Горбунова В.Н., Савельева-Васильева Е.А., Красильников В.В. Молекулярная неврология. Заболевания нервно-мышечной системы
Подождите немного. Документ загружается.

н
ого и мозгового типа, различающиеся по структуре первого
экзона, одновременно присутствуют в нейронах коры и гип-
покампа, и дополнительный полноразмерный транскрипт иден-
тифицирован в клетках Пуркинье. В последнем случае транс-
крипт также отличается от двух известных продуктов по струк-
туре первого экзона, который оказался локализован в большом
интроне между мышечным промотором и вторым экзоном гена.
Все образующиеся при этом формы дистрофина имеют неболь-
шие отличия в N-концевом участке белка.
В немышечных тканях и в клеточных линиях найдено
множество укороченных форм дистрофина — апо-дистрофи-
нов, образующихся за счет альтернативной транскрипции —
Dp40, Dp71, Dpi 16, Dpi40, Dp260. Цифры в названии этих
белков соответствуют молекулярным весам соответствующих
апо-дистрофинов. Dp71, или апо-дистрофин-1, по-видимому,
является основным продуктом гена DMD в немышечных тка-
нях, включая ткани мозга (Lederfein et al., 1993а; 1993b; Bar
etal., 1990). В Dp71 присутствует С-концевой домен дистро-
фина и часть цистеин-богатого, а также 7 дополнительных
аминокислот на N-конце белка. Содержание этого белка в
некоторых тканях сопоставимо с количеством дистрофина в
мышцах, и особенно обильно он представлен в клетках Шван-
на, где дистрофии отсутствует (Blake et al., 1992). В скелет-
ных мышцах взрослых Dp71 отсутствует, тогда как в мышцах
плода наряду с дистрофином экспрессируется и апо-дистро-
фин-1. Уровень мРНК Dp71 в культуре дифференцирующих-
ся миобластов сохраняется стабильным, тогда как содержа-
ние белка в процессе дифференцировки значительно возра-
стает, что указывает либо на его большую стабильность по
сравнению с полноразмерным дистрофином, либо на возмож-
ность изменений в эффективности трансляции (Tennyson et
a
'-i 1996). мРНК-транскрипт Dp71 более устойчив по сравне-
нию с полноразмерным транскриптом, так как период его
полураспада составляет около 20 часов, тогда как для мРНК
Дистрофина — только 16 часов (Tennyson et al., 1995). Иден-
тифицированы различные изоформы апо-дистрофина-1, об-
51
разующиеся за счет альтернативного сплайсинга последних
экзонов гена DMD. Характерным для Dp71 является сплай-
сирование экзона 78.
Dpi 16 и Dpi40 и Dp260 в дополнение к С-концевым
доменам имеют дистальные области стержневого домена
дистрофина различной протяженности. Все эти белки так-
же связаны с клеточными мембранами, однако их функ-
ции остаются в значительной степени неясными. Апо-дис-
трофин-2 — Dpi 16, преимущественно экспрессируется у
взрослых в периферических нервах. Размер мРНК самого
маленького апо-дистрофина-3 (Dp40) составляет 2.2 кб
(Tinsley et al., 1993). Чисто нейрональной изоформой дис-
трофина является Dpi40 (Morris et al., 1995). Наиболее
интенсивная экспрессия этого белка в мозге наблюдается
в эмбриональном периоде. Трансляция Dpi40 начинается
с 51 экзона гена дистрофина, а промотор и первый экзон
этой последовательности локализованы в 44 интроне гена
DMD. Показано, что снижение интеллекта, которое харак-
терно для части больных с миодистрофией Дюшенна, до-
стоверно коррелирует с делециями в районе экзонов 40-
52 гена DMD. Эти делеции могут затрагивать синтез или
функции Dpi40. В плексиформном слое сетчатки глаза
избирательно экспрессируется изоформа дистрофина
Dp260. Этот апо-дистрофин участвует в контроле нормаль-
ной электрофизиологической функции сетчатки (D'Souza
et al., 1995). Апо-дистрофины, так же как и полноразмер-
ные дистрофины, способны взаимодействовать с белками
дистрофин-ассоциированного комплекса. Однако из-за
отсутствия в их структуре N-терминальной части дистро-
фина эти белки не способны взаимодействовать с акти-
ном и потому их функции не связаны с поддержанием це-
лостности мембран мышечных волокон.
