Нечитайло В.А., Кучерява Л.Ф., Погребенник В.П. Систематика вищих рослин. Лабораторний практикум
Подождите немного. Документ загружается.

В.А. Нечитайло, Л.Ф. Кучерява, В.П. Погребенник
46
Систематика вищих рослин. Лабораторний практикум
47
Клітини у них без хлорофілу, з рівномірно потовщеними оболонками.
Під епідермою в районі гідатоди розміщена особлива тонкостінна па-
ренхіма, яка прикриває жилку. Вода з жилки крізь цю тканину потрапляє
до водяних продихів і виділяється назовні — відбувається гутація. З ве-
чора до ранку в сонячну погоду краплини води добре помітні у хвоща по
краю піхв верхніх вузлів. Рослина ніби "плаче".
У пазухах листків закладаються бруньки, з яких розвиваються гілки
завдяки прориванню трубки листків. Бічні гілки і стебло зеленого кольо-
ру і виконують функцію асиміляції. Детально розглядаючи стебло в лу-
пу, помічаємо гребені і улоговинки, які чергуються з гребенями і улого-
винками вище і нижче розташованих міжвузлів.
На поперечному зрізі міжвузля зеленого стебла видно: епідерму, ко-
ру, провідні пучки і центральну порожнину (рис. 12.3-4). Епідерма скла-
дається із щільно зімкнених клітин із звивистими суміжними стінками.
Зовнішня оболонка цих клітин дуже потовщена і зовні вкрита сосочка-
ми, бородавками, кульками. До складу оболонки клітин епідерми та
інших тканин пагона входить поряд з целюлозою кремнезем. На по-
верхні клітин епідерми кремнезем утворює безперервний шар, вкритий
зовні тонкою кутикулою з восковим нальотом. Він відіграє механічну і
захисну функцію. В епідермі, яка вистелює улоговинки, розташовані ря-
дами продихи. Кожен продих має чотири клітини, розміщені попарно од-
на над одною. Зверху над двома замикаючими клітинами розташовані
дві побічні клітини, які прикривають їх. Всі чотири клітини утворюються
з однієї материнської. Побічні клітини можуть щільно замикати продихо-
ву щілину завдяки наявності замкового механізму, який діє по принципу
замка "блискавка", коли кремнієві вирости однієї побічної клітини вхо-
дять у проміжки другої клітини.
Первинна кора утворена ділянками механічної та фотосинтезуючої
тканин — хлоренхіми (рис. 12.3-4). В гребенях розташована механічна
тканина з великою кількістю кремнезему. Це головна механічна опора
стебла. Хлоренхіма знаходиться в улоговинках або на їх схилах, де ба-
гато продихів. Внутрішню частину первинної кори складає основна па-
ренхіма з тонкостінних, нещільно розташованих округлих клітин. В ній
під улоговинками розташовані валекулярні (пат. VаIIе8 — долина, улого-
вина, тут — напроти улоговин стебла) порожнини, наповнені на початку
свого розвитку водою, а потім повітрям. Зона провідних пучків — цент-
ральний циліндр — відмежована від інших тканин кільцем одношарової
ендодерми (рис. 12.3-4). На поперечному розрізі в зоні провідних пучків
під гребенями видно закриті (без камбію) колатеральні пучки з каріна-
льними (лат. сагіпа — кіль, тут — напроти гребенів стебла) порожнина-
ми у вигляді каналів. Ці канали утворюються внаслідок розчинення де-
яких провідних елементів ксилеми. Дозовні від каналу розташована
флоема, по боках і до середини — ксилема. Флоема складається з сито-
нидних елементів і паренхімних клітин, ксилема — з трахеїд. Основна
ік.іиина — паренхіма — в центрі стебла при його рості утворює централь-
ну порожнину, заповнену спочатку водою, пізніше повітрям. Ця порожни-
м.і разом з валекулярними відіграє важливу роль в газообміні рослини з
н.ніколишнім середовищем. У вузлах об'єм ксилеми збільшується.
Розглядаємо органи розмноження хвоща польового. За допомогою
кореневищ хвощ польовий розмножується вегетативним шляхом. Від
ч.ісіин кореневища утворюються клони дочірніх рослин. Хвощ розмно-
жусіься також спорами, які утворюються на спороносних пагонах. За
р.іхунок поживних речовин кореневища та бульб в кінці квітня — на по-
ч.пку травня з'являються безхлорофільні рослини, що несуть на кінці
і ісбел стробіли від 1 до 3,5 см завдовжки (рис.12.7-8). Стебло у цих
рослин нерозгалужене, утворює в мутовках по 6-16 бурих луско-
подібних зрослих листків, прикріплених у вузлах. Розглянемо стробіли
хиоща в лупу, відпрепаруємо спорангієфори, які мають вигляд шести-
кутних щитків на ніжках.
Знімемо пінцетом окремий щиток. На його внутрішньому боці розташо-
п.іш 6-13 спорангіїв, прикріплених до щитка і звисаючих уздовж ніжки (рис.
ІУ 8). Стінка спорангія багатошарова. Всередині формується спорогенна
ікинина, материнські клітини якої дають тетради гаплоїдних спор. Кожний
спорангій після дозрівання спор розкривається поздовжньою щілиною,
розташованою на внутрішньому боці. Якщо залишити на папері зібрані
спіжі стробіли, то через кілька годин із спорангіїв висипляться спори у ви-
шиді сірувато-зелених клубочків. Розглянемо сухі спори в мікроскоп. Май-
же' круглі зеленуваті спори крім двох звичайних оболонок — інтини і екзи-
ни — мають третю — периній, або епіспорій, яка при дозріванні спори роз-
рипається по спіралі на дві стрічки з розширеними кінцями (рис. 12.9-10).
