циркуляції надходять лише зрілі клітини. Функцію
розпізнавання зрілих клітин виконують клітини, які «обгортають» венозний синусоїд —
адвентиційні клітини («клітини-ворота»), які мають морфологію, характерну для ретикулярних
клітин з великими відростками. Вони є «пропускним пунктом» для виходу клітин з кісткового мозку
в кров. Ендотеліальні клітини судин мають лабільні контакти між собою, що забезпечує здатність
утворювати пори, крізь які проходять клітини.
В-лімфопоез відбувається у прошарку сполучної тканини навколо дрібних артеріол. Починається
розвиток В-клітин біля ендосту, де проходить попереднє комітування СКК. У міру дозрівання
попередники В-лімфоцитів мігрують у напрямі до синусоїдів, постійно підтримуючи контакти з
ретикулярними клітинами.
Стадії диференціювання В2-лімфоцитів у кістковому мозку. Основними подіями під час
антигеннезалежного диференціювання В-лімфоцитів у кістковому мозку є перебудова генів
імуноґлобулінів, унаслідок чого на поверхні В-клітини має експресуватися повноцінний
імуноґлобуліновий рецептор. Залежно від стадії перебудови генів імуноґлобулінів
антигеннезалежний розвиток В-клітин у кістковому мозку поділяють на чотири стадії.
1. Ранні про-В-клітини (стадія А).
2. Пізні про-В-клітини (про-В1)* (стадія В). *У дужках позначено альтернативні назви цих стадій,
що також трапляються у спеціальній літературі.
3. Великі пре-В-клітини (великі пре-ВІІ) (стадія С).
4. Маленькі пре-В-клітини (маленькі пре-ВІІ) (стадія D).
Процес антигеннезалежного диференціювання В-клітин у кістковому мозку закінчується
утворенням клітин з повністю сформованим ВкР. Цю стадію називають стадією незрілих клітин
(стадія Е). Незрілі В-клітини виходять із кісткового мозку і заселяють селезінку, де завершуються
процеси їх дозрівання, тобто набуття здатності відповідати на розпізнавання антигену. Закінчується
цей процес формуванням зрілих наївних лімфоцитів (стадія F), які переходять до рециркуляції та
заселяють вторинні лімфоїдні органи.
Розглянемо докладніше стадії антигеннезалежного диференціювання В-клітин (мал. 90).
Ранні про-В-клітини. Ранні про-В-клітини — це попередники В-лімфоцитів, які ще не мають
рецепторів до антигену, але вже визначили свій подальший розвиток. У цих клітинах починає
відбуватися реаранжування D-J-сеґментів у локусі генів важкого ланцюга імуноґлобулінів. При
цьому D-J-реаранжування не зазнає алельного виключення, тобто воно може відбуватися в обох
хромосомах. Важливою передумовою активації процесів реаранжування генів є експресія ферментів
рекомбіназ RAG1 і RAG2 у цих клітинах. Оскільки D-сеґмент потенційно може транслюватися в
трьох різних рамках зчитування, то, ймовірно, ніякого контролю за «правильністю» перебігу D-J-
реаранжування в клітинах не відбувається.
Слід зазначити, що після з'єднування сегментів D і J може активуватися промотор перед сегментом
D, внаслідок чого може експресуватися мРНК-транскрипт неповного гена важкого ланцюга. Однак
функціональне значення такої експресії залишається невідомим, крім того, не встановлено, чи існує
білковий продукт, що може зчитуватися з такого мРНК-транскрипту.
Стадія розвитку ранніх про-В-клітин характеризується експресією на поверхні CK45R (В220), СК10,
СК19, СК38, c-Kit, ІL-7R. Деякі з них характерні ще для стовбурових клітин-попередників (СК34
СК45, c-Kit, IL-7R), а інші – лише для В-клітин (СК10, СК19, СК38).
Крім того, в цитоплазмі таких клітин починають екс пресуватися молекули, необхідні для
формування пре-В-клітинного рецептора: молекули для передавання сигналу Іgα і Іgβ, молекули λ5 і

55
VpreВ, які складають інваріантний легкий ланцюг рецептора, а також кілька транскрипційних
факторів – ЕВF, Е2А, Рах5 (див. далі).
Ймовірно, ранні про-В-клітини є зворотною стадією розвитку, оскільки ознаки D-J-реаранжування в
Н-локусі генів імуноглобулінів можна виявити навіть у лейкоцитах мієлоїдного ряду.
Пізні про-В-лімфоцити. Після завершення D-J-реаранжування в Н-локусі починається V-DJ-
реаранжування, тобто приєднання V-генів до об'єднаних раніше DJ-сеґментів. З V-DJ-
реаранжування починається стадія пізніх про-В-лімфоцитів. V-DJ-реаранжування
підпорядковується правилу алельного виключення, тобто відбувається лише в одній з пари
гомологічних хромосом. Крім активації продуктів генів RAG1 і RAG2 V-DJ-реаранжування
потребує участі ферменту ТdТ, яка вносить додаткові нуклеотиди у зону з'єднання V- і DJ-сеґментів.
Важливо, що під час утворення В-клітин в ембріональній печінці, а також у перші дні після
народження фермент TdT не активується. Крім того, активація TdT не відбувається під час розвитку
В1-клітин.
Головним наслідком V-DJ-реаранжування має стати повноцінний ген важкого ланцюга
імуноґлобулінів. Для цього потрібно, щоб V-, J- і С-сеґменти знаходилися у необхідній рамці
зчитування. Оскільки процес соматичної рекомбінації під час реаранжування може відбуватися з
помилками, слід контролювати «якість рекомбінації». Тому після V-DJ-реаранжування клітина
експресує утворений ген важкого ланцюга. Отже, пізні про-В-клітини можуть експресувати лише
один з поліпептидних ланцюгів, що входять до складу антигенспецифічних рецепторів.
Для того щоб на поверхні В-клітин міг експресуватися повний імуноґлобуліновий рецептор, має
відбутися об'єднання важкого ланцюга з легким. Оскільки гени легких ланцюгів залишаються ще не
перебудованими, пізня про-В-клітина використовує інваріантний (сурогатний) легкий ланцюг для
утворення повної імуноґлобулінової молекули. Сурогатний легкий ланцюг складається з двох
молекул: λ5-субодиниці, що аналогічна С-домену легкого ланцюга, і VpreB-субодиниці, що
аналогічна V-домену легкого ланцюга. Комплекс двох молекул Н-ланцюга з двома молекулами λ5 і
двома VpreB називають пре-ВкР. Якщо гени важкого ланцюга були перебудовані правильно, то пре-
ВкР збирається і може експресуватися на мембрані разом з молекулами Igα і Igβ. Сигнал від пре-
ВкР необхідний для проходження наступних стадій диференціювання. Однак невідомо, чи розпізнає
будь-який ліганд пре-ВкР. Ймовірно, що ні, оскільки перехресне зв'язування пре-ВкР на поверхні
про-В-клітин за допомогою МкАТ проти λ5-ланцюга чи μ-ланцюга не впливає на розвиток про-В-
клітин in vitro Можливо, що сам факт експресії пре-ВкР на мембрані є сигналом до позитивної
селекції про-В-лімфоцитів. Як показали С. Janeway і співавтори, у мишей, не здатних переключити
експресію мембранних імуноґлобулінів на синтез розчинних антитіл, подальший розвиток про-В-
клітин блокується. Очевидно, лігандом, який розпізнають про-В-лімфоцити за допомогою пре-В-
клітинного рецептора, є розчинні антитіла, синтезовані іншими попередньо активованими В-
лімфоцитами.
Якщо V-DJ-реаранжування відбулося з помилками, внаслідок чого з'являється нефункціональний
важкий ланцюг, який не може утворити комплекс із сурогатним легким ланцюгом, то в клітині
включається реаранжування локусу Н-генів в іншій хромосомі. Ймовірність вдалої рекомбінації
генів важкого ланцюга на одній хромосомі становить 1/3, тому ймовірність того, що хоча б на одній
із хромосом процес відбудеться правильно, становить 2/3. Отже, близько 30 % пізніх про-В-кліти
гине внаслідок нездатності правильно реаранжувати гени важкого ланцюга імуноґлобулінового
рецептора.
Пізні про-В-клітини експресують ті самі поверхневі маркери, що й ранні про-В-клітини, і ще ряд
додаткових маркерів – CD20, CD40. Крім того, вони втрачають експресію рецептора c-Kit до
фактора стовбурових клітин і починають експресувати CD25 - α-ланцюг рецептора до ІЛ-2. Лише
частина цих клітин експресує поверхневий рецептор пре-ВкР, але у цитоплазмі всіх клітин
виявляється білковий продукт трансляції генів важкого ланцюга.
Великі пре-В-лімфоцити. Після вдалого завершення процесів перебудови сегментів в Н-локусі
генів імуноґлобулінів попередники В-клітин переходять до наступної стадії диференціювання.
Природа намагається зберегти такі клітини і збільшити їхню кількість, оскільки процес
реаранжування генів не завжди приводить до продуктивного результату. Тому клітини з
перебудованими генами важкого ланцюга починають розмножуватися й утворювати клон. Під час
розмноження пре-В-клітини набувають морфології бластних клітин, тому вони мають великі
розміри, через що й дістали свою назву. У великих пре-В-клітннах не спостерігається наявність
активних ферментів-рекомбіназ RAG1 і RAG2 та ферменту TdT, хоча в цитоплазмі міститься мРНК
цих ферментів. Кожна така клітина може здійснити близько шести поділів, а отже, утворити клон з
64 однакових пре-В-клітин. На поверхні великих пре-В-клітин завжди міститься пре-ВкР, хоча
функціональне значення його експресії залишається невідомим. Такі клітини продовжують
експресувати рецептори до ІЛ-2 і ІЛ-7, а також набувають нових поверхневих маркерів —
амінопептидазу ВР-1, молекулу CD24 тощо.
Маленькі пре-В-клітини. Після розмноження великих пре-В-клітин вони зменшують свої розміри і
починають реаранжувати гени легкого ланцюга імуноґлобулінів. Для цього знову активується
експресія генів RAG1 і RAG2. Спочатку відбувається перебудова генних сегментів у κ-локусі генів
легкого ланцюга. При цьому спостерігається явище алельного виключення. Як правило, перша
рекомбінація наближує певний Vκ-сегмент до Jκ1-сегмента. Правильно перебудований κ-ланцюг
експресується разом з важким μ-ланцюгом на поверхні В-клітини. Якщо така експресія стає
можливою, подальші процеси перебудови припиняються, а якщо перша V-J-рекомбінація генів κ-
ланцюга виявляється непродуктивною, — включається рекомбінація генів на іншій хромосомі. У
разі непродуктивності рекомбінації в обох κ-локусах включається перебудова генів у λ-локусі. Тому
у мишей співвідношення імуноґлобулінів з κ-ланцюгом до імуноґлобулінів з λ-ланцюгом становить
10:1. У людини це співвідношення приблизно 3:1. Отже, після завершення стадії малих пре-В-
лімфоцитів утворюються незрілі В-клітини, які мають сформовані IgM-рецептори до антигену, але
ще не пройшли негативного відбору. IgM-рецептор експресується на мембрані разом з молекулами,
необхідними для передавання сигналу (Igα і Igβ, фосфатазою CD40R, корецептором CD 19), а також
МНСІІ та іншими маркерами (CD20, CD24, CD40, ВР-1 тощо). Маленькі В-лімфоцити втрачають
експресію рецепторів до інтерлейкінів 2 і 7.
Отже, антигеннезалежний розвиток В-лімфоцитів пов'язаний з перебудовою генів їхніх рецепторів.
Головним наслідком його має бути експресія повноцінного білкового продукту, який може
зчитуватися з перебудованих генів та експресуватися на мембрані клітин. Про-В-лімфоцити
перебудовують гени важкого ланцюга імуноґлобулінів, а пре-В-лімфоцити — легкого. Під час цих
перебудов відбувається перевірка правильності перебудови, яку можна розглядати як позитивний
відбір В-лімфоцитів.
У миші за добу утворюється приблизно 5·10
7
незрілих В-клітин. В-клітини, які утворилися
внаслідок антигеннезалежного диференціювання, зазнають негативного відбору. Підраховано, що
на периферію надходить лише 10 % утворених у кістковому мозку клітин. Усі інші В-клітини, як
вважають, гинуть унаслідок апоптозу завдяки аутореактивності їхніх рецепторів.
Відбір репертуару В2-клітин. Розвиток В-клітин — досить складний процес і тому потребує
кількох «точок контролю». В таких точках контролю звіряється відповідність програми розвитку
лімфоцита і визначається його подальша доля. Якщо лімфоцит відповідає необхідним вимогам, він
продовжує подальший розвиток, а якщо ні, — то гине. Як і Т-клітин, під час розвитку В-лімфоцитів
відбувається їхній позитивний та негативний відбір.
Позитивний відбір можна назвати «контролем якості» клітин. Такий відбір зумовлює появу клітин з
функціонально активними рецепторами, оскільки клітини, які виявилися нездатними правильно
реаранжувати гени імуноґлобулінів, гинуть під час розвитку. Головним наслідком першого етапу
позитивного відбору є відбір клітин, що експресують функціональний важкий ланцюг
імуноґлобулінів у складі пре-ВкР, а другого — функціональний легкий ланцюг у складі ВкР. Донині
невідомо, що за сиґнал отримують попередники В-лімфоцитів від пре-ВкР. Не з'ясовано також, чи
існує певний ліганд, який має зв'язати пре-ВкР. Найімовірніше, що сигналом про правильну
перебудову генів важкого ланцюга рецептора є власне експресія пре-ВкР на мембрані незалежно від
зв'язування ліганду. На користь цієї гіпотези свідчить той факт, що МкАТ, специфічні до певних
компонентів пре-ВкР, а саме, λ5, VpreB або μ-ланцюга, не впливають на розвиток пре-В-клітин.
Водночас антитіла проти μ-ланцюга повністю блокують розвиток незрілих В-лімфоцитів, у яких уже
сформований IgM-peцептор.
Негативний відбір В-лімфоцитів можна назвати «контролем безпечності» клітин, оскільки під час
цього процесу знищуються небезпечні та небажані клітини, які несуть специфічні до власних
антигенів рецептори. Існування негативної селекції передбачав ще Бернет у своїй теорії селекції
клонів.
Негативний відбір В-лімфоцитів може відбуватися у два етапи — на стадії маленьких пре-В-клітин
у кістковому мозку та на стадії незрілих В-клітин у селезінці, куди вони мігрують з кісткового
мозку і де завершується їхнє дозрівання. Відповідно перший етап негативного відбору називають
центральною толяризацією В-клітин, а другий — периферичною.
Якщо незрілі В-клітини зв'язують власні антигени в кістковому мозку чи в селезінці, вони не здатні
активуватися, а швидше загинуть чи залишаться назавжди нездатними активуватися. Однак існує
механізм, який може «залишити живими» аутоспецифічні незрілі В-клітини, що розпізнали
аутоантиген у кістковому мозку, і змінити специфічність їхніх рецепторів. Деякі В-клітини
повторно можуть активувати експресію ферментів-рекомбіназ RAG1 і RAG2 і здійснювати
подальше реаранжування генів легких ланцюгів рецептора. Якщо першими до Vκ-сеґмента
приєднується Jκ1-сегмент, то подальші перебудови генів легкого ланцюга можуть використовувати
інші Vκ-сегменти і з'єднувати їх з Jκ2-, Jκ3-, Jκ4-сеґментами тощо. Такий процес подальшої
перебудови генів легкого ланцюга називають процесом редагування рецепторів. Ці процеси
характерні також для розвитку Т-клітин, коли після високоафінного розпізнавання аутоантигенів у
тимусі може відбуватися додаткове реаранжування генів α-ланцюга ТкР.
У деяких випадках взаємодія рецепторів незрілих В-клітин з аутоантигенами призводить до
диференціювання таких В-клітин у В1-клітини, які продукують поліспецифічні антитіла.
12.3.2. Розвиток В1-клітин.
На відміну від традиційного розвитку В2-клітин у кістковому мозку розвиток В1-клітин
відбувається на периферії. Вважають, що комітовані попередники В1-клітин закладаються у
фетальній печінці у процесі ембріонального розвитку, після чого заселяють черевну і плевральну
порожнини, а також великий і малий сальники, де і підтримують свій подальший розвиток. Такі
клітини можуть відновлювати свою популяцію, тобто здатні до розмноження без диференціювання.
Мабуть, саме тому більшість відомих лімфом є переродженими В1-клітинами, які експресують
поверхневий маркер CD5.
В1-клітини у своєму розвитку проходять етапи, аналогічні розвитку В2-клітин. Однак під час
перебудови генів важкого ланцюга В1-клітини використовують лише V-генні сегменти, які
знаходяться поблизу D-сегментів. Крім того, в таких клітинах не активується фермент ТdТ, а отже,
місця з'єднання V-, D- і J-сегментів не мають вставок додаткових нуклеотидів. Тому різноманіття
рецепторів В1-клітин більш обмежене, ніж рецепторів В2-клітин, але їхні рецептори здатні зв'язати
ширший спектр антигенів.
12.4. ТРАНСКРИПЦІЙНІ ФАКТОРИ, ЩО ВИЗНАЧАЮТЬ ДИФЕРЕНЦІЮВАННЯ
ЛІМФОЦИТІВ.
Транскрипційні фактори, що визначають розвиток лімфоїдних клітин, перебувають у певній
ієрархії, тобто ті фактори, що активувалися раніше, активують фактори, необхідні для проходження
наступних стадій диференціювання.
На підставі експериментальних даних, які були накопичені за останні роки, Т. Enver і М. Greaves
зробили припущення, що для комітування СКК до певного розвитку потрібно, щоб відбулася
активація специфічних для цієї лінії розвитку генів і стійка репресія генів, що є характерними для
інших ліній розвитку. Властивість активувати та репресувати різні генетичні програми може мати
навіть один транскрипційний фактор, але, як правило, для цього необхідно кілька різних
транскрипційних факторів.
Роль фактора PU. 1 в комітуванні стовбурових клітин. Першим транскрипційним фактором, що
значно обмежує потенціал СКК, є фактор PU.1, який експресується лише в гематопоетичних
клітинах, що можуть дати початок усім білим клітинам крові (лейкоцитам), але не еритроцитам та
мегакаріоцитам. Нокаутні за PU.1 миші гинуть через 18 діб ембріонального розвитку, і їхні
ембріони позбавлені макрофагів, гранулоцитів та лімфоцитів, хоча мають нормальну кількість
еритроцитів і мегакаріоцитів. Іноді у таких ембріонах виявляють попередники Т-клітин.
Встановлено, що фактор PU.1 активує і підтримує експресію на СКК рецепторів до ІЛ-7, що
необхідні для розвитку лімфоїдних клітин, та рецепторів до М-КСФ, необхідних для розвитку
лейкоцитів мієлоїдного ряду (переважно макрофагів).
Отже, відкриття фактора PU. 1 свідчить про те, що, можливо, лейкоцити мієло- та лімфоїдного рядів
більше «споріднені» між собою, ніж це передбачає загальноприйнята схема гематопоезу.
Транскрипційні фактори, що контролюють розвиток В-клітин. Специфічними молекулярними
маркерами комітування СКК до розвитку в В-лімфоцитарному напрямі, як уже зазначалося, є перші
ознаки перебудови генів важкого ланцюга імуноґлобулінів (D-J-рекомбінація) та експресія генів
сурогатного легкого ланцюга, тобто генів, що кодують модулі передавання сигналу Igα та Igβ від
майбутнього ВкР. Для розвитку В-клітин необхідна попередня активація відповідних
транскрипційних факторів ще до того, як з'являються перші з наведених ознак комітування.
Активація транскрипційних факторів Е2А і EBF під час розвитку В-клітин. Перші етапи В-
лімфопоезу визначають два транскрипційних фактори — Е2А і EBF, які разом активують генетичну
програму розвитку В-клітин.