Уровень транскрипции полноразмерного мышечного
дистрофина в процессе дифференцировки миобластов воз-
растает примерно в 10 раз. Однако активность мышечного
промотора в зрелых миофибриллах в 30 раз ниже, чем в
52

незрелых мышечных волокнах — миотубулах, что указы-
вает на существование дополнительных транскрипционных
элементов, вовлеченных в регуляцию экспрессии гена DMD
s мышцах (Klamut et al., 1996). Одним из них является 5-кб
энхансер, локализованный внутри первого мышечного ин-
трона. Этот элемент специфическим образом активирует-
ся в мышцах. В генно-инженерных опытах in vitro было по-
казано, что мышечный DMD-энхансер обеспечивает повы-
шенный уровень экспрессии генов-репортеров независи-
мо от своего положения и ориентировки. По-видимому,
существуют и дополнительные подобные элементы в дру-
гих частях гена DMD.
Высококонсервативные последовательности шести эк-
зонов, кодирующих С-конец дистрофина, альтернативно
сплайсируются, образуя несколько структурно различающих-
ся изоформ дистрофина, осуществляющих различные функ-
ции. Очевидно, что продукты дистрофинового гена могут вза-
имодействовать со множеством различных белков и не толь-
ко в мышечных и нейрональных тканях (Monaco, 1989). В
настоящее время функции многочисленных изоформ дистро-
фина, обильно экспрессирующихся в различных специали-
зированных тканях и способных взаимодействовать со мно-
жеством белков и не только мышечного или нейронального
происхождения, изучены явно недостаточно.
6. Риртрпфин-ассоииированный комплекс белков
Связь цитоскелета мышечной клетки с экстраклеточ-
ным матриксом осуществляется за счет взаимодействия
полноразмерного дистрофина с дистрофин-ассоциирован-
ным комплексом белков. При разрушении этого комплек-
са мышечные волокна теряют свою структурную целост-
ность и погибают. Кроме того, связывание различных изо-
Ф°рм дистрофина с сарколеммой необходимо для их нор-
мального функционирования в центральной нервной сис-
теме. Оказалось, что дистрофин-ассоциированный ком-
плекс белков имеет гораздо более сложную структуру, чем
53
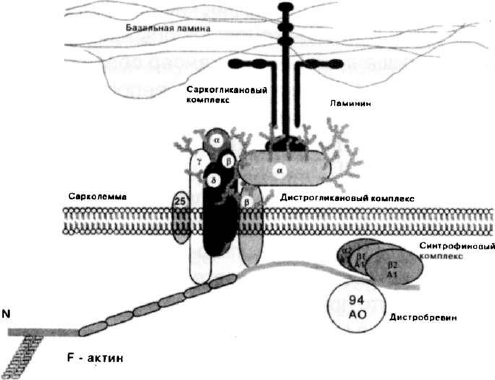
это представлялось вначале, и в нем могут быть выделены
три отдельных субкомплекса — дистрогликановый, сарког-
ликановый и синтрофиновый (рис. 7).