Ці стрічки називаються елатерами. Вони хрестоподібно прикріплюються
до спори в одному місці. У сухому стані елатери розгорнені і відстоять від
спори. Будучи надзвичайно гігроскопічними, при збільшенні зволоження
••іі.иори швидко скручуються навколо спори. Зміна вологості приводить
спори в рух. Якщо легко подихати на предметне скло зі спорами, що роз-
ніядаються в мікроскоп, то можна спостерігати цікаве явище, коли спори
"і.шцюють" завдяки гігроскопічності елатер. Елатери розпушують масу
спор, сприяють розсіванню їх цілими фупами, а не поодинці. Це забезпе-
чу!; сіагеве розмноження хвоща.
У сприятливих умовах — на вологому затіненому фунті або у воді —
«пори швидко проростають, інші втрачають здатність проростати. Спора
і іюч.ііку набухає, екзина розтріскується, після поділу утворюється ри-
ч»д і заросткова клітина (клітина гаметофіта). Остання багаторазово
дній 11.ся і дає початок зеленому гаметофіту, який набуває вигляду одно-
имропої пластинки, що наростає завдяки верхівковій меристемі. Пізніше
нитті частина гаметофіта збільшується, стелиться по поверхні грунту і
В.А. Нечитайло, Л.Ф. Кучерява, В.П. Погребенник
48
Систематика вищих рослин. Лабораторний практикум
49
ВІДДІЛ ПАПОРОТЕПОДІБНІ, або
ПОЛІПОДІОФІТИ (РОЬУРООЇОРНУТА)
Здебільшого багаторічні, рідше однорічні трави, часом дерева, з са-
мостійними гаплоїдним та диплоїдним поколіннями і переважанням в
циклі розвитку спорофіта.
Листки у папоротеподібних здебільшого багатократно розчленовані,
ірідка цілісні, утворились в процесі еволюції з теломів шляхом зростан-
ня і подальшого сплющення. Як і стебло, листки папоротей наростають
исрхівкою, що свідчить про їхню стеблову природу.
Друга за кількістю фупа серед вищих спорових рослин. До неї нале-
жить понад 12 тис. видів, поширених по всій земній кулі, але переваж-
но — в тропіках.
За однією із сучасних класифікацій у відділі виділяють сім класів, чо-
іири з яких об'єднують викопні папоротеподібні, три — нинішні. Най-
примітивніші папоротеподібні належать до класу Вужачковидні
(Орпюдіоззорзіа'а), а найчисельніші та філогенетично просунуті — до
класу Папоротевидні (РоІуросНорзіда).
КЛАС ВУЖАЧКОВИДНІ, або ОФІОГЛОСОПСИДИ
(ОРНІОСЬ0880Р8ЮА)
Характерними особливостями вужачковидних є: диморфізм листків,
ршноспоровість, товстостінні спорангії без кільця, багаточисленні спори
її одному спорангії (від 1 500 до 15 000), великі (до 6 см завдовжки) га-
метофіти з мікоризою. Все це свідчить про древність і примітивність.
Це невеликі багаторічні, здебільшого наземні рослини, рідше —
епіфіти ( в тропіках). Листки вужачковидних розділені на дві частини —
нерхню спороносну (фертильну) і нижню асиміляційну (хлорофілоносну,
•перильну). Ні перша, ні друга частини листків у молодому віці не бува-
ині> равликоподібно згорнуті.
До класу Вужачковидних належить один порядок та одна родина.
Порядок Вужачкові (Оркіодіоззаіез)
Родина
Вужачкові
(Оркіодіохзасеае)
Родина з трьома родами, які включають понад 80 видів. В Україні
іусірічаються зрідка лише п'ять видів, які ростуть на вологих фунтах в
іш ах, на луках, серед чагарників.
Матеріал для заняття: гербарні зразки вужачки та гронянки,
і/чіи
<ниіний
матеріал,
мікропрепарати.
Л.шдання: розглянути гербарні зразки вужачки та гронянки, звернути
ун.і.у 11, і листки, спорангії, кореневища, корені, виготовити препарати.
на ній утворюються прямостоячі зелені пластинки до 1-4 мм завдовжки
(рис. 12.11-12, 15). До фунту гаметофіт прикріплюється безбарвними
ризоїдами до 1 см завдовжки. Гаметофіти (заростки) хвоща зелені, жив-
ляться автотрофно. Складаються вони з паренхімних клітин і є пластин-
ками різної форми (рис.12.15) переважно одностатевими, але бувають і
двостатеві. Архегонії розвиваються у виїмках між лопатями вертикаль-
них пластинок, черевце їх занурене в тканину гаметофіта, шийка висту-
пає назовні. Антеридії утворюються на значно менших за розміром чо-
ловічих гаметофітах і занурюються повністю в тканину гаметофіта (рис.
12.12а). Жіночі гаметофіти дуже гіллясті, можуть стати двостатевими.
Взагалі, у сприятливих умовах утворюється значно більше жіночих за-
ростків, ніж чоловічих. У несприятливих умовах чоловічі гаметофіти пе-
реважають над жіночими. Сперматозоїди у хвоща багатоджгутикові —
джгутиків до 100 шт. (рис. 12.14). Запліднення відбувається при наяв-
ності краплиннорідкої води і забезпечується завдяки тому, що спори хво-
ща, зчіплені елатерами, поширюються ірупами. Після запліднення на
жіночому гаметофіті з зиготи формується зародок, а далі проросток з ли-
стковими піхвами, здатний самостійно живитися. Проросток нарешті пе-
ретворюється на літній пагін з кореневищем.
Гаметофіти хвоща можна виростити в лабораторії, якщо висіяти в чашку
Петрі на вологий пісок спори із зібраних стробілів і накрити їх склом. Через
1 -1,5 місяці заростки розвиваються до стадії формування статевих органів.
