Березов Т.Т., Коровкин Б.Ф. Биологическая химия
Подождите немного. Документ загружается.

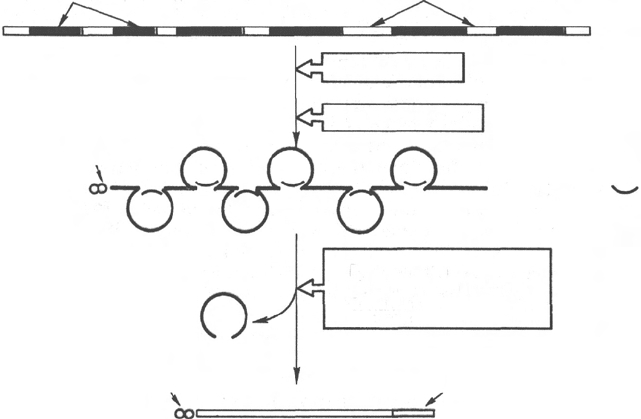
Рис. 13.5. Биогенез мРНК у эукариот.
вероятнее всего, служат сайтами (местами) узнавания для ферментов
сплайсинга. Поскольку 5'-ГУ ... АГ-3' последовательности не открыты
в молекулах предшественников тРНК, было высказано предположение
о существовании по меньшей мере двух типов ферментов сплайсинга;
одного для мРНК и другого для тРНК. Имеются, кроме того, достоверные
данные о том, что интроны часто оказываются длиннее экзонов и что
внутри гена на интроны приходится значительно большая часть нуклеотид-
ных пар. Подсчитано, например, что ген овальбумина содержит 7 интронов,
в общей сложности насчитывающих 7700 пар оснований, в то время как
сформировавшаяся после сплайсинга мРНК насчитывает всего 1859 ос-
нований. Почти во всех эукариотических клетках синтезированные на
структурных генах первичные транскрипты подвергаются процессингу,
прежде чем выполнят свои уникальные функции в белковом синтезе. Во
многих случаях процессинг имеет место главным образом в ядре, хотя этот
процесс продолжается и после транспортировки молекул РНК из ядра
в цитоплазму: например, терминальные реакции полиаденилирования и
метилирования остатков нуклеозидов.
Химический смысл кэпирования сводится к присоединению остатка
7-метилгуанозина посредством трифосфатной группы к 5'-концу молекулы
транскрипта, метилированию 2'-ОН-группы первого и второго нуклеотидов
на 5'-конце мРНК. Полиаденилирование 3'-конца первичного транскрипта
включает ряд стадий и участие эндонуклеазы и полиаденилатполимеразы.
Эндонуклеаза расщепляет мРНК вблизи специфической сигнальной после-
довательности (5')ААУААА(3'), отличающейся высокой консерватив-
ностью. Полиаденилатполимераза синтезирует поли-А-конец (от 20 до 250
нуклеотидов) начиная с точки распада.
Функции 5'-кэп и 3'-поли-А раскрыты недостаточно полно. Показано,
что 5'-кэп, соединяясь со специфическим белком, принимает участие в свя-
491
Интроны
Экзоны
ДНК
Транскрипция
5'-Кэпирование
Кэп
Первичный
транскрипт
3'мРНК
мяРНК
Расщепление
3'-Полиаденилирование
Сплайсинг
Кэп
Поли-А
Зрелая мРНК
зывании мРНК с рибосомой, способствуя инициации синтеза белка. До-
пускают, что основное назначение 5'-кэп и поли-А – защита мРНК от
энзиматического распада. Известно также, что не все цитоплазматические
мРНК содержат участки поли-А на 3'-концах и что в цитоплазме клеток
животных происходит как присоединение, так и удаление участка поли-А из
молекулы мРНК. Следует отметить, что размер молекулы цитоплазма-
тической мРНК даже после удаления 3'-поли-А оказывается все же намного
большим, чем требуется для синтеза кодируемого белка. В частности,
размер мРНК белка глобина (эритроциты кролика) составляет 550 нуклео-
тидов, в то же время кодирующий участок состоит из 430 нуклеотидов
(размер поли-А – 40 нуклеотидов). Другой пример: размер мРНК тяжелого
иммуноглобулина (из клеток миеломы мышей) составляет 1800 нуклеотид-
ных остатков, а кодирующая часть – 1350 нуклеотидов (размер поли-
А – 150–200 нуклеотидов). Интересно, что большинство указанных про-
цессов, если не все, могут регулироваться независимо, изменяя уровень
экспрессии гена. Более того, даже после завершения формирования мРНК
изменения ее стабильности могут оказывать существенное влияние на
экспрессию гена.
