Алексеев В.В. Физическое и математическое моделирование экосистем
Подождите немного. Документ загружается.

веществ поколениями организмов
с
последующим возвращением
их
в
водную
среду,
либо вызывается деятельностью человека,
на-
пример,
в
случае
поступления биогенов (фосфаты, нитраты)
в
во-
доем
со
сточными водами предприятий.
Процесс перестройки природного сообщества
при
биогенной
сукцессии очень сложен. Ниже
будет
исследована простейшая
модель сукцессии, описывающая изменение видового состава
био-
ценоза
в
зависимости
от
общего
за-
паса лимитирующего биогенного
элемента
в
системе.
Рассмотрим модельный биоце-
ноз,
состоящий
из
двух
конкури-
рующих трофических цепочек «про-
дуцент— консумент
I
порядка
—
консумент
II
порядка»
[62]. Счи-
таем,
что
консументы строго
спе-
циализированы
по
питанию. Схема
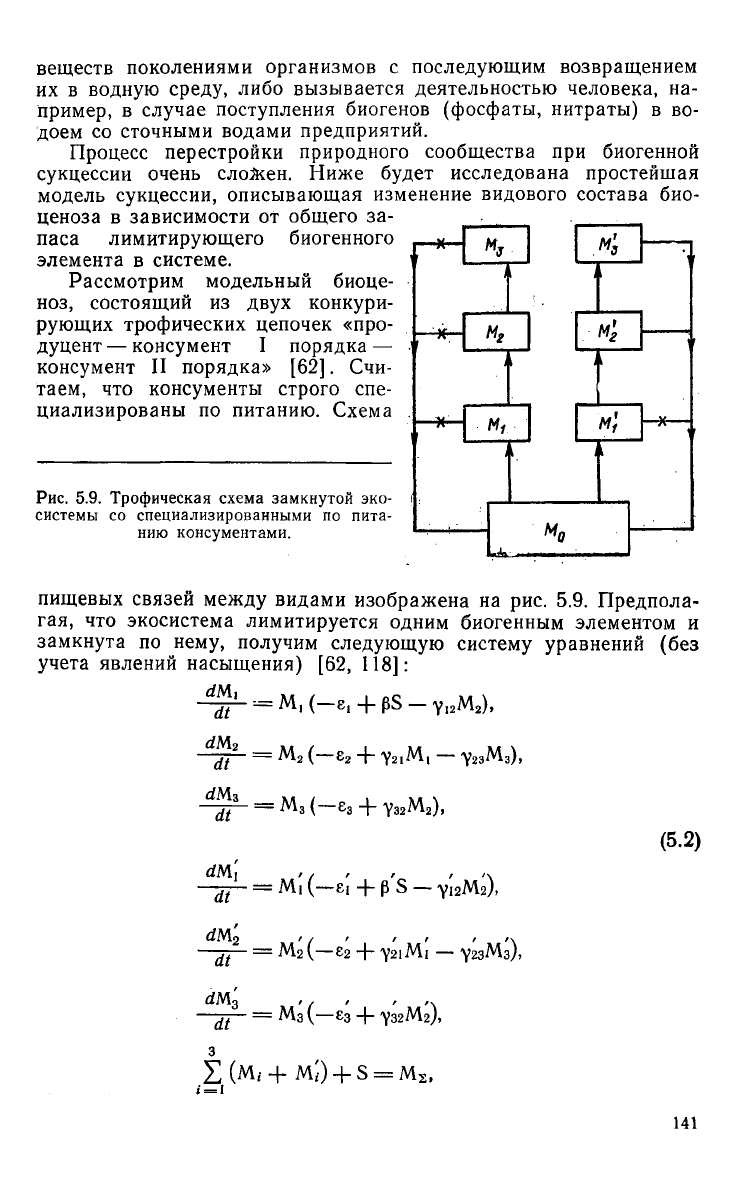
Рис.
5.9.
Трофическая
схема
замкнутой
эко-
системы
со
специализированными
по
пита-
нию консументами.
J
м,
Mi
i
1
м)
*1
1
пищевых связей
между
видами изображена
на
рис. 5.9.
Предпола-
гая,
что
экосистема лимитируется одним биогенным элементом
и
замкнута
по
нему, получим
следующую
систему уравнений
(без
учета
явлений насыщения)
[62, 118]:
dt
=
M
2
(—e
2
•
-
=
M
3
(—e
3
•
(5.2)
4
-_
= Mi(-e,+pS--
-~
=
M
2
( —
62
+
Y2lM!