Хвощ польовий — бур'ян, який масово розвивається на полях з кис-
лими фунтами. Лікарська рослина. Пагони містять кремнієву кислоту,
алкалоїди, флавоноїди, сапоніни, каротин, аскорбінову кислоту. Раніше
населення Євразії і Північної Америки вживало в їжу бульби хвоща по-
льового після спеціальної обробки.
Розглядаємо гербарні зразки інших видів хвощів.
Хвощ великий (Ещиізеїит Іеїтаіеіа) має два типи пагонів. Росли-
на досягає 1 м заввишки, зустрічається зрідка по заболочених місцях, у
тінистих лісах та на схилах. Релікт третинного періоду. Рідкісна зникаю-
ча рослина, що охороняється.
Хвощ лісовий (Е. зуЬгабсит) зустрічається в чагарниках, тінистих
лісах, на узліссях. Рослина має розгалужені бічні пагони і стебла, спороносні
після опадання стробілів зеленіють і галузяться. Звичайна рослина.
Хвощ болотний (Е. раїизіге) поширений на болотах, уздовж берегів
водойм, на вологих луках. Звичайна рослина. Спороносні та вегетативні
пагони однакові. Після опадання стробіла рослина продовжує рости і
фотосинтезувати.
Хвощ зимовий (Е. Ьуетаїе) відрізняється від інших жорсткими вічно-
зеленими пагонами, на яких немає бічних гілок. Спороносні та вегетативні
пагони однакові. Поширений в лісах, на вологих піщаних місцях.

В.А. Нечитайло, Л.Ф. Кучерява, В.П. Погребенник
Рис. 13. Вужачкові: 1-6 - вужачка звичайна (Орпіодіоззит уиїдаіит)-
1 - загальний вигляд (а - вегетативна частина листка, б - спороносна)
2 - ділянка листка зі спорангіями (а), 3 - заросток (гаметофіт) з архегоніями і
антеридіями та початковим розвитком спорофіта (с), 4 - антеридій, 5 - спер-
матозоїд, 6 - архегоній; 7-8 - гронянка півмісяцева (ВоігусЬіит Іипагіа):
7 - загальний вигляд (а - вегетативна частина листка, б - спороносна)
8 - ділянка листка зі спорангіями (а)
50
Систематика вищих рослин. Лабораторний практикум
Рід Вужачка {Орпіодіоззит)
Об'єднує близько 45 видів, поширених майже по всій земній кулі,
особливо в тропіках. У нашій флорі є лише один вид з цього роду.
В. звичайна (О. уиїдаїит) (рис. 13.1) дуже рідко зустрічається на
пуках і серед кущів майже всієї лісової зони України.
Розглянемо морфологічні особливості вужачки звичайної. Це неве-
ника рослина (5-20 см заввишки) з коротким косим кореневищем і тов-
сїими мичкуватими коренями (рис. 13). Листків 1-2, з нерозчленовани-
ми частинами. Стерильна частина листка цілісна, гола, довгаста або
панцетна. Спороносна частина довша за стерильну, подібна до
стробіла, несе від 12 до 40 пар спорангіїв. Спорангії розкриваються по-
перечною щілиною (рис. 13.2)
Гаметофіти (рисі3.3) підземні, циліндричні двостатеві, до 6 см за-
пдовжки, сапрофітні, розвиваються дуже повільно (1-20 років). Архе-
юнії і антеридії занурені в тканину гаметофіта. Сперматозоїди мають
численні джгутики (рис. 13.5).
Вужачка звичайна — рідкісний вид, який охороняється.
Рід Гронянка (Воігуспіит)
Включає 36 видів, поширених в помірній зоні північної півкулі. У флорі
України є чотири рідкісних види з цього роду.
Гронянка півмісяцева, або ключ-трава (В. Іипагіа) (рис. 13.7-8).
Дуже рідко можна зустріти цей вид на сухих луках, у світлих лісах та ча-
ирниках Карпат, Криму, Полісся і Лісостепу.
Розглянувши наочний матеріал, відмічаємо, що гронянка півмісяце-
п.і - це невелика (5-20 см заввишки) рослина. Відрізняється від вужач-
ки перисторозсіченими листками. Стерильна частина їх периста, з
цпіісними півмісяцевими або клиновидними сегментами, по 2-9 з ниж-
нього боку. Спороносна частина листка двічі-перисторозсічена, волоте-
подібна, на довгій ніжці.
Гронянка півмісяцева — рідкісна рослина, охороняється. Занесена в
Червону книгу України (1996).
КЛАС ПАПОРОТЕВИДНІ, або ПОЛІПОДІОПСИДИ
{РОЬУРООІОРЗША)
Гі.нагорічні, рідше однорічні рослини. Стебла у вигляді прямостоячих
і юнЬурів у деревовидних папоротей, у трав'янистих — кореневища. Ли-
< іки різноманітні за розмірами, формою. Спорангії в сорусах, здебільшо-
ІІІ і одношаровими стінками та кільцем. Найбільший за кількістю видів
ічі.и. близько 12 тис. видів, що об'єднуються в три підкласи, 6-8 по-
ридкін іа 15-30 родин (за різними системами).
П кпасі Поліподіопсиди розрізняють такі підкласи: Поліподііди
(Гпіуіпніисіае), Марсилііди (Магзіїеісіае) та Сальвінііди (За/у/п/'/с/зе).
