Ильичев В.Г. Устойчивость, адаптация и управление в экологических системах
Подождите немного. Документ загружается.


118
образом как функция k . Теперь заметим следующее:
а) согласно допущению
(
)
1W
находится выше
1
J ;
б) при малых
()
0+→kk
поверхность
(
)
kJ
1
не исчезает, а остальные
(
)
kJ
1
исчезают. Значит,
()
0+W
заполняет все
n
R
+
и, в частности, включает в себя
1
J .
Далее, проведем в
n
R
+
"биссектрису" (прямую
n
yyy
=
=
= K
21
).
Единственную точку пересечения
W
и биссектрисы назовем "центром" границы
клина. В силу непрерывности деформации, при некотором промежуточном
*
k
центр клина пересекает (или касается) поверхность
1
J в какой-то точке
*
Y из
n
R
+
.
Такая ситуация полностью соответствует случаю 1, который нереализуем.
Р
ис. 4.4. Клин (затемнен) и специальная точка
*
Y .
Итак, в обоих случаях возникает противоречие. Поэтому сделанное
допущение неверно. Значит,
1
J лежит выше всех остальных изоклин.
119
ГЛАВА 5.
ПЕРЕСТРОЙКА ПЕРЕМЕННЫХ КАК МЕХАНИЗМ АДАПТАЦИИ.
АНАЛИЗ ЭКОСИСТЕМЫ АЗОВСКОГО МОРЯ
В каждой области исследования люди cтремились
понять явление путем поиска его элементов. В
каком-то смысле эпоха машин была крестовым
походом, а его Святым Граалем стал Элемент.
(Акофф Р.Л. О менеджменте )
Для получения устойчивых характеристик в поведении экологических
систем целесообразно проводить модельные расчеты на длительную перспективу.
Действительно, асимптотические значения биологических компонент, как правило,
не зависят от неизбежных погрешностей в выборе начального состояния.
Далее, в отличие от технических систем в экологических системах
действуют различные механизмы адаптации и микроэволюции. Их необходимо
учитывать при проведении компьютерных расчетов на длительную перспективу, в
противном случае в модели будут получаться заниженные оценки биологической
продуктивности.
Обсудим основные виды приспособительных реакций (Гапочка,
1981;Тимофеев - Ресовский, Свирежев, 1966; Шилов, 1985; Dobzhansky, 1970). Они
бывают двух типов:
1) изменение переменных, связанное с переходом из старого равновесия в
новое. Модель такого механизма адаптации водорослей к содержанию биогенных
веществ будет продемонстрирована в разделе 5.1;
2) изменение параметров в результате действия процесса естественного
отбора (см. ниже главу 6).
Последний вариант адаптации является эколого - эволюционным, и
позволяет по - новому рассмотреть проблему моделирования динамики экосистем.
А именно, в экосистемах изменение переменных сопряжено с динамикой
параметров. Поэтому при описании поведения экосистем следует
руководствоваться не традиционными моделями типа:
),(
α
xfx
=
&
,
120
а более корректными моделями типа:
),(
α
yFy
=
&
,
),(
α
ε
α
yG
=
&
,
где y - вектор переменных, являющийся расширением вектора x с учетом
внутривидового полиморфизма природных популяций;
α
- вектор параметров; F -
"естественное" расширение оператора f, учитывающее процессы внутривидовой
конкуренции; G - оператор, производящий селекцию параметров;
ε
- скорость
микроэволюции (поскольку скорость изменения параметров существенно меньше
скорости изменения переменных, то
ε
мало).
Достоинством последней схемы является то, что она (в идеале) не имеет
параметров. По - существу, вектор параметров
α
сам "автоматически"
идентифицируются в процессе длительной работы модели.