—
Y23M3),
йЩ
,., , ,
-gj—
=
M3
(—e
3
+
Y32M2),
Z
(Mi
-f
Mi)
+
S
=
M
2>
141
где
Mi,
Mi
—
биомассы
двух
видов продуцентов;
М
2
,
Мг
—
био-
массы консументов
I
порядка;
Мз,
Мз
—
биомассы консументов
II
порядка
(все
биомассы выражены
в
единицах лимитирующего
биогенного элемента);
M
s
—
общий запас лимитирующего
эле-
мента
в
системе;
S —
концентрация
его в
среде;
е,-,
е*—скорости
распадных процессов;
р,
Р'— коэффициенты фотосинтеза;
Yi2, V12»
723, Т2з
—
коэффициенты потребления;
721. 721. 7зг,
7зг—коэффи-
циенты
усвоения пищи консумен-
тами. Очевидно,
что
между
коэф-
фициентами
системы имеют место
неравенства
г i
Y2I
<
YI2!
Y2I < Y12-
(5.3)
Y32
<
Y23!
Y32 < Y23.
поскольку прирост биомассы попу-
ляции
не
может превышать коли-
чества
съеденной пищи.
Рис.
5.10.
Трофические
структуры,
устой-
чивое существование которых
в
системе
(5.2) невозможно.
Рассмотрим, какие стационарные состояния возможны
в си-
стеме
(5.2) при
различных значениях запаса биогенного элемента
М
2
(биологический смысл имеют только положительные значения
биомасс видов
и
коэффициентов модели). Устойчивость
будет
ис-
следоваться
по
отношению
к
малым отклонениям
от
стационарных
значений
биомасс:
(!)
Tit АД (2) Т7' »«'
\i/
= М/ —
М
;
-;
(А/ =
М/
— М/.
В общем виде уравнения
для
отклонений
ц/
1)
имеют
вид [60]
du.
{
P
\"i (
дМ.Р dMJ
0
\
'—
=-.
у I J |_ -г- I
dt
.£—>.
\
(ЭМА
dM.fr
)
L\ am
(/=1,3;
i =1,2).
(5.4)
Решение
системы
(5.4)
имеет
вид ц}
1)
=
exp(W),
где
%
k
—
корни
характеристического уравнения. Стационарное состояние
си-
стемы
(5.2)
асимптотически устойчиво
при
условии Re^<0.
Нетрудно видеть,
что
экосистема, описанная уравнениями
(5.2), может иметь
12
стационарных состояний. Трофические
структуры
и
стационарные значения переменных приведены
142

в
табл. 5.1 (для симметричных
структур
указана только одна из
двух
возможных).
Четыре трофические структуры, представленные на рис. 5.10,
в
стационарном состоянии не
существуют,
так как системы алгеб-
раических уравнений для этих стационаров оказываются вырож-
денными.
Невозможность сосуществования
двух
продуцентов, кон-
курирующих за один компонент питания (рис. 5.10 а), отражает
выполнение
в системе принципа Гаузе. Невозможность стационар-
ных структур, показанных на рис. 5.10 6, в, г, по-видимому, свиде-
тельствует
о том, что стабилизирующее влияние, которое обычно
оказывает присутствие в экосистеме растительноядного консу-
мента, в определенной степени нейтрализуется наличием хищника
II
порядка.
Рассмотрим, какие трофические структуры оказываются устой-
чивыми в системе (5.2) при медленном увеличении параметра
М
2
— общего запаса лимитирующего биогенного элемента.
Структура 1. Содержание биогенного элемента в среде может
быть слишком малым, чтобы обеспечить устойчивое существова-
ние
даже
одной популяции продуцентов. Единственно возможным
в
этом
случае
является состояние с нулевыми значениями биомасс
видов. При исследовании на устойчивость корни характеристиче-
ского уравнения системы (5.4) получаем в виде
ei;
A,
2
=
P'M
£
—ei; А,
3
°
==
— 4'
}
; Х
2
=—г
2
;
А.з=—
ез,
откуда
следует,
что нулевое состояние устойчиво в интервале зна-
чений
параметра М
£
0<Mz<M
1Kp
,
(5.5)
где MiK
P
= min(ei/P; el/P').