51

В.А. Нечитайло, Л.Ф. Кучерява, В.П. Погребенник
.52
Систематика вищих рослин. Лабораторний практикум
Рис 14. Страусове перо та безщитник: 1-3 - страусове перо звичайне
(Маііеиссіа зігиіпіоріегіз), 1 - загальний вигляд (а - стерильний листок — тро-
фофіл б - фертильний листок - спорофіл), 2 - фрагмент спороносного ли-
стка 3 - частина сегмента листка з сорусами; 4-6 - безщитник жіночий
(Аіпугіит Шх-іетіпа), 4 - загальний вигляд; 5 - листок, 6 - частина листка з
сорусами (а)
53
ПІДКЛАС ПОЛШОДПДИ (РОЬУРООІШАЕ)
Сюди належать лише рівноспорові папоротевидні з різних життєвих
форм, різних розмірів і різної екології. Рослини вражають своїм різно-
маніттям, їх систематичними ознаками є характер стебла, листків, со-
русів, покривальця (індузія), кільця спорангіїв тощо. Гаметофіти двоста-
теві. Близько 12 тис. видів. В Україні зростає 52 види з 15 родин.
Порядок Багатоніжкові (Роїуройіаіеь)
Найчисельніша група в класі Папоротевидні. Наземні або епіфітні,
рівнинні або гірські. Спорангії тонкостінні, зібрані в соруси здебільшого
на звичайних листках, рідше на окремих спорофілах, прикритих
індузіями різної форми, або без індузія.
До порядку відносять за різними системами від 5 до 20 родин. Ряд
родин розглядається в ранзі підродин.
Родина
Оноклеєві
(Опосіеасеае)
Рослини з диморфними, рідко однаковими листками. Соруси при-
криті загнутим краєм листкової пластинки, розташовані по жилках.
В родині 10 видів і 3 роди, з них в Україні — 1 вид.
Матеріал для заняття: гербарні зразки страусового пера.
Завдання: розглянути гербарні зразки, звернути увагу на соруси.
Рід Страусове перо (Маііеиссіа)
Страусове перо звичайне [Маііеиссіа зігиіпіоріегіз=5ігиіпіоріе-
гів ЛІісазігит) (рис. 14.1-3) зустрічається в Карпатах, на Поліссі та в
Лісостепу, в тінистих лісах, у вологих чагарниках і вільшняках, по воло-
гих ярах. Поширене в Європі та Азії. Багаторічна рослина 60-170 см зав-
вишки, з товстим кореневищем і пучком диморфних листків. Вегетативні
листки зелені, двічі перисті, з широкодовгастою гострою пластинкою,
спороносні бурі, перисті, менші за вегетативні, розвиваються всередині
утвореної вегетативними листками лійки. Пера першого порядку у цих
листків лінійні, цілокраї, майже циліндрично згорнуті. Соруси спочатку
округлі, потім безперервні, розташовані на потовщеннях жилок, прикриті
дуже опуклим, по краю розірваним покривальцем, що рано опадає.
Граціозна декоративна рослина, яку вирощують у тінистих парках.
Лікарська рослина. Рідкісний вид, підлягає охороні.
Родина
Безщитникові
(Аіпугіасеае)
Кореневища у рослин вкриті листковими темно-червоними лусками.
Черешки з двома судинними пучками. Індузії продовгуваті, нирко-
подібні, гачкуваті, підковоподібні або взагалі відсутні. Спори бобо-
подібні. На всій земній кулі нараховують чотири-п'ять родів і близько
600 видів. В Україні з них зустрічається 8 видів.
Матеріал для заняття: гербарні зразки жіночої папороті.
В.А. Нечитайло, Л.Ф. Кучерява, В.П. Погребенник
Систематика вищих рослин. Лабораторний практикум
55
і продихами) з хлорофіловими зернами, зелену паренхіму між ними та
провідний пучок на місці перерізаної жилки (рис.15.3). Вивчаючи внутрішню
пудову кореневища, відмічаємо, що його провідна система (диктіостела) скла-
дається з багатьох концентричних пучків, в центрі яких розташована ксилема,
оточена флоемою. До складу ксилеми входять драбинчасті і кільчасті судини,
флоема, побудована з ситовидних трубок з сітчастими бічними стінками.
Соруси (ф. 50Г05 — купка) у чоловічої папороті та багатьох інших ут-
ворюються на нижньому боці листка і прикриті тонким безбарвним покри-
вальцем — індузієм (лат. іпдизіит — верхня туніка), що виконує захисну
функцію. Соруси чоловічої папороті круглі, розташовані по обох боках се-
редньоїжилки листкових часток — пер другого порядку. Кожен сорус при-
критий покривальцем бобоподібної форми (рис. 15.2). Отже, листки чо-
ловічої папороті є одночасно і спорофілами, і трофофілами, тобто вико-
нують функції асиміляції та спороношення.
Далі розглянемо сорус в розрізі (рис. 15.3). Під нижньою епідермою ба-
чимо масивний виріст листка, що називається плацентою. На верхівці пла-
цента переходить в розширений індузій, під яким розташовано кілька спо-
рангіїв на довгих ніжках. На поверхні кожного спорангія добре помітне кільце,
яке починається від ніжки, облямовує спорангій і закінчується на другому боці
недалеко від ніжки. Знімемо спорангії з сухих листків папороті і розглянемо їх
в мікроскоп. Відмічаємо, що оболонка клітин кільця неоднакового кольору і
товщини. Товста коричнева оболонка кожної клітини кільця має ІІ-лодібну
форму в місці примикання до сусідніх клітин кільця та до стінки спорангія.
Вільна зовнішня частина оболонки клітин кільця тонка, легко пропускає воду.
Під час падіння осмотичного тиску всередині клітин кільця гх тонкі стінки втя-
гуються, при цьому різко скорочується верхній бік кільця. Таке явище відбу-
вається в результаті дозрівання і висихання спорангія. В суху погоду за допо-
могою кільця, що випрямляється або вигинається в протилежний бік, стінка
спорангія розривається в місці, де кільце не доходить до ніжки і де клітини
тонкостінні. Тут утворюється отвір — стома (лат. зіота — рот) (рис. 15.4).
