Middleton G.V. (Ed.) Encyclopedia of Sediments and Sedimentary Rocks
Подождите немного. Документ загружается.


BIOGENIC SCDIMCNTARY STRUCTURES
General Traits
Common to
Brackish
Water
Ichnofacies
kolilhos^CyUndrichnus
Assoc.
nch
simple,
veftiail hitrnnvs are
common to estiiarine
sands and
sand-dominated
heterolithic beds
typicalJortm:
Skolithos.
Cylindrichnus &
.Atvnicolites
Figure B17 General brackish waler Irace fossil assemblaf^c. (Moditicd from Pomborlon ct .il.,
20021.
reported from a wide range of freshwater-terrestrial environ-
ments, in both ancienl and recent settings (Ekdale ctitl.. 1984).
At present, nonmarine environments have been characterized
by ihi'cc main ichnofacies: the Scoyetiiit. the Mfrniui. and the
Icrniitichiuis ichnofacies. Frey clal. (19^4) concluded that the
Scoyenia ichnofacies remains a valid concept within appro-
priate limits and suggests deposition on the shore of ephemeral
lakes and the overbank of sluggish rivers. The Scoyenia
assemblage is characterized by: (1) small horizontal, lined,
backfilled feeding btirrows; (2) curved to tortuous feeding
burrows; (3) sinuous crawling traces: (4) vertical, nnlined.
cylindrical to irregular shafts: and (5) vertebrate and arthro-
pod tracks and trails. Buatois and Mangano (ly^J.^) concluded
that the Mcruiia ichnofaeies typifies unconsolidated, fine-
grained, permanent subaqueous substrates, and well-oxyge-
nated, low-energy lake bottoms. The Mcrtnia assemblage is
characterized by: (I) dominance of horizontal to sub-liorizon-
lal grazing and feeding (races produced by mobile deposit
feeders: (2) subordinate occurrence of locomotion traces: O)
generally high to moderate diversity and abundance: and (4)
low abundance of specialized grazing structures. Smith ciat.
(1993) named the Termitichnus ichnofacies as a distinct
terrestrial assemblage characterizing paleosols. The Termitich-
nits ichnofacies is characterized by: (I) vertebrate tracks and
trails:
(2) low diversity and abundance:
(?•}
dominated by bee,
beetle, and termite nests; (4) few feeding structure; and (5)
rhizoliths. coprolites. and leaf miners. Prospects for the
recognition of additional archetypal nonmarine ichnofacies
remain encouraging. For example, Fkdale ct id. (19S4) and
Frey and Pemberton (1987) noted that distinct suites of traee
fossils characterize eolian dunes, fluvial, paleosol, and lake
environments, amonu others.

76 BIOCFNIC SFDIMFNTARY STRUCTURES
Brackish water ichnofacies
Brackish marginal marine environments are widespread in the
modern, and are being documented with more frequency from
the roek record. The general brackish water iraee fossil
assemblage (Figure B18) rellects inherently iluctuating envir-
onmenlal parameters (salinity, exposure, sedimentation rates,
temperature, and shifting substrates). These faetors suggest
that the benthie biota of braekish water environments is indeed
unique and in contrast to Bromley and Asgaard (1991) the
influenec of salinity gradients can hardly be divorced from the
ichnocoenosc concept. For the same reasons that distinct non-
marine associations are now being reeognized. unique brackish
water assemblages arc also widespread. In summary, brackish
water environments tend to be eharacterized by: (I) an
impoverished marine suite of benthie organisms; (2) traee-
making infaunal animals are more abundant than epifaunal
animals; (3) soft bodied organisms are more prevalent than
shelled organisms; (4) some animals show a distinet size
reduction; (5) some animals are bathymetrically displaced; (6)
a higher percentage of the assemblage of benthie organisms
are trophie generalists whose netivities result in morphologi-
cally simple burrows; and (7) although diversity is reduced,
high individual densities may be attained (Pemberton el al.,
1992).
Marine soft ground ichnofacies
The Psilonichmis ichnofacies is associated with supralittoral/
upper littoral, moderate to low-energy marine and/or eolian
conditions typically found in beach to backshore to dune
environments. The comments by Bromley and Asgaard (1991)
notwithstanding, the Fsilonichinis iehnofacies was founded on
fossil e.xamplcs (Frey and Pemberton. 1987) and is no more
theoretical than any other reeiirrent iehnofacies; rather, the
modern ichnocoenoses were emphasized to show the richness
that one might reasonably expect to have existed for various
ancient ichnofaunas. Furthermore, one of the major tenets of
ichnofaeies reconstruction is that the namebearer need not be
present in every occurrence of the iehnofacies; thus, just as
Crti-iana is rare in post-Paleozoic oeeurrenees of the Ciuzi-
anii ieiinofaeies. Psilonichmis may well be absent in pre-
iVleso7oic occurrences of the Psilonichmis ichnolacies.
The Skolilhos ichnofaeies (Figure BI9) is generally asso-
ciated with high-energy, sandy, shallow-marine environments.
The traee fossils are characterized by: (1) predotninantly
vertical, cylindrical or U-shaped burrows; (2) few horizontal
structures; (3) few structures produced by mobile organisms;
(4) low diversity, although individual forms may be abundant;
and (5) mostly dwelling burrows eonstrueted by suspension
feeders or passive earnivores.