Выражения ei/p и ei/p' имеют смысл минимальных концентра-
ций
лимитирующего биогенного элемента в среде, необходимых
для существования изолированных популяций продуцентов Mi
и
Mi.
Будем считать для определенности, что
ei/p < el/p', (5.6)
т. е. наиболее неприхотливым по отношению к дефициту биоген-
ного элемента S является продуцент Mj.
Структура 2. При значениях М
2
, больших Мир, становится
возможным существование в системе популяции фотосинтезирую-
щих растений вида М
ь
Структура 2 оказывается положительно
определенной и устойчивой при выполнении неравенства (5.6) на
отношения
е/р в интервале значений параметра М
2
М
1кр
< М
2
< М
2КР
(5.7)
(значение
М
2к
р приведено в табл. 5.1).
143
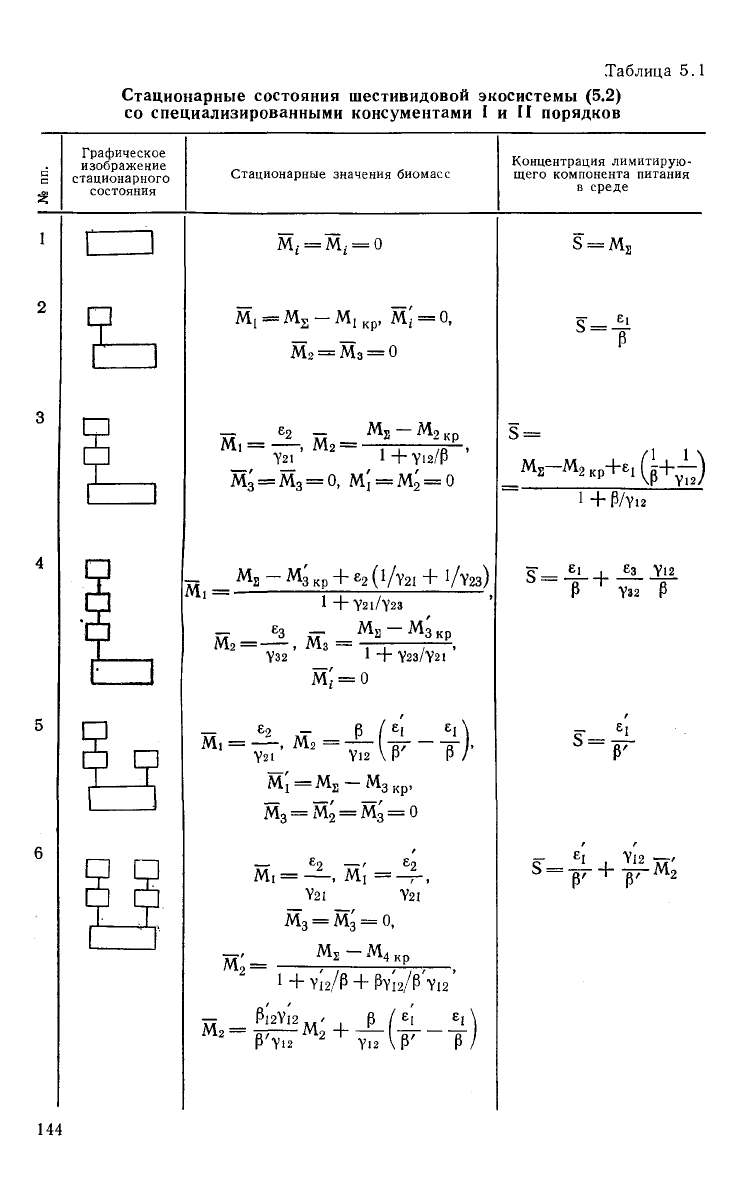
Таблица 5.1
Стационарные состояния шестивидовой экосистемы (5.2)
со специализированными консументами I и II порядков
Графическое
изображение
стационарного
состояния
Стационарные значения биомасс
Концентрация лимитирую-
щего
компонента питания
в
среде
S = —
е
2
М
а
-М2
Кр
Mi
=
—-,
М
2
= . , ,» ,
Y21
1 +
Y12/P
с
__
+
P/Y12
-
Мз
кр
+ е
2
(1
1/угз)
P
~
1+Y21/Y23
rj
е
З .-г-
М
а
-Мз
К
р
М
2
= , М
3
= -г-, ;—-.
