Горбунова В.Н., Савельева-Васильева Е.А., Красильников В.В. Молекулярная неврология. Заболевания нервно-мышечной системы
Подождите немного. Документ загружается.

тический анализ семей, в которых наблюдалась сегрега-
ция по сцепленной с полом форме дилатационной кардио-
миопатий, показал, что ген, ответственный за развитие
заболевания, локализован в той же области Хр21, где на-
ходится ген DMD (Towbin et al., 1993). Спустя некоторое
время у ряда больных с семейной Х-сцепленной формой
дилатационной кардиомиопатий, так же как и в нескольких
спорадических случаях у мужчин, были идентифицирова-
ны мутации в гене DMD. Это были либо делеции,затраги-
вающие регуляторную область и первый экзон DMD, либо
сплайсинговые мутации, локализованные в начале перво-
го интрона гена дистрофина (Yoshida et al., 1993; Muntoni
et al., 1995b; Milasin et al., 1996). У трех пациентов с Х-сцеп-
ленной дилатационной кардиомиопатией из двух неродст-
венных японских семей была обнаружена уникальная ин-
серция транспозоно-подобного элемента L1 в первом эк-
зоне гена DMD (Yoshida et al., 1998). В дальнейшем были
определены начальные этапы молекулярного патогенеза
подобных форм дилатационной кардиомиопатий. Оказа-
лось, что идентифицированные мутации приводят к полно-
му отсутствию мышечной изоформы дистрофина у боль-
ных. Кроме того, в миокарде пациентов отсутствует экс-
прессия двух других полноразмерных изоформ мРНК дис-
трофина, транскрибирующихся с промоторов мозгового
типа. В отличие от этого, в скелетных мышцах наблюдает-
ся достаточно высокий уровень двух полноразмерных изо-
форм дистрофина мозгового типа и потому сохраняется
дистрофин-гликопротеиновый комплекс. В сердечной мыш-
це не происходит компенсаторной экспрессии дистрофина
с альтернативных промоторов и дистрофин-гликопротеино-
вый комплекс полностью разрушен, несмотря на присутст-
вие там апо-дистрофина-1. В результате у пациентов раз-
вивается дилатационная кардиомиопатия без каких-либо
проявлений слабости скелетной мускулатуры.
Вскоре вслед за описанием молекулярного дефекта
п
Ри Х-сцепленной дилатационной кардиомиопатий у боль-
61
ных с аутосомно-рецессивными формами заболевания
были обнаружены мутации в двух группах генов. Одни гены
оказались связаны с белками дистрофин-ассоциированного
комплекса, такими как а-дистрогликан, а- и у-саркоглика-
ны, другие гены — с белками цитоскелета мышечной клет-
ки: метавинкулином, актином, мышечным белком, участ-
вующим в позитивной регуляции миогенеза, так называе-
мым LIM-белком (Towbin, 1998). Примечательно, что иден-
тифицированные у пациентов мутации в актиновом гене
приводили к дефекту того домена актина, который участ-
вует во взаимодействии с дистрофином. Дефект 5-сарког-
ликана найден в линии хомячка BI014.6 с кардиомиопати-
ей. Все эти факты свидетельствуют о центральной роли дис-
трофин-ассоциированного комплекса в функционировании
кардиомиоцитов. Не исключено, что и приобретенные фор-
мы дилатационной кардиомиопатии, доля которых дости-
гает 30% от общего числа дилатационных кардиомиопатии,
связана с нарушением нормальной работы дистрофина.
Энтеровирусы, и в частности вирусы Коксаки, способны
индуцировать дилатационную кардиомиопатию. Оказалось,
что единственным белком человека, который расщепляет-
ся вирусной протеазой 2А, является дистрофин. Это было
предсказано на базе компьютерного анализа и подтверж-
дено в опытах in vitro (Badorf et al., 1999). Дистрофин так-
же расщепляется в культивируемых миоцитах, зараженных
вирусом Коксаки, и in vivo в сердечной мышце инфициро-
ванных мышей. При этом в инфицированных миоцитах от-
сутствует не только дистрофин, но и ассоциированные с
ним гликопротеины — а-саркогликан и р-дистрогликан.
Очевидно, что разрушение дистрофинового комплекса яв-
ляется одним из центральных звеньев в этиологии дилата-
ционных кардиомиопатии.