Активація факторів Е2А і EBF є необхідною, але недостатньою умовою для розвитку В-клітин із
СКК. Ці фактори діють скоординовано, оскільки порушення генів кожного з них призводить до
повного блокування утворення В-клітин. Мішенями їхньої дії є гени сурогатного легкого ланцюга
імуноґлобулінів — λ5 і VpreB, гени рекомбіназ RAG1 і RAG2, гени сигнальних молекул рецептора
Igα (CD79α) і Igβ (CD79β). Ці фактори також регулюють експресію перебудованого μ-ланцюга
імуноґлобулінів, можливо, навіть після D-J-рекомбінації. Отже, фактори Е2А і EBF контролюють
синтез молекул, які необхідні для експресії пре-ВкР на мембрані та передавання сигналу від нього.
В-клітини Е2А-дефектних мишей не експресують корецептор CD19, крім того для них не характерні
перебудови в Н-локусі генів імуноґлобулінів. В-клітини EBF-дефектних мишей не експресують
сурогатний легкий ланцюг імуноґлобулінів і також не здатні активувати процеси перебудови генів
імуноґлобулінів.
Після факторів Е2А і EBF активується експресія транскрипційного фактора Рах5, який визначає
подальший перебіг процесів диференціювання попередників В-клітин.
Роль транскрипційного фактора Рах5 у диференціюванні В-клітин. Як і фактори Е2А і EBF,
фактор Рах5 починає експресуватися на ранніх стадіях розвитку попередників В-клітин і продовжує
експресуватися навіть до стадії плазматичних клітин. Нокаутування гена Рах5 зумовлює
припинення розвитку попередників В-клітин на стадії ранніх про-В-клітин. Такі клітини мають
перебудовані D-J-сеґменти генів у Н-локусі та експресують ряд генів, характерних для В-клітинного
диференціювання, за винятком CD19 та Igα. Ці клітини можуть проліферувати як завгодно довго під
дією ІЛ-7 та стромальних клітин кісткового мозку, але не виявляють при цьому ознак подальшого
диференціювання. Якщо ген Рах5 ввести в клітини за допомогою ретровірусного вектора, вони
починають нормально диференціюватися у зрілі В-клітини.
Однак з'ясувалося, що у Рах5
–
/
–
мишей про-В-клітини відрізняються від про-В-клітин мишей дикого
типу. Про-В-клітини Рах5
–
/
–
мають багато ознак СКК і можуть дати початок різним популяціям
лейкоцитів. Так, додавання до середовища культивування Рах5
–
/
–
-про-В-клітин in vitro М-КСФ
(замість ІЛ-7) призводить до утворення повноцінних макрофагів, ГМ-КСФ – ДК, Г-КСФ –
гранулоцитів, ІЛ-2 — НК-клітин. Після внесення таких про-В-клітин у тимус вони дають початок Т-
лімфоцитам. Цікаво, що у мишей дикого типу про-В-клітини здатні диференціюватися лише у
напрямі В-лімфоцитів.
У Рах5
–
/
–
-про-В-клітин залишаються активованими кілька генів, які є характерними для інших
клітинних ліній, зокрема гени рецепторів до різних гемопоетичних факторів. Отже, Рах5 необхідний
не лише для подальшого розвитку про-В-клітин, а й для блокування інших шляхів розвитку
попередника В-клітин, тобто для репресії генів, які специфічно експресуються іншими типами
лейкоцитів. Зокрема, молекулярними методами було підтверджено, що Рах5 дійсно репресує гени
рецептора до гемопоетичного фактора М-КСФ. Ймовірно, функцію репресії певних генів фактор
Рах5 виконує не сам, а разом з іншими корепресорами, наприклад корепресорами родини Groucho.
Що індукує активацію фактора Рах5 у попередниках В-клітин, залишається невідомим. Нині
припускають, що експресія фактора Рах5 активується в клітинах спонтанно, але з невеликою
ймовірністю, і саме такі клітини диференціюються на В-лімфоцити. Отже, визначення процесів
розвитку СКК у В-лімфоцитарному напрямі можна розглядати як стохастичний процес, який
відбувається автономно на рівні кожної клітини.
Транскрипційні фактори, що регулюють розвиток Т-клітин. На відміну від В-клітин
транскрипційні фактори, що регулюють розвиток Т-клітин, менш вивчені. З'ясовано, що
найважливішим сигналом для комітування СКК у бік Т-клітинного ряду є сиґнал від рецептора
Notch 1, який ці клітини отримують саме в тимусі.
Рецептори родини Notch належать до родини рецепторів, які можна розглядати як попередники
транскрипційних факторів. Після зв'язування ліганду позаклітинною частиною рецептора його
внутрішньоклітинна частина протеолітично відщеплюється та мігрує в ядро, де виконує функцію
фактора транскрипції після з'єднання з іншими ядерними факторами, наприклад CBF1/RBP-jκ.
У тимусі експресується ліганд Jagged2 для рецептора Notch1 тимоцитів. Саме взаємодія Jagged2 з
Notch1 зумовлює комітування СКК до розвитку на Т-лімфоцити. Сиґналінг від Notchl активує
експресію генів Hes (Hairy enhancer of split), продукти яких репресують активність транскрипційних
факторів Е2А і EBF (переважно E2A), необхідних для розвитку В-клітин. Якщо в СКК зруйнувати
гени Notch1, розвиток тимоцитів закінчується на стадії попередників CD44
+
CD25
–
(елімінація
експресії Notch на стадії CD25
+
CD44
–
розвитку тимоцитів не впливає на утворення і позитивний
відбір CD4
+
CD8
+
клітин). Крім того, в тимусі накопичуються В-клітини, розвиток яких у цьому
органі за нормальних умов інгібується внаслідок сиґналінгу від Notch1.
Якщо в клітини кісткового мозку внести вектор, що експресує лише цитоплазматичний домен
рецептора Notch1, тобто власне ділянку з активністю транскрипційного фактора, то розвиток Т-
клітин може відбуватися навіть у кістковому мозку незалежно від тимуса.
Показано, що сигнал від рецептора Notch1 в основному сприяє утворенню Тαβ та СD8Т-клітин і
незначною мірою — Тγδ та СD4Т-клітин. З цього випливає, що для стимуляції розвитку γδТ- і CD4
Т-клітин мають бути додаткові, поки що невідомі транскрипційні фактори.
Отже, виходячи з сучасних уявлень про механізми активації транскрипційних факторів, які
контролюють розвиток Т- і В-лімфоцитів, можна зробити висновок, що диференціювання
лімфоїдних попередників на В-клітини відбувається «за дефолтом», а на Т-клітини — під дією
«інструктивних сигналів» тимуса.
На ранньому етапі В-лімфопоезу два фактори — Е2А і EBF разом активують генетичну програму,
необхідну для розвитку В-клітин. Фактор Рах5 репресує випадкову експресію інших генів, що
остаточно комітує попередників для розвитку на В-клітини. Вважають, що активація фактора Рах5
відбувається стохастично з певною ймовірністю у клітинах кісткового мозку, а отже, розвиток В-
клітин детермінований лише внутрішніми процесами, що відбуваються у клітині, тобто не залежить
від зовнішніх впливів. На відміну від В-клітин розвиток Т-клітин контролюється інструктивними
сигналами, які клітина-попередник отримує від тимічного оточення через рецептор Notch1.
12.5. МІГРАЦІЯ ЛІМФОЦИТІВ У ПЕРИФЕРИЧНІ ЛІМФОЇДНІ ОРГАНИ І ТКАНИНИ.
Після дозрівання в центральних лімфоїдних органах лімфоцити виходять із них і заселяють
периферичні. Перед виходом із кісткового мозку і тимуса Т- та В-лімфоцити починають
експресувати молекули, необхідні для здійснення міжклітинних взаємодій і проникнення у
периферичні органи й тканини. Серед них молекула хомінгу — селектин L, що експресується у
великій кількості на поверхні Т- і В-лімфоцитів, а також рецептори для хемокінів. Крім того,
лімфоцити зазнають інших змін, що є важливими для їх виживання і функціонування на периферії.
Наприклад, на Т-лімфоцитах модифікуються вуглеводні залишки у складі мембранних
глікопротеїнів, які можуть розпізнаватися пектиновими рецепторами макрофагів. На зрілих

56
монопозитивних тимоцитах кінцеві цукри блокуються залишками сіалової кислоти (завдяки
активації сіалілтрансферази), що забезпечує захист Т-клітин від фагоцитів після еміграції з тимуса.
12.5.1. Еміграція лімфоцитів із центральних лімфоїдних органів і заселення периферії.
З тимуса першими виходять γδТ-клітини, які утворюються ще в ембріогенезі. Через 14 — 15 діб (у
мишей) ембріонального розвитку тимус залишають Vγ3 Т-клітини, а незабаром і Vγ4 Т-клітини, які
мігрують відповідно в епідерміс шкіри та у слизові язика і репродуктивних органів самок. Популяції
цих Т-клітин самопідтримуються місцево впродовж усього життя мишей. Інші Т-клітини — Vγ2 і
Vγ1 а також Vγ9, що становлять більшість периферичних Тγδ, емігрують із тимуса переважно
відразу після народження — до 13-ї доби постнатального розвитку.
У дорослих мишей Тγδ, що утворюються в тимусі, становлять менш як 1 % усіх тимоцитів. Однак
на периферії їх вміст значно більший (10 — 15% усіх Т-лімфоцитів), що пояснюється
самопідтриманням у слизових та шкірі популяції Т-клітин, які виселилися на периферію в
ембріогенезі. Крім того, Vγ4δ4 Т-клітини взагалі розвиваються поза тимусом у кишках або печінці з
кістковомозкових попередників. Більшість (99 %) Т-лімфоцитів, що утворилися в тимусі,
становлять Тαβ. Еміграція Тαβ з Тимура починається через 6 діб після народження. За добу з тимуса
емігрує 1·10
6
— 2·10
6
Т-клітин (1 % загальної кількості тимоцитів).
Серед В-клітин на периферії першими з'являються B1-клітини, що несуть маркер CD5. У людини
ранні В1-клітини — «іммігранти» — виявляються в селезінці та лімфовузлах на 17-му тижні
внутрішньоутробного розвитку плоду. В дорослому організмі основним джерелом В1-клітин є
сальник і перитонеальна порожнина, де популяція їхніх попередників самопідтримується впродовж
усього життя. Частка В1-клітин становить 20 % загальної кількості В-лімфоцитів у дорослому
організмі. В постнатальному періоді в кістковому мозку утворюються і виселяються з нього лише
В2-клітини, що становлять більшість периферичних В-лімфоцитів. За добу з кісткового мозку
мишей емігрує 1,5-5·10
7
В2-клітин.
Вийшовши з кісткового мозку і тимуса, лімфоцити з венозною кров'ю рухаються до серця, де
переходять в артеріальну кров і з нею потрапляють до периферичних лімфоїдних органів і тканин.
Залежно від характеру лімфоїдних утворень та їх переважної участі у тому чи іншому виді імунної
відповіді в них вибірково мігрують певні популяції та субпопуляції лімфоцитів. Так, лімфатичні
вузли, що більше спеціалізуються на розвитку реакцій клітинного типу, заселяють переважно Т-
лімфоцити (70%), серед яких приблизно 40% становлять СD4Т-клітини і близько 30% CD8Т-
клітини. В органах, основним призначенням яких є продукування антитіл (селезінка, пейєрові
бляшки), Т-лімфоцити становлять не більше 30 %.
У периферичних лімфоїдних органах Т- і В-лімфоцити локалізуються у спеціалізованих ділянках,
які відповідно називають
тимусзалежними (Т-клітинними) та
тимуснезалежними (В-клітинними)
зонами (далі Т- і В-зони). В селезінці Т-
зонами є періартеріолярні муфти, а в
лімфовузлах — паракортикальна ділянка.
В-зонами в селезінці є біла пульпа, а в
лімфовузлах — кортикальна ділянка. В
тимуснезалежних ділянках В-лімфоцити
утворюють фолікули.
Проникнення лімфоцитів з кров'яного
русла в органи чи тканини (екстравазація)
пов'язане з подоланням ними
ендотеліального бар'єра, а міграція в Т- і
В-зони — з переміщенням усередині
тканини (хемотаксис). Процес
екстравазації лімфоцитів детально
висвітлено в розд. 1. Розглянемо основні
події посттрансендотеліальної міграції
лімфоцитів та роль хемокінів і
позаклітинного матриксу в цьому процесі.
Нагадаємо, що лімфоцит проходить чотири послідовні стадії під час затримки в кровоносній судині
та проникненні через неї: ролінг, активація, адгезія, діапедез (мал. 91).
Ролінг реалізується за допомогою лімфоцитарних селектинів (CD62L та ін.) та їхніх лігандів на
ендотелії лімфовузлів (CD34, GlyCAM-1) і слизових оболонок (MadCAM-І). Активацію інтегринів
на поверхні лімфоцита (α
L
β
2
під час міграції у лімфовузли та α4β7 і α
L
β
2
– у пейєрові бляшки)
опосередковують експресовані на ендотелії хемокіни. Адгезію зумовлюють активовані інтегрини,
зв'язуючись з їхніми лігандами на ВЕВ лімфовузлів (ІСАМ-1, ІСАМ-2) та слизових оболонок
(MadCAM-І). У діапедезі беруть участь експресовані на лімфоциті молекули CD31, які також
забезпечують контакт між клітинами ендотелію.
Після проникнення через ВЕВ у лімфовузлах і пейєрових бляшках або потрапляння з маргінальної
зони в білу пульпу селезінки лімфоцити мігрують у дискретні ділянки лімфоїдної паренхіми — Т- і
В-зони.
Зауважимо, що В-лімфоцити, як буде
зазначено далі, набувають здатності
рециркулювати і заселяти В-зони
периферичних лімфоїдних органів
лише після завершення дозрівання у
селезінці, куди вони мігрують з
кісткового мозку не повністю зрілими.
Розглянемо процес міграції
лімфоцитів у Т- і В-клітинні зони на
прикладі заселення лімфатичних
вузлів (мал. 92).
Слід зазначити, що Т- і В-лімфоцити
спочатку мігрують до Т-зон
лімфовузлів. Більшість Т-лімфоцитів
залишається у Т-зонах і лише
незначна частка їх мігрує
безпосередньо до В-зон. Серед
останніх CD4Т-клітини чисельно
переважають над CD8T-клітинами.
Більшість В-лімфоцитів залишають Т-
зони, однак подальша доля їх
залежить від активації у Т-зонах.
Якщо В-лімфоцити активуються в Т-
зонах, вони мігрують до В-зон, де
утворюють зародкові центри.
Неактивовані в Т-зонах В-лімфоцити
також мігрують до В-зон, але не
залишаються там, а проходять через
Т-зони та медулу і виходять із
лімфовузла через еферентні
лімфатичні судини. Аналогічно Т-
лімфоцити, що залишилися в Т-зонах, але не були активовані, з часом мігрують через медулу з
лімфовузла в еферентну лімфу. Отже, більшість наївних Т-клітин проходить усередині лімфовузлів
значно коротший шлях, ніж наївні В-клітини.
Механізми, що зумовлюють різні шляхи міграції лімфоцитів через Т- і В-компартменти лімфовузлів
та інших лімфоїдних органів, поки що остаточно не з'ясовані. Очевидно, що в цьому процесі значну
роль відіграють різні фактори, серед яких важливими є склад позаклітинного матриксу та хемокіни.
12.5.2. Роль хемокінів у міграції лімфоцитів.
Хомінг як Т-, так і В-лімфоцитів до вторинних лімфоїдних органів (лімфовузлів та пейєрових
бляшок) реалізується, як вважають, за участю одних і тих самих молекул адгезії, але різних
хемокінів. За залучення наївних Т-клітин до лімфовузлів, пейєрових бляшок та селезінки відповідає
хемокін, який називають хемокіном вторинних лімфоїдних органів – CCL21 (SLC). CCL21
експресується на ВЕВ вторинних лімфоїдних органів і взаємодіє з експресованим на Т-клітинах
рецептором CCR7. Крім наївних Т-клітин SLC залучає субпопуляцію центральних Т-клітин пам'яті
та ДК, що експресують CCR7. У CCR7-дефектних мишей значно знижена кількість наївних Т-
клітин у вторинних лімфоїдних органах і спостерігаються структурні аномалії Т-зон. Порушення
міграції наївних Т-клітин до лімфоїдних органів характерно також для мишей з дефектами
продукування SLC та іншого ліганда CCR7 — CCL19 (ELC — ліганд рецептора, індукованого
вірусом Епштейн-Барр). SLC та ELC продукуються локалізованими в Т-зонах зрілими дендритними
та стромальними клітинами. Вважають, що обидва хемокіни забезпечують близьку локалізацію
CCR7
+
Т-клітин, що мігрували до Т-зон, з АПК, сприяючи їх взаємодії та ініціюванню імунної
відповіді. Хемокіновий рецептор CCR7 експресується також на деяких В-клітинах, хоча і в меншій
кількості, ніж на Т-клітинах. Ці В-клітини можуть заселяти Т-зони.
Міграцію В-лімфоцитів до вторинних лімфоїдних органів та локалізацію в них визначає хемокін
CXCL13 (ВСА-1 — хемокін 1, що активує В клітини; у мишей називається BLC), який взаємодіє з
рецептором CXCR5 В-клітин. CXCL13 експресується на ВЕВ фолікулярних судин та судин
фолікулярної мантії і відповідає за формування фолікулів у селезінці, лімфовузлах та пейєрових
бляшках. Крім В-клітин CXCR5 експресує також мінорна субпопуляція центральних CD4Т-клітин
пам'яті, що несуть і рецептор CCR7. Ці CXCR5
+
Т-клітини локалізуються у фолікулярних ділянках і
допомагають in vitro В-клітинам у синтезі антитіл. Тому їх називають фолікулярними В-хелперними
Т-клітинами (Тфх). Взаємодії цих Т- і В-клітин, можливо, можуть відбуватися у фолікулярних
ділянках, куди вони залучаються ВСА-1, однак експресія Т-клітинами CXCR5 не є постійною.
Крім систем SLC - CCR7 (або ELC - CCR7) і ВСА-1 - CXCR5, у регуляції заселення вторинних
лімфоїдних органів лімфоцитами беруть участь також інші хемокіни та їхні рецептори. Так,
СХCL12 (SDF-1, фактор 1, похідний від стромальних клітин) — хемоатрактант В-лімфоцитів,
залучає до вторинних лімфоїдних органів периферичні наївні В- і В-клітини пам'яті, але не В-
клітини зародкових центрів, що, можливо, пов'язано зі зниженою експресією на останніх його
рецептора — CXCR4. Інший хемокін CCL18 (DC-CK1 — хемокін 1 дендритних клітин)
продукується ІДК та ФДК (локалізованими відповідно в Т- і В-зонах) та впливає на наївні Т- і Т-
клітини пам'яті, що були залучені хемокіном SLC до міграції через ВЕВ у лімфоїдні органи.
Припускають, що продукований ІДК DC-CK1, так само, як і ELC, залучає наївні Т- і Т-клітини
пам'яті у Т-зони. DC-CK1, що продукується ФДК, скеровує міграцію Т-клітин до зародкових
центрів, де він, очевидно, підтримує активацію Т-клітин пам'яті та стимулює взаємодію Т- і В-
клітин.
Роль хемокінів у регуляції руху лімфоцитів у нелімфоїдних тканинах, на відміну від лімфоїдних,
вивчено недостатньо. Отримано дані про контроль хемокінами хомінгу Т-клітин пам'яті до шкіри та
кишок. Міграцію CLA
+
Т-клітин пам'яті до шкіри визначає хемокіновий рецептор CCR10, що
експресується переважно на цих клітинах, і його ліганд CCL27 (СТАСК — шкірний хемокін, що
атрактує Т-клітини). У регуляції хомінгу CLA
+
Т-клітин задіяні також експресований на їхній
поверхні CCR4 та наявний на ендотелії кровоносних судин нормальної та запаленої шкіри його
ліганд CCL17 (TARC — хемокін, регульований активацією та тимусом). Хомінг α4β7
+
Т-клітин
пам'яті до кишок контролює хемокін CCL25 (ТЕСК — хемокін, еспресований тимусом), що
взаємодіє з рецептором CCR9 Т-клітин.
