Смирнов В.М. Физиология человека: Учебник
Подождите немного. Документ загружается.


Подушка получает главные входы от ко-
ленчатых тел и неспецифических ядер тала-
муса. Эфферентные пути от нее идут в височ-
но-теменно-затылочные зоны коры, участ-
вующие в гностических (узнавание предме-
тов,
явлений), речевых и зрительных функ-
циях (например, в интеграции слова со зри-
тельным образом), а также в восприятии
«схемы тела». Электростимуляция подушки
доминантного полушария приводит к рече-
вым нарушениям типа аномии (нарушение
называния предметов), разрушение подушки
вызывает нарушение «схемы тела», устраняет
тяжелые боли.
Медиодорсальное ядро получает входы от
гипоталамуса, миндалины, гиппокампа, тала-
мических ядер, центрального серого вещест-
ва ствола. Проекция этого ядра распростра-
няется на ассоциативную лобную и лимби-
ческую кору (поля 12, 18). Оно участвует в
формировании эмоциональной и поведен-
ческой двигательной активности, а также,
возможно, в механизмах памяти. Разрушение
этих ядер устраняет у больных страх, тревогу,
напряженность, страдание от боли, но возни-
кает лобный синдром: снижение инициати-
вы,
безразличие, гипокинезия.
Латеральные ядра получают зрительную и
слуховую импульсацию от коленчатых тел и
соматосенсорную импульсацию от вентраль-
ного ядра. Интегрированная сенсорная ин-
формация от этих источников далее проеци-
руется в ассоциативную теменную кору и ис-
пользуется в ее функциях гнозиса, праксиса,
формирования схемы тела.
В.
Неспецифические ядра составляют эво-
люционно более древнюю часть таламуса,
включающую интраламинарную (внутриплас-
тинчатую)
ядерную группу: центральное сре-
динное ядро (п.СМ), парафасцикулярное
(n.Pf) и субпарафасцикулярное (n.sPf) ядра,
центральное латеральное (n.Cl) и медиальное
(п.Cm) ядра, парацентральное ядро (п.Рс).
Кроме того, к неспецифическим ядрам отно-
сят ретикулярное ядро (n.R). Эти ядра содер-
жат преимущественно мелкие многоотрост-
чатые нейроны и функционально рассматри-
ваются как производное ретикулярной фор-
мации ствола мозга.
Неспецифические ядра имеют многочис-
ленные входы как от других ядер таламуса,
так и внеталамические: по латеральному
спиноталамическому, спиноретикуло-тала-
мическому трактам, текто- и тегментотала-
мическому трактам, проводящим преимуще-
ственно болевую и температурную чувстви-
тельность. В неспецифические ядра посту-
пает непосредственно или через ретикуляр-
ную формацию также часть импульсации по
коллатералям от всех специфических сен-
сорных систем. Кроме того, в неспецифи-
ческие ядра поступает импульсация из мо-
торных центров ствола (красное ядро, чер-
ное вещество), ядер мозжечка (шатра, проб-
кообразного), от базальных ганглиев и гип-
покампа, а также от коры мозга, особенно
лобных долей. Неспецифические ядра
имеют эфферентные выходы на другие тала-
мические ядра, кору большого мозга как не-
посредственно, так и через ретикулярные
ядра, а также нисходящие пути к другим
структурам ствола мозга.
Благодаря этим связям неспецифические
ядра таламуса выступают в роли интегрирую-
щего посредника между стволом мозга и моз-
жечком, с одной стороны, и новой корой,
лимбической системой и базальными ган-
глиями, с другой стороны, объединяя их в
единый функциональный комплекс. На кору
мозга неспецифический таламус оказывает
преимущественно модулирующее влияние.
Разрушение неспецифических ядер не вызы-
вает грубых расстройств эмоций, восприятия,
сна и бодрствования, образования условных
рефлексов, а нарушает только тонкую регу-
лировку поведения. В связи с этим модули-
рующее влияние неспецифических ядер тала-
муса, обеспечивающее «плавную настройку»
высшей нервной деятельности, считается их
главной функцией.
8.3.2. ГИПОТАЛАМУС
Гипоталамус является вентральной частью
промежуточного мозга. Макроскопически он
включает в себя преоптическую область и об-
ласть перекреста зрительных нервов, серый
бугор и воронку, сосцевидные тела. Микро-
скопически в гипоталамусе выделяют, по
данным разных авторов, от 15 до 48 парных
ядер,
которые подразделяются на 3—5 групп.
Многие авторы выделяют в гипоталамусе
4 основные области, включающие в себя не-
сколько ядер (по О.Г.Баклаваджану, 1981):
• преоптическая область — медиальное и
латеральное преоптические ядра;
• передняя область — супрахиазматическое,
супраоптическое, паравентрикулярное и
переднее гипоталамическое ядра;
• средняя (или туберальная) область —
дор-
сомедиальное, вентромедиальное, аркуат-
ное (инфундибулярное) и латеральное ги-
поталамические ядра;
• задняя область — супрамамиллярное, пре-
мамиллярное, латеральное и медиальное
1
5*

мамиллярные ядра, заднее гипоталамичес-
кое и перифорниатное ядра, субталами-
ческое ядро Луиса.
Важной физиологической особенностью
гипоталамуса является высокая проницае-
мость его сосудов для различных веществ, в
том числе и для крупных полипептидов. Это
обусловливает большую чувствительность ги-
поталамуса к сдвигам во внутренней среде
организма и способность реагировать на ко-
лебания концентрации гуморальных веществ.
В гипоталамусе по сравнению с другими
структурами головного мозга имеются самая
мощная сеть капилляров (1100—2600 капил-
ляров/мм
2
) и самый большой уровень ло-
кального кровотока.
Ядра гипоталамуса образуют многочис-
ленные связи друг с другом (ассоциативные),
с парными одноименными ядрами противо-
положной стороны (комиссуральные), а
также с выше- и нижележащими структурами
ЦНС (проекционные). Главные афферент-
ные пути гипоталамуса идут от лимбической
системы, коры больших полушарий, базаль-
ных ганглиев и ретикулярной формации
ствола. Основные эфферентные пути гипота-
ламуса идут в ствол мозга — его ретикуляр-
ную формацию, моторные и вегетативные
центры, в вегетативные центры спинного
мозга, от мамиллярных тел к передним ядрам
таламуса и далее в лимбическую систему, от
супраоптического и паравентрикулярного
ядер к нейрогипофизу, от вентромедиального
и инфундибулярного ядер к аденогипофизу, а
также имеются эфферентные выходы к лоб-
ной коре и полосатому телу.