Y32
1
+
Y23/Y21
М-
= 0
6з
V'
2
Y32
P
М'=
2
i=Mj.
—М
3кр
,
М, = —, Mi=-=-,
Y21
Y21
М
3
=
М3
= О,
4
кр
144
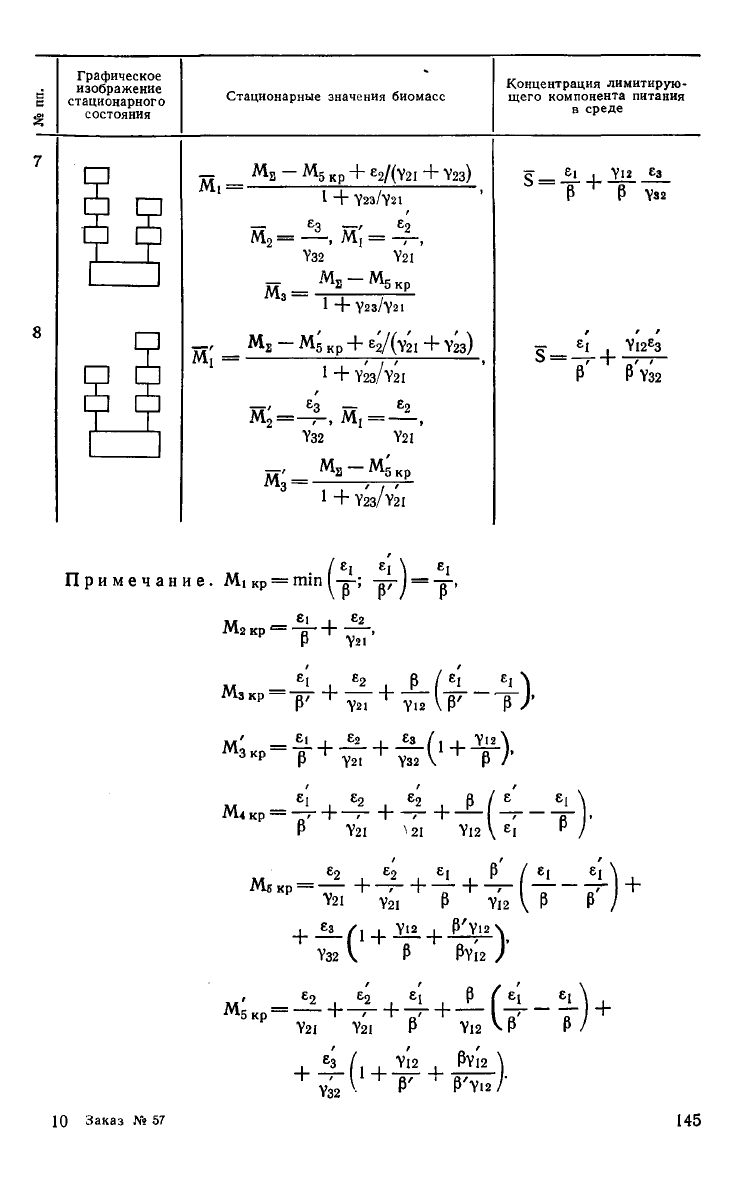
Графическое
изображение
стационарного
состояния
Стационарные значения биомасс
Концентрация лимитирую-
щего
компонента питания
в
среде
М
а
— М
5 кр
+
£
2
/(Y2l
+
Y23)
:—I ;
1
+
Y23/Y21
Б
ч
, Ео
М
2
=—,
М,
= -?-,
Y32
Y21
_ M
S
-M
5KO
-
^Е
1
+Y23/Y21
-
М
5
кр
;—г—,
1
+ Y23/Y2I
М
2
=
——,
М,= ,
Y32
Y21
—,_
—
М
д
кр
1
+ Y23/Y2I
о-_
е
' 1. Yia Ез
Т
+
Р Y32
О
==
г~ "Т" ~~,—~,
Р
PY32
Примечание. Mi
K
p
= min -=-;
\
P
..
ei . 62
M
2
кр
=
-R-
+ —,
"ог)
=
"о">
e
l
.
e
2 .
e
2 . P /
e
P'
Y21 \2I
Yi2\eJ
D
O2
&2 *^I P
^21
Y2I P Yl2
Y32
V P
ч
м,
•5
кр
1
itl
Y2i Y2I P' Yl2 W P
Y32
10 Заказ
№ 57
145

Альтернативное стационарное состояние, когда в системе при-
сутствует
только второй вид продуцентов Mi, при выбранном
знаке
неравенства (5.6) оказывается
всегда
неустойчивым.