Экспрессия дистрофина и Dp260 обнаружена во внеш-
нем плексиформном слое сетчатки глаза, где фоторецеп-
торные клетки взаимодействуют с нейронами ганглиев, в
то время как другой апо-дистрофин — Dp71 — экспресси-
62
пуется на внутренней ограничивающей мембране сетчат-
ки и на ее сосудах (Fillers et al., 1993; Howard et al., 1998).
у некоторых пациентов с миодистрофией Дюшенна /Бек-
кера и у мышей генетических линий mdx с мутациями в
гене Dmd выявлен аномальный характер электроретино-
грамм, причем степень аномалий зависит от типа мутаци-
онных повреждений.У трансгенных мышей с направленно
разрушенным экзоном 52 гена Dmd и отсутствием экспрес-
сии Dp260 изменена локализация р-дистрогликана и обна-
руживаются аномалии на электроретинограмме (Катеуа
et al., 1998). Показано, что присутствие каждого из двух
аподистрофинов — Dp71 и Dp260 — необходимо для пра-
вильной локализации р-дистрогликана, а это взаимодейст-
вие, в свою очередь, имеет важное значение для исполне-
ния нормальной электрофизиологической функции сетчат-
ки. Не исключено, что некоторые мутации в гене DMD, со-
провождающиеся образованием дефектных форм дистро-
фина и аподистрофинов Dp71 и Dp260, могут быть ответ-
ственны за развитие Х-сцепленной формы ночной слепо-
ты, ген которой также картирован в области Хр21.
Кроме того, полноразмерный дистрофии и его укоро-
ченная форма Dpi 16 активно экспрессируются во внутрен-
них и внешних волосковых клетках улитки, где эти белки
преимущественно располагаются на латеральной поверх-
ности и в синаптических районах (Dodson et al., 1995). Ген,
ответственный за развитие одной из моногенных форм ней-
росенсорной тугоухости, также картирован в области Хр21
недалеко от DMD (Lalvani et al., 1994). Возможно, дистро-
фины участвуют в трансдукции акустического сигнала в
нейрональный и некоторые их дефекты или дефекты дис-
т
Рофин-гликопротеинового комплекса ответственны за
°пределенные типы нейросенсорной тугоухости. Не исклю-
чено также, что делеций в 44-м интроне, «горячей» точке
Вн
Утригенной рекомбинации, специфически разрушающие
п
Ромоторную область для наиболее интенсивно экспрес-
Си
Рующегося в раннем эмбриогенезе нейронального апо-
63
дистрофина Dpi40, могут приводить к одной из форм
Х-сцепленной изолированной умственной отсталости, ген
которой также локализован в области Хр21.
Предполагается, что среди семей с миодистрофией
дюшенновского типа в 6.8% случаев наследование осуществ-
ляется по аутосомно-рецессивному типу (Zatz et al., 1989).
По оценкам этих же авторов, доля мальчиков с аутосомно-
рецессивным типом наследования среди изолированных слу-
чаев миодистрофии Дюшенна составляет 2.5-4%. Аномалии
дистрофин-связанных белков в сарколемме — первый об-
щий шаг в патологическом процессе, ведущем к некрозу мы-
шечных клеток, не только при Х-сцепленной форме миодист-
рофии Дюшенна, но и при тяжелых аутосомно-рецессивных
дюшенно-подобных миодистрофиях, так же как и при миоди-
строфии Фукуяма.
Дюшенно-подобные формы миодистрофии представ-
ляют собой генетически гетерогенную группу аутосомно-
рецессивных заболеваний. Чаще всего такие семьи выяв-
ляют потому, что в них наблюдаются девочки с клиникой
миодистрофии Дюшенна. При этом у больных девочек не
находят каких-либо структурных аномалий, затрагивающих
Х-хромосому. В частности, необычно высокая частота тя-
желой прогрессирующей мышечной дистрофии с аутосом-
но-рецессивным характером наследования описана в Ту-
нисе (Ben Hamida et al., 1983). Из 93 диагностированных
пациентов 75 принадлежали 17 семьям, в которых присут-
ствовали больные обоих полов, и 18 принадлежали 11 се-
мьям, в которых больными были только девочки. Оказа-
лось, что в большинстве случаев аутосомно-рецессивные
дюшенно-подобные формы миодистрофии обусловлены
мутациями в генах, кодирующих белки дистрофин-ассоци-
ированного комплекса. В табл. 7 суммированы данные по
локализации таких генов и их связи с наследственными
заболеваниями.