Гипоталамус является многофункцио-
нальной системой, обладающей широкими
регулирующими и интегрирующими влия-
ниями. Однако важнейшие функции гипота-
ламуса трудно соотнести с его отдельными
ядрами. Как правило, отдельно взятое ядро
имеет несколько функций, а отдельно взятая
функция локализуется в нескольких ядрах.
В связи с этим физиология гипоталамуса
рассматривается обычно в аспекте функцио-
нальной специфики его различных областей
и зон:
Гипоталамус является важнейшим цент-
ром интеграции вегетативных функций, регу-
ляции эндокринной системы, теплового ба-
ланса организма, цикла «бодрствование —
сон» и других биоритмов; велика его роль в
организации поведения (пищевого, полово-
го,
агрессивно-оборонительного), направ-
ленного на реализацию биологических по-
требностей.
8.4. МОЗЖЕЧОК
Мозжечок — отдел головного мозга, обра-
зующий вместе с мостом задний мозг. Со-
ставляя 10 % массы головного мозга, мозже-
чок включает в себя более половины всех
нейронов ЦНС. Это свидетельствует о боль-
ших возможностях обработки информации и
соответствует главной функции мозжечка как
органа координации и контроля сложных и
автоматизированных движений. В осущест-
влении этой функции важную роль играют
обширные связи мозжечка с другими отдела-
ми ЦНС и рецепторным аппаратом.
8.4.1.
ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ
И СВЯЗИ МОЗЖЕЧКА
Выделяют три структуры мозжечка, отражаю-
щие эволюцию его функций (по Дж. Джан-
сенду, А. Бродалу, 1954).
Древний мозжечок (архицеребеллум) состо-
ит из клочка и узелка (флоккулонодулярная
доля) и нижней части червя. Он гомологичен
мозжечку круглоротых, передвигающихся в
воде с помощью змеевидных движений тела.
Старый мозжечок (палеоцеребеллум)
включает в себя верхнюю часть червя и пара-
флоккулярный отдел. Он гомологичен моз-
жечку рыб, передвигающихся с помощью
плавников.
Новый мозжечок (неоцеребеллум) состоит
из полушарий и появляется у животных,
передвигающихся с помощью конечностей.
Межнейронные связи в коре мозжечка,
его афферентные входы и эфферентные вы-
ходы многочисленны. Грушевидные нейроны
(клетки Пуркинье), образующие средний
(ганглиозный) слой коры, являются главной
функциональной единицей. Ее структурной
основой являются многочисленные ветвя-
щиеся дендриты, на которых в одной клетке
может быть до 100 тыс. синапсов. Количест-
во клеток Пуркинье у человека, по разным
источникам, — от 7 до 30 млн. Они являются
единственными эфферентными нейронами
коры мозжечка и непосредственно связыва-
ют ее с внутримозжечковыми и вестибуляр-
ными ядрами. В связи с этим функциональ-
ное влияние мозжечка существенным обра-
зом зависит от активности клеток Пуркинье,
что в свою очередь связано с афферентными
входами этих клеток. Главными мозжечковы-
ми афферентами являются системы лиано-
видных и моховидных волокон. В последнее
время изучается и третья система — моноа-
минергические афференты.
132

Основным источником лиановидных
волокон являются нейроны нижней
оливы продолговатого мозга. Информация к
ним поступает от мышечных, кожных рецеп-
торов и нейронов основания задних рогов
спинного мозга (по спинооливному пути), а
также от двигательной коры головного мозга.
Каждое лиановидное волокно устанавливает
синаптические контакты с дендритами (ме-
диатор аспартат), как правило, одной клетки
Пуркинье, но число их может достигать трех-
сот, что объясняет исключительно сильное
возбуждающее действие этого афферентного
входа (большой ВПСП, на вершине которого
формируется 3—5 потенциалов действия).
Кроме того, они оказывают на них и тормоз-
ное (более слабое) влияние через корзинча-
тые и звездчатые клетки поверхностного (мо-
лекулярного) слоя коры мозжечка.
По моховидным волокнам в
кору мозжечка поступает информация от
коры большого мозга (преимущественно ас-
социативной, по корково-мостомозжечково-
му пути), проприорецепторов опорно-двига-
тельного аппарата (по спинно-мозжечковым
путям), вестибулярных рецепторов и ретику-
лярной формации. Моховидные волокна об-
разуют возбуждающие синапсы на дендритах
клеток-зерен внутреннего (гранулярного)
слоя коры. Через аксоны клеток-зерен этот
афферентный вход оказывает непосредствен-
ное возбуждающее влияние (медиатор — глу-
тамат) на тормозные клетки Пуркинье и
опосредованно (через корзинчатые и звездча-
тые клетки) тормозное действие на клетки
Пуркинье. Активность клеток-зерен регули-
руется через тормозные клетки Гольджи
внутреннего слоя по типу возвратного тормо-
жения (медиатор —
ГАМ
К).
Третью, афферентную (моноаминергичес-
кую)
систему мозжечка начали изучать в пос-
леднее время. Через эти пути передается вли-
яние на мозжечок некоторых структур ствола
головного мозга, например голубого пятна и
ядер шва. Нейроны голубого пятна осущест-
вляют влияние на мозжечок через медиатор
норадреналин, а ядер шва — через серото-
нин. Стимуляция этих ядер тормозит фоно-
вую активность клеток Пуркинье и модули-
рует их ответы на активацию моховидных и
лиановидных афферентов. Как прямое (через
лиановидные волокна), так и опосредован-
ное (через моховидные волокна и клетки-
зерна) афферентное влияние на клетки Пур-
кинье является возбуждающим.
Но поскольку клетки Пуркинье являются
тормозными нейронами (медиатор ГАМК),
то с их помощью кора мозжечка превращает
возбуждающие сигналы на входе в тормозные
сигналы на выходе. Таким образом, эффе-
рентное влияние коры мозжечка на после-
дующее нейронное звено (в основном это
внутримозжечковые ядра) осуществляется не
по механизму запуска нейронной активнос-
ти,
а по механизму более или менее сильного
сдерживания этой активности, т.е. в мозжеч-
ке доминирует тормозный характер управле-
ния [Фанарджян В.В., 1992].