В последние годы интенсивно исследуются структура и назначение
нетранслируемых участков генов – интронов. Они различаются по числу,
размерам и топографии. Показано, например, что ген сывороточного
альбумина хотя и содержит всего 6 интронов, но на их долю приходится до
80% этого гена; интроны имеют размеры от 90 до 20000 нуклеотидных пар.
Ген коллагена содержит более 50 интронов. Исключение составляют лишь
гены, кодирующие гистоны, не содержащие интронных структур. Раз-
личают 4 класса интронов. Первый класс открыт как в ядерных, так
и в митохондриальных генах, кодирующих рибосомные рРНК; второй
класс интронов открыт в первичных транскриптах митохондриальных
матричных мРНК. Оказалось, что оба эти класса интронов не нуждаются
ни в источнике энергии, ни в участии ферментов, но наделены способностью
самосплайсинга. Третий – самый большой класс интронов обнаружен в
первичных транскриптах ядерных мРНК, подвергающихся созреванию.
Сплайсинг требует наличия комплекса белков и особой группы клеточных
РНК, названных малыми ядерными РНК (мяРНК). Выделено и охарак-
теризовано 5 групп богатых уридином мяРНК, соответственно обозна-
чаемых U1, U2, U4, U5 и U6, размерами от 100 до 200 нуклеотидов.
Комплексы мяРНК и белков, названные малыми ядерными нуклеопро-
теинами, объединяются в единую систему – сплайсосому, координиру-
ющую весь процесс сплайсинга. Предполагают, что мяРНК соединяются
с обеими концами интрона, способствуя формированию специфической
конформации, необходимой для узнавания ее участвующими в процессе
ферментами, сближению двух экзонов, удалению интронов и воссоеди-
нению кодирующих экзонов. Четвертый класс интронов открыт в ряде
тРНК. Сплайсинг этой группы интронов требует доставки энергии и
присутствия эндонуклеаз и лигаз, катализирующих соответственно разрыв
фосфодиэфирных связей с 5'- и 3'-концов интрона и соединяющих два
экзона.
Укажем также на весьма интересные и новые данные о существовании
в структуре мРНК-предшественника, помимо экзонов и интронов, особых,
так называемых альтернативно сплайсируемых, последовательностей. Вы-
явлены примеры неоднозначного протекания сплайсинга для ряда генов.
Результат альтернативного сплайсинга – появление нескольких продуктов
при экспрессии одного гена. Так, получены доказательства, что экспрессия
492
одного и того же гена тропомиозина позволяет получить семь изоформных
белков, специфичных для разных групп мышц (гладких и поперечно-
полосатых) или для фибробластов и миобластов. В то же время известны
примеры формирования одного белкового продукта (например, олиго-
мерного фермента глюкозо-6-фосфатдегидрогеназы) при экспрессии двух
разных генов. Все эти данные свидетельствуют о том, что альтернативный
сплайсинг может играть существенную роль в функционировании генома
клеток высших организмов.
В нетранскрибируемых последовательностях генома перед экзон-
интронами открыты специфические участки, названные промоторами, а
также энхансерами (повышающие уровень транскрипции) и силан-
серами (ослабляющие уровень транскрипции). При взаимодействии с
белками они выполняют функции регуляторных сигналов при транскрип-
ции. Этот способ регуляции широко используется клетками эукариот как
в процессах дифференцировки, так и при индукции репрессии (см. главу 14).