Отже, хемокіни регулюють трансендотеліальну міграцію лімфоцитів та їхнє переміщення в тканині
лімфоїдних органів. Експресовані на ендотеліоцитах хемокіни зумовлюють активацію інтегринів та
затримують ті лімфоцити, що несуть відповідні хемокінові рецептори. Після екстравазації
лімфоцитів певні хемокіни в тканинах спрямовують їхній рух у позаклітинному матриксі. Чутливі
до дії певних хемокінів лімфоцити завдяки реорганізації цитоскелета та адгезивним взаємодіям з
позаклітинним матриксом рухаються за градієнтом концентрації цих хемокінів.
12.5.3. Роль позаклітинного матриксу в міграції лімфоцитів.
Після екстравазації лімфоцити потрапляють з двовимірного простору на ендотелії судин у
тривимірний — у міжклітинне середовище, так званий позаклітинний матрикс (ПКМ). Основним
компонентом ПКМ є волокна колагену різних типів, поперечно зшиті з іншими компонентами,
такими як фібронектин, ламінін, гіалуронан, які здатні адсорбувати хемокіни та інші цитокіни і
містять ліганди, з якими зв'язуються рецептори лімфоцитів.
У лімфоїдних органах ПКМ формує стабільну високоорганізовану тяжеподібну сітку з порами
діаметром 3—15 мкм, що нагадують каналоподібні структури. Ці структури зорієнтовані в
лімфовузлах у бік фолікулярних В-клітинних зон, а в пейєрових бляшках — у напрямі склепіння
фолікула (в зону презентації антигену) і є провідними шляхами міграції лімфоцитів. Подібні
структури виявлено і в шкірі.
Міграція лімфоцитів (як і інших типів лейкоцитів — нейтрофілів, моноцитів) через ПКМ, так само,
як і переміщення через поверхню судинного ендотелію, здійснюється за принципами амебоїдного
руху і пов'язана з поляризацією клітин. Поляризація індукується градієнтами хемоатрактантів,
розчинними та іммобілізованими на ПКМ, або певними його лігандами і скеровується в напрямі
градієнта. В результаті поляризації відбувається активація і керований цитоскелетом перерозподіл
поверхневих рецепторів, передусім хемокінових, та молекул адгезії до різних компартментів
поляризованої клітини. Докладніше вивчено міграцію в ПКМ Т-лімфоцитів.
Для дослідження молекулярних механізмів міграції Т-лімфоцитів у тканинах використовують
штучно створені дво- і тривимірні моделі з фізіологічних компонентів ПКМ як з волокон колагену в
чистому вигляді, так і з вмістом фібронектину, ламініну тощо. Перерозподіл поверхневих
рецепторів реєструють міченням їх антитілами, забарвленими флуоресцентними барвниками, а роль
окремих молекул у міграційному процесі — блокуванням їх антитілами.
У штучному тривимірному матриксі Т-клітини в процесі переходу від нерухомого до міграційного
стану змінюють свою форму і набувають вигляду завислої краплі з лідерним кінцем, основною
частиною (тілом) клітини округлої форми та замикальним кінцем — уроподом. На лідерному кінці
поляризованої Т-клітини акумулюються кіназа фокальних контактів, рецептори клітинної адгезії
LFA-1 і CD45RO, а в активованих клітинах і хемокінові рецептори — CCR2, здатний зв'язувати
CCL2, 7, 13 (відповідно МРС-1, -3, -4), та CCR5, лігандами якого є CCL4 і CCL5 (відповідно МІР-1β
і RANTES). Цей компартмент забезпечує сприйняття хемотаксичних сигналів, реорганізацію
цитоскелета та утворення точкових контактів у місці взаємодії клітини з субстратом. В основному
тілі клітини містяться організаційний центр мікротрубочок та протеїнкіназа С, відповідальна за
ініціювання хемокініндукованої міграції. Уропод формується цитоплазматичними виростами,
містить численні елементи цитоскелета й молекули адгезії: β1-інтегрини, ІСАМ-1, -З, CD44, CD43
(антиадгезійна молекула), можлива тимчасова експресія LFA-1 і CD45. Уропод є місцем взаємодії
поверхневих рецепторів клітини з цитоскелетом і виконує адгезивну та якірну функції під час
міграції.
Поляризація забезпечує можливість послідовної та поетапної взаємодії певних поверхневих
структур лімфоцитів із субстратом (ПКМ), що зумовлює циклічне прикріплення і відкріплення
клітин та сприяє їх переміщенню. Хоча точні механізми міграції лімфоцитів у ПКМ ще остаточно не
з'ясовані, проте отримані експериментальні дані свідчать про важливу роль у цьому процесі самого
матриксу.
Як виявилося, умови міграції Т-клітин через штучно створений тривимірний матрикс відрізняються
від таких через двовимірний субстрат. У двовимірних моделях, у яких взаємодія клітини з плоским
субстратом обмежується лише базальним боком, неактивовані Т- лімфоцити не здатні рухатися,
хоча інтенсивно змінюють форму здійснюючи «біг на місці». У разі внесення Т-лімфоцитів у
тривимірне штучне середовище, в якому клітини з усіх боків оточені матриксом, вони можуть
активно мігрувати навіть за відсутності активації β1-інтегринів завдяки численним точковим та
ланцюговим контактам із субстратом.
Міграція Т-клітин у тривимірному оточенні залежить як від розміру, ригідності та функціонального
стану клітини, експресії нею різних інтегринів, так і від властивостей ПКМ і може бути як
інтегриннезалежною (базовою), так і інтегринзалежною. Серед інтегринів провідну роль як
рецепторів адгезії відіграють інтегрини β1-родини. Зокрема, інтегрини α
1
β
1
, α
2
β
1
, α
3
β
1
, зв'язуються
переважно з колагеном, α
4
β
1
тa α
5
β
1
— з фібронектином, α
6
β
1
— з ламініном, a α
v
β
3
здатний
взаємодіяти з багатьма компонентами ПКМ. Крім інтегринів у цьому процесі беруть участь члени
родин імуноглобулінів, наприклад LFA-1/ІСАМ-1, та кадхеринів. Важливим індуктором β1-
інтегринзалежної міграції є хемокіни CCL4 і CCL5. Хемокіни можуть ініціювати міграцію Т-клітин,
впливаючи на сигнальну систему активації цитоскелета й посилюючи адгезивні контакти з
субстратом.
Швидкість міграції Т-лімфоцитів залежить від сили адгезії до субстрату, яка визначається
щільністю (і афінністю) рецепторів на клітині та їхніх лігандів на субстраті. Оптимальна швидкість
спостерігається за середньої та низької сили адгезії, оскільки саме така адгезія забезпечує легке
прикріплення лідерного та відкріплення замикального кінця клітини. Слід зазначити, що у
неактивованих та більшості субпопуляцій активованих Т-лімфоцитів рецептори зв'язування
інтегринів мають низьку афінність, що зумовлює низьку силу адгезії клітин до більшості субстратів.
Сигнали, що індукують інтегриннезалежну поляризацію та міграцію Т-клітин, виникають при
перехресному зшиванні α4-інтегринів, LFA-1, ІСАМ-3 або CD43, активації протеїнкінази С та
кінази фокальних контактів. При цьому інтегрини індукують міграцію Т-клітин через
фосфорилювання за тирозином кінази фокальних контактів, а хемокінові рецептори — через G-
білок та протеїнкіназу С.
За опосередкованими рецепторами механізмами відбувається міграція Т-клітин у хронічно
запалених тканинах, де у високих концентраціях містяться хемокіни та інші цитокіни й достатньо
адгезивні ділянки. Водночас показано, що Т-клітини можуть мігрувати у тривимірному матриксі й
за відсутності лігандспецифічних зв'язувань завдяки зміні форми клітини, утворенню філоподій та
виростів мембрани, контактів за участю заряджених залишків вуглеводів, гідрофобних зв'язків.
Такий спосіб міграції характерний для неактивованих Т-клітин у ретикулярних тканинах за
відсутності запалення та всередині фіброзної сітки лімфатичних органів.
Таким чином, ПКМ відіграє важливу роль у міграції Т-лімфоцитів і здійснює контактну регуляцію
цього процесу як за наявності, так і за відсутності хемотаксичних градієнтів.
Дозрівання лімфоцитів на периферії. У периферичних лімфоїдних органах лімфоцити, передусім
популяції В, зазнають подальшого розвитку, оскільки вони виходять із центральних лімфоїдних
органів неповністю зрілими. Незрілі В-клітини після стадій дозрівання в кістковому мозку мігрують
до периферичних лімфоїдних органів, здебільшого до селезінки. В селезінці В-клітини проходять
останні стадії дозрівання, після чого надходять для рециркуляції і заселяють В-зони вторинних
лімфоїдних утворів. Однак не зовсім зрозуміло, як саме відбувається дозрівання В-лімфоцитів у
селезінці. Вважають, що в селезінці незрілі В-клітини затримуються близько чотирьох діб. За цей
час має відбутися толяризація В-клітин «на периферії». Якщо незрілі В-клітини зустрічають
антиген, до якого специфічні їхні рецептори, вони не активуються, а гинуть. Наслідком цього є
толяризація В-лімфоцитів до аутоантигенів, які не містилися в кістковому мозку, але були достатньо
поширені в організмі. Після дозрівання в селезінці з незрілих В-клітин утворюються наївні В-
лімфоцити, які мають досить тривалий термін життя — близько 15 тижнів. За цей час вони
рециркулюють в організмі й «очікують» зустрічі з чужорідним антигеном. На відміну від незрілих
В-лімфоцитів наївні В-клітини експресують на своїй мембрані не лише IgM-, а й IgD-рецептор.
Функціональне значення експресії IgD-рецептора на наївних В-клітинах залишається невідомим,
оскільки миші з нокаутом гена Сδ не зазнають ніяких порушень у розвитку та активації В-клітин.
У периферичних лімфоїдних органах завершується також процес негативного відбору (периферична
толяризація) Т-клітин та їх функціональне дозрівання. Зауважимо, що Т-лімфоцити виходять із
тимуса у вигляді наївних Т-клітин, які є попередниками ЦТЛ і Т-хелперів. Важливу роль у
дозріванні Т-клітин на периферії відіграють секретовані тимічним мікрооточенням гормони, що
циркулюють у крові. Гормони тимуса сприяють підвищенню функціонального потенціалу Т-клітин
завдяки посиленню їхньої цитокінпродукувальної здатності. Зокрема, показано, що зрілі
периферичні Т-клітини є більш ефективними продуцентами ІЛ-2, ніж клітини заключної стадії
диференціювання в тимусі — медулярні тимоцити.
У цьому відношенні на особливу увагу заслуговує гіпотеза про існування механізму підтримання
життєздатності та проліферації Т-клітин на периферії. Допускається, що так само, як під час
позитивного відбору тимоцитів у тимусі, сигнали від комплексів своїх пептидів з власним МНС на
АПК підтримують життєздатність і навіть проліферацію Т-клітин на периферії. Ця проліферація
підтримується на низькому рівні і, як і в тимусі, не пов'язана з синтезом і секрецією цитокінів (та
розвитком імунореактивності).
Наївні Т- і В-лімфоцити — це повністю зрілі клітини, які вже пройшли контроль на безпечність для
організму, але ще не зустрілися з антигеном. Саме на цьому етапі стає можливим контакт Т- і В-
клітин з чужорідним антигеном і залучення антигенспецифічних клітин до імунної відповіді. Після
зустрічі з антигеном відбувається остання стадія диференціювання Т- і В-клітин, яку називають
постактиваційним диференціюванням. При цьому В-клітини також проходять низку важливих
етапів розвитку, під час яких можуть відбуватися як процеси зміни афінності їхніх рецепторів, так і
процеси додаткової селекції В-клітин з найбільш афінними рецепторами. Останні етапи є
характерними виключно для розвитку В-клітин і не мають аналогів у розвитку Т-клітин (див. розд.
11).
ВИСНОВКИ.
Лімфоцити – головні клітини імунної системи – утворюються з гемопоетичних стовбурових клітин.
Т-лімфоцити утворюються в тимусі, В-лімфоцити – в кістковому мозку (у птахів – у фабрицієвій
сумці). Під час роз0витку Т- і В-лімфоцитів зазнають перевірки на правильність будови
антигензв'язувальних рецепторів. У тимусі підтримуються для подальшого розвитку Т-лімфоцити,
які здатні впізнавати антигенні пептиди в асоціації з власними молекулами МНС (позитивний відбір
за МНС-рестрикцією), і елімінуються Т-лімфоцити, що потенційно здатні виявляти реактивність
проти комплексів власних МНС-пептид (негативний відбір на аутотолерантність). В-лімфоцити
зазнають позитивного і негативного відбору в кістковому мозку, але ліганд, який вони розпізнають
під час позитивного відбору; залишається нез'ясованим. У результаті двохетапного відбору
формується репертуар Т- і В-рецепторів, що спроможні розпізнати всі можливі антигени, з якими
може контактувати організм. Відібрані лімфоцити залучаються до проліферації й утворюють клони
(нащадки однієї клітини), які становлять популяції Т- і В-лімфоцитів.
Сформовані клони Т- і В-лімфоцитів виходять із центральних лімфоїдних органів та мігрують до
периферичних органів і тканин, де поселяються в дискретних спеціалізованих ділянках — Т- і В-
клітинних зонах. Міграція пов'язана з проходженням через ендотелій (екстравазація) та
переміщенням у тканині (екотаксис). Основою міграції є взаємодія молекул адгезії лімфоцита з
їхніми лігандами на ендотелії та позаклітинному матриксі. Міграція лімфоцитів регулюється
системою хемокінів, які, діючи як хемоатрактанти, спрямовують рух лімфоцитів. Мікрооточення Т-
і В-клітинних зон створює умови для активації лімфоцитів та кооперації Т- і В-клітин під час
розвитку імунної відповіді на антиген, що надходить у лімфоїдну систему.
Контрольні запитання.
1. Яке біологічне значення має постійне оновлення лімфоїдних клітин в організмі?
2. Якими властивостями характеризуються стовбурові клітини, яка їх роль в онтогенезі та які
існують методи їх аналізу?
3. Як відбувається рециркуляція стовбурових клітин і яка її роль?
4. Схарактеризуйте етапи диференціювання Т- і В-лімфоцитів, простежте появу та зміни експресії їх
основних функціональних маркерів, розділення на субпопуляції.
5. Які зміни генетичного апарату відбуваються в процесі розвитку Т- і В-клітин?
6. Які аналогії та відмінності у стадіях розвитку Т- і В-лімфоцитів?
7. Які транскрипційні фактори визначають напрям диференціювання клітин імунної системи?
8. Які клітини формують мікрооточення Т- і В-лімфоцитів у різних компартментах тимуса і
кісткового мозку та яка їхня роль у дозріванні лімфоцитів?
9. Що таке позитивний та негативний відбір Т- і В-клітин і як він здійснюється?
10. Яке значення мають генноінженерні методи для вивчення процесів відбору лімфоцитів?
11. Як здійснюється розселення Т- і В-лімфоцитів у периферичні лімфоїдні органи та які
молекулярні механізми міграції їх у спеціалізовані ділянки — Т- і В-клітинні зони?
РОЗДІЛ 13. ІМУННА ТОЛЕРАНТНІСТЬ.
Лімфоїдна система вищих тварин потенційно здатна здійснювати імунну відповідь проти безмежної
кількості чужорідних антигенів та відторгати тканини будь-яких генетичне чужорідних донорів.
Однак власні тканини вона не відторгає і в нормі імунних реакцій проти власних антигенів не
здійснює. Імунна система відрізняє «своє» від «не свого», тобто толерантна до власних речовин, але
відторгає речовини всіх інших особин як свого, так і інших біологічних видів. Відсутність
57
реактивності щодо власних речовин називають природною імунною толерантністю. Слово
«толерантність» (від лат. tolerantis — терплячий) означає «терпимість». Природна толерантність
відіграє важливу роль у збереженні антигенної індивідуальності особини. Крім природної
розрізняють набуту, або індуковану, імунну толерантність. Іноді набуту толерантність називають
імунним паралічем. Отже, природна толерантність (аутотолерантність, self-tolerance) — це
толерантність до своїх антигенів, а індукована толерантність — це толерантність до чужорідних
(non-self-tolerance) антигенів. Толерантність можна відтворити в експерименті. Тому такий тип
індукованої толерантності називають штучною толерантністю.
Значна кількість хвороб пов'язана як із втратою природної толерантності (різні аутоімунні
захворювання), так і з небажаним набуттям толерантності до інфекційних агентів або пухлин,
внаслідок чого імунна система стає неспроможною ефективно протистояти інфекції або злоякісному
захворюванню. Крім того, розкриття механізмів природної толерантності дало б змогу розробити
підходи для тривалого приживлення трансплантатів, що зумовило б значний прогрес у
трансплантаційній медицині. Однак наука нині ще не здатна з впевненістю відповісти на більшість
питань стосовно молекулярних механізмів, що лежать в основі формування природної
толерантності. Значна кількість експериментальних підходів для індукування штучної
толерантності та її перенесення іншим тваринам дали змогу детально описати різні її прояви, однак
невідомо, наскільки ці експериментальні моделі відображають механізми, що лежать в основі
формування природної толерантності.
Вважають, що природна толерантність, як і штучна, індукована антигенами, але не чужорідними, а
власними, з якими лімфоїдна система контактує під час індивідуального розвитку та утворення
нових лімфоцитів у первинних лімфоїдних органах. Тому, за визначенням експертів ВООЗ,
толерантність є специфічною імунною ареактивністю, що індукована антигеном. У межах цього
визначення толерантність є протилежністю вторинній імунній відповіді.
Антигени, що індукують толерантність, називають толерогенами. Оскільки толерантність може
мати різні ступені прояву, то її можна визначити як індуковану антигеном втрату або зниження
здатності лімфоїдної системи здійснювати специфічну імунну відповідь проти цього самого
антигену. Імунну толерантність слід відрізняти від генетично детермінованої неспроможності
відповідати на певні антигени внаслідок відсутності у цих антигенів імуногенних властивостей,
відсутності необхідних алелів МНС чи репертуару рецепторів. Толерантність потрібно також
відрізняти від інших форм пригнічення імунної відповіді — імунодефіцитних станів та станів
імунної депресії. На відміну від останніх імунна толерантність завжди специфічна, тобто
відсутність відповіді на певний антиген (толероген) не позбавляє організм здатності нормально
відповідати на введення інших антигенів. Отже, імунна толерантність є фізіологічною функцією
імунної системи, а імунодефіцити — патологією.
Природна імунна толерантність — регуляторний механізм імунної системи, що ґрунтується на
елімінації або інактивації лімфоїдних клітин, специфічних до антигенів певного організму. Цей
механізм запобігає розвитку аутоімунних реакцій.
На існування механізмів, які б запобігали утворенню антитіл проти власних антигенів організму,
звертали увагу ще П. Ерліх та інші першовідкривачі антитіл. У 1938 р. Трауб, перещеплюючи вірус
лімфоцитарного хоріоменінгіту на курячих ембріонах, помітив, що птахи, які розвиваються із
заражених ембріонів, не розвивають імунну відповідь до цього вірусу, тоді як інтактні кури стійкі
до спричиненої ним інфекції. В 1945 р. Р. Оуен установив, що в крові дизиготних телят-близнюків,
які в ембріональному періоді мали спільний плацентарний кровотік, містяться еритроцити, що
належать до двох різних груп крові. Крім того, такі телята легко переносили переливання крові від
одного до іншого. Ці спостереження наводили на думку, що формування природної толерантності
імунної системи до всіх власних антигенів відбувається в ембріональний період. У 1949 р. Ф. Бернет
і ф. Феннер висловили припущення, що толерантність до власних антигенів не є генетично
детермінованою або природженою, а формується в період дозрівання імунної системи внаслідок
контакту незрілих лімфоцитів із власними антигенами, які потім у постембріональний період
сприймаються імунокомпетентними клітинами як «своє».