При изучении связей коры мозжечка с его
ядрами были выделены три симметричные
вертикальные зоны. Клетки Пуркинье меди-
альной червячной зоны и флоккулонодулярной
доли проецируются на ядра шатра. Нейроны
этих ядер связаны с моторными центрами
ствола, от которых к спинальным моторным
центрам идут вестибуло-, рубро- и ретику-
лоспинальные тракты, регулирующие тонус
мышц. Клетки Пуркинье промежуточной
зоны мозжечка проецируются на промежуточ-
ные (шаровидное и пробковидное) ядра. Ак-
соны нейронов этих ядер идут к красному
ядру среднего мозга, от которого начинается
руброспинальный тракт, стимулирующий
через спинальные моторные центры тонус
мышц-сгибателей.
Есть путь от промежуточного ядра к вент-
ролатеральному ядру таламуса и оттуда к дви-
гательной коре. Клетки Пуркинье латераль-
ной зоны коры проецируются на зубчатые
ядра мозжечка, от которых мощные тракты
направляются к вентролатеральному ядру та-
ламуса, нейроны которого в свою очередь
проецируются на моторные зоны коры боль-
шого мозга. Итак, три парных ядра мозжечка
являются главным эфферентным выходом
мозжечка на стволовые и корковые моторные
центры. Нейроны этих ядер имеют высокую
фоновую активность (50—100 имп/с). Они
находятся под тормозным влиянием клеток
Пуркинье и возбуждающим влиянием аффе-
рентных входов, которые по коллатеральным
ответвлениям поступают в эти ядра.
8.4.2. ФУНКЦИИ МОЗЖЕЧКА
Функции мозжечка формируют три главных
его влияния на организм [Орбели Л.А., 1940]:
на двигательный аппарат, афферентные сис-
темы и вегетативную нервную систему.
А. Двигательные функции мозжечка заклю-
чаются в регуляции мышечного тонуса, позы
и равновесия (1), координации позы и вы-
полняемого целенаправленного движения
(2),
программировании целенаправленных
движений (3).
133

1.
Регуляция мышечного тонуса, позы и
равновесия осуществляется преимуществен-
но древним мозжечком (флоккулонодуляр-
ная доля) и частично старым мозжечком,
входящими в медиальную червячную зону.
Получая и обрабатывая импульсацию от вес-
тибулярных рецепторов (по вестибуломоз-
жечковому пути), от проприорецепторов ап-
парата движения и рецепторов кожи (по
спинно-мозжечковым путям), от зрительных
и слуховых рецепторов (через четверохолмие
по тектоцеребеллярным путям), мозжечок
способен оценить состояние мышц, положе-
ние тела в пространстве и через ядра шатра,
используя вестибуло-, ретикуло- и руброс-
пинальный тракты, произвести перераспре-
деление мышечного тонуса, изменить позу
тела и сохранить равновесие. Нарушение
равновесия является наиболее характерным
симптомом поражения архицеребеллума. Из
связей древнего мозжечка со структурами
ствола мозга наиболее выражены его двусто-
ронние связи с вестибулярными ядрами.
Мозжечок осуществляет тонкую настройку
вестибулярных рефлексов, в том числе реф-
лекторное поддержание антигравитацион-
ной (вертикальной) позы. Функцию древне-
го мозжечка врач оценивает, определяя рав-
новесие у пациента в позе Ромберга: верти-
кальная поза со сдвинутыми ступнями и за-
крытыми глазами, вытянутыми горизонталь-
но (вперед) руками.
2.
Координация позы и выполняемого целе-
направленного движения осуществляется ста-
рым и новым мозжечком, входящими в про-
межуточную (околочервячную) зону. В кору
этой части мозжечка поступает импульсация
от рецепторов аппарата движения, а также
импульсация от моторной коры (программа
произвольного движения). Анализируя ин-
формацию о программе движения (из мотор-
ной коры) и выполнении движения (от про-
приорецепторов), мозжечок способен через
свое промежуточное ядро, имеющее выходы
на красное ядро и моторную кору, осущест-
вить координацию позы и выполняемого це-
ленаправленного движения в пространстве, а
также исправить направление движения. Об
этом свидетельствует то, что нейроны про-
межуточного ядра продолжают генерировать
импульсы в течение всего времени выполне-
ния движения. Нарушение координации дви-
жения (атаксия) является наиболее характер-
ным симптомом нарушения функции про-
межуточной зоны мозжечка. Эта функция
мозжечка может быть исследована, напри-
мер,
пальценосовой или пяточно-коленной
пробами.
3.
Участие в программировании целена-
правленных движений осуществляется новым
мозжечком
—
той частью полушарий мозжеч-
ка, которая входит в латеральную зону. Кора
этой части мозжечка получает импульсацию
преимущественно из ассоциативных зон
коры большого мозга через ядра моста. Эта
информация характеризует замысел движе-
ния. В коре нового мозжечка (а также в ба-
зальных ганглиях) она перерабатывается в
программу движения, которая через зубчатое
ядро мозжечка и вентральное латеральное
ядро таламуса попадает в кору, обрабатывает-
ся в премоторной и моторной коре большого
мозга и через пирамидную, экстрапирамид-
ную системы осуществляется как сложное
целенаправленное движение. Контроль и
коррекция более медленных программиро-
ванных движений осуществляется мозжечком
на основе обратной афферентации преиму-
щественно от проприорецепторов, а также от
вестибулярных зрительных, тактильных ре-
цепторов. Коррекция быстрых (баллистичес-
ких) движений из-за малого времени их вы-
полнения осуществляется по другому меха-
низму. Если не достигается результат, кор-
рекция движений осуществляется путем из-
менения их программы в латеральном моз-
жечке, т.е. на основе обучения и предшеству-
ющего опыта. Об этом говорит и прекраще-
ние импульсации в зубчатых ядрах мозжечка
сразу после начала движения. К таким дви-
жениям относятся многие спортивные дви-
жения (например, бросок мяча), игра на му-
зыкальных инструментах, «слепой» метод пе-
чатания и др. Врач может оценить эту функ-
цию мозжечка пробой с адиадохокинезом
(например, быстрой сменой ротационных
движений рук), при которой необходимо бы-
стро менять программу движения.
4.