Нельзя не упомянуть об открытии рибозимов, т.е. молекул РНК,
выступающих в качестве катализатора. Пожалуй, это единственные из
известных макромолекул, которые наделены как информационной, так
и каталитической функцией. Открытие каталитических РНК поколебало
само понятие «фермент». Оказалось, что некоторые РНК осуществляют
посттранскрипционный процессинг, катализируя самосплайсинг, т.е. участ-
вуют в разрезании и удалении интронов. Наделенные рядом свойств
истинных и эффективных катализаторов рибозимы участвуют в двух типах
реакций: в гидролизе (разрыве) фосфодиэфирной связи и в реакциях
трансэтерификации. В качестве субстрата могут служить, помимо собствен-
ного, предшественник (про-РНК) и другие молекулы РНК. Сейчас ин-
тенсивно изучается третичная структура рибозимов, а первичная и вторичная
структуры ряда из них уже расшифрованы. Эти исследования, несомненно,
интересные сами по себе, могут пролить свет и на пути развития биоло-
гической эволюции.
Для полного понимания молекулярных механизмов сложного процесса
биогенеза мРНК предстоит решить множество вопросов. В частности,
необходимо выделить в чистом виде и охарактеризовать белковые фак-
торы, принимающие участие в этой регуляторной системе. Далее следует
раскрыть механизмы узнавания промотора, терминации и антитерминации,
избирательного метилирования, а также тонкие молекулярные механизмы
регуляции сплайсинга. Решение указанных проблем будет, несомненно,
способствовать лучшему пониманию сущности механизмов регуляции
экспрессии генов эукариотических клеток в норме и при патологии.
Биогенез транспортных РНК
Транспортные РНК в клетке выполняют адапторную функцию при тран-
сляции информации мРНК в первичную структуру белка. Как было указано
в главе 3, в молекуле тРНК содержится 8–10% необычных («минорных»)
азотистых оснований в составе нуклеотидов. Молекулы тРНК как у
эукариот, так и у прокариот синтезируются в виде больших предшествен-
ников, часто содержащих последовательности более одной тРНК, которые
затем подвергаются нуклеолитическому процессингу при участии специ-
фических рибонуклеаз. Гены некоторых тРНК содержат вблизи участка
ДНК, ответственного за синтез антикодоновой петли, интронные после-
довательности (около 18 нуклеотидов). Эти участки также транскриби-
493
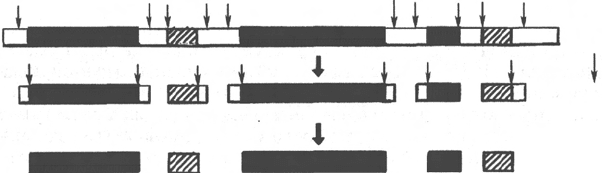
руются, поэтому процессинг тРНК включает, помимо удаления 18-членного
рибонуклеотидного интрона, также необходимый сплайсинг антикодоновой
области. Дальнейшая модификация включает присоединение триплета
ЦЦА и образование акцепторного участка (на 3'-конце молекулы), к
которому присоединяется аминокислота. Имеются данные, что метили-
рование предшественников тРНК у эукариот осуществляется в ядре, в то
время как ферментативные процессы удаления интрона и присоединения
триплета ЦЦА происходят, скорее всего, в цитоплазме. Помимо акцеп-
торного и антикодонового участков, тРНК содержит специфичные участки
узнавания для ферментов – аминоацил-тРНК-синтетаз, а также участки для
связывания с большими субчастицами рибосом (см. далее главу 14).
Биогенез рибосомных РНК
У прокариот синтез 23S, 16S и 5S рРНК осуществляется из более крупного
30S предшественника, получившего название прерибосомной РНК (пре-
рРНК). Под действием специфических нуклеаз и метилаз из этого общего
предшественника в результате процессинга сначала образуются проме-
жуточные рибосомные РНК, которые, подвергаясь дальнейшей нуклеазной
атаке и метилированию, превращаются в зрелые молекулы (рис. 13.6).
Для ряда эукариотических клеток доказана транскрипция двух вы-
сокомолекулярных рРНК (18S и 28S) и одной низкомолекулярной рРНК
(5,8S) из одного общего предшественника (45S); последний представляет
собой продукт генов рРНК, более тысячи копий которых содержит клетка.