5.1. Модели адаптации водорослей к содержанию азота и фосфора в среде
В модели круговорота азота (N) и фосфора (P), предназначенных для
исследования асимптотического поведения экосистем, необходимо включать
механизмы адаптации водорослей к содержанию данных биогенных веществ в
среде. В противном случае при модельных расчетах может “возникать”
неограниченный рост N или P. Покажем это. В качестве иллюстрации проведем
анализ динамики N и P в озерной упрощенной экосистеме “водоросли – азот и
фосфор”. Основными элементами баланса веществ являются:
1) приходная часть. Считаем, что в озеро впадает река - носитель
N
и
P
в
количестве и (в единицу времени);
N
c
P
c
2) расходная часть. Часть отмерших водорослей (и соответственно, N и P)
поступает в донные отложения.
Положим =
N
Z
N
минеральный
+
N
органический
+
N
в водорослях
.
Очевидно, величина характеризует суммарный запас азота в экосистеме.
Аналогично, определим переменную , которая отвечает за суммарный фосфор.
N
Z
P
Z
Обозначим и - содержание
N
k
P
k
N
и
P
в единице биомассы водорослей.
Тогда в
рамках непрерывной схемы суммарные запасы азота и фосфора
описываются соотношениями
121
, , (1.1)
DkcZ
NNN
−=
&
DkcZ
PPP
−=
&
где - cкорость захоронения (части) водорослей в донные отложения.
)t(DD =
Положим и
PPNN
k/Zk/Zw −=
PPNN
kckc //
−
=
δ
. Из (1.1) вытекает
δ
=
w
&
.
Очевидно, при имеем
PPNN
k/ck/c >
0>
δ
и, значит, . Поэтому
суммарный азот неограниченно возрастает. Напротив, при
∞→w
PPNN
k/ck/c
<
.
величина суммарного фосфора стремится к бесконечности.
В обзоре Фурсовой П.В. и Левича А.П. (2002) рассмотрены различные
модельные схемы адаптации водорослей к содержанию азота и фосфора.
В данной работе рассмотрен новый механизм адаптации, основанный на
концепции Ф. Добжанского (Dobzhansky, 1970) о жестком полиморфизме
природных популяций. Следуя этой идеи, всякая (”исходная”) популяции
водорослей состоит из набора близких субпопуляций, каждая из которых имеет
“свои” параметры . При этом потребление веществ азота и фосфора i-той
субпопуляцией пропорционально их внутриклеточному содержанию (т.е.
величинам ). Субпопуляцию с наибольшим будем называть
“азотолюбивой”, а с наибольшим - “фосфоролюбивой”. Исход конкуренции
данных субпопуляций зависит от содержания азота и фосфора в среде. Это
взаимодействие должно быть согласовано с принципом компенсации Ле – Шателье
– Брауна. Так, при больших
},{
PN
kk
i
P
i
N
kk ,
i
N
k
i
P
k
N
должна доминировать азотолюбивая субпопуляция,
поскольку в этом случае происходит наиболее сильное потребление N.
Аналогично, при больших
P
должна доминировать фосфоролюбивая
субпопуляция.
В качестве базовой модели динамики концентраций (
x
) популяции
водорослей используем однородную схему Пирcа (Pearce, 1970)
)N/xk,P/xk(xfx
NP
=
&
. (1.2)
Здесь
P
и
N
- концентрация минеральных форм фосфора и азота в воде;
гладкая трофическая функция убывает по каждой переменной.
),( vuf
В модели экосистемы Азовского моря (Ильичев, 1986а, 1986б, 1995а)
122
использовался частный случай схемы Пирса - модель Контуа, в которой
)vu1/(1)v,u(f +++−=
β
. В развернутой форме здесь получаем
)]PkNk(xPN/[PN1)N/xk,P/xk(f
NPNP
+
+
+
−
=
β
. (1.3)
Здесь
β
- скорость роста водорослей, зависящая от температуры и солености
среды; скорость смертности равна -1.
Пусть условия среды постоянные, тогда при
1>
β
в модели (1.2) с правой
частью (1.3) имеется единственное, положительное равновесие , где
)k,k(X
PN
]k/Pk/N/[)1()k,k(X
PNPN
+
−
=
β
. (1.4)
Данное равновесие глобально устойчиво в .