Рис. 7. Дистрофин-ассоциированный комплекс белков
В первый субкомплекс входят два гликопротеина, яв-
ляющиеся коровыми белками для всего комплекса — боль-
шой экстраклеточный белок ct-дистрогликан (156DAG) и не-
посредственно взаимодействующий с ним трансмембранный
р-дистрогликан (43DAG или АЗа). Заякоревание дистрофина
на внутренней поверхности сарколеммы обеспечивается за
счет его связывания с цитоплазматическим хвостом р-дис-
трогликана (Suzuki et al., 1994). а-дистрогликан, взаимодей-
ствуя с компонентом базальной мембраны ламинином 2 (или
мерозином), обеспечивает связь между внеклеточным мат-
риксом и сарколеммой, а через дистрофин — и с цитоскеле-
том клетки. Ламинин-связанный а-дистрогликан, соединен-
ный с экстраклеточным участком р-дистрогликана, форми-
54
рует дистрогликановый субкомплекс (Suzuki et al., 1995). Уча-
сток связывания дистрофина с р-дистрогликаном, начинаю-
щийся в цистеин-богатом районе дистрофина и захватываю-
щий часть концевого С-домена, имеет огромное значение для
функционирования полноразмерных и укороченных изоформ
дистрофина. Так, одна из редких миссенс-мутаций в гене
DMD — C3340Y, сопровождающаяся заменой консерватив-
ного цистеина в дистрогликан-связывающем домене, приво-
дит к тяжелой форме миодистрофии Дюшенна в комплексе с
умственной отсталостью и отсутствием b-волны на электро-
ретинограмме (Lenk et al., 1996). Участок связывания дис-
трофина с р-дистрогликаном содержит три пептидных моти-
ва, найденных в других неродственных дистрофину белках —
WW, EF, ZZ. Эти мотивы кодируются экзонами (63), (65-66),
(68-69), соответственно. Интересно, что р-дистрогликан спо-
собен связываться с src-гомологичным доменом SH3 одного
из адапторных белков (Grb2). SH3 — высококонсервативная
некаталитическая последовательность, участвующая в обра-
зовании стабильного комплекса между scr-киназами и дру-
гими сигнальными молекулами, включая рецепторы. Пред-
полагается, что b-дистрогликан взаимодействует с WW-участ-
ком дистрофина посредством атипичных ЭНЗ-связывающих
модулей (Yang et al., 1995). EF-последовательность может уча-
ствовать в связывании кальция. ZZ-мотив способен связывать
бивалентные катионы металлов, в частности цинка.
Показано, что а- и р-дистрогликаны являются альтер-
нативными продуктами одного гена DAG1, картированного в
области 3р21 (Ibragimov-Beskrovnaya et al., 1992; 1993). Со-
ответствующий белковый продукт, транслирующийся с мРНК
размером 5.5 кб, расщепляется на две нековалентно-ассо-
Циированные субъединицы — аир. Ген DAG1, состоящий из
Двух разделенных большим интроном экзонов, экспрессиру-
втся во множестве типов тканей. В опытах на мышах пока-
зан высокий уровень экспрессии этого гена в раннем эмбри-
огенезе. Дистрогликаны обнаруживаются в децидуальной тка-
н
и уже на периимплантационной стадии развития (Yotsumoto
55
et al., 1996). По-видимому, в этот период они выполняют роль
медиаторов для адгезии между децидуальными клетками или
между децидуальными и трофобластными клетками. Присут-
ствие дистрогликанов необходимо для формирования одной
из первых оболочек зародыша — Рейхеротовской мембра-
ны (Williamson et al., 1997). Роль дистрогликанов в различных
специализированных тканях во многом остается неясной.
В нейромышечных соединениях а-дистрогликан явля-
ется функциональным рецептором для агрина — нейрональ-
но секретируемого гликопротеина, индуцирующего кластери-
рование ацетилхолиновых рецепторов и последующую реор-
ганизацию постсинаптического цитоскелета (Sealock,
Froehner 1994). В ряде работ было показано, что в культиви-
руемых миотубулах антидистрогликановые антитела ухудша-
ют кластерирование ацетилхолиновых рецепторов (Campanelli
et al., 1994; Gee et al., 1994). Показано, что домен агрина,
ответственный за кластерирование ацетилхолиновых рецеп-
торов, физически отделен от домена, осуществляющего связь
с дистрогликаном (Gesemann et al., 1996). Поэтому представ-
ляется более вероятным, что дистрогликан трансдуцирует
сигнал от агрина для начала кластерирования ацетилхолино-
вых рецепторов через промежуточный белок, которым, по
всей видимости является перлекан (Peng et al., 1999). Не ис-
ключено, что именно через эту связь осуществляется учас-
тие дистрофина в синаптогенезе.
Другой саркогликановый субкомплекс формируется
четырьмя трансмембранными белками и включает а-сарко-
гликан (50DAG или адхалин или А2), р-саркогликан (43DAG
или АЗЬ), у-саркогликан (35DAG или А4) и 5-саркогликан
(35DAG). В этот субкомплекс, по-видимому, входит 25DAP (А5
или саркоспан). Связь саркогликанового субкомплекса с дис-
трофином осуществляется посредством его взаимодействия
с цитоплазматическим участком 35DAG. Для сохранения ста-
бильности всего саркогликанового комплекса особое значе-
ние имеет карбокси-терминальный район у-саркогликана. В
настоящее время все гены, кодирующие саркогликаны, кар-
56
тированы и клонированы. Мутации в саркогликановых генах
являются причиной развития различных аутосомно-рецессив-
нь1Х
дюшенно-подобных и конечностно-поясных миодистрофии.