Первичный транскрипт 45S рРНК высокометилирован, причем метили-
рованию в ядрышках подвергаются только те участки первичного транс-
крипта, из которых в процессе реакций процессинга образуются рРНК. Сам
механизм процессинга первичного транскрипта резко отличается от про-
цессинга гяРНК при образовании мРНК. Образовавшаяся, например, мо-
лекула 28S рРНК еще в ядрышке подвергается дальнейшему метили-
рованию, затем она взаимодействует с синтезированными в цитоплазме
рибосомными белками и формируется 60S рибосомная субчастица; 18S
рРНК аналогичным способом участвуют в формировании 40S рибосомной
субчастицы. Обе субчастицы стабильны в делящихся клетках и нестабильны
в неделящихся клетках.
Следует указать, что синтез РНК при участии ДНК-зависимой РНК-
полимеразы специфически тормозится антибиотиком актиномицином D,
который обладает способностью связываться водородными связями с ДНК
Рис. 13.6. Постсинтетическая модификация пре-рРНК прокариот (по Николову).
494
Пре-16S РНК
Пре-23S РНК
Пре-5S РНК
Пре-рРНК
Места эндо-
нуклеазной
атаки
16S РНК
тРНК
23S
РНК
5S РНК
тРНК
по месту остатков гуанина. Актиномицин D тормозит синтез РНК в ин-
тактных клетках. Он нашел широкое применение при определении про-
цессов, зависящих от транскрипции ДНК.
Синтез РНК на матрице РНК
ДНК-зависимая РНК-полимераза может осуществлять транскрипцию ДНК
нормальных клеток и ДНК-вирусов. Как же осуществляется синтез РНК
у тех вирусов, которые в геноме вместо ДНК содержат РНК? Оказывается,
в этих случаях вирусная РНК индуцирует образование в клетках хозяина
(например, у Е. coli) РНК-зависимой РНК-полимеразы, которая участвует
в репликации вирусной РНК (отсюда второе название фермента – РНК-
репликаза). Фермент также используется нуклеозидтрифосфаты для син-
теза одноцепочечной вирусной РНК. Этот синтез должен пройти через
стадию образования репликативной формы. Следовательно, на I стадии
РНК-репликаза на матрице РНК-вируса специфически строит комплемен-
тарную, с противоположной полярностью цепь РНК. Последняя на II
стадии служит матрицей для синтеза РНК, совершенно однотипной ис-
ходной вирусной РНК. Обе стадии катализируются одним и тем же
ферментом, хотя в каждой из них участвуют различные белковые факторы.
Следует особо подчеркнуть, что, поскольку РНК-репликаза имеет от-
ношение только к вирусам, очевидно, на этом основании могут быть
разработаны эффективные антивирусные лекарственные препараты.
Синтез РНК из нуклеозиддифосфатов. М. Грюнберг-Манаго и С. Очоа
в 1955 г. в клетках Е. coli открыли особый фермент – полинуклеотид-фос-
форилазу. Этот фермент наделен способностью синтезировать in vitro
полимерную молекулу РНК из однотипных или разных рибонуклеозид-
дифосфатов (НДФ). Реакция, являющаяся обратимой, протекает по урав-
нению:
(НМФ)
n
+ НДФ –> (НМФ)
n+1
+ Н
3
РO
4
.
Рибонуклеозидтрифосфаты и дезоксирибонуклеозидтрифосфаты не яв-
ляются субстратами фермента. Фермент не нуждается в матрице, однако
для синтеза необходима затравочная цепь РНК (НМФ)
n
со свободной
3'-гидроксильной группой, к которой присоединяются остатки моно-
нуклеотидов. Образовавшаяся полимерная молекула РНК не имеет за-
данной специфической последовательности мононуклеотидов, но содержит
3'–>5' фосфодиэфирные связи, легко разрываемые рибонуклеазой. Отно-
сительно биологической роли этого фермента у бактерий предполагают,
что он катализирует, скорее всего, обратную реакцию – расщепление мРНК
с образованием нуклеозиддифосфатов.
Полученные в лаборатории С.С. Дебова данные свидетельствуют о
более широком распространении полирибонуклеотид-фосфорилазы в жи-
вых организмах, чем это признавалось ранее. Фермент открыт также
в клетках животных. Кроме того, получены экспериментальные дока-
зательства синтетической функции полинуклеотид-фосфорилазы. Вполне
правомерно допущение, что этот фермент может принимать участие в
синтезе коротких полирибонуклеотидов в клетках эукариот в норме и
в некоторых экстремальных условиях. Кроме того, в лабораторных ус-
ловиях фермент может найти применение для синтеза РНК-праймеров,
используемых далее при синтезе ДНК.