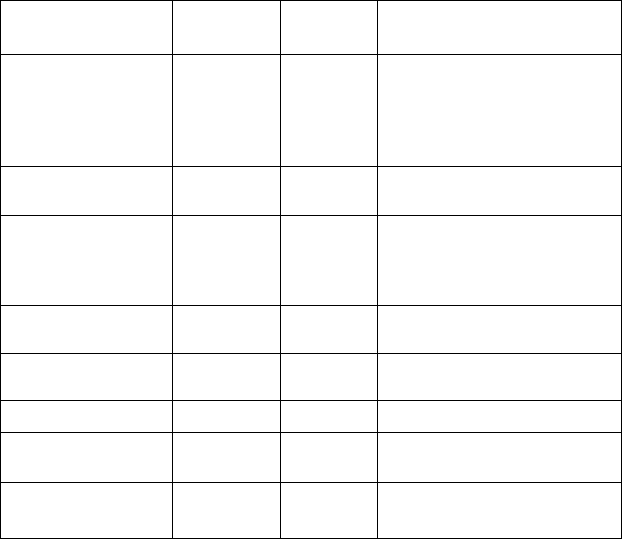
Таблица 7
Белок
Ген
Лока-
лизация
Заболевание
а-саркогликан,
адхалин
SGA, aSG,
ADL,
DAG2
17q21
первичная адхалинопатия;
SCARMD*); конечностно-
поясная миодистрофия 2D;
дилатационная
кардиомиопатия
р- саркогликан
SGB, pSG
4q12
конечностно-поясная
миодистрофия 2Е; ARMD**
у-саркогликан
SGG, ySG
13q12
конечностно-поясная
миодистрофия 2С;
SCARMD; дилатационная
кардиомиопатия
S-саркогликан
SGD, 8SG
5q33
конечностно-поясная
миодистрофия 2F
а- дистрогликан
DAG1
3p21
дилатационная
кардиомиопатия
р- дистрогликан
DAG1
3p21
не найдено
Дистробревин
DRP3
18q12.1-1
2.2
не найдено
Ламинин 2,
мерозин
LAMA
6q22-q23
миодистрофия врожденная
мерозин дефицитная
*) SCARMD — тяжелая детская аутосомно-рецессивная
мышечная дистрофия.
**) ARMD аутосомно-рецессивная мышечная дистрофия
Дюшенновского типа.
В ряде семей с тяжелой формой детской аутосомно-
Рецессивной мышечной дистрофией (SCARMD), впервые
описанной в большой тунисской семье, при сохранении нор-
мальных уровней дистрофина наблюдали выраженный дефи-
цит a-саркогликана, называемого иначе адхалином или
50DAG (Matsumura etal., 1992; Piccolo et al., 1995). Во многих
5
^каз №
170
65
Связь генов, кодирующих белки
дистрофин-ассоциированного комплекса,
с наследственными заболеваниями
таких семьях этот дефект является вторичным. В относитель-
но небольшом проценте семей подобный дефект является
первичным, то есть обусловлен мутациями в гене а-сарко-
гликана -aSG или ADL, картированном в области 17р21
(McNally et al., 1994).
Такая форма очень варьирующей по степени тяжести
аутосомно-рецессивной мышечной дистрофии получила на-
звание первичной адхалинопатии. Для пациентов с первич-
ной адхалинопатией характерным является преимуществен-
ное поражение проксимальных мышц, высокие уровни сыво-
роточной креатинкиназы, нормальное содержание дистрофи-
на, сохранный интеллект. Дебют и степень выраженности
клинических симптомов могут варьировать от ранних и тя-
желых форм до поздних с минимальными проявлениями
мышечной слабости. Примерно в 25% случаев у больных с
первичной адхалинопатией обнаруживается однотипная мис-
сенс-мутация R77C в гене ADL (Roberds et al. 1994). Пациен-
ты с этой мутацией были найдены в семьях различного этни-
ческого происхождения из Франции, Германии, Африки, США.
В дальнейшем эта форма миопатии была классифицирова-
на, как один из генетических вариантов конечностно-пояс-
ных миодистрофии, обусловленных наследственными дефек-
тами белков саркогликанового комплекса.