The Cruziana iehnofacies (Figure B2()) usually is associated
with infralittoral/shallow circalittoral marine substrates below
minimum wave base and above maximum wave base. The
trace fossils are characterized by: (I) high diversity; (2) low
individual densities; (3) a mixed association of vertical,
inclined, and horizontal structures; (4) presenee of struetures
produced by mobile organisms; (?) ihey are dominated by
feeding and grazing struetures eonstrueted by deposit feeders
some of whieh show quite eomplex behavior patterns resulting
in complex trace fossils (Pemberton cial.. 1992).
The Zoophycos ichnofaeies ideally is found in circalittoral to
bathyai. quiet-water marine tnuds or muddy sands, below
maximum wave base to fairly deep water in areas free of
turbidity Hows and subject to oxygen deficiencies. The trace
fossils are eharaeterized by: (1) low diversity, though
individual traces may be abundant; (2) grazing and feeding
structures produced by deposit feeders; and (3) horizontal to
gently inclined spreiten struetures (i.e., reptitious parellel or
concentric burrows or traces). However, the ichnofacies also
may occur in restricted intracoastal settings, particularly in
Paleozoic sequences; the ichnogenus Zoophycos possibly
represents greater depths of burrowing in Mesozoic and
Cenozoie deposits than in Paleozoic deposits (Pemberton
et al,,, 1992), hence the character of the Zoophycos ichno-
facies may vary from one part of the stratigraphic column lo
the next.
The Ncrciics ichnofacies typically is associated uiih bathyai/
abyssal, low-energy, oxygenated marine environments subject
to periodic turbidity flows. The trace fossils are characterized
by; (I) high diversity but low abundanee: (2) complex
horizontal grazing traces and patterned feeding/dwelling
structures; (3) numerous crawling/grazing traces and sinuous
feeal castings; and (4) structures produced by deposit feeders,
scavengers, or possibly harvesters (Ekdale ei al.. 1984). As
presently understood, the Nereites ichnofacies is restricted
primarily to flyseh or turbidite sequences; sediments in the
great expanses of sealloor beyond intluence of turbidity flows
eonsist chiefly of bioturbate texiures rather than discrete
traees hence there is no well-preserved record of specific
ichnocoenoses.
Substrate controlled ichnofacies
The remaining three ichnofacies represent speeialized,
substrate-controlled tracemakers and. environtnentally. are
very general in scope. The Glossifungites iehnofacies develops
in firm but unlithified substrates (i.e.. dewatercd muds) the
Trypaiiiies develops in fully eetnented substrates, and the Ter-
edoliics ichnotacies forms in woody substrates. Such horizons
may be critical markers in the evolving concept of sequence
stratigraphy (see Suhslraie-ConirolledIchnojacies),
Evaluation of the models
These archetypical models, particularly the marine ones, have
proven to be valuable indicators of general environmental
conditions. For instance, except for a capping layer of planar
bedforms, climbing ripples, or antidunes. primary physical
sedimentary structures of fluvial point bars may be strikingly
like those of estuarine point bars. However, biogenic
sedimentary structures are very different in the two settings.
Perhaps the most misunderstood aspect of these recurrent
ichnolacies is their use in paleobathymetry. Although some
workers have been complacent in this aspect of environmental
reconstruction (cf. Frey ctal,. 1990). various iehnologists have
long and persistently emphasized that local sets of environ-
mental factors are most important in controlling the distribu-
tion of traeemakers. whether or not these parameters oeeur at
speeilic water depths
Nevertheless, many kinds of environmental paiameters do
tend to change progressively with water depth and distance
from shore (grain size, energy levels, food resources, etc.). and
these gradients affect corresponding ehanges in the distribu-
tion of physical and biogenic sedimentary struetures. To
