Горбунова В.Н., Савельева-Васильева Е.А., Красильников В.В. Молекулярная неврология. Заболевания нервно-мышечной системы
Подождите немного. Документ загружается.

ной делецией 46% дистрофина. Эта огромная по протяжен-
ности делеция, целиком локализованная внутри стержнево-
го домена, практически не затрагивала функциональной ак-
тивности белка (England et al., 1990). После внутривенного
введения фрагментов гена Dmd в составе рекомбинантного
аденовируса наблюдали длительное присутствие экзогенной
ДНК в скелетных и сердечной мышцах животных (Srataford-
Perricaudet etal., 1992). Повышение экспрессии дистрофина,
происходящее у mdx-мышей в результате трансгеноза пол-
норазмерного гена Dmd, предотвращает развитие аномаль-
ных механических свойств мышц диафрагмы, связанных с
их дистрофией (Сох et al., 1993а). При этом 50-кратное уве-
личение уровня экспрессии дистрофина не вызывало ника-
ких побочных токсических эффектов.
Несмотря на эти очевидные успехи, проблема генно-
инженерной коррекции миодистрофии Дюшенна еще далека
от своего решения. До сих пор ведутся оживленные дебаты
исследователей о перспективности генной терапии миодист-
рофии Дюшенна по сравнению с клеточной терапией — пе-
ресадкой здоровых эмбриональных миобластов. Пока не ут-
верждена ни одна программа клинических испытаний гено-
терапевтического лечения миодистрофии Дюшенна. Основ-
ные проблемы генокоррекции данного наследственного за-
болевания связаны с (1) огромным количеством дистрофин-
положительных мышечных клеток, (2) сложным характером
регуляции экспрессии DMD гена, (3) необходимостью обес-
печения системы эффективной доставки гена дистрофина в
миофибриллы не только скелетных мышц, но, что особенно
важно, в мышцы сердца и диафрагмы, а возможно, и в раз-
личные отделы мозга, а также (4) с иммунологическими ос-
ложнениями, возникающими при использовании вирусных
векторов. В связи с этим наряду с совершенствованием пря-
мых, вирус-опосредованных и липосомных способов введе-
ния чужеродных ДНК разрабатываются иные подходы к ге-
нокоррекции миодистрофии Дюшенна, такие как: использо-
вание макрофагов, нагруженных плазмидами; модификация
6 Заказ № 170
81
культивированных сателлитных клеток, полученных с единич-
ных мышечных волокон; совместное выращивание фибро-
бластов и миобластов, их модификация и генетическая транс-
формация фибробластов в миобласты; конструирование
мини-хромосом, при котором ген дистрофина в составе YAC-
вектора вводится в сферобласты и проводится инфузия этой
конструкции в культивируемые миобласты.
Один из подходов в генотерапии миодистрофии Дюшен-
на основан на трансфекции в мышечные клетки антисмыс-
ловых РНК с целью посттрансляционной коррекции сдвига
рамки считывания (Matsuo, 1996). Эта технология основана
на том наблюдении, что делеции в гене DMD у пациентов с
мягкими формами миодистрофии Беккера, как правило, не
сопровождаются сдвигом рамки считывания. В опытах in vitro
показано, что в сарколемме культивируемых миотубул mdx-
мышей появляется экспрессия дистрофина после трансфек-
ции в них олигонуклеотидов, комплементарных З'-сайту сплай-
синга 22 интрона гена Dmd (Dunckley et al., 1998). Путем пря-
мого секвенирования соответствующих последовательностей
кДНК, полученных из этих клеток методом обратной ПЦР (RT-
PCR), показано, что при сплайсинге происходит точное вы-
резание области, включающей мутантный сайт и заключен-
ной между 22 и 30 экзонами. При этом образуется новый
РНК-транскрипт дистрофина, который может кодировать про-
дукт, достаточно функциональный для предотвращения раз-
вития наиболее тяжелых проявлений миодистрофии.