Аномально низкий уровень экспрессии саркогликанов
и дистрогликанов обнаружен у пациентов в Японии с врож-
денной аутосомно-рецессивной прогрессирующей мышеч-
ной дистрофией типа Фукуяма, сочетающейся с умствен-
ной отсталостью (см. ниже). Однако и в данном случае эти
дефекты являются вторичными, так как прослежено сцеп-
ление заболевания с маркерами длинного плеча хромосо-
мы 9 (9q31-q33).
Картированы и клонированы гены, кодирующие другие
белки саркогликанового субкомплекса. Мутации в этих генах
сопровождаются разрушением этого субкомплекса и реали-
зуются либо в виде различных форм миодистрофии плечево-
го и тазового пояса (Lim et al., 1995), либо в виде аутосомно-
66

рецессивной мышечной дистрофии (ARMD) дюшенновского
типа (Bonnemann et al., 1995).
DAG1 рассматривался в качестве кандидатного гена при
различных аутосомно-рецессивных формах миодистрофии.
Однако до сих пор в этом гене не обнаружено никаких мута-
ций, приводящих к наследственным миопатиям. По-видимо-
му, важная роль дистрогликанов в формировании оболочек
плода приводит к тому, что большая часть мутантов по DAG1
гену погибают в раннем эмбриогенезе. Мы уже упоминали о
том, что дефекты в а-дистрогликане обнаруживаются у неко-
торых пациентов с аутосомно-рецессивными формами дила-
тационной кардиомиопатий.
При иммуногистохимическом анализе биоптатов мышц
более 500 пациентов с различными формами миопатии, у
которых был сохранен нормальный уровень дистрофина, в
10% случаев наблюдали отсутствие окрашивания на ос-сар-
когликан (Duggan et al., 1997). В 60% этих случаев были об-
наружены мутации в саркогликановых генах. Большая часть
из них (58%) оказалась локализована в гене адхалина, 27% и
13% в генах р- и у-саркогликанов, соответственно. Чаще все-
го такие мутации обнаруживаются у пациентов с тяжелыми
дюшенно-подобными формами миодистрофии, дебютирующи-
ми в детском возрасте. Среди проксимальных форм с поздним
началом подобные мутации были найдены в 6% случаев.
8. Семейство DMD-родственных генов
Обнаружен аутосомный гомолог гена DMD, локализо-
ванный в области 6q24, — ген UTRN или DRP1, кодирующий
дистрофин-родственный белок с молекулярной массой в 395
кД — утрофин (табл. 8). Показано, что утрофин в ряде случа-
ев может выступать в качестве функционального гомолога
Дистрофина. Геномная область, занимаемая UTRN, почти
втрое меньше гена DMD и составляет 900 кб. Размеры РНК-
"Фанскриптов этих двух генов близки — величина мРНК DMD
составляет 14 кб, a UTRN — 13 кб (Blake et al., 1996). Ген
UTRN активно экспрессируется в нейромышечных и в мы-
67
шечно-сухожильных соединениях, в мозге и в дополнение к
этому во многих других висцеральных тканях, в первую оче-
редь — в легких. Интересно подчеркнуть, что регуляция экс-
прессии генов DMD и UTRN осуществляется, по-видимому,
аналогичным образом. Так, обнаружен 5.5-кб транскрипт это-
го гена, преимущественно экспрессирующийся в сенсорных
ганглиях и в мозге, кодирующий 113 кД белок G-утрофин, го-
мологичный апо-дистрофину периферических нервов Dpi 16
(Blake et al., 1995). Описан также 80 кД утрофин, который,
по-видимому, является аутосомным гомологом Dp71
(Fabbrizio et al., 1995a).
Кроме того, в области Xq22 идентифицирован другой
небольшой ген, гомологичный З'-концу DMD — DRP2 (Roberts
et al., 1996). Этот ген занимает 45 кб геномной ДНК, его ко-
дирующая область разделена на 24 экзона. Продуктом гена
DRP2 является 110-кД белок, состоящий из 956 аминокислот.
Ген DRP2 преимущественно экспрессируется в головном и в
спинном мозге, и его продуктом является белок, родствен-
ный Dpi 16. Прослежена эволюционная связь между дистро-
фин-родственными генами млекопитающих и их предшест-
венниками у беспозвоночных (Roberts,Bobrow, 1998). По-ви-
димому, у большинства беспозвоночных имеется один ген,
кодирующий высококонсервативный дистрофин-родственный
белок, в дополнение к гену дистробревина (DRP3), имеюще-
му более отдаленное сходство с дистрофином. Дистрофин-
родственный ген и ген дистробревина произошли от гена-пред-
шественника путем более ранней дупликации. У позвоноч-
ных дистрофин-родственный ген подвергался серии дупли-
каций, приведших к образованию покрайней мере трех от-
дельных генов (DMD, UTRN и DRP2), способных выполнять
различные специализированные функции.