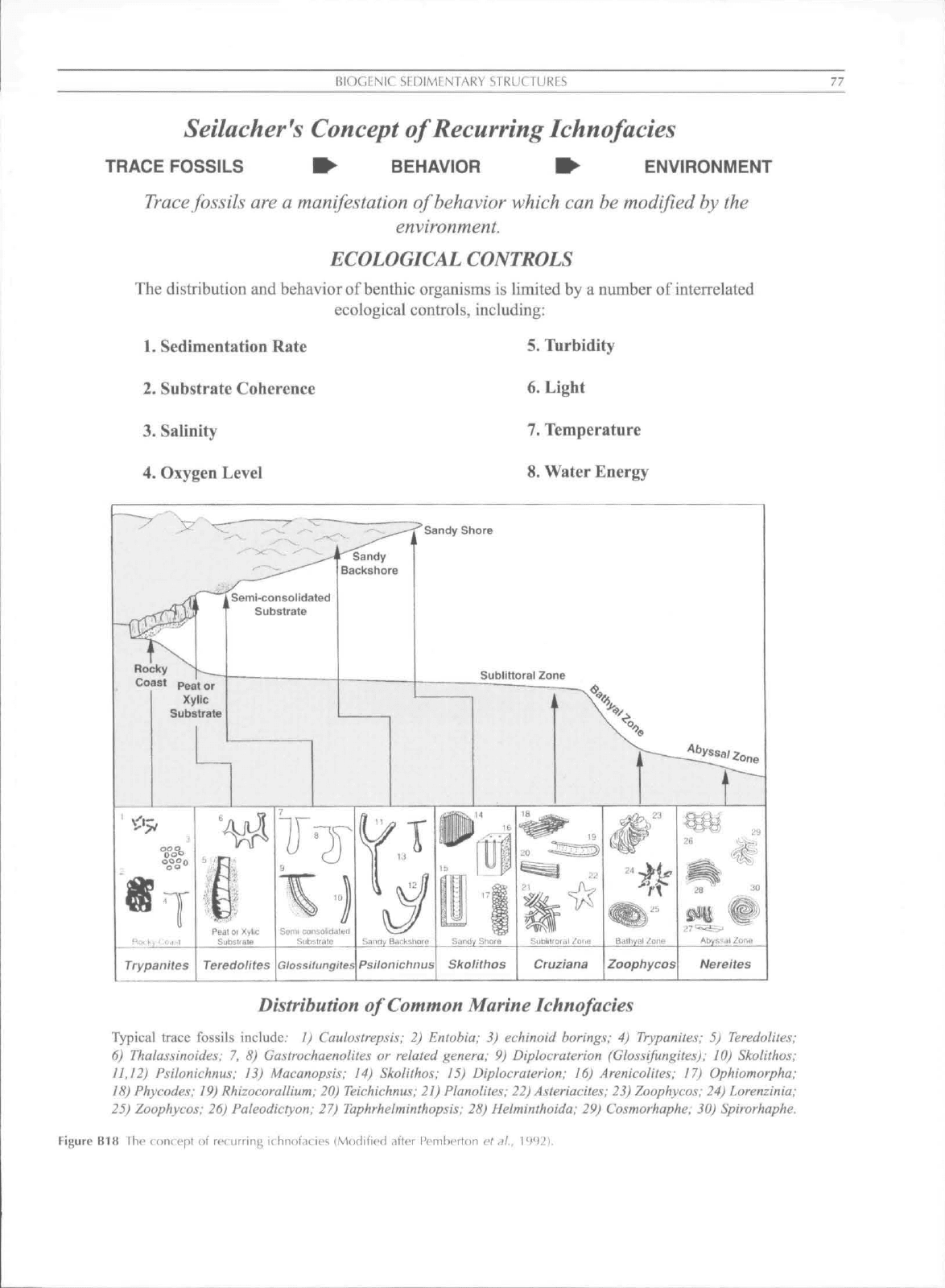
I^IOGFNIC
SEDIMFNTAKY STKUCTURFS 77
Seilacher's
Concept
of Recurring Ichnofacies
TRACE FOSSILS ^ BEHAVIOR W ENVIRONMENT
Trace fossils are a
manifestation
of behavior
which
can be modified by the
environment.
ECOLOGICAL
CONTROLS
The distribution and behavior of benthic organisms is limited by a number of interrelated
ecological controls, including:
1.
Sedimentation Rate
2.
Substrate Coherence
3.
Salinity
4.
Oxygen Level
5. Turbidity
6. Light
7. Temperature
8. Water Energy
Rocky
Coast Peat or
Xylic
Substrate
Distribution of
Common
Marine Ichnofacies
Typical trace fossils include." 1) Caulostrepsis; 2) Entohia; 3) echinoid borings; 4) Trypanites; 5) Teredolites;
6) Thalassinoides; 7. 8) Ga.strochaenolites or related genera; 9) Diplocraterion (Glossifungites); 10) Skolithos;
11.12) Psilonichnus; 13) Macanopsis; 14) Skolithos: 15) Diplocraterion; 16) Arenicolites; 17) Ophiomorpha;
18) Phycodes: 19) RhizocoralUum; 20) Teichichnus; 21) Planolites; 22) Asteriacites; 23) Zoophycos; 24) Lorenzinia;
25) Zoophycos; 26) Paieodictyon; 27) Taphrheiminthopsis; 28) Helminthoida; 29) Cosmorhaphe; 30) Spirorhaphe.
Figure B18 The concfpi of recurring ithnotaties (Modified ,ifter Pemberton et ,il., 1992).

78
BIOGENIC SEDIMENTARY STRLICTURES
Association
Skolithos
Ichnofacies
irenicolites
variabilis
'Skolithos linearis
Figure B19 The different expressions of the Skolithos ichnofacies (Modified from Pemberlon w ^/.,
20021.
thai extent, trace fossil associations arc indeed useful in
paleobathymetry.
Equally important is the long temporal duration of most
kinds of trace fossils. These basic benthic behavioral
patterns arc more nearly like stable ecologic niches than
individualistic records of particular animal species (Frey and
Scilachcr. 1980). As long as the functional niche remains
advantageous under given environmental conditions, many
different animal species, over long intervals of geologic time.
may be expected to exploit it: their preserved traces are
strikingly similar and have equivalent significance. Hence,
although we conveniently speak of the "SkolUhos animar" as
the architect for a particular kind of dwelling structure,
numerous dilTercnt animal species actually were involved. The
longevity of recurrent ichnofacies thereby exceeds the long-
evity ol recurrent biofacies by a considerable margin, and are
correspondingly more useful as archetypical models not only
for environmental interpretation but also for comparisons of
depositionat environments of widely differing ages.
The purpose for recognizing these recurrent ichnofacies must
not be overlooked. Interpreted in terms of the original trace
(bssil assemblage, these iehnofacies are merely archetypal
facies models with which the local ichnofacies may be
compared. The archetypes are intended to supplement, not
supplant, local ichnofacies designations, some of which are
quite distinctive.
The idealized ichnofacies succession works well in most
"iiormar" situations (Krey and Pemberton. 1987). including
distributions according to salinity gradients (cf. Bromley and
Asgaard. 1991); yet one should not be surprised to find
nearshore assemblages in offshore sediments, and vice versa,
for example, if these accumulated under conditions otherwise
like those preferred by the tracemaking organisms. The basic
consideration rests not with such inanimate backdrops as
water depth or distance from shore, or some particular tectonic
or physiographic setting, but rather with such innate, dynamic
controlling factors as substrate consistency, hydraulic energy.
rates of deposition, turbidity, oxygen and salinity levels, toxic

BIOCCNIC SEDIMENTARY STRUCTDRFS
79
CRUZIANA ICHNOFACIES
Figure B20 SchenLJlic diagram of
^i
typit.il Mesozoic example of the
Cruz'hin.i
ichnofacies (Modified from Pcniherton
cl.?/.,
H){M).
substances, the quality and quantity of available food, and the
ecologic or ichnologic prowess of tracemakers themselves.