Наряду с этим полушария мозжечка
осуществляют функцию
инициации
движения.
Было показано, что изменение активности
нейронов мозжечка (зубчатое и промежуточ-
ное ядра, клетки Пуркинье) на 0,1—0,3 с
предшествует началу движения, а охлаждение
зубчатого ядра задерживает на 0,1 с актива-
цию нейронов моторной коры и начало дви-
жения. Эти данные объясняют затруднения
вызова движений, которые испытывают
больные с поражением мозжечка. Вместе с
тем основные симптомы нарушения функ-
ций мозжечка в большей или меньшей степе-
ни выявляются при поражении каждого из
трех отделов мозжечка, что свидетельствует
об их функциональном перекрытии.
Из изложенного выше видно, что нейроны
мозжечка не имеют прямого выхода на спи-
134

нальные мотонеироны, а действуют на них
через корково-стволовые моторные центры.
С этим, вероятно, связана высокая степень
пластичности головного мозга по компенса-
ции нарушенных функций мозжечка. Извест-
ны случаи врожденного отсутствия мозжечка
или медленного разрушения его опухолью,
при которых у человека не определялись
симптомы нарушения движения.
Б.
Афферентная функция мозжечка. Выше
изложены в основном афферентные связи
мозжечка с проприорецепторами, кожными
и вестибулярными рецепторами. Известно
мнение Ч. Шеррингтона о мозжечке как
«главном ганглии» проприоцептивной чувст-
вительности. Однако, как показали электро-
физиологические исследования, изменения
активности клеток Пуркинье возникали при
стимуляции практически всех рецепторов —
зрительных, слуховых, висцеральных и др.
Вместе с тем исследования Л.А. Орбели лю-
дей с ранениями мозжечка показали измене-
ния пороговых величин различных видов
чувствительности. Эти данные свидетельст-
вуют о сложных двусторонних связях моз-
жечка и сенсорных систем. В реализации
влияния мозжечка на афферентные системы
организма, без сомнения, большую роль иг-
рают проекции ядер мозжечка на специфи-
ческие и неспецифические ядра таламуса как
главного центра переключения в сенсорных
системах. Можно предположить, что меха-
низмы влияния мозжечка на сенсорные
функции связаны с его влиянием на эффе-
рентный контроль активности рецепторного
аппарата и центров переключения в сенсор-
ных системах.
В последнее время исследуется участие
мозжечка в процессах мышления. Обнаруже-
но,
что при мыслительных операциях (на-
пример, арифметическом счете) увеличивает-
ся локальный кровоток не только в ассоциа-
тивной коре, но и в полушариях мозжечка.
Локальная биоэлектрическая активность в
полушариях мозжечка была отмечена в про-
цессах слышания и видения. Предполагается,
что мысль и движение контролируются одни-
ми и теми же нейронными системами [Ito
М., 1993].
В.
Роль мозжечка в регуляции вегетатив-
ных функций изложена в разделе 9.9.
8.5.
ЛИМБИЧЕСКАЯ СИСТЕМА
Термин «лимбическая система» (limbus —
край) был предложен П. Мак-Лином (1952) и
отражает особенность расположения ее в ви-
де кольца на границе новой коры, отделяю-
щей ее от ствола мозга. Под лимбической си-
стемой понимают функциональное объеди-
нение различных структур конечного, про-
межуточного и среднего мозга, обеспечиваю-
щее эмоционально-мотивационные компо-
ненты поведения и интеграцию висцераль-
ных функций организма. В эволюционном
аспекте лимбическая система сформировалась
в процессе усложнения форм поведения ор-
ганизма, перехода от жестких, генетически
запрограммированных форм поведения к
пластичным, основанным на обучении и па-
мяти.
8.5.1.
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ
ОРГАНИЗАЦИЯ ЛИМБИЧЕСКОЙ СИСТЕМЦ
В более узком понимании в лимбическую
систему включают образования древней коры
(обонятельная луковица и бугорок, периа-
мигдалярная и препириформная кора), ста-
рой коры (гиппокамп, зубчатая и поясная из-
вилины), подкорковые ядра (миндалина,
ядра перегородки). По отношению к гипота-
ламусу и ретикулярной формации ствола этот
комплекс рассматривается как более высо-
кий уровень интеграции вегетативных функ-
ций. В настоящее время преобладает понима-
ние лимбической системы в более широком
плане: кроме вышеназванных структур, в нее
также включают зоны новой коры лобной и
височной долей, гипоталамус и РФ среднего
мозга.
Важнейшие афферентные входы
в лимбическую систему осуществляются от
различных областей головного мозга, а также
через гипоталамус от РФ ствола, которая счи-
тается главным источником ее возбуждения.
Важным стимулирующим афферентным вхо-
дом являются также импульсы от обонятель-
ных рецепторов, поступающие по волокнам
обонятельного нерва (п. olfactorii, I) в структу-
ры лимбической системы — обонятельные лу-
ковицу и бугорок, параобонятельную зону
Брока, препириформную кору, миндалину.