Ф. Бернет і Ф. Феннер припустили, що субстанції, які утворюються в opгaнiзмі в ембріональному
періоді, контактують із лімфоїдною системою, розпізнаються як «свої», і до них виникає імунна
толерантність, яка стає нормою після завершення дозрівання імунної системи. Вони також
висловили припущення, що чужоріді антигени, введені в організм тварини в ембріональний або
ранній постнатальний період, будуть сприйняті незрілою імунною системою як власні і до них
сформується толерантність, тобто організм навчається розпізнавати «своє» і «чуже» ще у процесі
розвитку. Однак, працюючи з антигеном, до якого толерантність формується важко (вірус грипу), Ф.
Бернет не довів цього геніального припущення.
Здатність імунної системи диференціювати «своє» і «чуже» запобігає розвитку аутоімунних реакцій
проти компонентів власних тканин. Механізм імунної толерантності перешкоджає клонам
імунокомпетентних клітин, специфічних до антигенів, які постійно містяться в організмі у високих
концентраціях, зайняти домінуюче положення у популяції лімфоїдних клітин та спричинити
нерегульовану експансію цих клонів в організмі.
Щоб перевірити припущення Ф. Бернета і Ф. Феннера, Р. Білінгхем, Л. Б. Брент і П. Медавар (1953)
провели дослідження з трансплантацією ембріонам мишей клітин алогенної кровотворної тканини.
Ці дослідники довели можливість штучного відтворення стійкої імунної толерантності до алогенних
трансплантатів.
П. Медавар зі співавторами показали, що введення алогенних лімфоїдних клітин ембріонам мишей
зумовлює зниження або повну втрату дорослими організмами, що розвинулися з таких ембріонів,
здатності вилучати антигени лінії донора (синтезувати специфічні антитіла, відторгати
трансплантат, реагувати в ЗКЛ). Це було продемонстровано приживленням чорних клаптиків шкіри
мишей лінії C57BL на білих мишах лінії
А, в яких було індуковано імунну
толерантність у пренатальний період
введенням кровотворних клітин від
мишей лінії C57BL (мал. 93). В
експериментах П. Медавара і співавторів
тривале збереження толерантності
пояснювалося тим, що тварини ставали
«імунними химерами», в яких тривалий
час персистували лімфоїдні клітини від
двох різних ліній.
У дослідах М. Гашека у дорослих птахів,
які в ембріональному періоді перебували
в стані парабіозу, трансплантати
приживалися і введені внутрішньовенно
еритроцити партнера за парабіозом не
елімінувалися. Трансплантати від інших
тварин та еритроцити інших донорів
елімінувалися у звичайні строки. Така
закономірність пояснюється тим, що
здатність розрізняти власні й чужорідні
антигени у тварин, як правило, виникає
постнатально. В ембріональному і
ранньому постнатальному періоді незріла
лімфоїдна тканина сприймає за «свої»
введені в організм чужорідні антигени.
Доказом того, що природна толерантність до власних тканиноспецифічних антигенів є набутою,
можуть бути також дані Е. Тріплета (Е. Triplett; 1962), який пересаджував гіпофізи, видалені у
пуголовків, жабам, що з них розвинулися. У цих дослідах аутологічні гіпофізи відторгалися. Це
означало, що навіть антигени, які кодуються власними генами цих жаб, сприймалися як чужорідні,
якщо під час індивідуального розвитку лімфоїдна система з ними не контактувала.
Досліди П. Медавара і співавторів стали яскравим доказом справедливості геніальної здогадки Ф.
Бернета. Це дало змогу Ф. Бернету в 1957 р. сформулювати клонально-селекційну теорію імунітету,
одним з постулатів якої був висновок про те, що аутореактивні клони елімінуються в процесі
розвитку імунної системи внаслідок контакту з власними антигенами. За свої відкриття П. Медавар і
Ф. Бернет у 1960 р. були удостоєні Нобелівської премії.
Експерименти П. Медавара та інших дослідників поклали початок широкому вивченню імунної
толерантності до чужорідних антигенів, трансплантаційного імунітету, природної толерантності та
пригнічення імунної реактивності. Нині цим питанням присвячено багато важливих
експериментальних і клінічних спостережень.
13.1. ШТУЧНА ТОЛЕРАНТНІСТЬ.
Головною експериментальною моделлю для вивчення механізмів природної толерантності є
індукування штучної толерантності у піддослідних тварин.
Толерантність до алогенних і ксеногенних клітин. Як показав П. Медавар зі співробітниками,
штучну толерантність до алогенних тканин можна створити внутрішньовенним введенням
ембріонам живих лімфоцитів. Оскільки толерантність до алогенних клітин і тканин залежить від
сильних антигенів тканинної сумісності, які містяться на лімфоцитах, створення толерантності до
лімфоцитів зумовлює толерантність щодо антигенів інших тканин того самого донора. За здатністю
індукувати стан толерантності більш активними у мишей є клітини лімфатичних вузлів, селезінки,
кісткового мозку, менш активні — лімфоцити крові й тимоцити. Найефективніше індукує
толерантність введення алогенних кровотворних клітин у кров, оскільки це забезпечує надходження
антигену в усі лімфоїдні органи. Алогенні лімфоїдні клітини донора, трансплантовані ембріонам,
приживаються в організмі реципієнта і проліферують. У цьому випадку тварина є імунною
химерою, кровотворна тканина якої складається з власних і генетично чужорідних клітин.
Толерантність залежить від кількості клітин донора, що прижилися в організмі реципієнта. Чим
більша кількість живих клітин донора виявляється у лімфоїдних тканинах реципієнта, тим
триваліше зберігається толерантність до пересадженого трансплантата. Якщо клітини донора в
організмі реципієнта не приживаються, то трансплантат відторгається.
Здатність розвивати імунну відповідь формується в ембріональний період не до всіх антигенів
одночасно. Ембріони овець після 75-ї доби внутрішньоутробного розвитку відторгають
трансплантати шкіри, що свідчить про раннє дозрівання лімфоїдної системи у цих тварин. Однак
антитіла на дифтерійний анатоксин і корпускулярні антигени черевнотифозної палички в ягнят не
утворюються і впродовж перших місяців постнатального розвитку.
Імунну толерантність у деяких видів ссавців і птахів можна індукувати не лише в ембріональному
періоді, а й у перші дні постнатального розвитку. Час, упродовж якого можна створити
толерантність, називають адаптивним періодом. У кролів адаптивний період закінчується ще до
народження, у мишей він зберігається 1 — 2 доби, а в собак, щурів і качок — від 2 до 5 діб після
народження. Адаптивний період значною мірою залежить від характеру антигенів, їхніх доз,
антигенних відмінностей між донором і реципієнтом. Щодо алогенних антигенів він триваліший,
ніж до ксеногенних. При відмінностях лише за слабким локусом гістосумісності адаптивний період
у мишей може збільшуватися до двох тижнів. У дорослих тварин сталу штучну толерантність
можна зумовити багаторазовим уведенням великих доз алогенних клітин лімфатичних вузлів і
селезінки або інших клітин, що містять ядро. Однак це можливо лише у разі незначних генетичних
відмінностей донора та реципієнта за слабкими локусами гістосумісності. Максимальна подібність
донора і реципієнта за антигенами гістосумісності — одна з основних умов створення толерантності
до алотрансплантатів у тварин.
Створити повну толерантність щодо алогенних трансплантатів можна лише впродовж адаптивного
періоду після трансплантації великої кількості живих клітин лімфоїдної й мієлоїдної тканин, що
здатні до приживання та проліферації. При введенні недостатньої кількості клітин розвивається
часткова толерантність, а при введенні ще меншої кількості — сенсибілізація. Виникнення
толерантності залежить також від кількісного співвідношення введених живих лімфоцитів або
неклітинних антигенів і лімфоцитів реципієнта. Тривалість життя алогенного трансплантата прямо
залежить від кількості введених лімфоїдних клітин.
Для створення стійкої толерантності потрібна колонізація лімфоїдної тканини реципієнта
лімфоїдними клітинами донора. Після введення зруйнованих, або незруйнованих, але вбитих
лімфоїдних клітин пригнічується антитілогенез, але відбувається відторгнення трансплантатів;
тобто виникає часткова, неповна толерантність. Цей стан зберігається впродовж коротшого періоду,
ніж повна толерантність. При трансплантації еритроцитів реактивність до інших тканин
зберігається, хоча може бути й ослаблена.
Вважають, що з введенням новонародженим тваринам алогенної лімфоїдної тканини в організм
реципієнта потрапляють алогенні стовбурові кровотворні клітини (СКК). Частина цих алогенних
СКК може заселяти тимус, де вони диференціюються не лише на тимоцити, а й на клітини
мієлоїдного ряду — дендритні клітини і макрофаги тимуса. Саме тому алогенні клітини беруть
участь у процесах негативної і позитивної селекції тимоцитів, яка відбувається в тимусі реципієнта.
Отже, для створення і тривалого підтримання механізму толерантності необхідною умовою є
формування химерного тимуса, до складу якого входять клітини двох різних генотипів. Такий
механізм пояснює також те, чому в ембріонів і новонароджених тварин толерантність індукувати
легше — саме в цей час у них відбувається заселення тимуса ембріональними СКК, які формують
мієлоїдні клітинні елементи тимуса.
Толерантність, отриману за допомогою живих кровотворних клітин, можна перенести з клітинами
селезінки сингенним тваринам у ранньому постнатальному періоді або статевозрілим тваринам
після опромінення — адоптивне перенесення. Кінетику індукування толерантності in vivo можна
вивчати методом перенесення клітин (відмитих лімфоцитів) від толерантних тварин опроміненим
або сингенним реципієнтам, яким вводять специфічний антиген.
Стійку толерантність щодо ксеногенних тканин зазвичай створити не вдається, але нестійка
толерантність між тваринами споріднених видів у деяких випадках відтворюється. Головною
перешкодою у створенні толерантності до ксеногенних тканин є участь механізмів неспецифічного
захисту у відторгненні ксеногенних тканин. Гостре відторгнення ксеногенного трансплантата
пов'язане переважно з утворенням тромбів, геморагіями, після чого настає некроз пересаджених
тканин. Причиною гострого відторгнення є атака природними антитілами реципієнта ендотелію
донора, внаслідок чого відбувається активація комплементу. Наприклад, під час трансплантації
органів свиней до вищих приматів природні антитіла приматів розпізнають епітоп Gall-3Gal, який
входить до складу деяких глікопротеїнів і гліколіпідів ендотелію свиней, але відсутній в організмі
вищих приматів. Якщо гострому відторгненню запобігти, наприклад, за допомогою молекулярно-
генетичних методів (див. далі), то можна досягти тривалішого приживання ксеногенного
трансплантата.
Поліклональна толерантність, тобто толерантність до багатьох антигенів, може бути індукована
багаторазовим введенням (100 ін'єкцій) алогенних клітин, які містять ядро, і великих доз розчинних
антигенів (гемоціанін, флагелін) упродовж тривалого часу. Це забезпечує тривале приживлення як
алогенних, так і ксеногенних трансплантатів. Таку толерантність важко розглядати як власне
толерантність, вона швидше нагадує імунодефіцитні стани.
Явище імунної толерантності, як і всі імунні реакції, є суворо специфічним. Тварина, толерантна до
тканин донора трансплантата, реагує нормальними імунними реакціями на трансплантати від того
самого виду, але іншої генетичної лінії.
Толерантність до бактеріальних і вірусних антигенів. Створити стан толерантності до
бактеріальних і вірусних антигенів в експериментальних умовах, як правило, не вдається. G. Nossal і
A. Abbot (1966) індукували стан толерантності у мишей до сальмонел уведенням у неонатальному
періоді Н-антигену. У дослідженнях J. Larsen (1969) вірус лімфоцитарного хоріоменінгіту у
новонароджених мишей зумовлював стан повної і тривалої толерантності за наявності вірусемії та
відсутності специфічних антитіл. Те, що досягти толерантності до вірусів і бактерій важко, можна,
очевидно, пояснити високою антигенністю та мозаїчністю корпускулярних мікробних антигенів,
оскільки толерантність вдається отримати до певних фракцій мікробних антигенів — інактивованих
білкових токсинів, мономерного флагеліну, певних адгезивних молекул, але не до цілих бактерій чи
вірусів.
Відсутність толерантності до більшості бактерій і вірусів пояснюється участю механізмів
природного імунітету в їх елімінації. Корпускулярні вірусні та бактеріальні антигени мають явні
ознаки чужорідності для організму і тому у відповідь на їх потрапляння до організму починають
розвиватися реакції запалення, зумовлені факторами неспецифічного захисту: активується
комплемент, антигенні субстанції розпізнаються лектиновими і Toll-like рецепторами, активуються
тканинні 6азофіли, макрофаги і дендритні клітини тошо. Саме активація дендритних клітин є
умовою запуску специфічних імунних реакцій. Тому за наявності активації ДК дуже важко
зумовити специфічну імунну толерантність. Активацією ДК можна пояснити також зрив
толерантності після введення толерогену з ад'ювантом.
За певних вад системи неспецифічного імунітету можливий розвиток толерантності і до
корпускулярних вірусних та бактеріальних антигенів.
Після введення корпускулярних мікробних антигенів новонародженим і дорослим тваринам
формується лише часткова ареактивність, тобто знижена реактивність на деякі з антигенів
складного бактеріального комплексу. Ступінь толерантності, що індукована до певної мікробної
речовини, у різних генетичних ліній мишей неоднакова.
Толерантність, індукована неклітинними антигенами. Толерогенна активність антигенних
речовин залежить від їхньої молекулярної маси й полімерності. Так, мономерний флагелін з
молекулярною масою 40 кДа виступає як Т-залежний антиген, а полімерний флагелін з
молекулярною масою 100 кДа — як Т-незалежний. Тому й механізми індукування толерантності до
цих речовин різні.
Важливою умовою толерогенності білкових молекул є їх розчинність. Агреговані й денатуровані
молекули толерантності не зумовлюють. Д. Дрессер (1962) встановив, що осадження макроаґреґатів
білка з альбуміну і γ-глобуліну сироватки після ультрацентрифугування при 20 000g призводить до
втрати білками антигенних властивостей і набуття толерогенних. Якщо осаджений під час
центрифугування білок додати до розчинного білка, то розчинний білок набуває імуногенних
властивостей. Цей ефект, який називають феноменом Дрессера, спостерігається при аналогічних
дослідах і з розчинами інших білків. Дезагреговані γ-глобуліни виявляють толерогенну дію в малих
дозах, тому їх застосовують для індукування толерантності. У дорослих мишей толерантність
виникає після внутрішньочеревного введення 10 — 20 мг дезагрегованого γ-ґлобуліну людини.
Повторні введення γ-ґлобулінів у малих дозах показали, що стан толерантності можна індукувати і
підтримувати малими дозами антигену. Однак чим вища доза розчинних білкових aнтигенів, які
вводять новонародженим тваринам, тим більш вираженою і триваліше є толерогенність.
Толерантність новонароджених мишей і кролів до алоантигенів білків сироватки зберігається 3 — 6
міс і може подовжуватися після повторних введень антигену.
Можливо, толерантність імунної системи до розчинних білкових антигенів є важливим механізмом
підтримання природної толерантності до власних антигенів плазми і лімфи. Розчинні білкові
антигени самі по собі не здатні індукувати активацію дендритних клітин, а отже, зумовити
ініціювання специфічних імунних реакцій. Крім того, мономерні білкові антигени не здатні
перехресно зв'язувати рецептори В-лімфоцитів, що є необхідною yмовою для їх інтерналізації і
презентації.
Експериментальними дослідження було показано, що введення тваринам аутологічних антигенів з
плазми крові в агрегованому стані зумовлює аутоімунні реакції і синтез антитіл проти введених
антигенів. В агрегованому стані аутоантигени можуть спричинити запалення, що може призводити
до розвитку аутоімунних реакцій. Крім того, агреговані аутоантигени можуть зумовити перехресне
зв'язування рецепторів аутоспецифічних В-клітин і їх активацію (за принципом Т-незалежних
антигенів).
Можливий ще один механізм толерогенної дії розчинних білкових антигенів – блокування
рецепторів В-клітин. Таке блокування може відбуватися як на рівні зрілих В-клітин, що веде до
ігнорування В-лімфоцитами зв'язаного антигену, так і на рівні попередників В-клітин у кістковому
мозку, що може призводити до загибелі цих клітин.
Індукування толерантності до гаптенів. Як правило, гаптени самостійно толерантність не
індукують. Толерантність до гаптенів залежить від толерантності до носія. Наприклад, поліпептиди,
які складаються з D-амінокислот, мають толерогенні властивості і якщо їх використовувати як носії,
то вони пригнічують імунну відповідь на гаптен, введений повторно навіть з імуногенним носієм.
Тварина, толерантна до носія, не продукує антитіл до гаптену, що кон'югований з цим носієм, або
дає ослаблену імунну відповідь, отже, є підстави вважати, що толерантність до кон'югату може бути
зумовлена ареактивністю до носія.
Толерантність може бути індукована до реакційноздатних простих хімічних речовин, які є
гаптенами, при введенні їх у вільному стані. Ці речовини, очевидно, in vivo утворюють комплекси з
аутологічними білками. Доведено, що після внутрішньовенного введення гвінейським свинкам
великих доз новарсенолу у них не вдається отримати шкірну сенсибілізацію до цієї речовини. М.
Чейз (Chase) (1946) встановив, що в результаті перорального введення гвінейським свинкам ДНХБ
пригнічується виникнення сенсибілізації до цієї речовини. Зазначені явища називають феноменом
Сульцбергера—Чейза.
Спостереження про те, що аутологічні білкові носії, як правило, індукують толерантність до
гаптенів, які з ними зв'язані, розкриває базовий механізм природної толерантності. Нагадаємо, що
гаптени можуть розпізнаватися лише рецепторами В-клітин, а білкові носії мають містити також
сайти розпізнавання рецепторами Т-клітин (Т-епітопи). В аутологічних носіях Т-епітопів немає,
оскільки аутореактивні Т-лімфоцити проходять негативну селекцію в тимусі. Отже, толерантності
на рівні Т-клітин (у багатьох випадках), достатньо для відсутності імунної відповіді на той чи інший
антиген.
13.1.1. Ефект дози антигенів.
Толерантність індукують дози антигенів, які відрізняються від імуногенних доз. Між величиною
дози, ступенем толерантності та її тривалістю існує певна залежність. Н. Мітчисон (N. Mitchison) в
експерименті на мишах установив, що толерантність до тимусзалежних антигенів у дорослих мишей
виникає при дозах антигенів, які вищі або нижчі за імуногенні, тобто існує так звана високо- і
низькозонна толерантність. Було показано, що повторні ін'єкції мишам як низьких (0,001 мг), так і
високих (5 мг) доз бичачого сироваткового альбуміну індукують стан толерантності, тоді як
проміжні дози зумовлюють імунні реакції (мал. 94). Вважають, що на формування толерантності
впливають дози антигену, щільність
детермінант на поверхні молекули, час між
ін'єкціями тощо.
За наявності попереднього імунітету
досягнення толерантності до гомологічного
антигену ускладнюється. Це пояснюється
тим, що імунізовані тварини елімінують
антиген значно швидше, ніж інтактні або
толерантні, а також тим, що в імунізованих
тварин за повторної антигенної стимуляції
легше відбувається активація клітин пам'яті,
ніж їх інактивація. Щоб досягти стану
толерантності в імунних мишей, потрібна
доза антигену в 10 разів більша, ніж для
отримання такого самого ефекту в інтактних
мишей.