495
Проблемы генетической инженерии. Генетическая инженерия, по опре-
делению А.А. Баева, представляет собой систему экспериментальных
приемов, позволяющих создавать в лаборатории (в пробирке) искусствен-
ные биологические структуры. В качестве инструментов для генно-
инженерных операций применяются созданные самой природой ферменты:
одни из них рассекают молекулу ДНК в строго определенных участках
(рестриктазы), другие, напротив, сшивают разрозненные участки в еди-
ное целое (лигазы). Конечной целью генетической инженерии является
получение организмов (животных и растений) с новыми наследственными
свойствами с помощью лабораторных приемов. Для достижения этой пока
еще отдаленной цели необходимо проведение огромной работы на уровне
отдельного гена или генов. Ген, представленный определенным участком
ДНК и соответствующий определенному белку, можно или выделить из
другого организма, или синтезировать химическим либо биологическим
путем. Впервые в 1969 г. из Е. coli был выделен участок ДНК с геном,
ответственным за синтез фермента, катализирующего усвоение молочного
сахара (лактозы),– так называемый лактозный оперон. Химический синтез
гена аланиновой тРНК впервые осуществил Хар Гобинд Корана в 1970 г.
Состоящий из 72 нуклеотидов, этот ген, однако, лишен функциональной
активности, так как в клетках тРНК синтезируется не в готовом виде,
а в форме предшественника. Эти данные послужили для Кораны основой
для синтеза гена-предшественника тирозиновой тРНК (из 126 нуклеоти-
дов), хотя сама тирозиновая тРНК состоит из 85 нуклеотидов. Ввиду
громоздкости, а также недостаточной эффективности химического синтеза
в последние годы все большее место занимают биологические методы
синтеза генов при помощи обратной транскриптазы (ревертазы).
Для этого необходимо иметь мРНК, с помощью которой можно вос-
произвести соответствующий ген. Синтезированы ДНК-копии на мРНК,
кодирующие синтез белка глобина (человека, кролика, мыши, голубя, утки),
иммуноглобулина, белка хрусталика глаза и др. Однако на этом пути
синтеза генов встречаются большие трудности, связанные с выделением из
огромного разнообразия клеточных мРНК, нужной для синтеза гена.
Следующий этап генетической инженерии – перенос генов в клетку –
осуществляется тремя способами: трансформацией (перенос генов
посредством выделенной из клеток и освобожденной от примесей ДНК),
трансдукцией (перенос генов посредством вирусов) и гибридизацией
клеток, полученных из разных организмов (высших животных, микро-
организмов и др.) (рис. 13.7, 13.8). Заключительный этап этих экспери-
ментов сводится к адаптации введенного гена в организме хозяина, но он
почти не зависит от искусства экспериментатора.
Исследования в области генетической инженерии могут служить основой
для решения практических задач здравоохранения и сельского хозяйства.
Полученные в лаборатории искусственные гены, помимо широкого ис-
пользования в микробиологической и фармацевтической промышленности
для приготовления кормового белка и лекарственных препаратов (инсулин,
интерферон, гормон роста, гормоны щитовидной железы, стимуляторы
иммунитета и др.), возможно, смогут применяться при лечении многих
наследственных заболеваний (их насчитывается около 5000), генетический
дефект которых точно известен пока только для небольшого числа (не более
50) болезней.
Первые попытки применения лактозного гена при галактоземии (на-
следственное заболевание, связанное с непереносимостью галактозы вслед-
ствие отсутствия фермента гексозо-1-фосфат-уридилилтрансферазы; см.