Дозрілі спори завдяки механізму, що діє як катапульта, з силою викидаються
і розсіваються вітром. Розкриття спорангіїв можна спостерігати в лабораторії
на занятті, якщо провести досить простий дослід. Зберемо фільтрувальним
папером частину води з препарату, на якому розглядаємо спорангії, і заміни-
мо воду краплею спирту. Спирт відтягує з клггин кільця воду, зменшує тургор,
і кільце різко випрямляється внаслідок розривання стінки спорангія.
У спорангіях папороті з археспоріальної тканини шляхом редукційно-
го поділу формуються гаплоїдні спори, що дають початок гаплоїдному
поколінню — гаметофіту. На вологому фунті спора проростає і утворює
іаросток — самостійно існуючу рослину. Це зелена пластинка серце-
подібної форми до 1 см діаметром, прикріплена до поверхні грунту ри-
юїдами (рис. 15.5). На нижньому боці заростка формуються архегонії та
.інтеридії, занурені в тканину заростка. Після запліднення яйцеклітини
54
Завдання: розглянути гербарні зразки і соруси на листках без-
щитника жіночого.
Рід Безщитник (Аіпугіит)
Налічує близько 200 видів, з них в Україні — лише 2.
Безщитник жіночий, або жіноча папороть (АіЬугіит ЯІіх-їетіпа)
(рис. 14.4-6). зустрічається в тінистих мішаних і хвойних лісах і чагар-
никах по всій території України Характерний для лісової зони Євразії та
Північної Америки.
Це багаторічна трав'яниста рослина 30-150 см заввишки, з коротким
потовщеним кореневищем. Соруси спорангіїв утворюються на нижньо-
му боці листка, прикриті ниркоподібним або зігнуто-серпоподібним
індузієм. Листкова пластинка двічі-тричі перисто-розсічена. Порівняно з
листком чоловічої папороті листок безщитника жіночого більше розріза-
ний, ажурний, з вузькими перами першого та другого порядків.
Безщитник жіночий —декоративна і лікарська рослина.
Родина Щитникові, або Аспідієві
(Пгуоріегійасеае, або АзрШіасеае)
Рослини з потовщеними, короткими або сланкими тонкими, вкрити-
ми матовими лусками кореневищами. Листки двічі-тричі перисто-
розсічені, шкірясті і трав'янисті. Соруси на нижньому боці листка з щи-
топодібним, серцеподібним індузієм або без нього.
В родині близько 25 родів та 800 видів, поширених в лісових та
гірських областях земної кулі. В Україні з них зустрічається 13 видів з
трьох родин.
Матеріал для заняття: гербарні зразки чоловічої папороті, ли-
стки з сорусами, анатомічні препарати кореневища, соруса та лист-
ка в розрізі, заростки.
Завдання: розглянути гербарні зразки щитника чоловічого, лис-
ток, сорус та кореневище, звернути увагу на характер розташуван-
ня сорусів на листках, архегоніїв та антеридіїв на заростку, будову
спорангіїв.
Рід Щитник (Огуоріегіз)
Включає близько 150 видів, з них в Україні — 6.
Щитник чоловічий, чоловіча папороть (Огуоріегіз ЛІіх-таз) (рис.
15). Багаторічна трав'яниста рослина з коротким кореневищем. На верхівці
кореневища розташований пучок листків, а вниз відходять численні корені,
які є додатковими. Листки закладаються на верхівці кореневища і протягом
першого літа запишаються зачатковими. На другий рік вони мають вигляд
спіралей, вкритих коричневими лусками і розміщених в середині пучка до-
бре розвинених листків. Навесні третього року вони швидко розгортаються,
звільняються від лусок і стають дорослими. В листку папороті розрізняють
черешок і рахіс, пера першого, другого і вищих порядків.
На поперечному зрізі листка помітні верхній та нижній епідерміс (останній

В.А. Нечитайло, Л.Ф. Кучерява, В.П. Логребенник
Рис. 15. Щитник чоловічий (Огуоріегіз Шх-таз): 1 - загальний вигляд
(а - черешок листка, б - рахіс), 2 - частина листка з сорусами на нижньому
боці, 3 - сорус в розрізі (а - плацента, б - індузій, в - спорангій, г - пластинка
листка в розрізі), 4 - спорангій (а - кільце, б - стома, в - ніжка)
56
Систематика вищих рослин. Лабораторний практикум
Рис. 15. (продовження): 5 - заросток (гаметофіт) з нижнього боку (а - ар-
хегоній, б - антеридій, в - ризоїди), 6 - архегоній, 7 - антеридій із спермаген-
ними клітинами, 8 - розкритий антеридій і сперматозоїди, 9 - сперматозоїд,
10 - молода папороть (проросток) (а) на-гаметофіті (б)
Родина
Аспленієві (
А$рІепіасеае)
Наземні та епіфітні трав'янисті листопадні та вічнозелені рослини,
що населяють переважно вологі місця. Кореневища вкриті лусками. Со-
руси з яйцеподібними або лінійними індузіями. В родині налічують
близько 700 видів, з них в Україні — 11 видів.
Матеріал для заняття: гербарні зразки аспленія волосовидного
та пистовика сколопендрового.
багатоджгутиковим сперматозоїдом розвивається зародок, що має стеб-
ло, корінь, листок, підвісок, який з'єднує зародок з тканиною гаметофіта.
Зародок, що розвивається з диплоїдної зиготи, є спорофітом. Первинне
його живлення відбувається за рахунок гаметофіта. Зародок перетво-
рюється на проросток, коли з'являються вегетативні органи і починають
функціонувати корінь, стебло, листки. Гаметофіт поступово відмирає.
Чоловіча папороть — лікарська і декоративна рослина.
57
В.А. Нечитайло, Л.Ф. Кучерява, В.П. Погребенник
Завдання: розглянути гербарні зразки, звернути увагу на особли-
вості будови сорусів
Рід Аспленій (Азріепіит)
Налічує 680 видів, в Україні є дев'ять.