Низькозонна толерантність не виникає у
мишей nude і після введення антигену
нормальним тваринам разом з
антилімфоцитарною сироваткою (АЛС). Стан
низькозонної толерантності можна зняти введенням лімфотропних вірусів, що вражають лімфоцити
тимусзалежних зон лімфатичних вузлів і селезінки (P. Baker, 1975). На основі цих даних було
зроблено висновки, що низькозонна толерантність зумовлюється Т-клітинами-супресорами. При
низькозонній толерантності в лімфоїдній тканині мишей виникає популяція антигенспецифічних
клітин-супресорів, які пригнічують гуморальний імунітет у системі адаптивного перенесення.
Оскільки низькозонна толерантність зумовлюється Т-клітинами-супресорами, то під впливом АЛС
вона не формується. Сучасні погляди на природу клітин-супресорів залишаються суперечливими.
Однак очевидно, що низькозонна толерантність формується на рівні Т-клітин, а високозонна — на
рівні як Т-, так і В-клітин.
Цікаво, чи відрізняються процеси низько- та високозонної толерантності під час формування
природної толерантності до власних антигенів? З метою пошуку відповіді на це запитання були
58
створені трансгенні миші, які експресували яєчний білок овальбумін у клітинах підшлункової
залози. Одна лінія мишей експресувала овальбумін на високому рівні, а інша — на низькому.
Трансгенним мишам обох ліній вводили сингенні цитотоксичні Т-лімфоцити, специфічні до
епітопів овальбуміну. У мишей з високим рівнем експресії овальбуміну перенесені Т-клітини
гинули внаслідок апоптозу, а в мишей з низьким рівнем — специфічні Т-клітини продовжували
існувати, але не розвивали імунних реакцій щодо підшлункової залози. Виділені з таких мишей Т-
лімфоцити могли розвивати нормальні імунні реакції проти клітин, що експресують овальбумін in
vitro, але в організмі мишей вони чомусь «мовчали». Це явище називають клональним ігноруванням.
Воно пояснюється тим, що Т-лімфоцити не отримують сигнал, достатній для активації, внаслідок
низької щільності специфічних детермінант на клітинах-мішенях. Отже, механізмом високозонної
толерантності в цьому експерименті була клональна делеція, а низькозонної — клональне
ігнорування.
Для досягнення толерантності досить інактивації або блокування клітин з високоафінними
рецепторами. Клітини, що несуть рецептори з низькою афінністю, при цьому можуть залишатися
нетолерантними. Отже, клітини-попередники з високою афінністю рецепторів чутливіші та більш
«ранимі» у період формування толерантності й тому елімінуються вже за невеликих доз антигену.
Чим більш високоафінні рецептори на клітинах-попередниках, тим менші дози толерогену необхідні
для їхньої елімінації.
При ін'єкціях високими дозами антигенів з ад'ювантами толерантність не виникає, оскільки
ад'юванти створюють в організмі «депо» антигену, внаслідок чого провокують реакції запалення,
які вкрай важливі для ініціювання специфічних імунних реакцій.
13.1.2. Фактори, що сприяють створенню штучної толерантності.
На формування толерантності до більшості антигенів впливають фактори, які неспецифічно
пригнічують імунітет: сублетальні опромінення йонізуючою радіацією, дренаж грудної лімфатичної
протоки, хіміопрепарати, що мають імунодепресивну дію, наприклад циклоспорин А, циклофосфан,
імуран, метотрексат, а також АЛС. Імунодепресанти вводять разом з антигеном або впродовж
короткого часу після ін'єкцій антигену.
Перитонеальні макрофаги, опромінені in vitro або в організмі донора, а потім сенсибілізовані
антигеном, на відміну від інтактних, не здатні індукувати синтез антитіл. Вважають, що
радіоактивне випромінювання інактивує процесинг антигену макрофагами, що може бути одним із
механізмів його імунопригнічувальної дії. Інше пояснення — це нездатність опромінених
макрофагів експресувати костимуляторні молекули.
Виникнення толерантності до чужорідних антигенів (трансплантатів) після опромінення реципієнта
або дії імунодепресантів пояснюється тим, що ці впливи гальмують процеси проліферації і
диференціювання активованих зрілих лімфоїдних клітин, зумовлюючи загибель клітин, що
проліферують, а незрілі лімфоїдні клітини, що утворюються зі стовбурових кістковомозкових
попередників, за наявності чужорідних антигенів стають толерантними до них.
Значний успіх сучасної трансплантології насамперед пов'язаний з відкриттям імунодепресивних
препаратів, що специфічно пригнічують функцію лише окремих субпопуляцій клітин. Так,
циклоспорин А пригнічує активацію Т-хелперів у відповідь на стимуляцію ІЛ-2 та власне інгібує
продукування ІЛ-2 в організмі.
Отже, індуковану імунну толерантність можна визначити як повне або часткове гальмування
вторинної імунної відповіді на гомологічний антиген або як незавершену імунну відповідь. По суті,
індукована імунна толерантність — це вторинна імунна відповідь, яка виявляється не підвищеною, а
зниженою реакцією організму на повторне введення чужорідного антигену або її повною
відсутністю.
Штучна імунна толерантність індукується:
1) за повторних введень малих доз антигену впродовж тривалого часу;
2) після введення антигену в надлишковій кількості;
3) у разі застосування розчинних антигенів, звільнених від макромолекулярних аґреґатів;
4) якщо антиген дуже повільно деградує і тому довго зберігається у тканинах, зокрема в
центральних лімфоїдних органах;
5) у разі неонатального введення алогенних стовбурових кровотворних клітин.
Механізми індукування толерантності до алогенних клітин і білкових антигенів, очевидно, різні. До
Т-незалежних антигенів небілкової природи, а також до вірусних і бактеріальних корпускулярних
антигенів толерантність майже не формується (не беручи до уваги імунодефіцитні стани).
Штучна толерантність може сформуватися за такими механізмами:
1) клонального ігнорування — коли клітини «не помічають» антиген, введений у невеликій
кількості;
2) клональної делеції — загибелі специфічних клітин під час розпізнавання великої кількості
антигенів;
3) блокування еферентної ланки — наявність розчинного антигену в сироватці, що блокує
рецептори сенсибілізованих лімфоцитів;
4) блокування аферентної ланки — гальмування активації В-клітин надлишком антитіл, пригнічення
макрофагальної реакції;
5) порушення диференціювання стовбурових клітин, яке зумовлене відсутністю загруднинної залози
або створенням стійкого химерного тимуса.
13.1.3. Відміна толерантності.
Імунокомпетентні клітини дорослих тварин толерантні (ареактивні) щодо антигенів власного
організму, з якими вони контактують в ембріональному періоді. Однак часте виявлення аутоантитіл
у здорових особин свідчить про те, що навіть природна толерантність не є абсолютною. Причини
відміни толерантності, як і механізми її формування, у багатьох випадках залишаються
нерозкритими.
В експерименті штучно індукований стан толерантності припиняється після введення в організм
толерантної тварини живих лімфоцитів лімфатичного вузла сенсибілізованих або нормальних
сингенних тварин. Відміну толерантності зумовлює і трансплантація лімфоцитів від тварин з ГСТ
до цього антигену.
Стан штучної толерантності порушується під впливом неспецифічних стимуляторів лімфоцитів,
наприклад ендотоксинів (ЛПЦ). Здатність до утворення антитіл, втрачена після опромінення,
відновлюється під впливом парентерально введених ендотоксинів.
Після зникнення толерантності здатність виробляти антитіла або відторгати трансплантат
відновлюється. Втрата толерантності до власних антигенів призводить до аутоімунних захворювань,
а втрата толерантності до чужорідних антигенів зумовлює сенсибілізацію. Порушення
толерантності до власних антигенів відбувається спонтанно у мишей лінії NZB. У віці понад 3 міс. у
них виявляються аутоантитіла до ендогенних антигенів: еритроцитів, рибонуклеопротеїдів,
кардіоліпіну, колагену та інших речовин, фактично ця лінія мишей використовується як
експериментальна модель для вивчення системного червоного вовчака. Виявлено лінії курей, в яких
у період статевої зрілості виникає тиреоїдит і виявляються антитиреоїдні аутоантитіла. Зниження
толерантності до власних антигенів переважно залежить від Т-клітин, оскільки не всі В-клітини у
нормі толерантні до всіх аутоантигенів.
Причини порушення толерантності різні. Одна з них — приєднання до власних білків чужорідних
гаптенів. При цьому гаптен може змінювати структуру білка так, що він стає чужорідним для
власного організму. Такий механізм утворення аутоантигенів при тромбоцитопенічній пурпурі
спостерігається у хворих, яких лікували седормідом.
Толерантності немає до антигенів, які в нормі недоступні для лімфоїдних клітин. Деякі місця в
організмі належать до так званих імунопривілейованих сайтів, куди не можуть потрапляти наївні Т-
лімфоцити. До таких сайтів належить, наприклад, центральна нервова система. Показано, що якщо
тварин імунізувати аутологічним білком мієліном, введеним у повному ад'юванті Фрейнда, то у них
розвивається аутоімунна реакція проти власної нервової тканини. Толерантність у центральних
лімфоїдних органах також не формується при пізній появі антигену (сперма), коли толерантність
індукувати вже важко. Так можна пояснити наявність аутоантитіл до сперми при певних патологіях.
Втрата толерантності може відбуватися під впливом ад'ювантів, перехресних антигенів, у результаті
соматичних мутацій У генах, що кодують власні білкові антигени, або в генах рецепторів
імунокомпетентних клітин. У власних антигенів можуть змінюватися фізико-хімічні властивості,
наприклад, агрегований γ-глобулін і вкритий вірусами еритроцит набувають антигенних
властивостей.
Особливо небезпечним є зрив толерантності внаслідок перенесених інфекцій. Вважають, що
інфекційні агенти можуть бути одним з етіологічних факторів у патогенезі багатьох аутоімунних
хвороб. Наприклад, добре відомо, що ревматизм іноді буває пов'язаний з перенесеними
стрептококовими інфекціями, артрит — з такими вірусними інфекціями, як кір і краснуха, тощо. Це
пояснюють перехресними детермінантами білків інфекційних агентів і власних білків. Компоненти
клітинної стінки або вірусного капсиду в цьому випадку виконують роль ад'юванту, стимулюючи
розвиток запалення та ініціювання специфічних імунних реакцій навіть до тих детермінант, до яких
у нормі імунна система була толерантною.
Експериментальною моделлю для вивчення механізмів зриву імунної толерантності після
перенесених інфекцій є трансгенні миші, які експресують білок капсиду вірусу грипу в клітинах
підшлункової залози. В нормі миші толерантні до цього білка і сприймають його як свій власний.
Однак після перенесеної грипозної інфекції у мишей з'являються цитотоксичні лімфоцити, які
атакують клітини підшлункової залози і в мишей розвивається цукровий діабет. Одним із можливих
механізмів зриву толерантності в цьому випадку може бути активація дендритних клітин у слизових
оболонках під дією живих вірусних частин. У нормі дендритні клітини не презентували антигенів
цього вірусу, оскільки він був локалізований лише в межах підшлункової залози. За інфекційного
процесу ДК, які фагоцитували вірусні частинки, активуються, переходять у лімфатичні вузли, де й
презентують вірусні пептиди цитотоксичним лімфоцитам. Отже, стан системи природного захисту
може істотно впливати на індукування чи зрив толерантності, що може виявитися небезпечним у
разі наявності у збудника перехреснореагуючих антигенних детермінант.
Залежно від конкретних умов стан імунної толерантності буває корисним або шкідливим. Природна
толерантність до власних антигенів — явище фізіологічне, що забезпечує гомеостаз. Порушення
толерантності спричинює виникнення аутоімунних процесів. Під час трансплантації індукування
толерантності до антигенів донора сприяє подовженню тривалості виживання трансплантата. За
толерантності до інфекційних агентів знижується резистентність організму до інфекції і тварини
гинуть від малих доз патогенних мікробів. Формування толерантності до антигенів пухлин підсилює
ріст цих пухлин.
13.2. МЕХАНІЗМИ ПРИРОДНОЇ ІМУННОЇ ТОЛЕРАНТНОСТІ.
Елімінація або супресія клонів. Питання про те, чи відбуваються у процесі толерантності
елімінація, інактивація або пригнічення функції клітин, що реагують на певний антиген, остаточно
ще не з'ясоване. Ймовірно, різні прояви толерантності забезпечуються різними механізмами (табл.
55). Як уже зазначалося, механізми індукування високозонної толерантності відрізняються від
низькозонної. Толерантність на рівні Т-клітин відрізняється від толерантності на рівні В-клітин.
Крім того, різняться механізми центральної і периферичної толерантності, толерантності до
розчинних антигенів, антигенів позаклітинного матриксу та мембранних антигенів живих клітин.
Очевидно, у підтриманні стану природної толерантності беруть участь багато різних механізмів.
Толерантність, яка формується під час розвитку лімфоїдних клітин у центральних лімфоїдних
органах, називають центральною. Вона переважно зумовлена елімінацією клонів аутореактивних
клітин. Елімінації насамперед зазнають клітини з високою афінністю рецепторів до
найпоширеніших аутоантигенів. Ті потенційно аутореактивні клітини, які виселяються з
центральних лімфоїдних органів на периферію, підлягають периферичній толяризації. Периферична
толерантність забезпечується великою кількістю різних механізмів, головні з яких — запобігання
розпізнаванню антигену (ігнорування), відсутність активаційних і костимуляторних сигналів
(анергія), специфічне пригнічення функцій (супресія), загибель аутореактивних клітин на периферії
внаслідок повторних стимуляцій антигеном (апоптоз). Вважають, що головні механізми
підтримання толерантності ще не відкрито.
13.2.1. Елімінація клонів при центральній толерантності
Перше теоретичне пояснення імунної толерантності було зроблено на основі клонально-селекційної
теорії імунітету. Припускали, що антиген виявляє депресивну дію на незрілі лімфоїдні клітини
специфічного клону, запобігаючи їх проліферації та диференціюванню. Пізніше уявлення про
механізми толерантності розширились, але теорія елімінації клонів залишилася основною догмою.
Вважають, що елімінація аутореактивних клонів (клональна делеція) є головним механізмом
центральної толерантності.
Незрілі аутореактивні Т-клітини елімінуються в тимусі, а незрілі аутореактивні В-клітини — в
кістковому мозку.
Роль тимуса у створенні й підтриманні толерантності. Як було розглянуто в попередньому
розділі, в тимусі попередники Т-лімфоцитів набувають здатності розпізнавати власні молекули
МНС І і МНС II класів, тобто стають рестриктованими за МНС. МНС-рестрикція може призводити
до появи клітин з підвищеною аутореактивністю до власних антигенів гістосумісності. Щоб
уникнути можливості потрапляння таких клітин на периферію, в тимусі відбувається негативна
селекція клітин, які розпізнають власні МНС з підвищеною афінністю чи авідністю.
Вважають, що толерантності до власних тканин тимоцити набувають у результаті елімінації в
тимусі клонів лімфоцитів, здатних достатньо афінно розпізнавати власні антигени МНС. Про роль
тимуса у створенні толерантності свідчить і те, що толерантність до деяких антигенів вдається
передати трансплантацією тимуса. У тимусі виявлено гормон (прегненолон), який має
імунодепресивну дію і сприяє виникненню толерантності.
Тривалість толерантності до алотрансплантатів шкіри часто збігається з перебуванням у тимусі
алогенних кровотворних клітин. Це дає змогу вважати, що тимус — основне місце формування
толерантності. Щодо клонів, які здатні реагувати проти власних антигенів, то можна припустити,
що їх знешкодження досягається внаслідок загибелі. Елімінація клітин-попередників, що зв'язали
велику кількість антигену, здійснюється в результаті апоптозу та фагоцитозу. Можливо, в деяких
випадках толерантність досягається тим, що специфічні клони під час розпізнавання антигену в
тимусі диференціюються до зрілих клітин без проліферації, що призводить до виснаження їх пулу
(теорія кінцевого диференціювання).
Практично неможливо, щоб усі власні антигени були наявні у тимусі в кількості, необхідній для
елімінації аутореактивних клітин. Підраховано, що всього в організмі людини синтезується
приблизно 50 тис. білків, з яких може утворитися до 30 млн антигенних пептидів. На кожній клітині
в середньому експресується близько 100 тис. молекул МНС. Тому ймовірність потрапляння на
поверхню клітини у комплексі з МНС для кожного пептиду в середньому становить 0,3%.
Відповідно, ймовірність презентації двох однакових пептидів ще менша. Безумовно, різні білки
мають неоднаковий рівень експресії в клітині, але збільшення ймовірності презентації одних білків
робить ще менш імовірною презентацію інших. Тому формування центральної толерантності в
тимусі відбувається переважно до тих білків, які мають високий рівень експресії.
Центральна толерантність В-клітин у кістковому мозку. Які наслідки очікують незрілі В-
клітини, якщо вони зв'яжуть власні антигени в кістковому мозку? Вважають, що незрілі В-клітини
не здатні активуватися після розпізнавання антигену, але можливі два наслідки такого
розпізнавання — загибель або редагування рецепторів.
Під час розвитку в кістковому мозку В-лімфоцити, що розпізнали власні антигени, гинуть унаслідок
апоптозу. Такий шлях є найімовірнішим для незрілих В-клітин. Він зумовлює клональну делеціію
аутоспецифічних В-лімфоцитів. Вона є головним механізмом толяризації В-клітин до аутоантигенів.
Роль толерогенів у цьому випадку виконують антигени, яких дуже багато в організмі, та антигени,
які мають полімерну структуру з багатьма однаковими антигенними детермінантами. До таких
антигенів належать білки позаклітинного матриксу — колаген, ламінін, фібронектин, фібрин,
протеоглікани, гіалуронова кислота тощо. Такі антигени становлять небезпеку активації В-клітин на
зразок тимуснезалежних антигенів завдяки перехресному зв'язуванню поверхневих
імуноглобулінових рецепторів. Тому В-клітини, специфічні до цих аутоантигенів, мають бути
знищені у першу чергу. До групи таких антигенів можна також віднести молекули МНС та інші
мембранні глікопротеїни, які досить щільно експресовані на мембранах власних клітин організму.
Слід зазначити, що у разі зв'язування незрілою В-клітиною в кістковому мозку певного
аутоантигену вона не завжди гине. Природа винайшла спосіб економнішого використання
потенціалу В-клітин. Деякі В-клітини можуть проводити подальше реаранжування генів легких
ланцюгів рецептора. При цьому може змінюватися специфічність рецепторів і втрачатися
аутореактивність. Цей процес, що називається редагуванням рецепторів, докладніше описаний у
попередньому розділі. Він характерний також для Т-клітин під час їхнього розвитку в тимусі, але
вивчений у них недостатньо.
13.2.2. Механізми периферичної толерантності.
Делеція клонів на периферії. Після виселення з центральних лімфоїдних органів лімфоцити,
передусім В-клітини, певний час (кілька діб) залишаються незрілими. Дозрівання лімфоцитів
відбувається у периферичних лімфоїдних органах — В-клітини дозрівають у селезінці, а Т-клітини
— у Т-зонах лімфатичних вузлів. Під час такого дозрівання можлива додаткова толяризація
лімфоцитів. У разі надходження повторних антигенних стимуляцій від прилеглих тканин лімфоцити
програмуються на самознищення. Така клональна делеція на периферії може відбуватися і в інших
анатомічних структурах. Наприклад, В-клітини, що мігрують у лімфоїдні фолікули, можуть
затримуватися після розпізнавання своїми рецепторами аутоантигенів, які оточують клітини.
Несвоєчасне потрапляння В-лімфоцита в лімфоїдний фолікул також призводить до його загибелі.
Важливим механізмом, що контролює толерантність до власних тканин, є індукована активацією
загибель лімфоцитів (AICD — activation induced cell death) (див. розд. 10). Після активації на Т-
лімфоцитах збільшується кількість рецепторів Fas і FasL. Це має велике біологічне значення.