Finally, ihc models should not be divorced from associated
patterns of bioturbatioii. Numerous local ieiinofaeies. parti-
cularly those representing low-energy conditions cind slow
rates of deposition, are set in a eomplex biogcnie fabric.
Several generations of burrows may be discernible via their
crossctittinjj relationships, showing that the same volume of
sediment passed repeatedly through various styles of rework-
ing. In environinental reeonstruction, such ichnologic fabrics
may be equally as important as the individual, named trace
lossils (Frey and Pemberton. 19S4).
Paleoenvironmenta! significance of trace fossils
I he coneept of lu net lonal morphology,
a
basic premise employed
by ecologisls and palcoeeologisls in en\ironinenlal reconstruc-
tion, is equally applicable to iehnology. In faet, trace fossils are
unique in that they represent not only the morphology and
ethology of the tracemaking organism but also the physical
characteristics of the substrate; they are elosely linked to tlie
environmenlal eonditioiis prevailing at the time of their
eonstruetion. Frey and Seilacher (I'JSd) re-emphasized that sueh
variables as bathymetry, temperature, volumes of sediment
deposited or eroded, aeration of water and sediment, and
substrate coherence and stability, bave a profound effect on
resultant traee fossil distributions and morphology, and hence
can be used in the determination of original biological.
ethnological, and sedimcntologicai conditions.
The applieation of iehnology to paleoenvironmental analy-
sis goes far beyond the mere establishment of gross or
archetypal ichnolacies. For instance, shallou-water, eoastal
marine environments comprise a multitude of sedimentologieal
regimes, which are subject to fluctuations in many physical and
ecological parameters (see Coastal Sedimcniary Facies). In
order fully to eomprehend the depositional history of such
zones in tbe rock reeord. it is imperative to have some reliable
means of differentiating subtle ehanges in these parameters.
Detailed investigations ol' many of these coastal tnarine zones
in Georgia have shown the value of utilizing biogenic
sedimentary struetures (in eoneert with physieal sedimentary
structures) in delineating them (Frey and Pemberton, 1987).
The application of these studies in deeiphering paleoenviron-
tnents has also proven invaluable. For example, the shoreface
consists of a seaward-sloping sediment wedge extending from
the low tide mark, generally to the fairwealher (minimum)
wave base, corresponding to approximately l()-2()m of water
depth. The shorefaee setting is dominated by wave energy and.
as a result of decreasing uave interaction uith the substrate in
a seaward direetion. shows a pronouneed basinward fming.
The shorefaee is elassically div ided into three subenvironments
(from seaward to landward): the lower, the middle and the
upper shoreface. Tbe boundaries between these are not always
eiearly defined (Reiiison. 1984). Although the large-scale
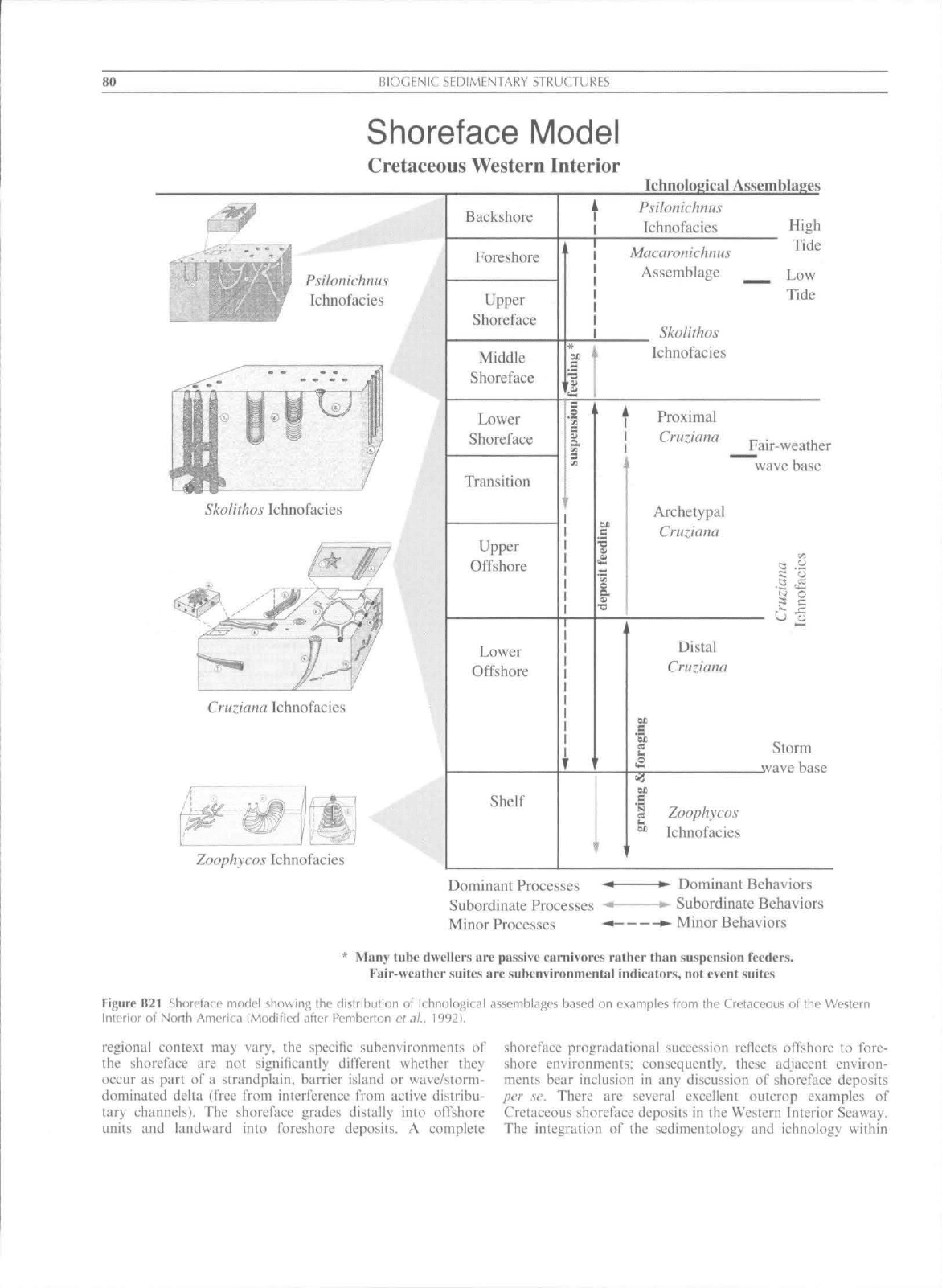
80
BIOCENIC SEDIMENTARY STRUCTURES
Shoreface Model
Cretaceous Western Interior
Ichnological Assemblages
Psilonichnus
Ichnofacies
Skolithos Ichnofacies
Cruziana Ichnofacies
Backshore
Foreshore
Upper
Shoreface
Middle
Shoreface
Lower
Shoreface
Transition
Upper
Offshore
Lower
Offshore
Zoophycos Ichnofacies
Psilonichnus
Ichnofacies
Mucaronichnits
Assemblage
Skolithos
Ichnofacies
High
Tide
Low
Tide
Proximal
Cruziana
Archetypal
Cruziana
Fair-weather
wave base
"3
Distal
Cruziitmi
Storm
_ivave base
S Zoophycos
^ Ichnofacies
Dominant Processes
Subordinate Processes
Minor Processes
Dominant Behaviors