496
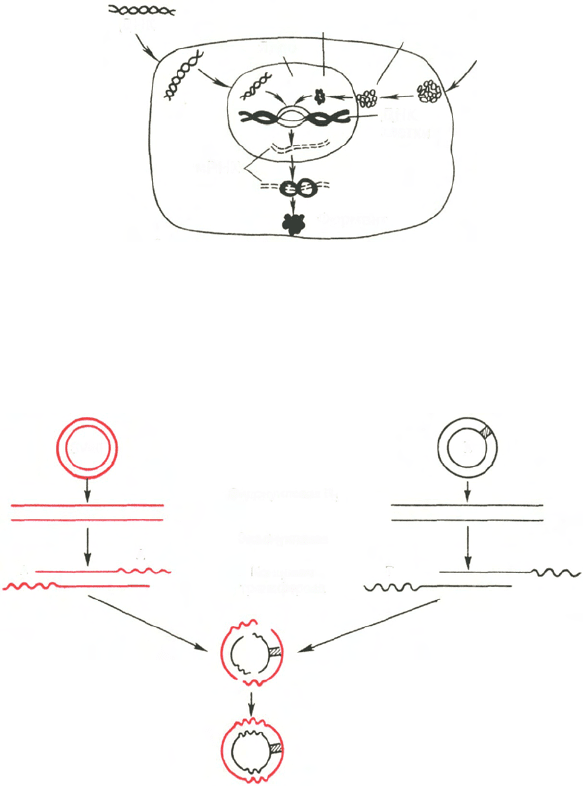
Рис. 13.7. Схематическое изображение двух способов введения генов в клетку –
трансформации и трансдукции (по А. А. Баеву).
Рис. 13.8. Получение гибридной молекулы, содержащей одновременно ДНК вируса
SV40, ДНК фага λ, и галактозный оперон (схема по А.А. Баеву).
Под действием эндонуклеазы R
1
Е. coli кольцевые ДНК разрываются в одной точке, в резуль-
тате образуются линейные нити. Под действием другого фермента - экзонуклеазы (из фага)
укорачиваются нити ДНК с противоположных концов. Далее при помощи фермента концевой
трансферазы наращиваются нити ДНК, причем у одной ДНК новые концы состоят из
адениловых (А), у другой - из тимидиловых (Т) остатков. При смешивании молекул концевые
остатки А и Т образуют комплементарные пары, замыкая линейные молекулы в кольца.
Вначале эти кольца содержат 4 разрыва, которые затем закрываются при участии еще одного
фермента - ДНК-лигазы.
Трансформация
Трансдукция
ДНК
ДНК
вируса
Белковая
оболочка
вируса
Вирус
Ядро
мРНК
ДНК
клетки
Фермент
Вирус
SV40
Фаг
λ
Галактозный
оперон
Эндонуклеаза R
1
Экзонуклеаза
Концевая
трансфераза
А
А
Т
Т
ДНК-полимераза
ДНК-лигаза
Гибридная молекула
(с 4 разрывами)
Гибридная молекула,
замкнутая в кольцо
главу 10) вселяют надежду на реальные практические возможности гене-
тической инженерии, хотя вполне обоснованы тревога и опасения, свя-
занные с вмешательством человека в сферу тончайших биологических
процессов наследственного аппарата целостного организма. В последние
годы, после бурного периода расцвета, в генетической инженерии наблю-
дается некоторый спад, обусловленный недостаточностью знаний о струк-
туре и функционировании генома клеток эукариот. Переход от иссле-
дований на клетках прокариот к исследованиям на клетках эукариот
оказался затруднен рядом технических сложностей вследствие мозаич-
ности структуры генов последних. В частности, открытие эк-
зонов и интронов в геноме, явления сплайсинга (формирование зрелой
матричной РНК) указывает на необходимость соблюдения высочайшей
точности процедуры вырезания необходимого гена из ДНК генома со-
ответствующими рестриктазами. В противном случае могут быть получены
не структурные транслируемые гены, а интроны или участки экзонов, не
кодирующие белок. После того как были разработаны методы искусствен-
ного синтеза и сшивки отдельных участков молекулы ДНК, появилась
возможность конструирования и создания новых, неизвестных ранее в
природе организмов с заранее заданными свойствами. Современная
биотехнология явилась логическим развитием этого направления науки. Она
сложилась на основе фундаментальных достижений биохимии, генетики
и микробиологии, открыв широкие возможности для создания новых
сортов растений, новых пород животных и т.п. Учитывая исключительную
важность биотехнологии для народного хозяйства, в 1985 г. в нашей стране
был создан и успешно работает межотраслевой научно-технический ком-
плекс (МНТК) «Биоген». Комплекс был призван обеспечить создание
и организацию промышленного производства новых биологически ак-
тивных веществ и препаратов для медицины, ветеринарии, растениеводства
на основе прогрессивных биотехнологических методов, в том числе методов
клеточной и генетической инженерии.