В 1995 г. исследования по генотерапии миодистрофии
Дюшенна начаты в рамках программы «Геном человека» и в
нашей стране. С использованием метода бомбардировки кле-
ток и тканей конъюгированными с плазмидами частицами зо-
лота или вольфрама, разогнанными до высоких скоростей,
полноразмерная кДНК дистрофинового гена была успешно
трансфецирована в скелетные мышцы мышей линии mdx
(Zelenin et al., 1997). Иммуногистохимический анализ пока-
зал, что в некоторых клетках мышц мутантных животных по-
является дистрофин (рис. 10). Доля дистрофин-положитель-
82
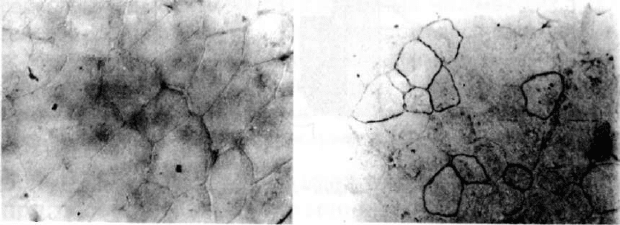
ных волокон в области бомбардировки поднимается при этом
до 2.5-5% (по сравнению с 0.5% в контроле) и сохраняется
на этом уровне, по крайней мере, в течение двух месяцев. В
некоторых экспериментах доля цельнокрашенных дистрофин-
положительных волокон в скелетных мышцах увеличивалась
от 27% через 2—3 недели до 82% через 2 месяца после бал-
листической трансфекции (Михайлов и др., 1998). По-види-
мому, в некоторых случаях происходит нерегулируемая экс-
прессия трансфецированного гена. Однако экспрессия дис-
трофина не нормализует полностью дифференцировку воло-
кон. Большой процент цельнокрашенных миофибрилл деге-
нерирует, в других сохраняется центральное положение ядер.
Рис. 10. Иммуногистохимический анализ дистрофина в
мышечных клетках мышей линии mdx до (а) и после (б)
введения кДНК-овых конструкций гена DMD
В отличие от липосомного способа введения, еще бо-
лее успешной оказалась трансфекция полноразмерной кДНК
дистрофина (pHSADy) с помощью синтетических микросфер
MF-2 (Баранов и др., 1998; Baranov et al., 1998). После одно-
кратной инъекции MF-2/pHSADy в четырехглавую мышцу бе-
дра введенная конструкция обнаруживается методом ПЦР
во многих тканях, включая скелетные мышцы, сердце, мозг,
легкие, причем присутствие нагруженных плазмидой микро-
сфер определяется методом флюоресцентной гибридизации
in situ (FISH) в 60—70% ядер. Неожиданно высоким оказы-
83
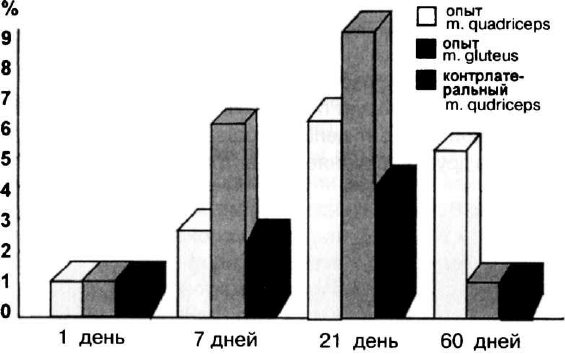
вается уровень дистрофин-положительных волокон в контра-
латеральной по отношению к инъецированной четырехглавой
мышце mdx-мышей спустя 60 дней после введения (рис. 11).
Рис. 11. Число дистрофин-положительных волокон
Многие проблемы в генотерапии миодистрофии Дюшен-
на могли бы быть преодолены при использовании принципи-
ально иного метода коррекции генного дефекта, основанно-
го на специфической активации у больных аутосомного го-
молога DMD — гена UTRN (Tinsley, Davies 1993; Blake et al.,
1996). Сходная ртратегия изменения регуляции экспрессии
фетального гемоглобина была использована для компенса-
ции дефекта глобиновых цепей у больных с серповидно-кле-
точной анемией (Perrine et al., 1993). Характер экспрессии
утрофина в раннем эмбриогенезе позволяет предполагать,
что этот белок может функционировать как фетальная изо-
форма дистрофина. Возможно при этом, что утрофин ока-
жется способен заменить дистрофин, если удастся изменить
характер экспрессии UTRN-гена у пациентов в тех клетках, в
которых в норме ген DMD экспрессируется. Для направлен-
84
ного изменения регуляции работы гена утрофина с помощью
определенных химических стимуляторов необходимо преж-
де всего детальное знание его промоторной области. Реали-
стичность такого подхода для лечения миодистрофии Дюшен-
на проверяется в настоящее время в опытах по трансгенозу
на модельных объектах, в первую очередь, на mdx-мышах.