В лимбической системе, таким образом, нахо-
дится корковый отдел древнего — обонятель-
ного — анализатора. Главные эфферент-
ные выходы из лимбической системы
осуществляются через гипоталамус (особенно
его мамиллярные тела) на ниже лежащие веге-
тативные и соматические центры ствола и
спинного мозга. Другой эфферентный выход
проводит возбуждение из лимбической систе-
мы в новую кору (преимущественно ассоциа-
тивную). Через нее лимбическая система
135
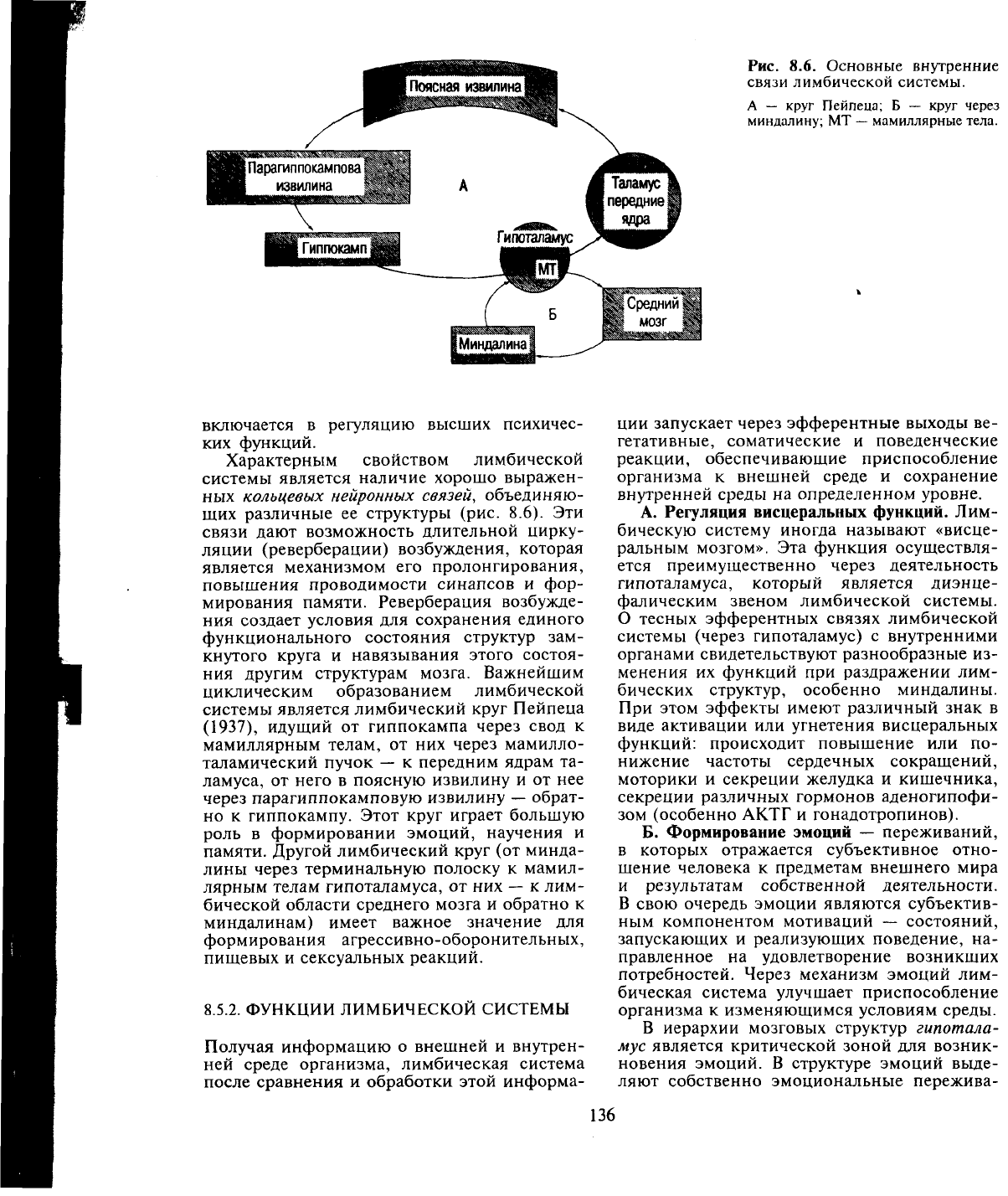
Рис.
8.6. Основные внутренние
связи лимбической системы.
А — круг Пейпеца; Б — круг через
миндалину; МТ
—
мамиллярные тела.
включается в регуляцию высших психичес-
ких функций.
Характерным свойством лимбической
системы является наличие хорошо выражен-
ных кольцевых нейронных связей, объединяю-
щих различные ее структуры (рис. 8.6). Эти
связи дают возможность длительной цирку-
ляции (реверберации) возбуждения, которая
является механизмом его пролонгирования,
повышения проводимости синапсов и фор-
мирования памяти. Реверберация возбужде-
ния создает условия для сохранения единого
функционального состояния структур зам-
кнутого круга и навязывания этого состоя-
ния другим структурам мозга. Важнейшим
циклическим образованием лимбической
системы является лимбический круг Пейпеца
(1937),
идущий от гиппокампа через свод к
мамиллярным телам, от них через мамилло-
таламический пучок — к передним ядрам та-
ламуса, от него в поясную извилину и от нее
через парагиппокамповую извилину
—
обрат-
но к гиппокампу. Этот круг играет большую
роль в формировании эмоций, научения и
памяти. Другой лимбический круг (от минда-
лины через терминальную полоску к мамил-
лярным телам гипоталамуса, от них
—
к лим-
бической области среднего мозга и обратно к
миндалинам) имеет важное значение для
формирования агрессивно-оборонительных,
пищевых и сексуальных реакций.
8.5.2. ФУНКЦИИ ЛИМБИЧЕСКОЙ СИСТЕМЫ
Получая информацию о внешней и внутрен-
ней среде организма, лимбическая система
после сравнения и обработки этой информа-
ции запускает через эфферентные выходы ве-
гетативные, соматические и поведенческие
реакции, обеспечивающие приспособление
организма к внешней среде и сохранение
внутренней среды на определенном уровне.
А. Регуляция висцеральных функций. Лим-
бическую систему иногда называют «висце-
ральным мозгом». Эта функция осуществля-
ется преимущественно через деятельность
гипоталамуса, который является диэнце-
фалическим звеном лимбической системы.
О тесных эфферентных связях лимбической
системы (через гипоталамус) с внутренними
органами свидетельствуют разнообразные из-
менения их функций при раздражении лим-
бических структур, особенно миндалины.
При этом эффекты имеют различный знак в
виде активации или угнетения висцеральных
функций: происходит повышение или по-
нижение частоты сердечных сокращений,
моторики и секреции желудка и кишечника,
секреции различных гормонов аденогипофи-
зом (особенно АКТГ и гонадотропинов).
Б.
Формирование эмоций — переживаний,
в которых отражается субъективное отно-
шение человека к предметам внешнего мира
и результатам собственной деятельности.
В свою очередь эмоции являются субъектив-
ным компонентом мотиваций — состояний,
запускающих и реализующих поведение, на-
правленное на удовлетворение возникших
потребностей. Через механизм эмоций лим-
бическая система улучшает приспособление
организма к изменяющимся условиям среды.