Третий синтрофиновый субкомплекс образован четырь-
мя ассоциированными с дистрофином цитоплазматически-
ми белками — тремя 59DAP, получившими название синтро-
финов или А1 триплета (а-А1, р1 -А1 и р2-А1), а также минор-
ными белками комплекса, относящимися к группе дистроб-
ревинов (АО). Синтрофины связываются с участком С-кон-
цевого домена дистрофина, кодируемым экзонами 73-74,
альтернативно сплайсирующимися во многих типах тканей
на разных стадиях развития (Suzuki et al., 1994; Ahn, Kunkel,
1995). В частности, р1-синтрофин специфически связывает-
ся с расположенной в концевом С-домене последовательно-
стью из 53 аминокислот, кодируемой 74 экзоном дистрофи-
на. а-синтрофин связывается с другим участком дистрофи-
на, локализованным в непосредственной близости от сайта
связывания р1-синтрофина (Suzuki et al., 1995). Возможно,
дистробревин участвует в связи между дистрофином и сар-
когликановым субкомплексом. Синтрофины, по-видимому,
являются сигнальными медиаторами, так как они содержат
протеин-связывающие домены (PDZ). Они могут опосредо-
вать соединение дистрофин-ассоциированного комплекса
белков с киназами, натриевыми каналами и nNOS. Предпо-
лагается также их участие в пострансляционных модифика-
циях DAP-комплекса в мышцах в ответ на изменение меха-
нического стресса. Синтрофины, локализованные в нейро-
мышечных соединениях, и в первую очередь р2-синтрофин,
по-видимому, принимают участие в синаптогенезе.
Дистробревины относятся к семейству дистрофин-
Родственных белков. Их гомология с дистрофином в обла-
сти цистеин-богатого домена достигает 57%. Ген дистроб-
Ревина картирован в области 18q12.1-12.2 и кодирует се-
мейство белков, главными изоформами которых в мыш-
Чах являются дистробревин-1 (94 кД), дистробревин-2 (62
КД) и дистробревин-3 (42 кД) (Sadoulet-Puccio et al., 1996;
57
Metzinger et al., 1997). Обнаружено не менее 5 различных
транскриптов гена дистробревина, три из которых присут-
ствуют в мозге. Фосфорилированный по тирозину дистро-
бревин с молекулярной массой 94 кД имеет 90% гомоло-
гии с 87 кД постсинаптическим белком электрического ска-
та Torpedo (Wagner et al., 1993). He исключено участие дис-
тробревинов в синаптогенезе, так как они вычищаются в
комплексе с ацетилхолиновыми рецепторами. Хотя у
Torpedo дистробревин также ассоциирован с синтрофина-
ми и дистрофином, прямого взаимодействия между этим
комплексом и ацетилхолиновыми рецепторами не обнару-
жено. Иммуногистохимические исследования показывают,
что уровни дистробревина в сарколемме пациентов с мио-
дистрофией Дюшенна и, в меньшей степени, Беккера рез-
ко снижены. Однако присутствия дистрофина недостаточ-
но для правильной локализации дистробревинов в сарко-
лемме, так как аналогичное снижение наблюдается и у
пациентов с конечностно-поясными мышечными дистро-
фиями, обусловленными дефектами в саркогликановых
генах.
В целом организация дистрофин-гликопротеинового
комплекса очень сходна со структурой кадхеринов или ин-
тегринов, а сам этот комплекс является важнейшим эле-
ментом архитектуры мышечных клеток. Очевидно, что во
многих типах клеток позвоночных белки дистрофинового
семейства выполняют критическую роль в поддержании ас-
социированных с мембраной комплексов в местах меж-
клеточных контактов. Но на этом их функция не ограничи-
вается. Наличие многочисленных сайтов фосфорилирова-
ния в цистеин-богатом домене, присутствующем во всех
дистрофин-родственных белках, и возможность его связы-
вания с кальцием, магнием и кальмодулином доказывают
активное участие этих белков в передаче сигналов через
мембрану мышечного волокна (Michalak et al., 1996).Для
большинства дистрофин-ассоциированных белков выпус-
каются в настоящее время коммерческие антитела.