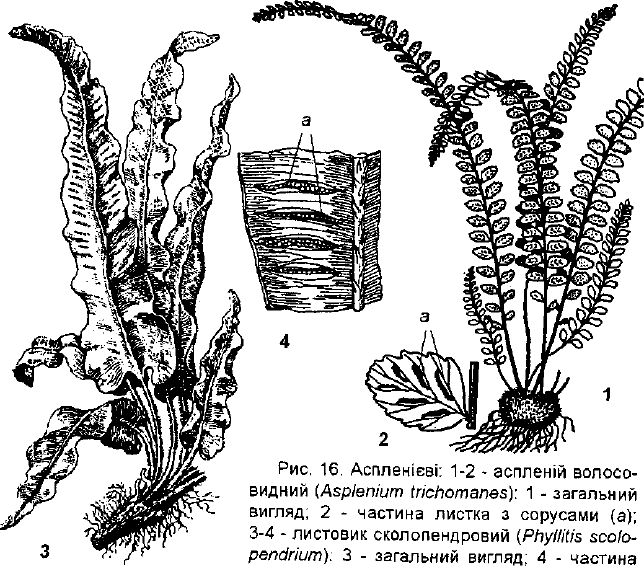
Аспленій волосовидний (Азріепіит ігіспотапез) (рис. 16.1-2)
росте на затінених скелях у Карпатах, на Правобережному Поліссі, в
Лісостепу, рідко в Степу та в Криму, трапляється на затінених схилах в
широколистяних лісах. Поширений в Європі, Азії і Північній Америці.
Багаторічна рослина 5-15 см заввишки з коротким прямостоячим коре-
невищем, вкритим темними лусками. Листки перисті; черешок і рахіс
листка темно-бурі, блискучі, голі. На обернено-яйцеподібноокруглих пе-
рах першого порядку розташовані довгасті або лінійні купки спорангіїв з
покривальцем, прикріпленим з одного боку.
Рослина дуже декоративна, використовується в затінених парках та
оранжереях. Застосовується в народній медицині. Аспленій стає
рідкісною рослиною і тому підлягає охороні.
листка з сорусами (а)
Систематика вищих рослин. Лабораторний практикум
58
59
Рід Листовик (РпуІШіз)
Налічує чотири види, в Україні — 1.
Листовик сколопендровий (РпуІІШз зсоїорепсігіит) (рис. 16.3-4)
поширений в Європі та Північній Америці. На території України зу-
стрічається зрідка в Карпатах, Криму, на Поліссі, на затінених кам'янис-
тих місцях. Багаторічна наземна рослина 30-60 см заввишки, з товстим
коротким кореневищем і нерозсіченими цілісними і цілокраїми довгасто
ланцетними слабо шкірястими листками, що мають стріло- або серцепо-
дібну основу (рис. 16.3). Цими листками листовик відрізняється від усіх ін-
ших папоротей, що мають розсічені листки. На нижньому боці листків роз-
ташовані косо упоперек у вигляді великих смуг лінійні соруси, парами по
обидва боки жилки (рис.16.4). Соруси прикриті двома покривальцями.
Рослина декоративна, культивується в оранжереях, в парках, на заті-
нених скелях. Листки використовуються в народній медицині та гомеопатії.
Родина Деннштедтієві, або Гіполепісові
(Реппзіейііасеае, або Нуроіерійасеае)
Великі рослини з горизонтальними кореневищами і двічі-тричі перистими
листками. Соруси крайові, прикриті індузієм і загорнутим краєм листка.
В родині 6 родів і близько 80-100 видів, поширених здебільшого в
тропічних і субтропічних областях, рідше в полярних широтах. З них в
Україні є два види.
Матеріал для заняття: гербарні зразки орляка звичайного,
мікропрепарати поперечного розрізу кореневища.
Завдання: розглянути гербарні зразки та препарати, звернути
увагу на соруси.
Рід Орляк (РІегіаЧит)
Включає один поліморфний вид, з якого часто виділяють 4-8 видів.
З них в Україні є два.
Орляк звичайний (Ріегісііит ациіііпит) (рис. 17) зустрічається у со-
снових та мішаних лісах, по чагарниках, майже по всій Україні. Космо-
політно поширений на всій земній кулі, крім пустель та тундри. Багаторічник
50-100 см заввишки, з довгим повзучим чорним кореневищем і великими
тричі перистими шкірястими листками на довгих черешках.На поперечно-
му перерізі кореневища розглядаємо типову диктіостелу та будову окремо-
го провідного пучка (рис. 17.3-4). Купки спорангіїв розташовані на нижньо-
му боці листків і у вигляді безперервної облямівки тягнуться по самому
краю листкових часток під прикриттям загорнутого вниз безбарвного війча-
стого краю листка і перетинчастого війчастого покривальця (рис.17.5-8).
У багатьох країнах орляк став бур'яном, що потребує спеціальних
методів боротьби. В житті людини відіграє також і позитивну роль. На
Далекому Сході, в Японії і Китаї молоді ніжні листки ще в черешках
вживають в їжу як овочі. Жителі багатьох країн споживають в їжу і коре-
невище, багате крохмалем. Орляк застосовується в медицині як про-

В.А. Нечитайло, Л.Ф. Кучерява, В.П. Погребенник
Рис. 17. Орляк звичайний (Ріегісііит ациП'тит): 1 - загальний вигляд;
2 - частина кореневища; 3, 4 - поперечний переріз кореневища (схема) та ок-
ремого провідного пучка (а - зовнішня кора, б - внутрішня кора, в - півкільце
механічної тканини, г - провідні пучки, д - паренхіма кори, е - ендодерма,
є - перицикл, ж - крохмалоносний шар, з - флоема, / - ксилема)
60
Систематика вищих рослин. Лабораторний практикум
Рис. 17 (продовження). 5, 6 - частина листка з сорусами по краю всіх сег-
ментів листка (а - соруси); 7 - кінцева частина пера першого порядку з ниж-
нього боку; 8 - соруси під загорнутим краєм листка.