В иерархии мозговых структур гипотала-
мус является критической зоной для возник-
новения эмоций. В структуре эмоций выде-
ляют собственно эмоциональные пережива-
136

ния и его периферические (вегетативные и
соматические) проявления. Эти компоненты
эмоций могут иметь относительную самосто-
ятельность в проявлениях: выраженные субъ-
ективные переживания могут сопровождать-
ся небольшими периферическими проявле-
ниями, и, напротив, при псевдоэмоциях
яркие периферические проявления (напри-
мер,
мимические и вегетативные реакции
при плаче или смехе) могут протекать без су-
щественных субъективных переживаний. Ги-
поталамус в этом плане представляется
структурой, ответственной преимущественно
за вегетативные проявления эмоций.
Кроме гипоталамуса, к структурам лимби-
ческой системы, наиболее тесно связанным с
эмоциями, принадлежат поясная извилина и
миндалина. Электрическая стимуляция мин-
далины у человека вызывает преимущест-
венно отрицательные эмоции — страх, гнев,
ярость. Напротив, двустороннее удаление
миндалин в эксперименте на обезьянах резко
снижает их агрессивность, повышает тревож-
ность, неуверенность в себе. У таких живот-
ных нарушается способность оценивать ин-
формацию (особенно зрительную и слухо-
вую),
поступающую из окружающей среды, и
связывать эту информацию со своим эмоцио-
нальным состоянием. В результате нарушает-
ся нормальное взаимодействие организма с
окружающей средой, в том числе зоосоци-
альные отношения с другими особями в
группе. Наряду с этим миндалина участвует в
процессе сравнения конкурирующих мотива-
ций, выделения из них доминирующей моти-
вации и, следовательно, влияет на выбор по-
ведения. Поясная извилина, имею-
щая многочисленные связи как с новой
корой, так и со стволовыми центрами, вы-
полняет, по мнению некоторых авторов, роль
главного интегратора различных систем моз-
га, участвующих в формировании эмоций.
Важную роль в регуляции эмоций играет
вентральная лобная кора, имею-
щая хорошо выраженные связи с миндали-
ной. Поражение лобной коры вызывает рез-
кие нарушения эмоций у человека. Характер-
ным является возникновение эмоциональной
тупости. В первую очередь нарушаются эмо-
ции, связанные с социальными отношения-
ми людей, творчеством, и растормаживаются
эмоции, связанные с удовлетворением био-
логических потребностей.
В.
Участие в формировании памяти и обу-
чения. Эта функция преимущественно связа-
на с основным лимбическим кругом Пейпеца
(см.
рис. 8.6). Вместе с тем в одной из форм
обучения (однократном обучении) важную
роль играет миндалина благодаря ее свойству
индуцировать сильные отрицательные эмо-
ции, что способствует быстрому и прочному
формированию временной связи.
Среди структур лимбической системы, от-
ветственных за память и обучение, весьма
важную роль играют гиппокамп и свя-
занные с ним задние зоны лобной коры. Их
деятельность необходима для консолидации
памяти — перехода кратковременной памяти
в долговременную. Повреждение гиппокампа
у человека вызывает резкое нарушение усвое-
ния новой информации, образования долго-
временной памяти.
Электрофизиологической особенностью
гиппокампа является то, что в ответ на сен-
сорное раздражение, стимуляцию ретикуляр-
ной формации и заднего гипоталамуса в гип-
покампе развивается синхронизация элект-
рической активности в виде тета-ритма (4—
7 Гц). При этом в новой коре, напротив, воз-
никает десинхронизация в виде бета-ритма
(14—30 Гц). Пейсмекером тета-ритма счита-
ют перегородку — ее медиальное ядро. Во-
прос о поведенческих проявлениях гиппо-
кампального тета-ритма до сих пор не решен.
Его считают индикатором участия гиппокам-
па в ориентировочных рефлексах, реакциях
внимания, настороженности, развитии эмо-
ционального напряжения (ритм напряжения,
по П.К. Анохину). Однако большинство ис-
следователей считают, что он связан с обра-
боткой мозгом информации и организацией
памяти. Другой электрофизиологической
особенностью гиппокампа является его уни-
кальная способность на стимуляцию отвечать
длительной (в течение часов, дней и даже не-
дель) посттетанической потенциацией, кото-
рая приводит к облегчению синаптической
передачи и является основой формирования
памяти. Ультраструктурным проявлением
участия гиппокампа в процессах памяти яв-
ляется увеличение числа шипиков на дендри-
тах его пирамидных нейронов, что свидетель-
ствует об усилении синаптической передачи
возбуждающих и тормозных влияний.
8.6. БАЗАЛЬНЫЕ ГАНГЛИИ
Базальные ганглии — совокупность располо-
женных в основании больших полушарий
трех парных образований: бледного шара
(pallidum), филогенетически более позднего
образования — полосатого тела (striatum) и
наиболее молодой части — ограды (claus-
trum).
Бледный шар состоит из наружного и
внутреннего сегментов; полосатое тело пред-
137

ставлено хвостатым ядром (п. caudatus) и
скорлупой (putamen). Ограда расположена
между скорлупой и островковой корой. В
функциональном отношении в базальные
ганглии включают некоторые структуры про-
межуточного и среднего мозга: субталамичес-
кое ядро и черное вещество (substantia nigra).
8.6.1.
ФУНКЦИОНАЛЬНЫЕ СВЯЗИ
БАЗАЛЬНЫХ ГАНГЛИЕВ
Афферентная импульсация поступает преиму-
щественно в полосатое тело в основном из
трех источников: от всех областей коры не-
посредственно и через таламус; от неспеци-
фических интраламинарных ядер таламуса;
от черного вещества. Среди эфферентных свя-
зей базальных ганглиев можно отметить три
выхода:
• от полосатого тела пути направляются к
бледному шару; от бледного шара начина-
ется самый важный эфферентный тракт
базальных ганглиев, идущий в таламус, в
его релейные вентральные ядра, от них
возбуждающий путь следует в двигатель-
ную кору (поля 4 и 6);
• часть эфферентных волокон из бледного
шара и полосатого тела идет к центрам
"ствола мозга (ретикулярная формация,
красное ядро и далее — в спинной мозг), а
также через нижнюю оливу в мозжечок;
• от полосатого тела тормозящие пути идут
к черному веществу и после переключе-
ния — к ядрам таламуса.
Оценивая связи базальных ганглиев в
целом, можно отметить, что эта структура яв-
ляется промежуточным звеном (станцией
переключения), связывающей ассоциатив-
ную и, частично, сенсорную кору с двига-
тельной корой.
В структуре связей базальных ганглиев вы-
деляют несколько параллельно действующих
функциональных петель, соединя-
ющих базальные ганглии и кору больших по-
лушарий [Александер Г. и др., 1986].