+
R
Важно еще раз напомнить, что в схеме Пирса правая часть является
однородной функцией первой степени от переменных
N
,
P
,
x
. Поэтому переход от
валовых значений содержания водорослей, азота и фосфора к их концентрациям
“не портит” коэффициенты правой части модели (см. глава 1). Это позволяет без
изменения переносить идентифицированные в лабораторных экспериментах
“свободные” коэффициенты функции на модели природных экосистем.
f
Теперь построим расширение модели (1.2), включающее в себя три
субпопуляции ( ). Определим внутриклеточное содержание азота и
фосфора в данных субпопуляциях следующим образом:
321
x,x,x
1) и ;
NN
kk =
1
PP
kk =
1
2) и ;
ε
NN
kk =
2
ε
/
2
PP
kk =
3) и .
ε
/
3
NN
kk =
ε
PP
kk =
3
Здесь
ε
- некоторое число, большее 1. Первая субпопуляция – носитель параметров
исходной популяции; вторая – азотолюбивый мутант; третья - фосфоролюбивый
мутант. Взаимодействие всех субпопуляций описывается системой (“Триадой”)
, (1.5)
)N/Sk,P/Sk(fxx
i
N
i
Pii
=
&
где ;
3,2,1=i
321
xxxS ++
=
. Пусть - трофическая функция Контуа, тогда в
“Триаде” выполняется простое
f
Утверждение 5.1. При
PN
k/Pk/N
ε
>
123
вторая (азотолюбивая) субпопуляция вытесняет остальные.
При
PNP
k/P)/1(k/Nk/P
ε
ε
<
<
первая субпопуляция вытесняет остальные.
При
NP
k/Nk/P
ε
>
третья (фосфоролюбивая) субпопуляция вытесняет остальные.
В имитационной модели “водоросли – азот и фосфор” Азовского моря
представлены три основных вида - диатомовые, пирофитовые и синезеленые -
водорослей. Каждый вид водорослей описывается модельной конструкцией
"Триада". Действие "Триады" не устраняет возможности возникновения дисбаланса
азота и фосфора в среде, а лишь смягчает его.
Заметим, что нелинейный рост в модели Контуа
)(
NP
PkNkxPN
PN
++
β
,
является некоторым “приближением” для мультипликативной формулы
NP
xkN
N
xkP
P
++
β
.
В ряде моделей совместное действие и
N
P
задается выражением
},min{
NP
xkN
N
xkP
P
++
β
.
Отметим, что согласно утверждению 5.1 при заданном соотношении
в модели выжывает лишь одна субпопуляция. В этом случае теряются адаптивные
свойства триады. Так, при изменении величины в среде уже будет
невозможна целесообразная перестройка модельной “Триады”. В этой связи,
правую часть “Триады” целесообразно дополнить следующим образом
P/N
P/N
,
μ
+= )N/Sk,P/Sk(fxx
i
N
i
Pii
&
где ;
3,2,1=i
μ
- положительно и мало. Присутствие малого числа
μ
позволяет
навечно закрепить трехвидовую структуру в машинной модели и в тоже время
сохраняется прежний характер доминации (см. утверждение 4.2).
5.2. ”Парадоксальные“ модельные эксперименты и механизм
возникновения дисбаланса азота и фосфора в Азовском море
124
Соединения азота и фосфора являются необходимым кормовым ресурсом
для водорослей, которые, в свою очередь, служат пищей для более высоких
трофических уровней экосистем. С точки зрения теории автоматического
регулирования процессы питания осуществляют прямые связи, а процессы распада
отмерших организмов образуют обратные связи в общем круговороте веществ в
экосистеме (Новосельцев, 1989). Когда в море отношение общих запасов азота
(N
общ
) к общим запасам фосфора (P
общ
) близко к значению азот/фосфор в клетках
водорослей (N
кл
/P
кл
), то в биологическом круговороте веществ активно “работают”
две обратные связи (как по азоту, так и по фосфору). В главе 1 показано, что при
избыточном накоплении в среде
N
ослабляется действие обратной связи по азоту.
Сокращение же числа активных обратных связей ”опасно”, поскольку может
привести к дестабилизации динамики водной экосистемы.