Активовані лімфоцити, що знаходяться поруч, можуть вбивати один одного. Таке явище називають
«братовбивчою війною» лімфоцитів, унаслідок чого більшість активованих лімфоцитів гине
впродовж кількох діб. Отже, випадково активовані клони аутореактивних Т-клітин можуть загинути
ще до того, як зможуть спричинити певні ушкодження. Показано, що у мишей з дефектами генів Fas
і FasL виникає лімфопроліферативний синдром, який характеризується певними аутоімунними
ушкодженнями.
Ігнорування антигену. Якщо антиген міститься в організмі у невеликій кількості копій, то Т- і В-
клітини можуть зовсім не реагувати на нього, навіть якщо їх рецептори специфічні саме до цього
антигену. Такий стан ареактивності лімфоцитів називають ігноруванням антигену. Суть ігнорування
антигену полягає в тому, що для активації лімфоцитів, як правило, необхідне зв'язування багатьох
поверхневих рецепторів (40 — 100 для Т-клітин і 100 — 1000 для В-клітин). Крім того, розчинний
моновалентний антиген у невеликій концентрації не здатний зумовлювати активацію В-клітин,
оскільки він не може перехресно зв'язувати рецептори.
Більшість аутоантигенів експресується в клітині на рівні, недостатньому для розпізнавання Т-
клітинами, і тому вони не можуть спричинювати аутоімунні реакції. Малоймовірно, щоб антигенні
пептиди, які утворилися з білків, що синтезуються клітиною у невеликій кількості копій, могли бути
наявні у достатній кількості з однаковими молекулами МНС на клітинній поверхні. Для того щоб
активувати Т-клітину, треба, щоб клітина-мішень мала хоча б кілька десятків (не менше 40)
однакових Т-епітопів з однаковими молекулами МНС. У разі меншої їх кількості взаємодія з
рецепторами Т-клітин може відбутися, але сигналу, достатнього для активації, Т-клітина не
отримає. Тому більшість аутоантигенів Т-клітини просто «ігнорують». Вірусні антигени, навпаки,
синтезуються в інфікованій клітині у надмірній кількості копій і тому можуть знаходитися на
поверхні клітини з високою щільністю, достатньою для активації Т-клітин.
Іншим прикладом ігнорування є недоступність антигену для взаємодії з лімфоцитами. Це
насамперед стосується антигенів, які знаходяться в імунопривілейованих сайтах. Порушення
спеціальних бар'єрів, що відокремлюють сайти від лімфоїдної системи, призводить до розпізнавання
цих антигенів лімфоцитами та розвитку аутоімунних реакцій. Таке порушення може відбуватися
внаслідок місцевого запалення або травмування.
Відсутність «професійної» допомоги. Роль Т-хелперів. У регуляції стану імунної толерантності
важливу роль відіграють Т-хелпери. При толерантності не відбувається кооперативна взаємодія Т- і
В-лімфоцитів, яка необхідна для імунної відповіді В-клітин на тимусзалежні антигени, оскільки В-
клітини не можуть відповідати на антиген, до якого Т-хелпери толерантні. Саме тому для створення
толерантності необхідна елімінація аутореактивних Т-хелперів, які потенційно можуть розпізнавати
епітопи власних білкових антигенів у комплексі з МНС II.
Імунна толерантність до власних тканин, очевидно, залежить від відсутності Т-клітин, специфічних
до власних антигенів. Аргументацією на користь такого припущення є такі встановлені факти:
толерантність легше формується в популяції Т-клітин, ніж у популяції В-клітин. У Т-клітин її
можна індукувати меншими дозами антигену за більш короткої експозиції, але зберігається вона
впродовж тривалішого часу. До тимуснезалежних антигенів толерантність створити складніше,
оскільки нетолерантні В-клітини залучаються в імунну відповідь без участі Т-клітин.
Білоквмісні компоненти власних тканин є тимусзалежними антигенами. Вони можуть виявити свої
антигенні властивості лише в асоціації з продуктами МНС класу II. Тому за відсутності
специфічних Т-хелперів В-клітини, що розпізнали такі аутоантигени, не активуються. Отже, навіть
специфічні до власних антигенів В-клітини, які презентують продукти МНС класу ІІ на поверхні
разом з епітопами аутоантигенів, перебувають у стані «імунного мовчання». В-клітини, які
розпізнали і поглинули внаслідок ендоцитозу певні власні антигени, Т-хелперами не активуються,
оскільки негативна селекція рецепторів Т-хелперів більш сувора. В-клітини, які не отримують
допомоги від Т-хелперів, перебувають у стані анергії і через кілька діб загинуть у результаті
апоптозу. Толерантні тварини можуть мати В-клітини, що несуть рецептори, специфічні до
толерогену. Показано, що in vitro лімфоїдні клітини мишей, толерантних до еритроцитів барана,
утворюють з ними розетки, тоді як антитіла у сироватці таких тварин відсутні. За відсутності
специфічних Т-хелперів будуть пригнічені також клітинні реакції, оскільки Тх1 активують наївні
цитотоксичні лімфоцити через утворений АПК місток. Отже, толерантність зумовлюється не тільки
елімінацією клонів лімфоїдних клітин, що специфічні до антигену, а й відсутністю стимулів,
необхідних для активації.
Роль дендритних клітин. Дендритні клітини (ДК) є ініціаторами специфічних імунних реакцій, а
тому вони відповідальні за дотримання стану толерантності до власних антигенів. У зв'язку з цим
ДК мають особливі функції, характерні для цього типу клітин. По-перше, активація ДК відбувається
лише за умов локального запалення. Умовою активації ДК є розпізнавання їхніми рецепторами
консервативних структурних мотивів, характерних для патогенних агентів. По-друге, в активованих
дендритних клітинах протеасома заміщується імунопротеасомою. Важливою і ще не з'ясованою
особливістю імунопротеасоми є ефективніший процесинг вірусних білків, ніж власних. Тому на
активованих ДК зменшується, якщо не припиняється зовсім, презентація власних білків. По-третє,
ДК здатні до перехресної презентації антигенів, тобто поглинуті антигени презентуються з МНС І і
МНС II. По-четверте, для остаточної активації ДК необхідна їх взаємодія з Т-хелпером (хоча б

59
наївним). По-п'яте, після активації ДК мігрують у вторинні лімфоїдні органи, де припиняють
процеси поглинання і процесингу антигенів, тобто інтердигітальні дендритні клітини лімфовузлів
представляють лише ті антигени, які вони поглинули в місці запалення.
Активовані ДК активують наївні Т-клітини (Т-хелпери і ЦТЛ), унаслідок чого запускаються
специфічні гуморальні та клітинні імунні реакції. Особливо важлива роль ДК при першому контакті
організму з антигеном, оскільки з усіх типів АПК лише вони, як вважають, можуть бути
ініціаторами імунної відповіді.
Відсутність костимуляції під час розпізнавання антигену. При кооперативних взаємодіях клітин
у системі Т-хелпер —АПК обидві клітини обмінюються костимуляторними сигналами, які разом із
сигналом про розпізнавання антигену необхідні для активації клітин (див. розд. 10). Нестача
костимуляторних молекул на активованих клітинах може бути наслідком збоїв під час активації або
залученням до активації незрілих клітин. За відсутності костимуляторних сигналів клітини можуть
загинути внаслідок апоптозу або набути стану анергії, яка через кілька діб закінчується апоптозом.
У стані анергії клітини не здатні здійснювати ефекторні функції, але можуть виконувати важливі
імунорегуляторні функції, зокрема у процесах підтримання толерантності. Наприклад, Т-клітини у
стані анергії можуть конкурувати з наївними Т-клітинами за місця зв'язування з АПК, а можливо,
навіть індукувати стан анергії у тих АПК, з якими вони взаємодіють. Це може призводити до
поступового збільшення частки алергічних клітин у популяції специфічних до певного антигену Т-
клітин і АПК. Хоча такий механізм є гіпотетичним, його можна вважати досить імовірним.
Блокування рецепторів імунокомпетентних клітин. Блокування рецепторів лімфоцитів можливе
у разі тривалої персистенції антигену в тканинах за його високої концентрації та низької
імуногенності. Вважають, що за таким механізмом підтримується природна толерантність до
розчинних білкових антигенів крові, лімфи та позаклітинної рідини. Крім того, припускають, що за
таким механізмом індукується штучна толерантність до розчинних білків (феномен Дрессера).
Деякі дослідники вважають, що низька імунна реактивність до ракових тканин, поряд з іншими
причинами, може бути також зумовлена толерантністю до пухлиноспецифічних антигенів унаслідок
блокування специфічних рецепторів лімфоцитів розчинними антигенами, які постійно надходять у
внутрішнє середовище.
Ефект блокування рецепторів намагаються застосувати для розроблення методів лікування певних
аутоімунних хвороб.
Роль інгібіторних рецепторів. Інгібіторні рецептори на поверхні імунокомпетентних клітин у
нормі виконують функцію негативних зворотних зв'язків у системі регуляції імунної відповіді,
тобто інгібують активацію клітин за великої кількості активованих специфічних клітин або антитіл
(див. розд. 10, 11). В-клітини сорбують цитофільні IgG з плазми крові за допомогою своїх FcγRII, і
такі рецептори можуть бути інгібіторами активації В-клітин за умови перехресного зв'язування їх та
IgM-рецепторів тієї самої специфічності відповідним антигеном.
Іншим добре вивченим інгібіторним рецептором є молекула CTLA-4, що експресується
активованими Т-лімфоцитами. Завдяки більшій, ніж CD28, афінності до стимуляторного ліганду
антигенпрезентувальних клітин В7.1(2) за недостатньої експресії останнього молекули CTLA-4
першими залучаються до взаємодії, що призводить до гальмування активації лімфоцитів.
Можливо, за деяких обставин інгібіторні рецептори можуть брати участь у підтриманні стану
толерантності до певних антигенів. Також можлива участь інгібіторних рецепторів у процесах
десенсибілізації організму.
Клітини-супресори. Чимало феноменів підтримання імунної толерантності пояснюють існуванням
спеціалізованої популяції клітин-супресорів. Про існування таких клітин свідчать певні
спостереження. За спонтанної втрати толерантності імунна відповідь у деяких випадках
здійснюється за вторинним типом. Поява імунітету після зникнення толерантності свідчить про те,
що остання була створена в такому випадку без елімінації клонів. Крім того, у лімфоїдних органах
толерантних тварин є морфологічні зміни, які подібні до морфологічних змін в імунних тварин. В
обох випадках спостерігається гіперплазія лімфоїдних елементів.
У мишей під час толерантності до еритроцитів барана кількість клітин, що розпізнають антиген, не
лише не зменшується, а й може збільшуватися. Виявлення клітин-супресорів дало б змогу
розглянути ще один механізм, за допомогою якого клітини перестають відповідати на цей антиген
без елімінації клону.
Дослідженнями на мишах показано, що стан толерантності зникає після їх опромінення. Тварини,
що втратили толерантність, набувають високої реактивності до повторного введення специфічного
антигену. На основі цих даних зроблено висновок, що толерантність зумовлюється супресорним
впливом специфічної популяції лімфоцитів, чутливих до опромінення. Доказом існування
толерогенної популяції лімфоцитів є такі дані. У разі одночасного введення опроміненим тваринам
суміші лімфоцитів від сингенних нормально імунізованих і толерантних тварин імунна відповідь
іноді стає пригніченою. Це пояснюється тим, що популяція лімфоцитів-супресорів толерантних
тварин пригнічує імунну відповідь антигенреактивних клітин. Однак спроби виділити Т-лімфоцити-
супресори в чистому вигляді та схарактеризувати їхній фенотип і понині залишаються
безуспішними. Спочатку функцію супресорів відводили певній популяції CD8Т-клітин. Однак ці
погляди були переглянуті. Ймовірно, що Т-лімфоцити-супресори — це Т-клітини фенотипу як CD8,
так і CD4, в яких у стані активації відсутня експресія певних костимуляторних молекул. Завдяки
цьому вони можуть індукувати стан анергії в АПК (дендритних клітинах, макрофагах, В-
лімфоцитах), і такі АПК самі можуть виконувати функцію супресії імунної відповіді.
У зв'язку з великою кількістю противників гіпотези про існування Т-супресорів у сучасній
літературі Т-клітини із супресорними властивостями часто почали називати Т-регуляторами, або
вето-клітинами.
З уявленнями про Т-супресори були пов'язані певні сподівання щодо створення специфічних вакцин
проти аутоімунних хвороб. Вважалося, що після імунізації такими вакцинами виникатиме стан
стійкої толерантності, зумовлений активністю Т-супресорів.
Багато проявів імуносупресії можна пояснити існуванням двох субпопуляцій Т-хелперів — Тх1 і
Тх2, що перебувають між собою в антагоністичних стосунках. Під час активації гуморальної ланки
імунітету під дією Тх2 може індукуватися толерантність клітинної ланки, і навпаки, за активації
клітинної ланки під дією Тх1 може спостерігатися відсутність антитілопродукування. Щонайменше,
антагонізм Тх1 і Тх2 добре пояснює явища розщепленої толерантності, яка часто спостерігається в
експерименті. Чи пов'язаний подібний механізм із підтриманням стану природної толерантності,
поки що сказати важко.
Явище антагонізму Тх1 і Тх2 намагаються використовувати для лікування аутоімунних
захворювань. З'ясувалось, що деякі аутоімунні захворювання пов'язані з дисбалансом Тх1 і Тх2.
Наприклад, спонтанний інсулінзалежний діабет у мишей NOD (експериментальна модель діабету),
як правило, зумовлюється аутоімунною атакою ЦТЛ, які розпізнають епітопи білка теплового шоку
Нsр60. Якщо мишей імунізувати пептидом р277 з цього білка, то аутоімунній патології вдається
запобігти. Виявилося, що імунізація пептидом стимулює активацію Тх2, які пригнічують активність
аутоімунних Тх1 і ЦТЛ.
Антиідіотипічна супресія. Тривалий час центральною догмою уявлень про природу
імунорегуляції була теорія ідіотипічних сіток. Вважали, що ідіотипи антитіл (антигензв'язувальні
центри) самі можуть виступати як антигенні детермінанти і зумовлювати синтез антиідіотипічних
антитіл, які блокують попередні антитіла. Змінюючись один за одним, різні антиідіотипічні антитіла
призводять до хвилеподібного згасання імунної відповіді. Можливо, певні стани хронічної
імуносупресії можна пояснити впливом антиідіотипічних антитіл.
Індукування антиідіотипічних антитіл також намагаються використати з терапевтичною метою для
лікування аутоімунних хвороб. Наприклад, анти-ДНК-антитіла у мишей лінії NZB мають спільний
ідіотип, який позначають 16/6Id. Пептиди, які мають послідовність, аналогічну CDR1-ділянці анти-
ДНК-антитіл, здатні індукувати припинення розвитку системного вовчака у NZB-мишей. Імовірно,
що такі пептиди індукують появу антиідіотипічних антитіл, що пригнічують аутоімунну відповідь.
Сучасні уявлення про механізми природної толерантності наведено в табл. 56.
ВИСНОВКИ.
Усі клітини специфічної складової імунної системи проходять негативний відбір під час розвитку в
первинних лімфоїдних органах. Завдяки негативному відбору популяція лімфоцитів позбавляється
клітин, які специфічні до найпоширеніших в організмі аутоантигенів. На думку більшості
дослідників, клональна делеція аутоспецифічних лімфоцитів є головним механізмом формування
природної толерантності.
На периферії створено умови для запобігання активації потенційно аутореактивних клітин, що не
зазнали негативного відбору. Це насамперед залежність активації Т-хелперів від дендритних клітин
і залежність В-клітин від Т-хелперів. Фактично розпізнавання кожного антигену має бути
потрійним: дендритними клітинами, Т-хелперами та ефекторними лімфоцитами. Крім того, має
бути висока локальна щільність антигену — за недостатньої щільності Т- і В-клітини його
ігнорують. Очевидно, що в реалізації периферичної толерантності беруть участь клітини з
супресорними властивостями. Теоретично супресію імунної відповіді можуть забезпечувати як Т-
клітини, так і АПК тоді, коли вони перебувають у стані готовності до взаємодії з наївними
клітинами, але не здатні забезпечити їх повноцінними костимуляторними сиґналами. Можлива
також роль антиідіотипічних взаємодій або інших, ще не встановлених механізмів у підтриманні
толерантності до власних антигенів.
Контрольні запитання
1. Які експериментальні спостереження стали основою для відкриття толерантності?
2. У чому полягає відмінність між термінами «толерантність», «ареактивність» та «імунодефіцитний
стан»?
3. Як пояснює механізми формування толерантності теорія Ф. Бернета?
4. Яка роль тимуса у формуванні природної толерантності?
5. Чому складно сформувати штучну толерантність до корпускулярних вірусних і бактеріальних
антигенів?
6. Що таке центральна і периферична толерантність?
7. В яких експериментальних системах штучна толерантність найбільш подібна до природної?
8. Які механізми толяризації аутореактивних клітин на периферії?
РОЗДІЛ 14. ЕВОЛЮЦІЯ ІМУНІТЕТУ.
У процесі еволюції живої матерії виникла, сформувалася, розвивалася і вдосконалювалася система
захисту організму від проникнення й розмноження носіїв чужорідної генетичної інформації та
контролю за збереженням цілісності власної генетичної інформації. У ссавців вона досягає
найвищого рівня і складається з природних і набутих факторів.
14.1. ПРИРОДНІ ФАКТОРИ ІМУНІТЕТУ.
Фактори природної резистентності, такі як покривні тканини, клітини із захисними функціями та
гуморальні фактори захисту, тромбування ран, коагуляція гемолімфи, інкапсуляція чужорідного
матеріалу, з'явилися на ранніх етапах еволюційного розвитку і більшість із них не лише
зберігаються впродовж еволюції, а й у міру ускладнення організмів значна частина з них
удосконалюється — у них виникають нові структури. Внаслідок цього з'являються відповідні
функціональні активності, які відіграють важливу роль у становленні й функціонуванні адаптивних
факторів імунної відповіді та регуляції багатьох функцій організму, що сприяло формуванню
багаторівневої структури імунної системи.
Покривні тканини. Одними з найбільш ранніх факторів природного захисту від проникнення
чужорідних об'єктів, що зберегли свою функціональну активність і донині, є бар'єри, які
формуються зовнішніми покривами. Вони виявляються вже у найпримітивніших живих істот.
Будова і функціональна активність зовнішніх покривів у процесі еволюційного розвитку
ускладнювалася; з'являлися допоміжні захисні фактори — зовнішні скелети, кальциновані
раковини, панцирі, луска, пір'я, зроговіння епітелію шкіри, шерсть тощо. Крім того, ускладнювалася
і функціональна активність — у зовнішніх покривах з'являються залози, які виділяють слизоподібні
речовини з бактерицидними факторами. Так, кишковопорожнинні та кільчасті черви виділяють
слиз, який містить бактерицидні речовини, в тому числі фактор, подібний до лізоциму.
Важливу роль у захисних реакціях відіграють слизові оболонки, особливо дихальних і травних
систем. Кишечник уперше з'являється у членистоногих з постійним виділенням слизу та деяких
бактерицидних речовин. Під час формування травного каналу та дихальної системи на слизових
з'являються війки, за допомогою яких та завдяки скороченню м'язів відбувається рух слизу, що
сприяє видаленню мікроорганізмів. У шкірі хребетних з'являються потові залози, які продукують
низку захисних факторів.
Клітинні фактори захисту. Фагоцитоз. Для знешкодження мікроорганізмів, що проникли в
організм живих істот, одним із найефективніших факторів є фагоцитоз. Захисна функція фагоцитів
виникла зі здатності деяких клітин здійснювати функцію травлення.