58
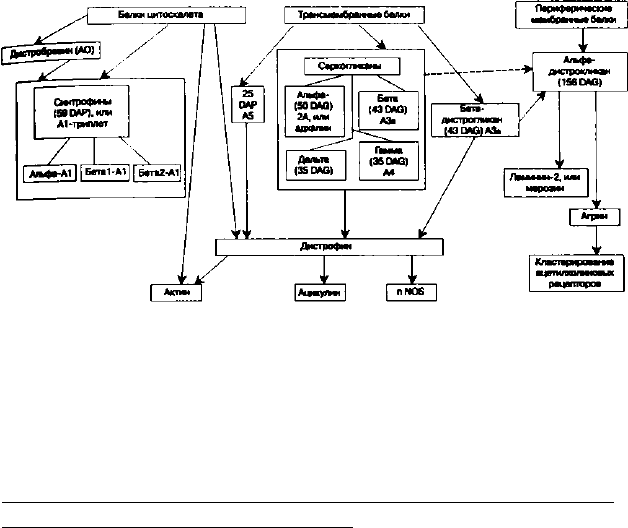
На рис. 8 схематически изображены связи между бел-
ками дистрофин-ассоциированного комплекса.
Рис. 8. Связи между белками дистрофин-ассоциированного
комплекса
7. Анализ корреляций генотип-фенотип. Дюшенно-подобные
аутосомно-реиессивные миопатии
Анализ корреляций между клиническим течением ми-
одистрофии Дюшенна/Беккера и делециями без сдвига рам-
ки считывания, затрагивающими специфические домены
белкового продукта DMD, позволяет строить своеобразную
«патофункциональную» карту дистрофина (Ahn, Kunkel,
1993). Показано, что N-концевые делеций приводят к гете-
рогенным, но достаточно серьезным формам Беккера.
Делеций проксимальной части стержневого домена связа-
ны с очень мягким или атипичным фенотипом, тогда как
Дистальные делеций этого же домена дают типичные фор-
59
мы Беккера. За некоторым исключением делеции двух С-
концевых доменов реализуются в виде миодистрофии Дю-
шенна.
У больных с миодистрофией Дюшенна/Беккера коли-
чественное содержание всех белков гликопротеинового
комплекса, в том числе и внеклеточного альфа-дистрогли-
кана (156DAG), значительно понижено и одновременно
повышен уровень кальция скелетных мышц, "аким обра-
зом, аномальная экспрессия дистрофина может оказывать
влияние на внешние компоненты мышечных вогокон вслед-
ствие разрушения дистрофин-ассоциированно'о комплек-
са, нарушения целостности или эластичности сарколеммы,
ведущего либо к ее механическому повреждению, либо к
изменению механизмов регуляции кальция. Потеря экс-
траклеточного гликопротеина является, по-видимому, пер-
вым шагом в молекулярной цепи патогенеза данных форм
миодистрофии.
Не исключено, что делеции в некодируюдих облас-
тях DMD, специфическим образом нарушающее экспрес-
сию определенных изоформ дистрофина вследствие изби-
рательного разрушения промоторных областей для соот-
ветствующих транскриптов, могут реализоваться в сцеп-
ленные с полом заболевания, не сопровождающиеся мы-
шечной дистрофией. Так, было показано, что делеции об-
ласти локализации мышечного промотора, а также точко-
вые мутации в 5'-сайте сплайсинга первого экзона DMD
являются причиной одной из форм тяжелой Х-сцепленной
дилатационной кардиомиопатии, протекающей без прояв-
лений симптомов мышечной слабости (Muntoni etal., 1995а;
Milasin et al., 1996). Распространенность дилатационной
кардиомиопатии в США составляет 36.5 на 100 000. При-
мерно для 20% подобных случаев показано менделевское
наследование различного типа — аутосомно-доминантное,
аутосомно-рецессивное и сцепленное с полом. При этом
не наблюдается никаких клинических различий между се-
мейными и спорадическими случаями заболевания. Гене-
60