Таким
образом, неравенство (5.6) на коэффициенты е/р опре-
деляет, какая из
двух
конкурирующих трофических цепочек полу-
чит преимущественное развитие, в частности, при выбранном
соотношении
коэффициентов первой развивается левая пищевая
цепь.
Структура 3. При увеличении запаса лимитирующего биогена
в
системе выше М
2К
р
структура
2 становится неустойчивой и по-
является
структура
3. Это стационарное состояние положительно
определено при Ms > М
2кр
. Корни характеристического уравнения
системы (5.4) имеют вид
h = —е
3
+
Y32M2;
%2
= —ei + P S,
(5.8>
Д.з=—
Е2!
Я4=—ез,
а
ЯБ,6
определяются из квадратного уравнения
2
(
4
0 (5.9>
и
всегда
имеют Re Я < 0.
Все корни (5.8) имеют отрицательные Re
К,
т. е. состояние 3'
устойчиво, если
М
2
кр<М
2
<
ггип(Мзк
Р
,
Мзкр).
(5.10>
Легко видеть, что Мзкр <
Мз
кр
при выполнении следующего нера-
венства для коэффициентов модели:
Как
будет
видно из дальнейшего изложения, неравенство
(5.11)
будет
определять характер развития трофической структуры эко-
системы при дальнейшем увеличении параметра M
s
.
Биологический
смысл неравенства
(5.11)
следующий. Разность
количественно характеризует конкурентное преиму-
щество, которое имеет продуцент Mi в отсутствие консументов.
Величина (ез/732)
(Y12/P)
отражает
«давление»
цепочки консумен-
тов М
2
— М
3
на продуцент Mi, ослабляющее конкурентные воз-
можности этого вида. Если
«давление»
консументов достаточно
велико,
в систему может заселиться второй вид продуцентов Mi,
в
противном
случае
вид Mi остается конкурентным доминантой
даже
при наличии выедания консументами.
Прежде чем перейти к дальнейшему рассмотрению модели от-
метим, что стационарное состояние, симметричное
структуре
3,
т. е. Mi — M2, при выполнении неравенства
(5.11)
всегда
не-
устойчиво.
146

Структура
4.
Рассмотрим стационарное состояние,
в
котором
«левая»
трофическая цепь, состоящая
из
видов
Mi, M
2
, М
3
, пол-
ностью вытесняет представителей второй трофической цепи.
Это
стационарное состояние
существует
при
М
2
>Мз
К
р. Характери-
стическое уравнение системы
(5.4) в
этом
случае
имеет корни
h
=
— ei
+
PS,
X
2
=
— е
2
,
(5.12)
три
других
корня удовлетворяют уравнению
а
а
Х
3
+ а
1
Х
2
+
а
2
Х
+ а
3
= 0,
(5.13)
где
а
о
=1,
ai =
pMi,
аг
==
Y23Y32M2M3
+ V21 (р + Y12)
М1М2,
аз
= Mi
M2M3PY32
(Y21
+ Y23).
Условие устойчивости Раусса—Гурвица
[60] для
уравнения
(5.13)
сводится
к
неравенству
Y12Y23
—
Y21Y32
> 0.
(5-14)
которое всегда выполняется
[см.
(5.3)].
Поэтому состояние
4
устойчиво
при
единственном условии
ReXi<0, которое выполняется, если выполнено неравенство
(5.11).
Таким
образом, существование цепочки видов
Mi —
М2
—
М3
при
условии
(5.11)
оказывается устойчивым
всюду
начиная
с
Мз =
Мзкр. Этот вариант сукцессии системы представлен
на
рис.
5.11
а.
Структура
5.
Положительно определена
при M
s
>
М
ЗКР
.
Три
корня
характеристического уравнения имеют
вид
h
=
—
ез
+
Y32M2,
Я-2
=
—е
2
-f
Y2iMi,
(5.15)
Я.З
= —ЕЗ,
а
три
других
корня удовлетворяют кубическому уравнению
a
0
X
i
+ a
l
X
s
+
a
2
X
+ a
3
= 0,
(5.16)
где
a
o
=l,
a
I
=
pMi
+
p'Mi,
а
2
=
(р+
712)721
MiМ
2
,
а
3
=
М1М2М1Р
Yi2Y2i.