Subordinate Behaviors
Minor Behaviors
* Many tube dwellers are pa.ssive earnivores rather than suspension feeders.
Fair-weather suites are subenvirunniental indicators, not event suites
Figure B21 Shoreface model showing the distribution uf Ichnological assemblages based on examples from Ihe Cretaceous of the Western
Interior of North America (Modified after Pemberton ef
.1/.,
1992).
regional context may vary, the specific subenvironments of
the shorefaee are not signifieantly different whether they
occur as part of a strandplain. barrier island or wave/storm-
dominated delta (free from interference from active distribu-
tary channels). The shoreface grades distally into offshore
units and landward into foreshore deposits. A complete
shorefaee progradational suceession reflects offshore to fore-
shore environments: eonsequently. these adjaeent environ-
ments bear inclusion in any discussion of shorefaee deposits
per se. There are several exeellent outerop examples of
Cretaceous shoreface deposits in the Western Interior Seaway.
The integration of the sedimentology and iehnolugy within

Crevasse Splay
Resriciec
Brackish Bay
Delta Front
^^Ulracklsh
Bay Model
Cot^monly recurring ichnofacies
Figure B22 Schem<ilic
et ,il
of typittit rwurring Irate fossil jssembltij;es in hr.u kish water li.iy-like environments iModified from Pemberton

82 BIOCENIC SEDIMi^NTARY STRUCTURES
these deposits affords the opportunity to ehanieterize the faeies
and facies suceessions (Figure B2!). and to explain the
observed facies variability.
Hudson elal. (1995) recently stressed that Jurassie benthie
mollusean assetnblages in the Great Estuarine Group of Great
Britain are controlled largely by salinity with different genera
or groups of organisms characterizing a spectrum of tnainly
brackish water environments. As pointed out by Pemberton
and Wightman (1992) salinity gradients also iittluenee the
nature of the ichnofauna. Although some behavioral patterns
may occur on each side of the salinity transition, distinct
ichnoeoenoses are present in freshwater, braekish water and
fully marine environments. Recently, marginal marine envir-
onments (including tidal channels, estuaries, bays, shallow
lagoons, delta plains, etc.) have been recognized with more
frequency in the rock record. Such environments character-
istically display steep salinity gradients, which, when combined
with corresponding changes in tetnperature. turbulence.
exposure, and oxygen levels, result in a physiologically
stressful environment for numerous groups of organisms.
Brackish water bay-like environments include bays, sounds.
lagoons, inter-distributary bays, and the central basins of
wave-dominated estuaries to name just a few (Figure B22).
One of iehnologys greatest strengths, the bridging of
sedimentology and paleontology, in some respects can be its
greatest liability. Sedimenlologists tend to tise a striet
uniformitarian approaeh to paleoenvironmental interpretation
and rely heavily on modern analogues. Paleontologists, on the
other hand, must temper their observations in the light of
organic evolution. Although trace fossils can be considered as
biogenie sedimentary structures and are difficult to classify
phylogenetically. they are constructed by biological entities
and are thus subjected at least to some degree to evolutionary
trends. For example, oeeurrenees of well-developed terrestrial
trace fossil assemblages are much more prevalent in post-
Cretaceous rocks. This development corresponds to the
evolutionary explosion of the insects, brought on by the
diversification of the angiosperms in the Late Cretaceous.