Мы уже упоминали о том, что при трансгенном введении кон-
струкции, обеспечивающей экспрессию в скелетных мышцах
укороченной формы утрофина, в линию дистрофин/утрофин-
дефицитных мышей, наиболее полно фенокопирующую мио-
дистрофию Дюшенна, наблюдается коррекция основных
клинических симптомов миодистрофии (Rafael et al., 1998).
Очевидно, что присутствие укороченной формы утрофина в
сарколемме мышц мутантных животных, не имеющих эндо-
генного дистрофина и утрофина, способно восстановить дис-
трофин-ассоциированный комплекс белков, нормальное раз-
витие и функционирование дефектных мышц. Значение этих
результатов для разработки программ генотерапии миодист-
рофии Дюшенна трудно переоценить.
б) Мышечная дистрофия врожденная
прогрессирующая с умственной отсталостью, тип
Фукуяма (MIM: 253800)
В 1960 г. впервые Фукуяма описал 6 семей с врожден-
ной прогрессирующей миодистрофией дюшенновского типа,
сочетающейся с выраженной умственной отсталостью
(Fukujama et al., 1960). Характер наследования этой формы
миодистрофии соответствовал аутосомно-рецессивному.
Представлены результаты клинико-генеалогического анали-
за более 200 семей с этим заболеванием в Японии (Fukujama
et al., 1981). Прогрессирующей мышечной слабости часто
сопутствуют мышечная гипотония, эпилептиформные припад-
ки, гидроцефалия. Патоморфологически обнаруживаются
микрополигирия, фиброглиальная пролиферация мягкой и
паутинной оболочек. Кроме миодистрофии типа Фукуяма
известны три клинически различающихся синдрома, при ко-
85
торых врожденная миопатия сочетается с пороками разви-
тия, такими как лиссэнцефалия, гидроцефалия, аномалии
глаз. Это синдром Валькера-Варбурга (гидроцефалия, агирия,
дисплазия сетчатки: HARD-синдром, MIM 236670), описанная
в Финляндии болезнь МЕВ (muscle-eye-brain disease, MIM
253280) и врожденная мышечная дистрофия с центральной
гипомиелинизацией, обусловленная дефектом мерозина (см.
ниже, MIM 156225).
При иммуногистохимическом анализе биоптатов мышц
больных с миодистрофией Фукуяма примерно в 13% случа-
ев обнаруживаются аномалии дистрофина (Beggs et al., 1992).
Для объяснения этого было высказано предположение, что
дистрофин и продукт гена FCMD (Fukujama type congenital
muscular dystrophy) взаимодействуют. Раннее начало и бо-
лее тяжелое течение заболевания наблюдается у пациентов,
которые одновременно гетерозиготны по FCMD и гемизигот-
ны по мутациям DMD-гена. Предполагается, что такой ком-
бинированный фенотип встречается среди мужчин в Японии
с частотой 1:175000.
У пациентов с миодистрофией Фукуяма обнаруживает-
ся аномально низкий уровень экспрессии многих белков дис-
трофин-ассоциированного комплекса, включая дистроглика-
ны — трансмембранный 43DAG и экстраклеточный 156DAG,
и саркогликаны (Matsumura et al., 1993). Кроме того, содер-
жание ламинина в скелетных мышцах и в периферических
нервах пациентов также оказывается сниженным. Однако
генетический анализ семей дает основания предполагать, что
эти нарушения являются вторичными по отношению к основ-
ному биохимическому дефекту, так как найдено сцепление
гена FCMD с микросателлитными маркерами, локализован-
ными в области 9q31 -q33 (Toda et al., 1993). Ближайшие флан-
кирующие маркеры D9S58, D9S59 и D9S2107. Не исключе-
но, что в этой области локализован ген, кодирующий еще один
белок дистрофин-ассоциированного комплекса (DAP), дис-
функция которого обусловливает клинику миодистрофии Фу-
куяма. Перекрывание клинических симптомов при миодист-
86

рофии Фукуяма и при мягких формах синдрома Валькера-
Варбурга и результаты анализа гаплотипов больных по мар-
керам, сцепленным с геном FCMD, не противоречат предпо-
ложению об аллельной природе этих двух заболеваний (Toda
et al., 1995).