Согласно наблюдениям (Бронфман и др., 1979) в Азовском море
среднемноголетняя величина np=N
общ.
/P
общ
составляла
≈
10. Указанное значение
np близко к величине N
кл
/P
кл
для основных видов азовских водорослей. После
сооружения Цимлянского водохранилища (1953 г.) сократился объем (
v
) и
деформировался химический состав ( ) донского стока. В результате в
Азовском море стало наблюдаться постепенно нарастание отношения . За
промежуток времени 1970-1980 гг. величина np достигла значения 20.
PN
c,c
np
≈
Представляет интерес поиск причин, вызывающих рост .
np
Напомним, что в модели “водоросли – азот и фосфор” Азовского моря
представлены три основных вида - диатомовые, пирофитовые и синезеленые -
водорослей (см. табл. 5.1). Каждый вид водорослей описывается модельной
конструкцией "Триада".
Для дальнейшего важно отметить что потребление
P
всеми видами
водорослей примерно одинаково, а потребление
N
существенно различно. Так,
наибольшее потребление азота осуществляется синезелеными водорослями.
Управляющим фактором во внутригодовой динамике водорослей является
температура воды (
θ
). Зависимость скорости роста
β
от
θ
задается с помощью
так называемого температурного интервала толерантности
]da,da[I
T
+
−= .

125
Когда
T
I∈
θ
, то неотрицательная функция
)(
θ
β
имеет колоколообразный вид с
максимумом при
a=
θ
. В случае
T
I
∉
θ
полагаем
0)( =
θ
β
. Каждый вид
водорослей имеет свой интервал (см. табл. 5.1).
T
I
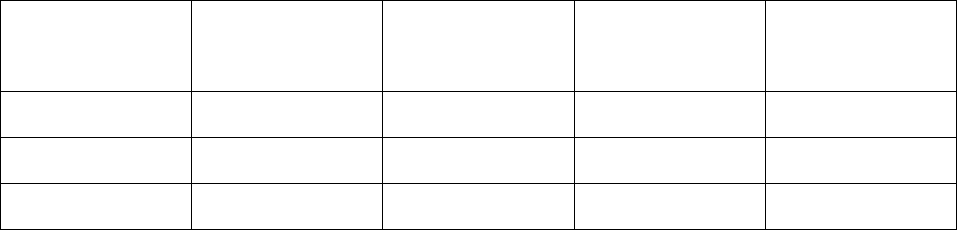
Таблица 5.1.
Экологические характеристики водорослей Азовского моря
(Алдакимова, 1976; Сергеев и др., 1979)
Содержание
*
в
клетках, в долях
Группа водорослей
Оптимальная
соленость,
00
0
/
Оптимальная
температура,
C
0
Азот Фосфор
Диатомовые 9 12-14 0,00525 0,00105
Пирофитовые 14 19-22 0,00750 0,00112
Синезеленые 6 22-24 0,01050 0,00120
*
Содержание в долях к сырому весу.
При наступлении неблагоприятных температурных условий (зима и др.)
активные клетки (
x
) водорослей переходят в пассивное состояние (споры). Когда
же температурные условия становятся благоприятными происходит возврат спор
(
s
) в активное состояние. Кратко напомним формализацию данного процесса из
главы 2. Пусть исходная модель имеет вид
),x(xfx
θ
=
&
.
Учет процесса спорообразования добавляет к данной модели следующую
линейную надстройку:
,s)(qx)(ps
,
s
)
(
q
x
)
(
p),
x
(
x
f
x
θθ
θ
θ
θ
−=
+
−
=
&
&
где функции и - скорости перехода из одного состояния в другое. Каждый вид
водорослей характеризуется своим температурным интервалом “живучести”
p q
I
Ж
=[b – c, b + c], в пределах которого его клетки сохраняют активность. Поэтому
будем полагать:
)(q
θ
- велико и
0)( =
θ
p
при
I
∈
θ
Ж
;
0)(
=
θ
q
и
)(p
θ
- велико при
I
∉
θ
Ж
.