Распад нуклеиновых кислот
Полимерные молекулы нуклеиновых кислот расщепляются в тканях
преимущественно гидролитическим путем при участии специфических фер-
ментов, относящихся к нуклеазам. Различают эндонуклеазы, разрывающие
внутренние межнуклеотидные связи в молекулах ДНК и РНК, вызывающие
деполимеризацию нуклеиновых кислот с образованием олигонуклеотидов,
и экзонуклеазы, катализирующие гидролитическое отщепление концевых
мононуклеотидов от ДНК и РНК или олигонуклеотидов. Помимо гидро-
литических нуклеаз, имеются ферменты, катализирующие распад нуклеино-
вых кислот, например, посредством трансферазной реакции. Они ката-
лизируют перенос остатка фосфорной кислоты от 5'-го углеродного атома
рибозы одного мононуклеотида ко 2'-му углеродному атому соседнего
мононуклеотида, сопровождающийся разрывом межнуклеотидной связи
и образованием фосфодиэфирной связи между 2'-м и 3'-м углеродными
атомами рибозы одного и того же мононуклеотида. К настоящему времени
открыты группы нуклеаз, катализирующие распад ДНК и РНК.
Дезоксирибонуклеазы I катализируют разрыв внутренних фосфодиэфир-
ных связей в одной из двух цепей молекулы ДНК между 3'-м углеродным
атомом дезоксирибозы и остатком фосфата с образованием низкомоле-
кулярных олигодезоксирибонуклеотидов:
498
ДНК + (n–1) Н
2
O –> n-Олигодезоксирибонуклеотиды.
Среди продуктов реакции открыты также моно- и динуклеотиды. Ти-
пичными представителями этих ферментов являются ДНКазы поджелу-
дочной железы. Одна из них (ДНКаза I) была получена в чистом виде,
расшифрована последовательность всех ее 257 аминокислотных остатков.
Фермент наиболее активен при рН 6,8–8,0, активируется двухвалентными
ионами Mg
2+
и Мn
2+
и ингибируется конечными продуктами фермен-
тативной реакции – олигонуклеотидами.
Дезоксирибонуклеазы II вызывают деполимеризацию молекулы ДНК
в результате парных разрывов фосфодиэфирных связей обеих цепей ДНК
с образованием более крупных олигодезоксирибонуклеотидов. Предста-
вителем их является ДНКаза II, выделенная из селезенки, имеющая мол.
массу 38000 и состоящая из 343 аминокислотных остатков. В составе этой
ДНКазы открыт глюкозамин. Фермент также активируется ионами ме-
таллов, ингибируется анионами; его оптимум рН между 5,5 и 5,8.
Помимо этих ферментов, открыты (преимущественно у микроорга-
низмов) еще экзодезоксирибонуклеазы, гидролизующие фосфодиэфирные
связи молекулы ДНК с отщеплением концевых 5'-дезоксирибонуклеотидов.
Например, из E. coli выделено четыре таких фермента, обозначаемых
экзодезоксирибонуклеазами I, II, III и IV.
Рестриктазы – ферменты ДНКазного типа действия – катализируют
распад чужеродной (в основном фаговой) ДНК в строго определенных
участках молекулы, имеющих структуру палиндромов. Из E. coli выделены
и охарактеризованы две такие рестриктазы, обозначаемые EcoRI и EcoRII
соответственно. Рестриктазы оказывают строго специфическое действие,
поэтому они используются для расшифровки последовательности нуклео-
тидных остатков в ДНК фагов и вирусов. Кроме того, это уникальное
свойство рестриктаз находит все большее практическое применение в
генетической инженерии при «вырезании» определенных фрагментов ДНК
и «встраивании» их в геном бактериальной ДНК (получение рекомби-
нантных ДНК). В результате клетке передается ряд не свойственных ей
прежде наследственных признаков. Теоретическое и главным образом
практическое значение подобных исследований трудно переоценить. Сви-
детельством огромного интереса к проблемам генетической инженерии
является создание и успешное выполнение в институтах Российской АН
и лабораторий ряда стран совместной комплексной программы – проекта
«Рестриктазы».