Скелетомоторная петля соединяет премо-
торную, первичную моторную и соматосен-
сорную области коры со скорлупой, импуль-
сация из которой идет в бледный шар, черное
вещество и далее через вентролатеральное
ядро таламуса возвращается в премоторную
область коры (поле 6). Считают, что эта
петля служит для регуляции таких парамет-
ров движения, как амплитуда, сила, направ-
ление. При этом путь через черное вещество
регулирует сокращение мышц лица.
Глазодвигательная петля соединяет облас-
ти коры, контролирующие направление
взгляда (поле 8 лобной коры и поле 7 темен-
ной коры), с хвостатым ядром базальных ган-
глиев. Оттуда импульсация поступает в блед-
ный шар и черное вещество, из которых она
проецируется соответственно в ассоциатив-
ное медиодорсальное и релейное вентральное
переднее ядра таламуса, а из них возвращает-
ся в лобное глазодвигательное поле 8. Эта
петля участвует в регуляции, например, скач-
кообразных движений глаз (саккад). Предпо-
лагается существование также сложных пе-
тель,
по которым импульсация из лобных ас-
социативных зон коры поступает в структуры
базальных ганглиев (хвостатое ядро, бледный
шар,
черное вещество) и через медиодорсаль-
ное и вентральное переднее ядра таламуса
возвращается в ассоциативную лобную кору.
Считают, что эти петли участвуют в регуля-
ции высших психических функций мозга:
контроля мотиваций, прогнозирования, по-
знавательной (когнитивной) деятельности
(см.
Таламолобная система в разделе 8.7.3).
8.6.2. ФУНКЦИИ БАЗАЛЬНЫХ ГАНГЛИЕВ
Полосатое тело. Основными объектами
функционального влияния полосатого тела
являются бледный шар, черное вещество, та-
ламус и моторная кора.
1.
Влияние полосатого тела на бледный
шар осуществляется преимущественно через
тонкие тормозные волокна (медиатор
ГАМК). Многие нейроны бледного шара
дают смешанные ответы (ВПСП и ТПСП), а
некоторые нейроны отвечают только ВПСП.
Эти данные свидетельствуют о том, что поло-
сатое тело оказывает на бледный шар двоя-
кое действие — тормозящее и возбуждающее
с преобладанием тормозящего.
2.
Влияние на черное вещество. Между
черным веществом и полосатым телом име-
ются двусторонние связи. Нейроны полоса-
того тела оказывают тормозящее влияние
(медиатор ГАМК) на нейроны черного веще-
ства. В свою очередь нейроны черного веще-
ства модулируют влияние (медиатор—дофа-
мин) на кортико-стриарные каналы связи.
До сих пор окончательно не установлен ха-
рактер этого влияния — тормозной, возбуж-
дающий или тот и другой. Дофаминергичес-
ким путям в базальных ганглиях принадле-
жит важная роль в связи с двигательными
расстройствами при паркинсонизме, при ко-
тором резко падает концентрация дофамина
в обоих ядрах полосатого тела — хвостатом и
138

скорлупе. Кроме влияния на полосатое тело,
черное вещество оказывает тормозящее дей-
ствие на нейроны таламуса (медиатор ГАМК)
и получает возбуждающие афферентные
входы от субталамического ядра.
3.
Влияние
на таламус. Раздражение участ-
ков таламуса вызывает синхронизацию элек-
троэнцефалограммы: появление в ней высо-
коамплитудных ритмов, характерных для фа-
зы медленного сна. Эти ритмы можно вызвать
раздражением не только таламуса, но и поло-
сатого тела. При этом из всех структур (черное
вещество, миндалина, бледный шар), посыла-
ющих прямые пути к таламусу, только раздра-
жение полосатого тела вызывает появление в
коре высокоамплитудных медленных ритмов.
Напротив, разрушение полосатого тела нару-
шает эту реакцию и уменьшает время сна в
цикле сон—бодрствование.
4.
Влияние
на моторную кору. Прямая сти-
муляция полосатого тела через хронически
вживленные электроды вызывает относи-
тельно простые двигательные реакции: пово-
рот головы и туловища в сторону, противо-
положную раздражению, иногда сгибание ко-
нечности на противоположной стороне. Сти-
муляция некоторых зон полосатого тела вы-
зывает задержку текущей поведенческой дея-
тельности — двигательной, ориентировоч-
ной, пищедобывательной. Животное как бы
«застывает» в одной позе. При этом на
электроэнцефалограмме развиваются мед-
ленные высокоамплитудные ритмы. Раздра-
жение других точек полосатого тела приводит
к подавлению ощущения боли.
Поражение полосатого тела
(хвостатого ядра) дает достаточно характер-
ную клиническую картину, в которой главны-
ми являются насильственные избыточные
движения (гиперкинезы). Больной как бы не
может справиться со своей мускулатурой. Эти
гиперкинезы обозначаются как хорея и атетоз.
При повреждении в эксперименте полосатого
тела у млекопитающих стабильно развивается
синдром гиперактивности: число бесцельных
движений в пространстве увеличивается в 5—
7 раз. Опыты с разрушением полосатого тела
показали его важную роль в организации по-
ведения: животные с удаленной корой, но ин-
тактными хвостатыми ядрами в упрощенном
виде могут осуществлять некоторые формы
поведения — передвижение в пространстве,
поиск пищи, сохраняются ранее выработан-
ные простые условные рефлексы. Дополни-
тельное разрушение хвостатых ядер прекра-
щает эти формы поведения.
Бледный шар. Получая из полосатого тела
преимущественно тормозное и частично воз-
буждающее влияние, бледный шар оказывает
модулирующее влияние на двигательную
кору, мозжечок, ретикулярную формацию,
красное ядро. При стимуляции бледного
шара у животных преобладают элементарные
двигательные реакции в виде сокращения
мышц конечностей, шеи и лица. При этом
выявлено влияние бледного шара на некото-
рые зоны гипоталамуса (центр голода и зад-
ний гипоталамус), отмечена активация пи-
щевого поведения. Разрушение бледного
шара сопровождается снижением двигатель-
ной активности: возникают адинамия, как
бы отвращение ко всякому движению, сон-
ливость, эмоциональная тупость, затрудня-
ются осуществление имеющихся и выработка
новых условных рефлексов.