Наблюдения показывают, что интервал
I
Ж
накрывает почти совпадает с
126
интервалом .
T
I
Перейдем к описанию параметров речного стока, использованных в
модельных экспериментах. До 1953 года среднемноголетний объем суммарного
речного стока (Дон + Кубань) в Азовское море составлял 41 км /год. Эта величина
равна 1/8 от объема Азовского моря. Поэтому речной сток играет главную роль в
судьбе экосистемы Азовского моря
3
Согласно данным (Бронфман и др., 1979) за период 1935-1975 гг.
содержание азота и фосфора в донской воде изменялось в диапазонах:
∈
N
c [1050, 2040] мг/м и
3
∈
P
c [155, 275] мг/м .
3
Соответствующие показатели для кубанских вод составляли
∈
N
c [2475, 2660] мг/м
3
и
∈
P
c [164, 710] мг/м
3
.
После сооружения Цимлянского и Краснодарского водохранилищ
произошли значительные потери (до 30%) объема речного стока в результате
испарения и водозабора на орошение. Попутно изменилось и содержание
N
и
P
в
речном стоке. Так, по данным 1973-1975 гг оно составило:
Дон ) мг/м и
2420≈
N
c
3
155
≈
P
c мг/м ;
3
Кубань ) мг/м
3
и 2475≈
N
c 164
≈
P
c мг/м .
3
Содержание азота и фосфора (биогенных веществ) в источниках водного
баланса Азовского моря приведено в таблице 5.2.
Таблица 5.2.
Биогенные вещества (мг/м ) в элементах водного баланса Азовского моря
3
Биогенный
элемент
Дон Кубань Атмосферные
осадки
Черное море
Общий
N
2000 – 3000 1900 1400 350
Общий
P
200 – 300 200 45 27
PN / 6,6 – 15 9,5 31 13
В результате деформации речного стока изменилось и отношение общих
запасов и
N
P
в Азовском море. Для сравнения приведем некоторые данные
1256/677np ≈=
(1958 г.) и
2843/1207np
≈
=
(1974 г.).
127
В том же направлении произошло изменение и отдельных форм
N
и
P
.
Приведем одно из “естественных” объяснений (химическая гипотеза ) роста
np, предложенное Бронфманом А. М. с соавторами (1979). А именно, в
Цимлянском водохранилище летом возникают исключительно благоприятные
условия для развития синезеленых водорослей (высокая температура + медленное
течение). Клетки этих водорослей способны к фиксации атмосферного N, поэтому
в результате их жизнедеятельности происходит дополнительное обогащение
цимлянских вод азотом. Содержание органического
P
в воде несколько снижается
в результате его частичного захоронения в донные отложения. В целом, указанные
процессы вызывают одновременное увеличение содержания азота ( ) и
уменьшение содержания ( ) в Цимлянском водохранилище, а затем и в водах
Нижнего Дона. Наконец, донские воды (”с большим np”) попадают в Азовское
море, и это порождает рост np в азовской экологической системе.
N
c
P
c
Отметим, что поступление в Азовское море донских вод с
“деформированным содержанием N и P” вызовет внутривидовую перестройку
азовских водорослей. Согласно принципу компенсации Ле-Шателье-Брауна
начнут доминировать виды водорослей с высокой потребностью в N . Такая
адаптивная реакция сообщества водорослей может “смягчить” рост N в
экосистеме Азовского моря.
Для исследования степени “смягчения” проведено компьютерное
исследования действие различных вариантов химического состава донского стока
на нижние трофические уровни экосистемы Азовского моря. Попутно
варьировался и объем донского стока. Так, задавались следующие диапазоны :
1)
v
изменялся в пределах 10-50 км
3
/год;
2) для < > рассмотрены четыре варианта
PN
cc ,
1 – <2000,200>; 2 – <2000,300>; 3 – <3000,200>; 4 – <3000,300> в мг/м .
3
Объем и содержание азота и фосфора в стоке реки Кубани строго
фиксированы и соответствуют современному состоянию.
Кроме того, были использованы среднемноголетние данные внутригодового
хода температурного и кислородного режимов.