10*
147
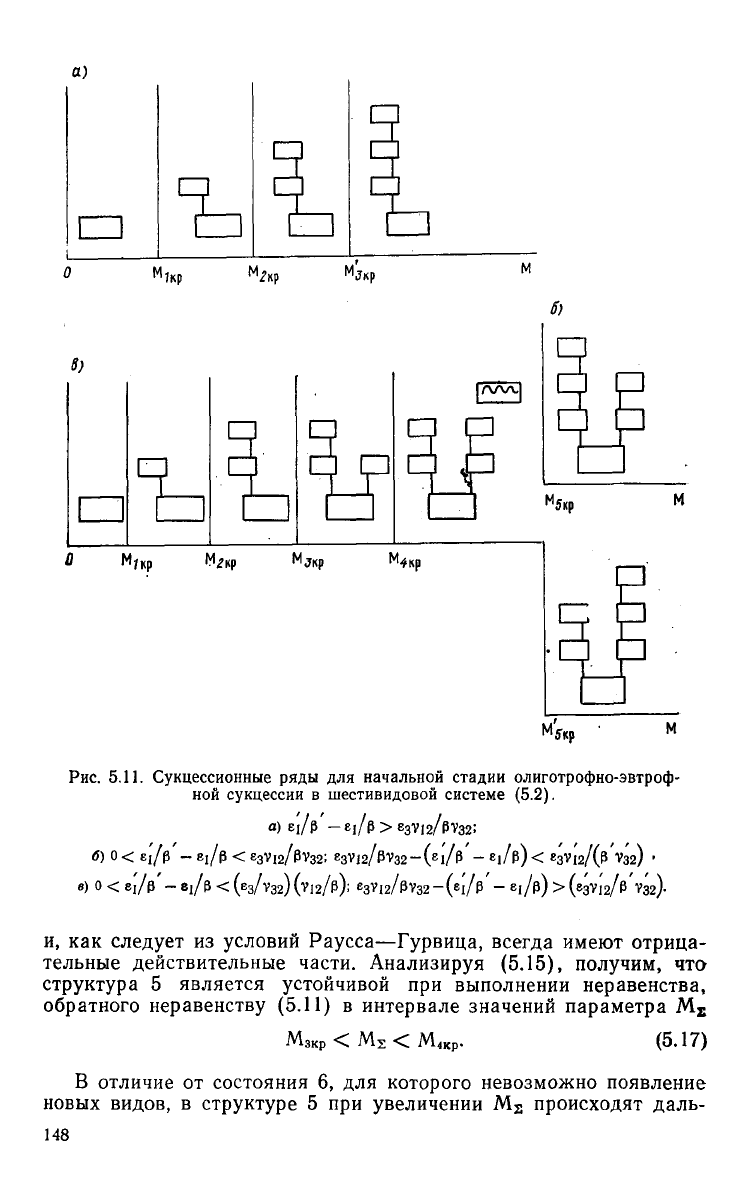
а)
ъ
1,
б)
8)
м
»кр
1JKP
Рис.
5.11. Сукцессионные ряды для начальной стадии олиготрофно-эвтроф-
ной
сукцессии в шестивидовои системе (5.2).
a) ei/p
-ei/p>e
3
Vl2/pV32:
в) о <
ei/p'-
ej/p <
(E
3
/V
3
2)(VI2/P);
)
P'-
8,/p)
>
и,
как
следует
из условий Раусса—Гурвица, всегда имеют отрица-
тельные действительные части. Анализируя (5.15), получим, что
структура
5 является устойчивой при выполнении неравенства,
обратного неравенству (5.11) в интервале значений параметра М
г
< M
s
< М
4Кр-
(5.17)
В отличие от состояния 6, для которого невозможно появление
новых видов, в
структуре
5 при увеличении M
s
происходят даль-
не

нейшие
сукцессионные изменения, выражающиеся в появлении
новых консументов, как показано на рис.
5.116,
в.
Структура 6. Следующим за структурой 5 при возрастании M
s
в
системе возникает сообщество, состоящее из
двух
цепочек, вклю-
чающих продуценты и консументы первого порядка.