61
тиглистний засіб, а також при інших захворюваннях — народною меди-
циною. В ряді країн орляк використовується як паливо, покрівельний ма-
теріал, добриво. З попелу одержують поташ для виробництва скла і зе-
леного мила. Листки мають бактерицидні властивості.
В. А. Нечитайло, Л.Ф. Кучерява, В.П. Погребенник
62
Систематика вищих рослин. Лабораторний практикум
68
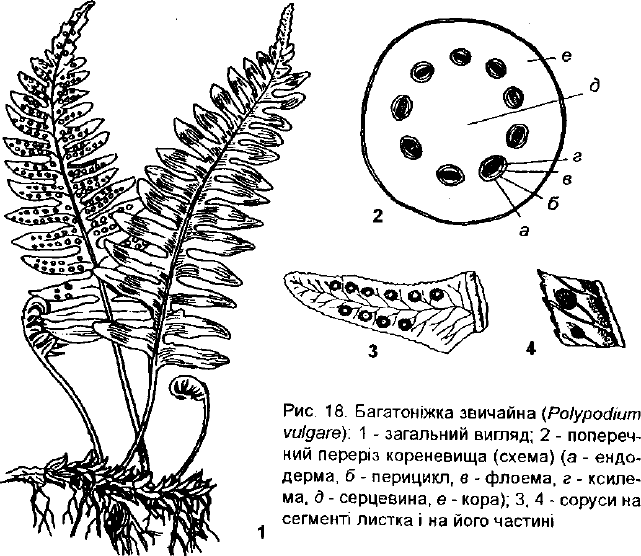
иищем (рис. 18, 1). Соруси розташовані у два ряди на нижньому боці
иисіків, округлі, без індузія (18, 3-4). Декоративна й лікарська рослина. В
кореневищі містяться глікозиди, яблучна кислота, сапоніни. В листках є
моїаш. Численні садові форми багатоніжки культивують у відкритому
Фунті та в оранжереях.
ПІДКЛАС МАРСИЛІІДИ (МАКЗІЬЕЮАЕ)
Характерними особливостями представників підкласу є: водний і зем-
і«жодний спосіб життя; різноспоровість, а звідси і різностатеві гаме-
тфіти; наявність особливих органів — спорокарпіїв, в кожному з яких є і
мікро-, і мегасоруси одночасно.
В підкласі є один порядок — Марсилієві (Магзіїеаіез) і одна родина
Марсилієві (Магзіїеасеае) з трьома родами і невеликою кількістю видів
(ппизько 80).
Порядок Марсилієві {Магзіїеаіез)
Родина
Марсилієві
{Магзіїеасеае)
Матеріал для заняття: гербарні зразки марсилії чотирилистої,
фіксований матеріал, спорокарпії, мікропрепарати.
Завдання: розглянути листки, стебла, спорокарпії, виготовити
мікроскопічні препарати.
Рід Марсилія (Магзіїеа)
Цей рід найчисленніший серед інших: нараховує близько 70 видів, по-
ширених майже по всій Землі, але здебільшого в північній півкулі. Це жи-
пчи водойм та заболочених місцезростань. В Україні з роду Марсилія є
пише один вид.
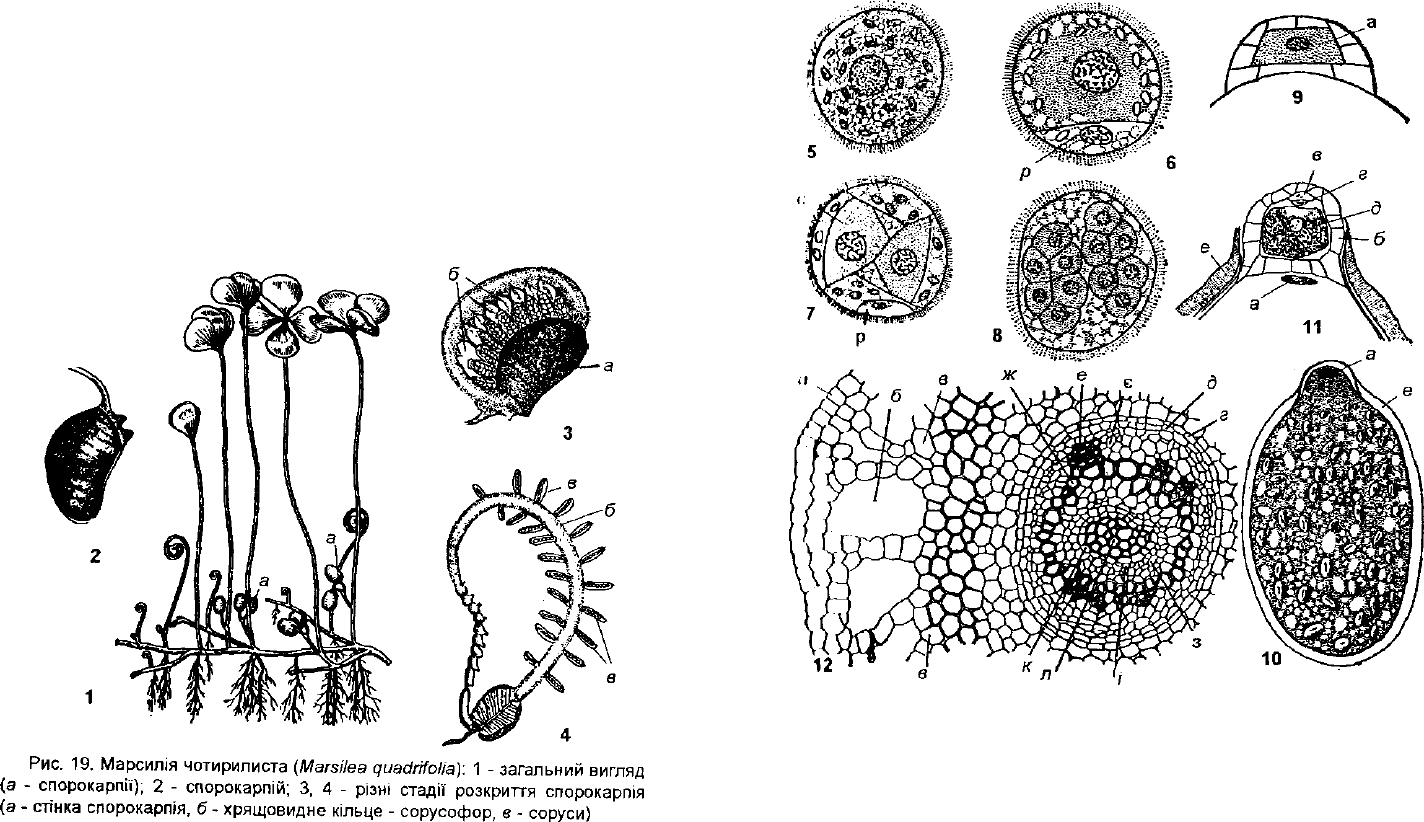
М. чотирилиста (М. циабгіїоііа) (рис. 19). Рослина зустрічається ду-
жо рідко на мілководдях водойм, на замулених берегах в пониззях
Днісіра та Дунаю, в Закарпатті і на Західному Поліссі. Як зникаючий
рппікговий вид вона внесена в Червону книгу України (1996).
Розглянемо морфологічні особливості марсилії чотирилистої. Рослина
мш; іонке га довге поверхневе кореневище, до якого у вузлах прикріплені
'іисішнні корені і листки довжиною від 10 до 80 см (рис. 19.1). На верхівці
чі'р»;шка розміщені чотири яйцевидно-клиновидні листочки; два з них чер-
пни і дна — супротивні. Листок марсилії дуже схожий на листок конюши-
ни Саме тому марсилію і називають водяною конюшиною. Молоді листки
марсилії згорнуті равликом. Розглядаючи уважно листки (рис. 19.1), в їх
оскомі помічаємо овальні спорокарпії, поодинокі або по кілька штук на че-
(пчику листка. В розкритих спорокарпіях можна побачити хрящовидне