Prior to this time terrestrial substrates may not have been as
extensively bioturbated due to a paucity of tracetnakers.
Likewise, patterned grazing traces, which characterize deep-sea
environments, show a trend toward more eomple.\ organiza-
tion through most of the Phaiierozoie. This trend may be
related to the evolution of more effieient foraging strategies
(Seiiaeher. 1986). For these reasons, paleoenvirontnental
interpretations based on trace fossils must be considered not
in striet uniformitarian terms, but rather, in actualistic ones.
Equally important, unique quantitative environmental in-
dieators are indeed rare in the geologieal reeord. and ichnology is
no exception (Frey and Seilacher. 1980). However, traee fossils
can supply a wealth of environmetUal information that eannot
be obtained in any other way and v\'hieh should not be ignored.
Their potential usefulness is accentuated when fully integrated
with other (chemieal. physical, and biological) lines of evidence.
Combined studies of physieal and biogenic sedimentary
structures constitute a powerful approach to facies analysis.
Summary
Iehnology and its significance to sedimentai'y geology is based
on the following concepts:
(a) Biogenic structures represent the activity of soft-bodied
organisms that are not generally preserved. Such
organisms (including many entire phyla) are commonly
the dominant component of the biomass of tnany
environments.
(b) Biogenie struetures are eommonly enhaneed by diagenesis
and ean be used in horizons where physical sedimentary
struetures have been masked. For example, in oil sands
deposits bitumen staining obliterates mosi physical struc-
tures,
but due to the coneeutration of clay minerals, it
enhances the visibility of biogenie structures.
(c) Biogenie struetures can be assoeiated with facies that do
not eontain any other fossils. In many silieiektstie regimes,
diagenesis dissolves most of the shelly fauna and trace
fossils represent the only clue as to the original biogenic
component of the unit.
(d) Biogenic structures can be used for the paleoecological
reconstruction of depositional environments.
(e) Biogenic struetures are sensitive to fluctuations in sedi-
mentary dynamics and are important in recognizing event
beds and distinet sedimentation patterns.
(0 Biogenie struetures are sensitive indieators of substrate eohe-
renee and substrate-controlled iehnofaeies are emerging as
important elements in genetie stratigraphie paradigtns.
(g) Biogenie struetures ordinarily eannot be transported and
therefore represent the original environmental position of
the trace making anitnal.
(h) Biogenic structures are sensitive to changes in certain
ecological paratiieters that are otherwise difficult to
ascertain sueh as salinity and oxygen levels.
(i) An integrated approach utilizing physieal, chemical.
biological, and ichnological lines of evidence constitutes
a powerful tool for facies Interpretation.
S. George Pemberton
Bibliography
Bromley. R.G.. 19'J6. inuc
Fossils.
Biohii;)-
tiiul laplioiumiv. 2tii.i edn.
London: L'nwiii Hyman.
linimlcy. R.G,. and Asgaard. LI,. 1991, Iclinofaeies: a mixiiirc of
lapliofacies ;ind biofacies. Lctluiiti. 24: 153 163.
BLiatois. L.A.. and Mangano. M.G., 1995. The pahioocnvironnicntal
and palacoccological significance of the Mermia idinotaeics: an
archtflypiil subaqui;ous non-marint" tract" fossil assemblage, kimos.
4: I5i-I6i.
Ekdale. A.A.. 1985. Palcoecology of the marine endobcntlios. Piihico-
i^c'ogrtiphv. P<ihii-orlimeitologY. Piil(icocci>lo!;v. 50: 63 81.
Ekdale. A.A.. Bromley. R.Ci.^aiid Pembt.'rlon. S.G., 1984.
Icliiiolos-y:
Till'
L'.sc
of
Trace
Fossils in St'dimi'iitolof'y and Stratigraphy. Society
of Economic Paleontologists and Mineralogists, Short Course
Notes 15-
F'rey. R.W.. and Pemberton. S.G,. 1987, The Psiloiikhnti.s ielinocoe-
iiosc and its tclationsliip lo adjacent marine and nonmarine
ichnocoenoses along the Georgia coast.
Bulletin oJ
Camtditiii Petro-
leum Geohi-v. 35: 333 357.
Frey. R.W.. and Seilacher. A.. 1980. Uniformity in marine invcrtebtate
ichnology, Lt'tliaia.n: 183-207.
Frey. R.W.. Pemberton. S.G.. and Fagerstrom, J.A.. 1984.
Morphologieal. ethologieal. and environmental significance ol" the
iehnogenera Saivenia and Aiuorivhniis. Jiiiiriialofl'dlcniitolof-v. 58:
511 528.
Froy. R.W.. Pcmbi-rton. S.G.. and Sauncfers. T.D.A.. 1990. Ichiio-
fiieies iind batliymctry: a passive relationship. Jaiiriuil of
Paleontology. 64: 155-158.
Gingras. M,K,. Pemberton, S.G.. Mendoza. C. and Henk. B.. 1^99.
Modeling fiuid flow in trace fossils; assessing the anisotropie
permeability of
G!o.s.\ifungilex
surfaces. Petroleum
GcoscU-nces.
5:
349-357.

ULACK SHALES
83
Hudson. J.D.. Clements.
R.(}..
Riding. J.B.. Waketicld. M.I.. and
Walter. W.. 1995. Jurassic paleosalinities and brnekish-water
eommiinities a ease study. Palaios. 10: 392-407.
Pemberton. S.G.. MacEachern. J.A.. and Frey. R.W.. 1992. Traee
tbssi! tacies models: environmental and allostratigrciphie signili-
canee. In Walker.