Уже найпростіші здатні поглинати чужорідні речовини, в тому числі й мікроорганізми, та
захоплювати їх у вакуолі травлення. У них виявляється вибіркова здатність поглинати певні
мікроорганізми. Найпростіші можуть відрізняти «своє» від «чужого», хоча механізми такого
розпізнавання ще не з'ясовано.
У найпримітивніших первинних багатоклітинних одношарових організмів — губок, у яких клітини
ще не об'єднані в тканини, з'являються великі рухомі, без постійної форми клітини мезоглії —
археоцити, або блукаючі амебоцити. Вони містять велику кількість лізосом і здатні розпізнавати «не
своє», поглинати та перетравлювати чужорідні об'єкти, зокрема мікроорганізми, та розносити
продукти метаболізму по всьому організму. Амебоцити беруть активну участь у процесах
регенерації ушкоджених ділянок і є факторами захисту від проникнення мікроорганізмів у
внутрішнє середовище.
Крім того, амебоцити здатні здійснювати характерний для природних кілерів лізис клітин,
інкапсулювати патогени великих розмірів, виділяти бактерицидні фактори і цитокіноподібні
речовини. Припускають, що амебоцити примітивних безхребетних стали родоначальниками клітин
лімфомієлоїдного комплексу.
У кільчастих червів з'являються замкнена кровоносна система і вторинна порожнина — целом. У
целомі виявляється кілька типів клітин з протективними властивостями. Серед останніх виділяють
гіалінові амебоцити — нейтрофіли-макрофаги з фагоцитарною активністю, здатністю розпізнавати
«чуже» і відторгати трансплантати — та подібні до незрілих лімфоцитів хребетних клітини, що
мають структури, подібні до маркерів Т-лімфоцитів ссавців, розпізнають «чуже» і є основними
факторами трансплантаційного відторгнення та адаптивного перенесення. У кільчастих червів
уперше здійснюється розподіл клітин з імунними потенціями на макрофаги-нейтрофіли та
лімфоцитоподібні клітини.
Серед молюсків виділяють дві основні групи клітин з імунними властивостями. Одні,
лімфоцитоподібні, клітини циркулюють у гемолімфі, утворюють розетки з еритроцитами барана,
під час контакту з ІЛ-2 та ІФН-γ активують свою цитолітичну активність. Інша група клітин має
фагоцитарну активність, бере активну участь в інкапсуляції бактерій.
У членистоногих виявлено шість типів клітин з певними проявами імунних властивостей. Серед них
виділяють макрофагоподібні клітини з фагоцитарною активністю (плазматоцити); подібні до малих
лімфоцитів прогемоцити; можливі попередники мастоцитів — адиногемоцити, а також оеноцити
(кристалічні клітини), що відіграють певну роль в інкапсуляції патогенів.
Серед представників голкошкірих у крові та целомі крім таких клітин, як у членистоногих, виявлено
вібруючі клітини з джгутиками і гранулами. У процесі ізоляції чужорідних корпускул вони
коагулюють і руйнуються. У голкошкірих на лімфоцитоподібних клітинах виявлено ІЛ-2 та
рецептор до нього, а також антигени CD7 та CD4, які характерні для хребетних.
У покривників, що займають проміжне положення між безхребетними та хребетними, фагоцитарну
активність мають багато клітин — макрофаги, грануло- й амебоцити, гіалінові клітини. Основними
клітинами гемолімфи є вакуолізовані клітини, що містять ванадій і нарівні з функцією перенесення
кисню здатні до деяких неспецифічних реакцій — інкапсуляції чужорідних часточок.
Лімфоцитоподібні клітини у покривників характеризуються помірною цитоцидною активністю,
беруть активну участь у відторгненні ало- та ксенотрансплантатів і подібні до лімфоцитів та до
стовбурових гемопоетичних клітин хребетних.
Одна з початкових стадій фагоцитозу — хемотаксис. Хемотаксичні речовини білкової природи з
молекулярною масою близько 10 кДа виявлено у кільчастих червів, молюсків, комах.
Аглютиніни, як фактори адгезії, виявлено у молюсків, а в членистоногих крім аглютинінів як
опсоніни виступають комплементподібні білки, активність яких пов'язана з фенолоксидазною
системою.
Процес поглинання чужорідних об'єктів у безхребетних подібний до такого у хребетних, але має
деякі особливості. У безхребетних біохімічні процеси кілінгу та утилізації чужорідних об'єктів
вивчені у молюсків і членистоногих. У членистоногих до бактерицидних речовин належать
пероксид гідрогену (водню), мієлопероксидаза, лізоцим, хітиназа, меланін та профенолоксидазна
активність. Під час активації фенолоксидазної системи утворюються активовані кисневі метаболіти,
хіноїди, меланін, оксид азоту (II) і пероксинітрит. Подібною активністю характеризуються і
гемоцити молюсків. Про активність пероксидази у молюсків існують різні дані — в одних молюсків
вона відсутня, а в інших — виявлена.
Природна цитотоксичність виявляється в усіх безхребетних організмів. У примітивних
безхребетних вона здебільшого реалізується блукаючими амебоцитами, а у більш організованих —
лейкоцитами, переважно лімфоцитоподібними клітинами. У риб лейкоцити периферичної крові
здатні убивати власні трансформовані клітини.
Лімфоцитоподібні клітини кільчастих червів, голкошкірих, покривників реагують проліферацією
на КонА, ФГА, ЛПЦ і за своїми властивостями наближаються до гемопоетичних стовбурових
клітин хребетних. У вищих безхребетних (членистоногих, покривників) виникає механізм
постійного відновлення популяцій гемоцитів унаслідок їх диференціювання зі стовбурових
кровотворних клітин.
Слід зазначити, що більшість лімфоцитоподібних клітин багатьох безхребетних мають рецептори до
чужорідних алоантигенів та деяких мікробних структур (ЛПЦ) і є мобільними резервними
клітинами, які постійно заповнюють загальний пул гемоцитів, а після вторгнення патогенів
активуються і зміцнюють захист організму. Вони нагадують «наївні» лімфоцити ссавців.
У крові примітивних хребетних (круглоротих) існують подібні до лімфоцитів і плазмоцитів клітини,
але їхня функція не з'ясована, оскільки в крові не виявлено імуноґлобулінів та відсутні органи
диференціювання цих клітин. За антигенної стимуляції кількість подібних до плазмоцитів клітин
збільшується — можливо, вони продукують антигензв'язувальні фактори комплементу, які раніше
вважали за антитіла. У круглоротих не виявлено ні генів Ig, ні ТкР, ні генів антигенів МНС І та
МНС II класів, ні білків, що сприяють реаранжуванню V-генів або транспорту пептидів до МНС. Ці
клітини реагують на Т- і В-мітогени. Основними учасниками відторгнення трансплантатів у
круглоротих є нейтрофіли та еозинофіли.
Мастоцити виявляються у примітивних хребетних — круглоротих. У крові риб містяться мастоцити
та базофіли. В крові амфібій виявлено дві популяції мастоцитів.
Гуморальні фактори природного імунітету. У безхребетних — кільчастих червів, молюсків,
членистоногих, голкошкірих — виявлено групу гуморальних факторів, здатних лізувати еритроцити
— гемолізини, які є поліморфною структурою з кількома компонентами, термолабільні, чутливі до
протеолітичних ферментів. Активність їх пригнічується хелатами і потребує наявності
двовалентних катіонів. Вони здатні пригнічувати життєдіяльність деяких бактерій і лізувати їх.
Природа літичної дії комплементоподібних факторів і гемолізинів неоднакова. Кількість
гемолізинів збільшується після введення в організм чужорідних об'єктів або ушкодження покривних
тканин. Серед інших лізинів у безхребетних найчастіше виявляються фактори з лізоцимподібною
активністю — у кільчастих червів, молюсків, членистоногих, голкошкірих. Лізоцим міститься в
гемоцитах і при їх активації фермент може викидатися у навколишнє середовище. Кількість
лізоциму значно зростає при введенні безхребетним чужорідних часточок, особливо
мікроорганізмів.
У більшості безхребетних виявлено аглютиніни, здатні аглютинувати різні живі істоти та клітини.
Деякі з них характеризуються певною специфічністю — вони здатні аглютинувати еритроцити
лише певних тварин і лише певні мікроорганізми. Кількість лізоциму, лізинів, аглютинінів та інших
протективних факторів збільшується після введення безхребетним еритроцитів, патогенів або
ушкодження покривів. Аглютиніни можуть бути не тільки внутрішньоклітинними компонентами, а
й клітинними рецепторами, що здатні злущуватися та існувати в гемолімфі. Аглютиніни крім
антибактеріальної активності виявляють також властивості опсонінів і хемоатрактантів. Аглютиніни
голкошкірих за будовою подібні до важких ланцюгів IgM.
У членистоногих, комах у разі ушкодження зовнішніх покривів або проникнення мікроорганізмів
індукується синтез ряду бактерицидних речовин — лізоциму (три варіанти), атацинів, цекропінів,
дефензинів тощо. Атацини мають сильну бактерицидну активність, але механізм дії невідомий.
Цекропіни (п'ять варіантів) — низькомолекулярні катіонні білки (близько 5 кДа) характеризуються
вираженою бактерицидною активністю. Кожний цекропін може лізувати кілька видів бактерій — як
грампозитивних, так і грамнегативних, але вони не діють на клітини еукаріотів і виявляють певну
специфічність до ряду патогенів. Механізм дії цекропінів подібний до дії системи комплементу або
перфоринів ссавців. Одні речовини синтезуються постійно, інші — в процесі активації. Індукування
синтезу протимікробних речовин відбувається швидко (впродовж кількох годин) і за механізмом
перебігу нагадує реакцію гострої фази у ссавців.
У голкошкірих є комплементоподібна система з двома білками — білок С3/С4/С5, що циркулює в
гемолімфі, і подібний до рецептора комплементу білок. У покривників система комплементу
ускладнюється — з'являються гомологічні С1r-, C1s- і C1q-структури та виникає лектиновий спосіб
активації комплементу. У членистоногих та інших безхребетних аналогом системи комплементу є
каскадна система активації профенолоксидази, що ефективно діє на гриби, деякі види бактерій і
навіть на багатоклітинні організми. Система комплементу існує в усіх хребетних, але в круглоротих
і деяких риб відсутні компоненти класичного шляху активації комплементу С1, С2, С4.
Цитокіноподібні речовини в незначній кількості виявляються на всіх етапах еволюції безхребетних і
хребетних. Так, у найпростіших виявлено молекулу Е2-1, яка подібна ІЛ-2 хребетних. У кільчастих

60
червів, молюсків, голкошкірих містяться речовини, подібні до ІЛ-1а, ІЛ-1b, ФНП та ейкозаноїди й
опіатні пептиди, що відіграють певну роль у регуляції імунної відповіді. У риб, амфібій і рептилій
відбувається синтез цитокіноподібних речовин.
Тромбування ран. Основними воротами проникнення мікроорганізмів у внутрішнє середовище
живих організмів є ушкодження цілісності покривів. Тому закриття (тромбування) ран є одним із
ефективних способів захисту від проникнення патогенів. Перша реакція організму безхребетних на
появу ран — скорочення м'язів навколо рани. Подальші реакції, пов'язані з закриттям ран, можуть
здійснюватися двома способами: або лише аглютинацією рухомих клітин, або аглютинацією
рухомих клітин і полімеризацією молекул міжклітинної рідини (коагуляція).
У кишковопорожнинних, кільчастих червів, молюсків, голкошкірих основним засобом тромбування
ран є крім скорочення м'язів індукування прискореного розмноження циркулюючих клітин, їхня
ефективна міграція до рани, агрегація, формування тромбу і закупорення рани. У голкошкірих і
покривників виявлено гуморальні фактори (цитокіноподібні), які беруть активну участь у залученні
клітин
;
що циркулюють, у місця ушкодження та їх агрегації, що сприяє швидкому утворенню
тромбів у рані.
Коагуляцію плазми в процесі загоювання ран досить детально описано у членистоногих. Джерелом
полімерних молекул є гранулярні амебоцити, які містять гранули з низькомолекулярними білками –
попередниками гелю і ферментами, що запускають їх полімеризацію. В деяких організмів до складу
гранул входять компоненти фенолоксидазної системи. Після виникнення певних сигналів
відбувається швидкий викид гранул та звільнення з них біологічно активних речовин, активація
ферментів, що призводить до полімеризації білків. Найефективнішим активатором гранулярних
амебоцитів є ЛПЦ клітинних стінок бактерій та 1,3-глюкан. У членистоногих у гранулах містяться
білки коагулін і фібриноген, які у процесі активації утворюють гель.
Інкапсуляція. У разі неспроможності поглинати великі чужорідні об'єкти, що потрапили всередину
організму, їх ізолюють за допомогою інкапсуляції один або кілька типів клітин. Механізм утворення
капсули подібний для всіх безхребетних. Так, у голкошкірих на поверхні чужорідного тіла
розпластуються фагоцити і формують багатошарову капсулу. Клітини, які щільно прилягають до
чужорідного тіла, часто руйнуються, внаслідок чого виділяються різні ферменти і бактерицидні
речовини, що призводить до руйнування інкапсульованого об'єкта. В тілі кільчастих червів,
молюсків, членистоногих нерідко виявляють так звані бурі тіла, які можуть існувати впродовж
усього життя і, очевидно, є інкапсульованими продуктами перетравлення чужорідних часточок. У
процесі взаємодії з чужорідними об'єктами фагоцити можуть виділяти речовини, які стимулюють
інші клітини та індукують хемотаксис.
У комах (членистоногі) у процесі інкапсуляції беруть участь два типи клітин. Гранулоцити
першими контактують із бактеріями, активуються і дегранулюються Вміст гранул зазнає
полімеризації та меланізації, що сприяє склеюванню бактерій і стимулює плазмоцити, які активно
аглютинуються й розпластуються на поверхні агрегатів, що призводить до утворення вузликів. У
деяких комах в інкапсуляції беруть активну участь кристалічні клітини, що містять фенолоксидазу в
неактивній формі, яка активується під час викиду вмісту клітин після їх контакту з чужорідними
об'єктами. В інших комах цю функцію виконують так звані тромбоцитоїди, які у процесі контакту з
чужорідними об'єктами розсипаються на маленькі фрагменти, подібно до мегакаріоцитів хребетних.
В інших комах, у гемолімфі яких міститься дуже мало клітин, відбувається утворення безклітинної
капсули — із полімеризованих фенолом білків.
14.2. ОРГАНИ ГЕМОЛІМФОМІЄЛОПОЕЗУ.
Уперше органи гемолімфопоезу з 'являються одночасно із замкненою кровоносною системою в
деяких видів кільчастих червів і розміщені у вигляді парних вузликів уздовж кишки. Тому на рівні
кільчастих червів імунна система поповнюється важливим еволюційним досягненням у вигляді
сконцентрованої лімфогемопоетичної тканини, де локально відбувається клітинне дозрівання.
Гемопоетичні утвори з'являються у більшості типів безхребетних і розміщуються в різних тканинах
і органах.
У молюсків цей утвір розміщений у целомі між перикардіумом та ниркою; при інфекціях він
збільшує продукування амебоцитів. Голкошкірі мають спеціальний орган — аксіальний, у якому
диференціюються фагоцити, лімфоцити, вібруючі та кристалічні клітини і надходять у судини
кровоносної системи. У покривників осередки гемопоезу виявлено в навкологлотковій ділянці, в
сполучній тканині кишечника і криптах целому. У нижчих хребетних важливим органом
кровотворення є нирки.
Перехід від водного до наземного способу існування став важливою віхою еволюції
гемолімфомієлопоезу. З одного боку, цей перехід зумовив формування трубчастих кінцівок і
відповідно кісткового мозку — в майбутньому основного джерела стовбурових клітин для
лімфопоезу, а з іншого, — стимулював подальший розвиток імунної системи завдяки широкому
контакту на суші з новими численними мікроорганізмами. В амфібій і рептилій у становленні клітин
імунної системи активну участь беруть кістковий мозок, печінка та ін. Вперше у рептилій
спостерігається відокремлення руху клітин по лімфатичних і кровоносних системах. У птахів
з'являються характерні лише для них імунно активні органи — сумка Фабриціуса і залоза Гарднера
та у весняно-літній період підшкірні лімфовузли з аферентними й еферентними судинами,
медулярною зоною та центрами розмноження.
Тимус або тимусподібний орган відсутній у всіх безхребетних. Він є лише у примітивних
хребетних і це одне з важливих явищ в еволюції імунної системи — з'являється специфічний орган,
де відбувається генералізація в онтогенезі формування Т-клітинного розвитку, що значно
підвищило ефективність функції всієї імунної системи.
Примітивні зачатки тимусподібних структур виявлено у представників класу круглоротих, крім
міксин, в яких вони відсутні. У більшості еволюційно більш досконалих представників круглоротих
— міног у ділянці зябрових щілин є лімфоїдна тканина, де відбувається місцеве скупчення
лімфоцитів з високим рівнем мітотичної активності, що дало змогу деяким авторам вважати його
прообразом тимуса.
У класі хрящових риб тимус уже диференційований на кіркову та медулярну зони і в деяких
підкласах зазнає вікової еволюції. У більшості представників класу кісткових риб та амфібій і
рептилій тимус розвивається з епітелію глоткових кишень. У кісткових риб тимус розміщений під
зябрами. Відмічається, хоча і не в усіх кісткових риб, наявність кіркової та медулярної зон тимуса,
але це виражено у них значно менше, ніж у ссавців. Для всіх риб характерна вікова еволюція
тимуса. В амфібій тимус формується в ембріональному періоді; у личинок утворюються кіркові та
медулярні зони. Для амфібій, як і для ссавців, характерний контакт лімфоцитів кори тимуса з НК,
макрофагами і дендритними клітинами та поява клітин, що нагадують клітини-няньки ссавців.
Тимус амфібій відіграє важливу роль в імуногенезі. Його відсутність призводить до послаблення
алотрансплантаційної реакції, пригнічує проліферацію лімфоцитів під час дії Т-клітинних мітогенів,
гальмує гуморальну імунну відповідь до Т-залежних антигенів. Тимус амфібій, подібно до тимуса
ссавців, здійснює позитивний відбір клонів тимоцитів, які розпізнають комплекс чужорідний
антиген — власні антигени МНС. Для рептилій характерне формування тимуса в анатомічній
близькості до прищитоподібної залози. Тимус рептилій зазнає вікової інволюції.
У птахів тимус знаходиться з обох боків шиї у вигляді парних семи структур, причому кожна частка
має кортикальну і медулярну зони. В тимусі птахів є субпопуляції Т-лімфоцитів з різними
функціями.
Селезінка. У круглоротих навколокишкова лімфоїдна тканина, очевидно, є попередником
селезінки. У хрящових риб у сполучній тканині кишечника є скупчення лімфомієлоїдної тканини з
кровотворною функцією. В стінці стравоходу між слизовою та м'язовою оболонками розміщений
орган Лейдіга, в якому здійснюється гранулоцитопоез. Селезінка з'являється у риб. У селезінці
амфібій продукуються всі типи клітин і вона є основним кровотворним органом.
Лімфовузли. У нижчих хребетних виявляються скупчення лімфоїдних клітин, подібних до
лімфовузлів ссавців, але зв'язані не з лімфатичною системою, а з кровоносною. У безхвостих
амфібій виявлено так звані лімфомієлоїдні органи — інкапсульовані скупчення лімфоїдної тканини,
місця знаходження яких в деяких видів різні. В цих органах відбувається накопичення антигенів,
здійснюється антигензалежне диференціювання лімфоцитів та фільтрування крові, але не лімфи.
У рептилій у стінках лімфатичних судин трапляються дифузні скупчення лімфоцитів, що нагадують
лімфовузли. У птахів виявлено дифузні скупчення лімфоцитів, окремі лімфоїдні фолікули та групи
фолікулів, які інколи називають неінкапсульованими лімфовузлами, але тільки у ссавців
формуються повноцінні лімфовузли.
Паралельно зі тановленням та вдосконаленням структурної організаії імунної системи формується
та вдосконалюється імунореактивність.