Трофическая
структура
6
существует
при M
s
> М
4к
р^ корни
характеристического уравнения системы (5.4) имеют вид
h = —
ез
+
Y32M2,
%2
= — е
3
+ Y32 Мг,
(5.18)
а четыре остальных корня определяются из уравнения
V + a,V +
a
2
l
2
+ а
3
Х + а
4
= 0,
(5.19)
где
сц = pMi + р'Мь а
2
=
M1M2Y21
(p + Y12) +
M'IM^I
(р' + 712),
а
3
= MiMi
(PY12Y21M2
+ Р
Yi2Y2i
M
2).
«4 =
Y21Y21M1M2M1M2
(YI2Y12
4- PY12 + Р Via)-
Для уравнения
(5.19)
возможно нарушение условий устойчиво-
сти Раусса—Гурвица, в этом
случае
система (5.2) не
будет
иметь
ни
одного устойчивого стационарного состояния. Поскольку общая
масса M
s
лимитирующего элемента ограничена, биомассы видов
должны совершать колебания. Подробно возможные режимы ав-
токолебаний в четырехвидовом биоценозе рассмотрены в п. 5.5.
В области значений коэффициентов, где критерий Раусса —
Гурвица для уравнения
(5.19)
выполняется, стационарное состоя-
ние
является устойчивым при ReA,i,2<0, откуда получаем интер-
вал значений
М
4кр
< М
2
< min(M
5
Kp,
MSKP).
(5.20>
Нетрудно показать, что Ms
K
p < Ms
K
p. если выполняется нера-
венство
е
з
VI2 . /j-
Структура 7. Эта трофическая
структура
возникает в экоси-
стеме при М
2
> М
5к
р и выполнении неравенства (5.21). Характе-
ристическое уравнение системы (5.4) имеет вид
X
5
+ а,Я.
4
+ а
2
Я.
3
+ a
3
V -f a
t
k +
a
s = 0, (5.22>
на

где
Y23Y32M2M3,
PY21Y32M1M3
+
+
P
Y12Y21M1M1
+
PY12Y21M1M1
Мг,
a
4
= (p +
Y12JY21Y23Y32M1M2M2M3
+
+
(Y12Y12
+ PY12 + P Y12)
Y2lY2lM?M2,
^5 =
(Y2!Ml
+
Y23M2)
PY12Y21Y32M1M1M2M3.
Для того чтобы состояние 7 было устойчиво, необходимо и доста-
точно выполнения условий Раусса — Гурвица
a,a
2
— а
3
> О,
(5.23)
а
3
(а
1
а
2
— а
3
) — а, (а
{
а
А
— а
5
) > 0.
Можно
показать, что при M
s
, значительно превышающем Мбкр,
неравенства
(5.23)
выполняются. При M
s
, немного превышающем
Мбкр,
возможно нарушение условий Раусса — Гурвица, в этом слу-
чае в системе возникают автоколебания.
Структура
8. Если выполняется неравенство на коэффициенты
системы, противоположное неравенству (5.21), то вместо струк-
туры
7 при M
s
> М
5 кр
появляется
структура
8 (см. рис. 5.11 в).
Поскольку
структурами 7 и 8 исчерпываются возможности мо-
дельной экосистемы (5.2), дальнейшей сукцессии при возрастании
запаса биогенного элемента M
s
не произойдет. Для модельной
экосистемы
большей размерности аналогичным образом можно
проследить развитие более сложного сообщества.
Таким
образом, анализ модели экосистемы (5.2)
дает
следую-
щие
результаты:
при
фиксированном значении запаса биогенного элемента М^
лишь
одно стационарное состояние системы оказывается положи-
тельно определенным и устойчивым;
каждая видовая
структура
сообщества
существует
и устойчива
в
определенном интервале значений М
2
, причем верхняя граница
устойчивости г-й структуры совпадает с нижней границей устой-
чивости более сложной (t+ 1)-й структуры;
последовательная смена устойчивых стационарных состояний
при
возрастании биогенной нагрузки M
s
образует сукцессионный
ряд,
имитирующий развитие системы при эвтрофикации. Сукцес-
сионные
ряды для системы (5.2) представлены графически на
рис.
5.11;
в
системе возможны несколько вариантов сукцессии в зависи-
мости от неравенств, связывающих коэффициенты. Неравенства
на
коэффициенты представляют собой соотношения
между
мини-
150