R.C}..
and James. N. (eds.),
Fa<U:\
\todi-ls:
Response
10SeaLcniCiiiini;i\ St. Johns. Newlbundknid: Geological
Association of Canada, pp. 47-72.
Pemberton. S.G.. Spihi. M.V.. Piilluim. A.J.. Sannders. T.. MaeEa-
chern. J.A.. Robbins. D.. and Sinclair. 1.. 2001.
Ichnology
and Sc-
dinn'moloiiy of Sludlow and Marginal Marine .SV.v/t'/».v.* Ben
Ni'vis and Avalon Reservoirs. Jeanne
D.4rc
Btisin. St. Johns. New-
t\iund!and: Geological Association
ol"
Canada. Shori Conrse Noles
15:
353 p.
Pemberton. S.G.. and Wightman. D.M.. IW2. lehnological charaeler-
istlcs of brackish water deposits. In Pemberton. S.G. (ed.).
.-ippli-
cations of Ichnoloi^y to Pctroh-iini Fxploralion. Inlsa: Society of
Economie Paleontologists and Mineralogists. Core Workshop 17.
pp.
141 167.
Reinson. G.F. 1984, Barrier-island and associated strand-plani
systems. In Walker. R.G, (ed.).
Facic.\
MoclcLs.
.si-cond
edition. Si.
Johns.
NewloLindland: Geologieal Assoeiation of Canada, pp, 119-
140.
Seilacher. A. l9S(i. Evolution of behavior as expressed in marine irace
fossils. In Nitecki. N.W.. and Kitchell. J.A. (eds,|. F-vohaionofAn-
imal Behavior. New York: O.Klbrd University Press, pp. 62 87.
Smith. R.M.H.. Mason. T.R.. and Ward. L.F.. 199.V Flash flood
sediments and ichnofacies of the Late Pleistocene Homeb Sills.
Kuiseb River. Namibia. SedimenkiryGeology. 85: 579 599.
Cross-references
Aman/GressK (1S14 1S65)
f-acies Models
Johannes Waltlier( 1860 1937)
Rudolf Richter (ISSI 1957) and the Senckcnbcrg Laboratory
Substi ale-Controlled Ichnofacies
Taphonomy: Sedimenlological Implications of Fossil Preservation
BLACK SHALES
HIack Shitlcs iirc line grained, gcncriilly orgatiic carboii-rich
scilimcntary rocks that primarily consist of a mixture of clay
iiiincriils. quartz silt, organic particles (mostly plaiiktonic algae
atid plant debris), and kerogen. They may also contain
viiriablc amounts of disseminated finely crystalline calcite
and dolomite, as well as phosphate (eommonly as concretions).
Most black shales arc found in marine seditncnts (Potter elal..
1980).
but they can also form prominent deposits in lacustrine
successions (Boliacs elal.. 2(K)()). Their black color is due to
Iwo constituents: (I) the contained organic matter, and
(2) finely disseminated pyrite. The reducing conditions
indicated by the latter have long led geologists to believe that
aneient black shales required anoxie bottom waters for their
formation, and were a typical deposit of the distal, deepest
portions of sedimentary basins (via comparisons with the
abyssal Black Sea where earbonaceous muds currently
accumulate).
The Black Sea is a stratified, silled basin, in which a lower
marine water body is overlain by a layer of brackish water due
large input of freshwater frotn rivers. Beeause of density
contrasts a halocline is developed at a depth of approxirnately
20(1
m, severely restricting vertieal advection of oxygen-rich
surface waters. Oxygen demand by decaying and descending
organic matter exceeds oxygen replenishment by \crtical
advection. rendering the water column beneath the haloeline
anoxie and facilitating the accumulation of organic-rich muds.
These observations have suggested to many geologists that
organic earbon enriehment in the bottom sediments was
basically a result of enhanced preservation (Wignall. 1994).
Although the Black Sea model dominated the study of blaek
shales for many years, it is of limited use vvhen studying
ancient black shales. A major difference between the Black Sea
and practically all black shales in the rock reeord is that the
substantial depth (-200()m) of the former, in conjunction
with a stable halocline. greatly restriets eireulation, whereas
ancient black shales accumulated in much shallower epicontj-
nental seas with a much larger width/depth ratio. Thus, on a
geologic timescale at least, stortns should have caused frequent
mi\ing of the water column. With continued research into the
origin of black shales, many occurrences that were initially
interpreted according to the Black Sea model, are now in the
process of re-evaluation.
Evolving views of the origin of the Late Devonian
Chattanooga Shale of the eastern US may serve as an example
for these changing perspectives in blaek shale deposition. On
the basis of well developed line lamination and apparently little
evidence of benthie life, it was iititially thought of as a very
good representative of a basin with a stratilied water column,
and enhanced preservation as the primary cause for black
shale accumulation. Recent research, however, has uncovered
abundant evidence for wave reworking and intermittent
erosion (Figure B23) of black shale (Schieber cial., 1998).
The observation that erosion surfaees of the type illustrated in
Figure B2? can be traced for hundreds of kilometers actually
forms the basis for a sequence stratigraphic inierpretation of
this succession (Schieber. 1998). Careful examination ol'shale
fabrics has also revealed SLibtlc but nonetheless widespread
evidence (bioturbatioii features triostly) for benlhic coloniza-
tion of the sealloor. The high organic carbon contents in the
Chattanooga Shale (up to 20 per cent) initially suggest that
preservation due to oxygen restriction may have been mainly
what controlled its accumulation. Yet. sedimentary and
bioturbation features suggest that anoxia, if they indeed
occurred, were intertnittent. due to frequent water cokmin
mixing by storms or seasonal temperature variations. In the
case of the Chattanooga Shale, therefore, surface productivity
probably was also an important factor for organic carbon
enrichment (Schieber t'/i//.. 1998).