14.3. АДАПТИВНА ІМУННА ВІДПОВІДЬ.
Достовірні експериментальні дані про можливість формування специфічної імунної гуморальної
відповіді у безхребетних відсутні. Антигензв'язувальні речовинну голкошкірих за своєю структурою
відрізняються від імуноґлобулінів. Структура інших антигензв'язувальних речовин ще не вивчена. У
формуванні адаптивної імунної відповіді основну роль відіграють Т- і В-лімфоцити.
Еволюція Т-лімфоцитів. Клітини з маркером Thy-1 виявляються на ранніх етапах еволюційного
розвитку — вони з'являються у кільчастих червів і здатні незначною мірою відповідати на Т-
клітинні мітогени. Лімфоцити примітивних хребетних — круглоротих, хрящових і кісткових риб на
Т-клітинні мітогени реагують дуже слабко, потребують великої дози мітогенів і їхня проліферація
не досягає рівня проліферації лімфоцитів ссавців.
Реакція змішаної культури лімфоцитів, яка свідчить про наявність на лімфоцитах структур з Т-
залежною антигенрозпізнавальною функцією, виявляється у голкошкірих на видовому рівні, але
відсутня під час алогенної відповіді. Це, можливо, зумовлюється відсутністю певних
костимулювальних молекул. Однак у морських зірок виявлено поверхневий білок, який
гомологічний β-ланцюгу ТкР ссавців. У примітивних хребетних (круглоротих і хрящових риб)
розвивається слабка реакція у змішаній культурі лімфоцитів. Так, у риб формуються основні
популяції клітин імунної системи, характерні для ссавців. Близько 70 % лейкоцитів риб є
лімфоцитами. У костистих риб з'являються популяції Т-лімфоцитів з хелперною та супресорною
активностями. У прояві функціональної активності клітин імунної системи важливу роль відіграють
клітини мікрооточення. У костистих риб та амфібій розвивається виражена реакція в змішаній
культурі лімфоцитів а в рептилій реакція змішаної культури лімфоцитів подібна до такої у птахів та
ссавців, але має сезонні коливання. Лімфоцити риб і амфібій продукують цитокіни з ІЛ-1-, ІЛ-2-,
МІФ-подібною активністю.
Еволюція В-клітин. На відміну від Т-системи, окремі ознаки якої виявляються навіть в
одноклітинних та примітивних багатоклітинних організмах, В-система розвивається лише у
хребетних. У крові міксин виявлено близько 65 % клітин з поверхневим імуноґлобуліном —
характерною ознакою В-лімфоцитів. Однак серед них не виявлено функціонально завершеної
форми В-лімфоцитів — плазмоцитів, здатних продукувати антитіла. У міног, більш організованих
представників круглоротих, є орган В-лімфопоезу — жирове тіло (зародок кісткового мозку),
розміщене над хордою. У личинок цю функцію виконує тифлозоль — прообраз селезінки і нирки,
які в процесі метаморфозу резорбуються, а їхня функція переходить до жирового тіла. В тифлозолі
та жировому тілі містяться плазмоцитоподібні клітини, але синтезу антитіл не виявлено.
У хрящових риб В-залежними лімфоїдними органами є селезінка, нирки, орган Лейдіга. В
ембріогенезі, як і в усіх ссавців, першим органом, що продукує антитіла, є печінка. У кісткових риб
відсутні функціонально активний кістковий мозок, сумка Фабриціуса, лімфовузли, а основним
джерелом стовбурових клітин для формування лімфоїдних і мієлоїдних клітин та первинним
органом для розвитку В-клітин є нирки, а потім селезінка. В крові близько 20 % лімфоцитів
належить до В-лімфоцитів. В амфібій виявлено функціонуючий кістковий мозок, який стає органом
формування В-клітин та основним місцем антититілоутворення. У рептилій джерело В-лімфоцитів –
печінка і кістковий мозок, а антитіла утворюються переважно в селезінці.
Основним місцезнаходженням В-клітин у птахів є сумка Фабриціуса — до 75 % всix лімфоцитів.
Антитілоутворювальні клітини, крім сумки Фабриціуса, знаходяться в селезінці, сліпій кишці,
залозах Гардеріана. Домінуюче зосередження В-лімфоцитів у сумці Фабриціуса, очевидно,
зумовлюється специфічним медіатором — бурсином, який здатний відновлювати
антитілоутворення у птахів з видаленою сумкою. У птахів кількість В-лімфоцитів подібна до такої у
ссавців.
Еволюція суперродини імуноґлобулінів. При розгляді питання про еволюцію антитіл привертає
увагу кілька фактів: усі молекули представників суперродини імуноґлобулінів (антитіла, ТкР і ВкР,
антигени МНС І та МНС II класів, ряд молекул адгезії) мають доменну структуру (V- і С-домени) і
статистично достовірну гомологію амінокислотних залишків відомим імуноґлобулінам. Крім того,
деякі білки цієї суперродини мають однодоменну структуру (Thy-1 антиген, Ро-білок мієліну, 2-
мікроґлобулін). Структури, подібні до однодоменних білків суперродини імуноґлобулінів,
трапляються на найнижчих щаблях еволюційної драбини — у найпростіших і навіть у деяких
бактерій і дріжджів. Однодоменні білки на самому початку еволюційного розвитку функціонували
як міжклітинні ліганди, що контролювали міжклітинний контакт та мітотичну активність.
Збільшення маси органів і кількості тканиноспецифічних білків ускладнювали функцію білків
суперродини імуноґлобулінів, що зумовило виникнення їх різноманіття та значного підсилення
їхніх рецепторних функцій.
V- і С-домени, очевидно, утворилися з одного загального попередника, структура якого подібна до
антигену Thy-1 та до антигенів гістосумісності І та II класів хребетних. Найдавнішим є домен С2,
який, очевидно, еволюціонував у двох напрямах – С2-V2-V1 та С2-С1. Білки з доменами С2
виявлено у різних тканинах губок, молюсків, членистоногих, і вони подібні до NCAM, NgCAM, KIR
хребетних. Деякі з них (VDV молюсків, гемолін та пероксидазин комах) продукуються гемоцитами і
мають захисні функції. Низка білків з доменами типу V2 — білок молюсків FREP,
преципітувальний білок деяких паразитів та білок комах амальган з доменами V2 і С2 відношення
щодо захисних функцій не мають, але могли бути родоначальниками для Т-клітинного рецептора та
окремих імуноґлобулінів. У результаті дуплікації первинного гена та транслокаційних перебудов
гени V- і С-доменів утворилися на одній хромосомі, що зумовило виникнення багатодоменних
білків — гомо- і гетеродимерів з певною спеціалізацією, в яких V-домени сумісне розпізнають певні
ліганди. Первинними лігандами рецепторів суперродини імуноґлобулінів були різні аутологічні
білки, нащадки яких частково збереглися на лімфоцитах як допоміжні рецептори лімфоцитів (CD4,
CD8 та ін.), виявляються на попередниках Т- і В-лімфоцитів у процесі їх диференціювання та
входять де складу антигенрозпізнавальних структур примітивних Тγδ- і В1-лімфоцитів і складу
антитіл нижчих хребетних. Припускають, що предками Т-клітинних рецепторів та імуноґлобулінів
були рецептори-димери клітинних мембран, що складалися з V- і С-доменів і сформували
антигензв'я зувальні ділянки. Останні були здатн розпізнавати невеликі детермінанти – примітивні
антигени гістосумісності. Це підтверджується спорідненістю С-доменів імуноґлобулінів, Т-
клітинного рецептора, антигенів І і II класів гістосумісності одночасна їх поява у ході еволюції (в
хрящових риб) та особливостями диференціювання Т-клітин. Основна функція предків Т-клітинних
рецепторів та імуноґлобулінів — мічення «свого». С-домени Т-клітинних рецепторів та
імуноґлобулінів, очевидно, розвинулися одночасно з V-доменами, коли С- і V-домени складали вже
один поліпептидний ланцюг, і це простежується в голкошкірих, здатних до специфічного
алотрансплантаційного відторгнення.
Отже, найвірогіднішою основою для виникнення імунної системи хребетних є алоогенна
гістонесумісність у безхребетних, що контролювала алогенний паразитизм, мімікрію мічення
«свого» ксеногенними патогенами та інбридинг. У покривників виявлено інгібіторні рецептори до
аутологічних антигенів гістонесумісності й пускові рецептори до чужорідних антигенів
гістонесумісності. Подібну подвійну систему рецепторів до антигенів МНС І мають Т-лімфоцити і
природні кілери хребетних, що свідчить про можливу роль деяких структур гістонесумісності
безхребетних у формуванні певних ланок імунної системи хребетних. Виявлення у покривників
білків суперродини імуноґлобулінів з доменами V1 і С1 та характерної для МНС Іа та МНС ІІа
послідовності дали змогу припустити, що генетичний локус гістонесумісності безхребетних Fu/HC
має спорідненість із комплексом МНС. Можливо, трансформовані з Fu/HC антигени МНС І
спочатку представляли аутологічні пептиди ПК, а потім почали презентувати й чужорідні пептиди
Т-лімфоцитам. Тому у покривників трансплантаційні імунні реакції наблизилися до таких у
хребетних, тоді як гуморальні реакції покривників не вийшли за межі природного імунітету, але
значно ускладнилися.
Становлення імунної системи хребетних пов'язане насамперед зі зміною способу життя і загрозою
агресії різних патогенів. Ранні хребетні в результаті появи у них органів дихання, травлення,
кровообігу стали чутливішими до агресії патогенів. Розміщення лімфоїдних органів біля основних
можливих воріт інфекції має протективний характер. Механізми природної резистентності за такої
ситуації не могли ефективно захищати організм від мікроорганізмів, що сприяло формуванню на
базі певних структур природної резистентності покривників зачатків адаптивної імунної відповіді у
хребетних. Унаслідок дуплікації та диверсифікації зародкових V-генів і їх соматичних мутацій
з'явилося різноманіття рецепторів лімфоцитоподібних клітин, що зумовлює формування
самостійних клітинних ліній лімфоцитів. Важливу роль у закріпленні клонального різноманіття
лімфоцитів за принципом одна клітина — один рецептор та збільшення варіабельності самих
рецепторів відіграв механізм соматичного реаранжування V-генів за допомогою RAG-1 і RAG-2,
які, як вважають, виникли з вірусних транспозаз-інтеграз. У нових умовах різко зменшується
загроза алогенного паразитизму, але зростає вірогідність бактеріально-вірусної агресії, що сприяло
перетворенню антигенів гістонесумісності на поверхневі пептидопрезентувальні антигени МНС І і
МНС II класів, а їхніх рецепторів — на рецептори лімфоцитів. Таким чином, ТкР і ВкР утворилися
із рецепторів, які втратили зв'язок з антигенами гістонесумісності.
Перетворення за допомогою алельного поліморфізму мічення «свого» на антигени тканинної
несумісності зумовило синхронну перебудову імунної системи -виникнення різноманіття рецепторів
та спеціальних механізмів адаптації їх до індивідуальних наборів антигенів гістосумісності. Цьому
сприяли дуплікації та виникнення різних генів і поділ V-генів нітронами на V-, D- і J-сеґменти та
вторинне збирання цих кластерів. Цей процес сприяв експресії у кожній клітині лише одного
варіанта поліпептидного ланцюга, відповідно рецептора однієї специфічності і, як результат, —
формування клональної структури лімфоцитів.
У безхребетних розвивалися домени, які становлять основу розпізнавальних структур варіабельних і
константних доменів Т-клітинних рецепторів та константних доменів МНС. У хребетних, за
винятком ссавців, виникають домени імуноглобулінів та розподіл (дивергенція) початкових доменів
ТкР, МНС і легких ланцюгів на різні типи доменів (варіабельні й константні домени
імуноґлобулінів та ін.). У ссавців відбувається дивергенція доменів на класи й підкласи
імуноґлобулінів. Кількість V-генів хребетних збільшується у міру ускладнення організації
хребетних. Це виявляється у збільшенні ізотипів та афінності антитіл у процесі розвитку імунної
відповіді у представників, які стоять вище на філогенетичному щаблі еволюційної драбини.
У найпримітивніших хребетних — круглоротих — у крові та деяких органах виявляються
лімфоцито- та плазмоцитоподібні клітини. Кількість останніх після введення чужорідних
еритроцитів збільшується, і вони продукують антигензв'язувальні речовини, які не належать до
імуноґлобулінів. Вважають, що плазмоцити продукують антигензв'язувальні компоненти
комплементу. Однак ні генів імуноґлобулінів, ТкР, МНС І і II, ні антитіл у сироватці крові, ні білків,
що зумовлюють реаранжування V-генів та транспортування пептидів до антигенів МНС, ні імунної
пам'яті у круглоротих не виявлено.
Перші чітко визначені імуноґлобуліни виявлено в еволюційно більш розвинених хрящових риб. У
них спостерігається синтез специфічних антитіл з різними ізотипами — IgH і IgL, ТкР обох типів,
МНС І і МНС II класів, соматичне перегрупування V-генів. Імуноґлобуліни хрящових риб мають
ефективну специфічність і є двох типів — з високою (900 кД) і низькою (180 кД) молекулярною
масою. Імуноґлобуліни з високою молекулярною масою складаються з п'яти ідентичних мономерів,
сполучених дисульфідними зв'язками. Н-ланцюги низько- і високомолекулярних форм мають
подібний амінокислотний склад, серологічну специфічність, вміст вуглеводів і гомологічні за
амінокислотним складом з Н-ланцюгами IgM ссавців, їхні гени гомологічні за нуклеотидною
послідовністю. Високомолекулярна пентамерна форма імуноґлобуліну гомологічна IgM ссавців.
У костистих риб виділяють кілька видів молекулярних форм імуноґлобулінів, але для більшості їх
характерна наявність тетра- і мономерної форм IgM-подібного імуноґлобуліну. Н-ланцюг в окремих
видів риб має різну молекулярну масу. Імуноґлобуліни костистих риб, незважаючи на існування
двох їх форм та двох варіантів Н-ланцюгів, належать до класу IgM і гомологічні IgM ссавців. У
костистих риб уперше відбувається поділ L-ланцюгів на два типи за молекулярною масою — 22 —
24 і 26 кД (можливо, κ- та λ-типів). У деяких випадках імунна відповідь риб супроводжується
формуванням імунної пам'яті, але здебільшого у слабкій формі. У міру розвитку первинної і
вторинної імунної відповіді у риб афінність антитіл не змінюється і формується мало ізотипів
імуноґлобулінів. Це свідчить про малу кількість V-генів, що працюють, імуноґлобулінів і,
відповідно, незначну кількість предетермінованих антигенспецифічних В-клонів. Однак
спостерігається і значний стрибок у становленні імунної системи порівняно з круглоротими, і в
цьому поступі, як припускають, важливу, а, можливо, основну роль відіграє горизонтальне
перенесення генів ретровірусами, внаслідок чого вірусні ретрогази перетворилися на RAG-1 та
RAG-2 (ферменти рекомбінази, що зумовлюють складання V-генів імуноґлобулінів і ТкР).
Певне значення має становлення імунної системи у хвостатих і безхвостих амфібій. Хвостаті амфібії
постійно живуть у воді і їхня імунна система мало відрізняється від імунної системи костистих риб.
Безхвості амфібії в процесі еволюційного розвитку живої природи вперше виходять на сушу і
стикаються з великою кількістю патогенних мікроорганізмів. Це потребувало вдосконалення
захисних сил організму, що сприяло виникненню функціонуючого кісткового мозку — джерела В-
клітин, формуванню ефективної імунної пам'яті, появі механізмів підвищення афінності антитіл та
виникненню додаткових ізотипів імуноґлобулінів. Ці нові ефективні захисні фактори еволюційно
закріплялися й удосконалювалися в рептилій, птахів і ссавців. У амфібій переважно виявлено три
ізотипи імуноґлобулінів: у всіх видів — високомолекулярний пентамер (900 кД), аналогічний IgM
ссавців, а в деяких виявлено ще два ізотипи — мономер IgY (очевидно, аналог IgG) та гексамер IgX
з молекулярною масою 80 кД. Характерно, що до складу молекули IgY входять глікозильовані та
неглікозильовані Н-ланцюги.
У рептилій також виявлено три типи імуноґлобулінів — високомолекулярний пентамер (900 кД), що
серологічно ідентичний IgM ссавців, імуноґлобулін IgY з проміжною молекулярною масою (175 кД)
та імуноґлобулін з низькою молекулярною масою (120 кД) і низьким вмістом вуглеводів (IgN).
У птахів трапляється чотири типи імуноґлобулінів — високомолекулярний пентамер (900 кД), який
серологічно ідентичний IgM ссавців, імуноґлобулін IgY з молекулярною масою 175 кД та IgN —
можливий аналог секреторного IgA2 ссавців, який виявляється у різних секретах курей,
плазмоцитах слизових кишок і має подібність за складом та послідовністю амінокислот. У качок
виявлено IgN, який становить основну масу сироваткового імуноґлобуліну.
Мономерні імуноґлобуліни сироватки крові характеризуються опсонізувальною активністю і
вищою афінністю до антигенів порівняно з IgM. Їх поява у хребетних корелює з появою імунної
пам'яті. IgE виявлено лише у ссавців, починаючи з сумчастих.
Імунна пам'ять. Більшість експериментальних даних про існування імунної пам'яті у безхребетних
(переважно за наявністю трансплантаційного відторгнення) малодостовірні, оскільки здебільшого
грунтуються на окремих, не перевірених результатах. Слабка і недовготривала імунна пам'ять під
час алотрансплантації виявлена у губок, кільчастих червів, членистоногих, голкошкірих, можливо,
зумовлюється пролонгованою антигенною стимуляцією. У разі існування імунної пам'яті у
безхребетних виникає питання про механізм її формування та прояв, оскільки безхребетні не мають
ні ТкР, ні імуноглобулінів, ні клонованих за спорідненістю до певних антигенів лімфоцитів. У
найбільш організованих безхребетних — покривників під час алотрансплантації виникає тривала
(до 2 міс.) специфічна пам'ять із активацією гемопоезу. У примітивних хребетних — круглоротих
при імунізації імунної пам'яті не виявлено; у костистих риб виявлено лише деякі В-лімфоцити
пам'яті. У риб трансплантати розпізнаються білками, які продукуються МНС і відторгаються
цитотоксичними лімфоцитами, і формується пам'ять до трансплантатів.
14.4. РОЗПІЗНАВАННЯ «СВОГО» І «ЧУЖОГО».
Одним із найдавніших факторів прояву захисних реакцій є розпізнавання «свого» і «чужого», бо
тільки визначивши «чуже», можна запускати захисні реакції проти нього. Розпізнавання «свого» і
«чужого» характерне для всіх багатоклітинних організмів, у тому числі й примітивних, хоча
механізм його в останніх відомий недостатньо. Припускають, що розпізнавання зумовлюється
певними поверхневими рецепторами захисних клітин. Підтвердженням цього є дані про наявність
відповідних розпізнавальних молекул на поверхні гемоцитів та в гемолімфі. Так, на гемоцитах
кільчастих червів, голкошкірих виявляються рецептори до КонА, що має подібність до певних
рецепторів лімфоцитів хребетних, а на гранулярних гемоцитах деяких членистоногих виявлено
рецептори до ЛПЦ бактерій. Роль розпізнавальних молекул безхребетних можуть відігравати й різні
аглютиніни та протеази, що містяться в гранулах гемоцитів і виводяться в гемолімфу екзоцитозом.
Протеази можуть специфічно розпізнавати ЛПЦ бактерій і β-1-3-глікани грибів і запускати ряд
захисних реакцій. Опсоніни, в тому числі комплементоподібні білки, можуть сприяти
розпізнаванню «чужого». У деяких членистоногих (мечохвостів) як розпізнавальні молекули
виступає споріднена з С-реактивним білком речовина лімулін, що розпізнає цукрид, який входить