Дальнейший генетический анализ, выполненный с ис-
пользованием техники микросателлитного картирования,
позволил сузить область локализации гена FCMD до менее
чем 100 кб. Кандидатным для FCMD рассматривается в на-
стоящее время расположенный в той же области ген мышеч-
ной рецепторной тирозинкиназы — MUSC (muscle specific
kinase) (Toda et al., 1996). Впервые гомолог этого гена —
MuSC — был идентифицирован у мышей (Valenzuela et al.,
1995). Показано, что MuSC специфическим образом экспрес-
сируется в скелетных мышцах и в нейромышечных соедине-
ниях. При разрушении этого гена путем трансгеноза у мы-
шей не формируются нейромышечные синапсы (DeChiara et
al., 1996). Предполагается, что кодируемая геном MUSC ре-
цепторная тирозинкиназа является критическим сигналом для
агрина, участвующего в активации всех ступеней каскада,
ведущего к образованию синапса, включая организацию пост-
синаптической мембраны, синапс-специфическую транскрип-
цию и пресинаптическую дифференцировку.
1.3. Конечностно-поясные
мышечные дистрофии
Это гетерогенная группа заболеваний с преиму-
щественной локализацией дистрофического процесса в
мышцах плечевого и тазового пояса. Выделяют следующие
клинические типы: тазобедренный Лейдена-Мёбиуса;
лопаточно-бедренный Эрба; поздний аутосомно-
доминантный и квадрицепс-миопатию (Вельтищев и др.
1998). Описана также необычная форма аутосомно-
рецессивной мышечной дистрофии плечевого и тазового
пояса в сочетании с буллезным эпидермолизом.
87
а) Миодистрофия конечностно-поясная аутосомно-
доминантная, типы: 1A(MIM:159100); 1В; 1С;
аутосомно-рецессивная, типы: 2А (MIM-.253600);
2В (MIM:253601); 2С (MIM:253700); 2D (ОМ1М:600119);
2Е (OMIM:600900); 2F (OMIM: 601400)
Заболевание начинается в юношеском возрасте с
11—20 лет, возможно и более раннее начало. Мышечная
слабость, утомляемость равиваются постепенно, поэтому
точно определить дебют заболевания не всегда удается.
Слабость сочетается с симметричными атрофиями мышц
тазового пояса. Больные начинают испытывать затрудне-
ния при беге, быстрой ходьбе, прыжках. Ограничение дви-
гательных возможностей начинает выделять их из группы
сверстников. Это особенно отчетливо выявляется на за-
нятиях физкультурой в школе. В дальнейшем появляются
затруднения при подъеме на лестницу, вставании со стула
и особенно с пола. Появляются так называемые вспомо-
гательные движения, меняющие формулу нормального дви-
гательного акта. Начинаясь с тазового пояса и ног, мышеч-
ная слабость распространяется на плечевой пояс и туло-
вище. Меняется осанка, опускаются надплечья, появляют-
ся «крыловидные» лопатки. Туловище отклоняется назад,
усиливается поясничный лордоз, выпячивается живот, по-
является «осиная» талия. Больной ходит раскачиваясь из
стороны в сторону, с трудом отрывая стопы от пола. Рань-
ше других атрофируются поясничные и ягодичные мышцы,
четырехглавая мышца бедра. Псевдогипертрофии, если и
бывают, то не столь выраженными, как при форме Дюшенна.
Лицевая мускулатура обычно не страдает. Сухожильные и
периостальные рефлексы снижаются, а затем полностью ис-
чезают. Снижение рефлексов начинается с коленных, а за-
тем с двуглавых мышц. Дистальные отделы конечностей еще
долго сохраняют свои функции. Течение заболевания про-
грессирующее, менее злокачественное по сравнению с
миодистрофией Дюшенна. Способность к самостоятельно-
му передвижению может сохраняться до 20—30 лет.