Although the "deep anoxie basin" model has enjoyed
prominenee for many years, when we look at blaek shales
more closely we find increasingly that they can actually form in
a wide range of depositional settings. Black shales do not
constitute a large proportion of the sedimentary rock column,
but they are of great eeonomic interest as the main source rock
for hydrocarbon production, as well as containing unusually
high metal concentrations (Potter ei al.. 1980). Fstimates
suggest that tnorc than 90 pereent of the world's recoverable
oil.
and gas reserves were generated from blaek shales
(Klemme and Lllmishek. 1991). Equally significant is the fact
that the latter are restricted to six stratigriiphic intervals that
represent a mere one third of Phanero/oic time (Silurian.
Upper Devonian-Tournaisian. Pennsylvanian-Lower Permian.
Upper Jurassic. Middle Cretaceous. Oligocene-Miocenc).
Because organic carbon results from photosynthesis by
plants and algae, each atom of carbon buried implies a
molecule of oxygen added to the atmosphere. Carbon burial is

BLACK SHALES
Figure B23 Erosion surfaces in the ChatLinoogd Shale (entire exposure consists of black shale). Picture shows truncation of black shale packages
by erosion surfaces (dashed white lines marked wilh \jr^e white arrows), and illustrates how succeeding black shale packages are draped
conformably over these prosion surfaces. Small white arrows point out truncation o! shale beds by erosion siirfaco. In places erosion has removed
in excess of
1
m ot black shale befween successive block shale packages (at a total Chaltanooga thickness of less than 10m).
linked to other global biogeochcmical cycles and is a critical
variable in our attempts to understand the history and
evolution of the oceans and atmosphere, as well as global
climate change. Over geologic time, carbon burial probably
was responsible for a gradual rise in atmospheric oxygen levels.
Also,
by reducing the greenhouse effect it can lead to lower
global temperatures and even iee ages (Berner. 1997). In that
context, considering that the geological record is punctuated
by global episodes of widespread black shale formation
(Klemme and Ulmishek. 1991). understanding what cornbina-
tions of variables are required to produce a "black shale
world"
is of considerable significance.
Three factors control organic matter accumulation in
sediments: (1) organic matter input: |2) organic matter decay:
and i?) dilution oforgauic matter by other ingredients. Factor
I. organic tiiattcr input, has two components, dctrilal organic
matter washed in by rivers and organie matler from primary
production in the water column. The latter is strongly
dependent on nutrient availability. Factor 2 relates to break-
down of organic matter, mostly by microbes. Its efficiency and
the types of bacteria involved depend on oxygen availability in
the water column and the sediment. Factor 3. dilution, has a
terrigenous component (fluvial and eolian contributions), and
an intrabasinal component of skeletal remains
(e.g..
coccoliths,
radiolaria, foraminlfera. diatoms, etc.). The fluvial cotnponent
is linked to climate and relief in the source region, and to sea
level fluctuations. Because the magnitude of this component
rises and falls with continental runotT, it is directly linked to
the nutrient input that impacts primary production. Relief and
sediment supply are linked to tectonism. which in addition can
affect subsidence rates, basin gcotnctry, and water depth
(Schieber
etal..
I99S). Because each factor is a function of
several other variables that are in part cross-linked to other
factors, determining the magnitude of each of these factors in
the rock record is far from trivial. Obviously, black shale
formation is a multifacetcd problem and depending on the
interaction of the variables, there are multiple scenarios that
are conducive to black shale formation (Tyson and Pearson,
1991). Microbial mats can also promote accumulation, and
preservation of organic-rich sediments in a wide range of
environments (Schieber, 1999). a currently under appreciated
fact (see Microbially IndmetlSeilinwulary Siruclures).
Black shales are not only common in thick, basinai.
successions, but also occur as thin tongues within basin-
margin successions. The latter are thought to owe their
existence to high stands of sea level when terrigenous input
was at a minimum, and as such may mark maximum flooding
surfaces (Creaney and Passey, 1993). Whether these shallow
water black shale tongues reflect deposition beneath a
nearshorc zone of oxygen restriction, or simply are the outer
edge of an expanding anoxie water body that tisually only
occupies the basin center (Wignall. 1994). is still being debated.
Nearshore oxygen restriction could, for example, be produced
by salinity stratiiication due to freshwater plumes, or by high
nearshore productivity fueled by nutrients in continental
runoff. Alternatively, rapid sea-level rise could have caused
expansion of central anoxie water bodies into shallow
nearshore waters. Recent efforts to integrate black shales into
a sequence stratigraphic context suggest that at the onset of
transgression, black shale deposition may commence in the
basin interior, that nearshore black shale deposition is possible
early in transgressions, and that black shales related to
maximum flooding can form during high stands of sea level
(Wignall.
1994: Schieber
elal..
1998). Sea level variations may
also have lead to conditions with recurring water column
mixing events and recycling ol" key nutrients (N, P) to surface
waters, making possible a '"productivity-anoxia feedback"
mechanism to maintain high rates of carbon burial (Ingall
i'lal..
1993).
Black shales are the end product of the complex interplay
among a range of geologic variables and processes, and there
are no easy answers and no "one size tits all" rnodels. The
seemingly drab and uniform nature of many of these rocks
poses a considerable challenge and requires us to extract