кільце (сорусофор) з сорусами, розміщеними в два ряди (рис.19.3-4).
Н кожному сорусі до плаценти прикріплені мегаспорангії на верхівці, а
по Гкжах численні мікроспорангії. В мікроспорангіях утворюється 64
Родина
Багатоніжкові
(РоІуроаЧасеае)
Листки розміщені дворядно на верхньому боці соковитих м'ясистих
кореневищ, вкритих лусками і волосками. Соруси округлі або еліптичні,
без покривця, розташовані на листках знизу. Представники родини
зустрічаються по всій земній кулі, але переважно в тропіках Старого
світу. В родині налічують близько 1000 видів та ЗО родів. З них в Україні
у природному стані є лише два види.
Матеріал для заняття: гербарні зразки багатоніжки звичайної.
Завдання: розглянути гербарні зразки, звернути увагу на соруси.
Рід Багатоніжка (Роїуродіит)
Налічує 75 видів, поширених здебільшого в тропіках (епіфіти на де-
ревах), також на скелях і пнях. У флорі України є два види.
Багатоніжка звичайна (Роїуродіит уиїдаге) (рис. 18) трапляється
спорадично майже по всій території України на замоховілих скелях, валу-
нах, на залісених схилах ярів, на стовбурах дерев, завжди в тіні. Пошире-
на в Європі, Азії, Північній Америці. Багаторічник 8-70 см заввишки, з
простими перистороздільними зимуючими листками і повзучим корене-
В.А. Нечитайло, Л.Ф. Кучерява, В.П. Погребенник
64
Систематика вищих рослин. Лабораторний практикум
І'ис 19 (продовження): 5 - мікроспора; 6, 7 - проростання мікроспори (р -
ри хидіїпьна клітина, с - спермагенна клітина); 8 - чоловічий гаметофіт з дво-
м.і ілпі'ридіями (в кожному утворюється по 6 сперматозоїдів); 9 - мегаспора
і ид|юм (./), 10-11 - проростання мегаспори і утворення архегонія на верхівці
момсмори (я - ядро мегаспори, б - клітини стінки архегонія, в - шийкова ка-
ниііміпііл клітина, г - черевцева канальцева клітина, д - яйцеклітина,
м оГкіїїонка мегаспори); 12 - поперечний переріз стебла (а - епідерміс,
я мііннрнна паренхіма (аеренхіма), в - кора, г - зовнішня ендодерма, д - зов-
нішній мприцикл, е - протоксилема, є - зовнішня флоема, ж - метаксилема,
» мнуцишші флоема, і - внутрішній перицикл, к - внутрішня ендодерма,
II і н|іЦиНИІІ,і)
65
мікроспори, а в мегаспорангіях — одна мегаспора. Спори протягом 48 го-
дин проростають і формують дуже редуковані гаметофіти. На чоловічому
гаметофіті утворюється ризоїдальна клітина і два антеридії, кожен з яких
має спермагенну клітину і по три клітини стінки. Отже, весь чоловічий га-
метофіт складається з дев'яти клітин (рис. 19.8).
Жіночий гаметофіт (рис. 19.9-11) утворюється в мегаспорі. На верхівці
мегаспори, ядро якої ділиться, формується один архегоній з яйцекліти-
ною, черевцевою і шийковою канальцевими клітинами (рис.19.11).