Недавно был изолирован ген морского ежа, который,
по-видимому, является предшественником генов дистрофи-
на и утрофина (Wang et al., 1998). Более чем 300-кД продукт
этого гена содержит покрайней мере 3 из четырех доменов
дистрофина. В этом гене также обнаружено несколько про-
68

моторов и его наименьший продукт, по-видимому, родстве-
нен Dpi 16. Предложена модель, согласно которой ген DRP2,
кодирующий Ор116-родственный белок, также эволюциони-
ровал от этого гена-предшественника путем дупликации его
небольшой части. В настоящее время никаких мутаций в ге-
нах UTRN, DRP2 и DRP3 не описано.
Таблица 8
Семейство DMD-родственных генов
Ген
Локализация
Размер
гена (кб)
Размер
мРНК
(Кб)
Основной
продукт
DMD
Хр21 2500
14
дистрофии
UTRN,
DRP1
6q24
900 13
утрофин
DRP2
Xq22
45 2.8
дистрофин-род-
ственный белок 2
DRP3
18q12.1-12.2
дистробревин
395-кД утрофин также относится к спектрин/а-актини-
новому семейству белков. Он имеет доменную организацию,
гомологичную дистрофину. При этом сходство его актин-свя-
зывающего, цистеин-богатого и С-концевого доменов с соот-
ветствующими доменами дистрофина достигает 85—88%. На-
ибольшие различия касаются стержневого домена, состоя-
щего в утрофине из 22 повторяющихся спектрин-подобных
сегментов, прерванных двумя петлеобразными районами.
G-утрофин содержит уникальный N-конец, небольшой фраг-
мент стержневого домена, включающий два спектрин-подоб-
ных повтора и последний петлеобразный район, а также цис-
теин-богатый и С-концевой домены утрофина. Показано, что
утрофин подобно дистрофину способен взаимодействовать
с компонентами дистрофин-гликопротеинового комплекса и
69
выполнять сходные функции, то есть дистрофин и утрофин,
по-видимому, являются функциональными гомологами. Дру-
гой дистрофин-родственный белок, кодируемый геном DRP2,
содержит короткий пролин-богатый район, два спектрин-по-
добных повтора стержневого домена и последовательность,
гомологичную последующим доменам дистрофина и утрофи-
на — цистеин-богатому и С-концевому. Найдено большое
сходство между Dpi 16 изоформой дистрофина и дистрофин-
родственным белком drp2 (Roberts et al., 1996). Хотя точные
функции G-утрофина и drp2 неизвестны, они могут подобно
апо-дистрофинам связываться с р-дистрогликаном.
В раннем эмбриогенезе утрофин присутствует в сарко-
лемме скелетных мышц, занимая сходное с дистрофином
положение (Clerk et al., 1993). По-видимому, высокая скорость
деления клеток в этот период не позволяет транскрибиро-
ваться такому крупному гену, как DMD. Однако в дальней-
шем дистрофин постепенно замещает утрофин в сарколем-
ме мышц и начиная с 26 недель беременности утрофин там
уже не обнаруживается, а экспрессируется только в нейро-
мышечных и мышечно-сухожильных соединениях, причем его
локализация в этих сайтах отлична от локализации дистро-
фина. В сердечной мышце утрофин находят в интерколяр-
ных дисках, тогда как дистрофин располагается преимущест-
венно по периферии кардиоцитов.
Можно считать доказанным участие утрофина в синап-
тогенезе. Показано, что взаимодействие сс-дистрогликана с
агрином и перлеканом, стимулирующее в дальнейшем клас-
терирование ацетилхолиновых рецепторов, осуществляется
в тех случаях, когда этот гликопротеин находится в комплек-
се с утрофином. На рис. 9 изображена структура утрофин-
ассоциированного комплекса белков в нейромышечных со-
единениях. Оказалось, что утрофин, в отличие от дистрофи-
на, способен взаимодействовать с рапсином, содержащим
43кД цинк-фингерный домен и принимающим непосредствен-
ное участие в перекрестной сшивке ацетилхолиновых рецеп-
торов с образованием микрокластеров (Phillips et al., 1993).
70