Многие сотни рестриктаз выделены в очищенном состоянии и уже
являются коммерческими препаратами.
Из ферментов, катализирующих гидролитический распад РНК, наиболее
изучены рибонуклеазы I. Они гидролизуют фосфодиэфирные связи внутри
молекулы РНК. Выделенная из поджелудочной железы многих животных
РНКаза состоит из 124 аминокислотных остатков во всех случаях, хотя
ферменты несколько различаются последовательностью аминокислотных
остатков; выяснена также третичная структура ряда РНКаз (см. главу 4).
Получен в гомогенном состоянии из плесневого гриба рода Aspergillus
фермент гуанилрибонуклеаза, катализирующая эндонуклеолитическое рас-
щепление РНК.
Из ферментов, осуществляющих распад ДНК и РНК не по гидро-
литическому пути, следует назвать полинуклеотид-фосфорилазу и группу
ДНК-гликозидаз. В настоящее время подробно изучены физико-химические
свойства и биологическая роль микробной полинуклеотид-фосфорилазы
499
в лаборатории С.С. Дебова; в той же лаборатории фермент открыт
в животных тканях.
Механизм действия фермента сводится к переносу нуклеотидных ос-
татков с РНК на неорганический фосфат, при этом образуется рибо-
нуклеотиддифосфат (РДФ):
РНК + Н
3
РО
4
—> (РНK)
n–1
+ РДФ
Предполагают, что in vivo фермент катализирует распад клеточных
РНК, преимущественно мРНК, до нуклеозиддифосфатов, участвуя тем
самым в регуляции концентрации клеточного неорганического фосфата.
Следует указать еще на одну не менее важную уникальную функцию
полинуклеотид-фосфорилазы – способность фермента катализировать в
опытах in vitro синтез из свободных нуклеозиддифосфатов (НДФ) поли-
рибонуклеотидов с заданной последовательностью. Этот фермент сыграл
выдающуюся роль в расшифровке кода белкового синтеза в лабораториях
лауреатов Нобелевской премии С. Очоа и М. Ниренберга (см. главу 15).
Открыта группа ДНК-гликозидаз, участвующих в реакциях отщепления
модифицированных пуриновых и пиримидиновых оснований (например,
урацила, образующегося при дезаминировании остатка цитозина в одной из
цепей ДНК).
Таким образом, ДНК-гликозидазы выполняют важную функцию в
процессах репарации (восстановление структуры) молекулы ДНК.
В результате последовательного действия разнообразных клеточных
экзо- и эндонуклеаз нуклеиновые кислоты подвергаются распаду до стадии
рибо- и дезоксирибонуклеозид-3'- и 5'-фосфатов. Дальнейший распад об-
разовавшихся продуктов связан с ферментативными превращениями моно-
нуклеотидов *, нуклеозидов и далее свободных азотистых оснований. На
I этапе гидролиза действуют 3'- и 5'-нуклеотидазы, катализирующие гидро-
литический распад мононуклеотидов до свободных нуклеозидов с от-
щеплением неорганического фосфата соответственно от С-3' или С-5'
атомов углеводного остатка. На II этапе происходит перенос остатка
рибозы от нуклеозида на свободную фосфорную кислоту с образованием
рибозо-1-фосфата и свободного азотистого основания.
Распад пуриновых нуклеозидов
Образовавшиеся при гидролизе пуриновые нуклеозиды – аденозин и гуано-
зин – подвергаются ферментативному распаду в организме животных
вплоть до образования конечного продукта – мочевой кислоты, которая
выводится с мочой из организма. У человека, приматов, большинства
животных, птиц и некоторых рептилий мочевая кислота является конечным
продуктом пуринового обмена. У других рептилий и некоторых млеко-
питающих мочевая кислота расщепляется до аллантоина и у рыб – до
аллантоиновой кислоты и мочевины. Последовательность всех этих пре-
вращений, катализируемых специфическими ферментами, можно предста-
вить в виде следующей схемы:
* АМФ может подвергаться в животных тканях обратимому дезаминированию в
инозиновую кислоту.
500