Ограда. Электрическое раздражение раз-
личных зон ограды вызывает разнообразные
соматические, вегетативные и поведенческие
реакции, например пищевые, ориентировоч-
ные и эмоциональные, сокращение мышц ту-
ловища, головы, жевательные и глотательные
движения и др. Двустороннее разрушение ог-
рады вызывает слабо выраженные нарушения
рефлексов позы, вегетативных реакций и ус-
ловных рефлексов, например дифференци-
ровки раздражителей.
В заключение отметим, что базальные
ганглии — это прежде всего центры органи-
зации сложнейших видов моторной актив-
ности организма. Самые общие предположе-
ния о функциональной роли базальных ган-
глиев сводятся к следующему: допускается
участие этого подкоркового комплекса в со-
здании программ целенаправленных двига-
тельных актов на основе информации, по-
ступающей к ним практически из всех об-
ластей коры. Обработанная информация
далее передается в ядра переднего таламуса,
где она объединяется с информацией, по-
ступающей из мозжечка. Совокупный ком-
плекс возбуждения поступает в лобную
кору, затем в двигательную кору, на нейро-
нах которой конкретная программа реализу-
ется, обеспечивая мозаичное возбуждение и
торможение множества моторных нейронов,
имеющих эфферентный выход на стволовые
и спинальные двигательные центры. Базаль-
ные ганглии контролируют такие параметры
движения, как сила, амплитуда, скорость и
направление. Кроме участия в формирова-
нии двигательных актов, базальные ганглии
включены в регуляцию цикла сон—бодрст-
вование, механизмы формирования услов-
ных рефлексов, сложные формы восприятия
внешней информации (например, осмысле-
ние текста).
139

8.7. НОВАЯ КОРА БОЛЬШОГО МОЗГА
Новая кора (неокортекс) представляет собой
слой серого вещества общей площадью
1500—2200 см
2
, покрывающий большие полу-
шария; она составляет около 72 % всей пло-
щади коры и около 40 % массы головного
мозга. В коре имеется около 14 млрд нейро-
нов,
количество глиальных клеток примерно
в 10 раз больше. Кора большого мозга явля-
ется в филогенетическом плане наиболее мо-
лодой нервной структурой; у человека она
осуществляет высшую регуляцию функций
организма и психофизиологические процес-
сы,
обеспечивающие различные формы пове-
дения.
8.7.1.
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ
ХАРАКТЕРИСТИКА
А. Нейронная организация новой коры. В на-
правлении с поверхности в глубь коры разли-
чают 6 горизонтальных слоев.
I — молекулярный слой имеет очень мало
клеток, но большое количество ветвящихся
дендритов пирамидных клеток, формирую-
щих сплетение, расположенное параллельно
поверхности. На этих дендритах образуют си-
напсы афферентные волокна, приходящие от
ассоциативных и неспецифических ядер та-
ламуса.
II — наружный зернистый слой составлен
в основном звездчатыми клетками и частич-
но малыми пирамидными клетками. Волокна
клеток второго слоя расположены преимуще-
ственно вдоль поверхности коры, образуя
кортико-кортикальные связи.
III
—
наружный пирамидный слой состоит
в основном из пирамидных клеток средней
величины. Аксоны этих клеток, как и зер-
нистые клетки II слоя, образуют кортико-
кортикальные ассоциативные связи.
IV — внутренний зернистый слой по ха-
рактеру клеток (звездчатые клетки) и распо-
ложению их волокон аналогичен наружному
зернистому слою. В этом слое имеют синап-
тические окончания афферентные волокна,
идущие от нейронов специфических ядер та-
ламуса; здесь отмечена наибольшая плот-
ность капилляризации.
V — внутренний пирамидный слой обра-
зован средними и крупными пирамидными
нейронами, причем в двигательной коре рас-
положены гигантские пирамидные клетки
Беца. Аксоны этих клеток образуют эффе-
рентные кортико-спинальный и кортико-
бульбарный двигательные тракты.
VI
—
слой полиморфных клеток образован
преимущественно веретенообразными клет-
ками, аксоны которых образуют кортико-та-
ламические пути.
Оценивая в целом афферентные и эффе-
рентные связи коры большого мозга, можно
отметить, что в слоях I—IV происходит вос-
приятие и обработка поступающих в кору
сигналов. Напротив, покидающие кору эф-
ферентные пути формируются преимуще-
ственно в V—VI слоях. Более детально деле-
ние коры на различные поля проведено на
основе цитоархитектонических признаков
(форма и расположение нейронов) К. Брод-
маном (1909), который выделил 52 поля;
многие из них характеризуются функцио-
нальными и нейрохимическими особеннос-
тями.
Гистологические данные показывают, что
элементарные нейронные цепи, участвую-
щие в обработке информации, расположе-
ны перпендикулярно поверхности коры.
Электрофизиологические исследования
[Маунткасл В., 1957] с погружением микро-
электродов перпендикулярно поверхности
соматосенсорной коры показали, что все
встречаемые на пути нейроны отвечают на
раздражитель только одного качества (вида)
(например, тактильный). Напротив, при по-
гружении электрода под углом на его пути
попадались нейроны разной модальности.
Был сделан вывод, что в коре мозга имеются
функциональные объединения нейронов,
расположенные в цилиндрике диаметром
0,5—1,0 мм. Эти объединения были названы
нейронными колонками. Они обнаружены в
моторной коре, в различных зонах сенсор-
ной коры. Соседние нейронные колонки мо-
гут взаимодействовать друг с другом.
Б.
Кортиколизация функций — возраста-
ние в филогенезе роли коры большого мозга
в анализе и регуляции функций организма и
подчинение коре нижележащих отделов
ЦНС.
Например, регуляция таких двигатель-
ных функций, как локомоция (прыжки, ходь-
ба, бег) и выпрямительные рефлексы у низ-
ших позвоночных (амфибий), полностью
обеспечивается стволом мозга, удаление
больших полушарий практически их не изме-
няет. У кошек перерезка ствола между сред-
ним и промежуточным мозгом существенно
не влияет на выпрямительные рефлексы, но
нарушает в остром периоде локомоцию, ко-
торая в дальнейшем частично восстанавлива-
ется. Выключение коры больших полушарий
у обезьян и у человека приводит к потере не
только локомоции, но и выпрямительных
рефлексов.
140