88

1. Общая генетическая характеристика
LGMD — (limb-girdle muscular dystrophy) — гетероген-
ная группа заболеваний с варьирующей экспрессивностью
мутантного гена. В настоящее время идентифицированы три
аутосомно-доминантных формы заболевания: LGMD1A —
LGMD1C и шесть аутосомно-рецессивных форм: LGMD2A —
LGMD2F (Bushby et al., 1995; Van der Kooi et al.,1997; Bashir
et al., 1998). Форма 1A связана с геном, расположенным в
длинном плече хромосомы 5 в области 5q22.3-31.3 (Speer et
al., 1992). Активно обсуждается возможность участия в кон-
троле этого заболевания гена LMNB1, расположенного в той
же цитогенетической области и кодирующего один из белков
семейства ядерных ламинов — В1. Ген, ответственный за мио-
дистрофию LGMD1B, ассоциированную с кардиологическими
дефектами, расположен в длинном плече хромосомы 1 — в
области 1q11-q22 (van der Kooi et al., 1997). В этой же области
локализован ген LMNA, кодирующий две другие альтернативно
сплайсирующиеся изоформы ядерных ламинов А и С. Показа-
но, что ген LMNA дефектен при аутосомно-доминантной фор-
ме мышечной дистрофии Эмери-Дрейфуса (Bonne et. AL,
1999).Несмотря на выраженные клинические различия между
этими двумя заболеваниями (отсутствие контрактур и преиму-
щественная слабость проксимальных отделов конечностей при
обсуждаемой форме конечностно-поясной миодистрофии), не
исключена их аллельная природа.Третья аутосомно-доминант-
ная форма миодистрофии плечевого и тазового пояса LGMD1С
обусловлена мутациями в гене кавеолина-3 (CAV3), располо-
женном в области Зр25 (Minetti et al., 1998).
Пациенты с аутосомно-рецессивными формами
LGMD2A и LGMD2B имеют мягкий фенотип и часто клиниче-
ски могут быть отнесены к типу Эрба. Форма 2А связана с
мутациями в гене CAPN3, расположенном в длинном плече
хромосомы 15. CAPN3 кодирует одну из субъединиц специ-
фической мышечной протеазы — кальпаина-3. Форма 2В ока-
залась аллельным вариантом миопатии Миоши и обусловле-
на мутациями в гене DYSF, локализованном в коротком плече
89

хромосомы 2. Продукт этого гена — дисферлин, являющийся
цитоплазматическим белком, способным связываться с эндо-
или саркоплазматическим ретикулумом, а также с ядерной мем-
браной. Гены, ответственные за более тяжелые формы ауто-
сомно-рецессивной конечностно-поясной миодистрофии
LGMD2C — LGMD2F, кодируют белки саркогликанового дистро-
фин-ассоциированного субкомплекса: у-, ос-, р- и 6-саркоглика-
ны, соответственно (Roberds et al., 1994; Lim et al., 1995;
Bonnemann et al., 1995; Noguchi et al., 1995; Nigro et al., 1996b).
При исследовании бразильских семей с аутосомно-ре-
цессивными формами тазово-поясной мышечной дистрофии,
в каждой из которых наблюдали не менее 3 пациентов, в 33%
семей болезнь была обусловлена мутациями в гене кальпаи-
на 3 (форма 2А), в 33% — в гене дисферлина (форма 2В), в
17% — в гене ос-саркогликана (форма 2D), в 10% семей — в
гене усаркогликана (форма 2С) (Passos-Bueno et al., 1996а).
Следует подчеркнуть, что дифференциальная диагностика раз-
личных генетических форм конечностно-поясных и аутосомно-
рецессивных дюшенно-подобных форм миодистрофии возможна
в настоящее время только на базе молекулярного анализа.
Генетическое разнообразие конечностно-поясных миоди-
строфии, по-видимому, не ограничивается описанными выше
формами. Так, на основании анализа большой финской семьи,
в которой одновременно наблюдали больных с двумя различ-
ными формами заболевания — тяжелой конечностно-поясной
и мягкой дистальной миопатией с поздним началом, высказано
предположение, что в первом случае это результат проявления
доминантного гена в гомозиготном, а во втором случае — в
гетерозиготном состоянии (Udd et al., 1992). Остановимся бо-
лее подробно на тех формах миодистрофии плечевого и тазо-
вого пояса, для которых гены уже идентифицированы.
2. Конечностно-поясная миодистрофия. аутосомно-
доминантная. форма LGMD1C
Мы уже упоминали о том, что форма LGMD1С обуслов-
лена мутациями в гене кавеолина-3 (CAV3), расположенном
в коротком плече хромосомы 3 (Minetti et al., 1998). Кавео-
90