Zelditch M.L. (и др.) Geometric Morphometrics for Biologists: a primer
Подождите немного. Документ загружается.

chap-13 4/6/2004 17: 28 page 348
348 GEOMETRIC MORPHOMETRICS FOR BIOLOGISTS
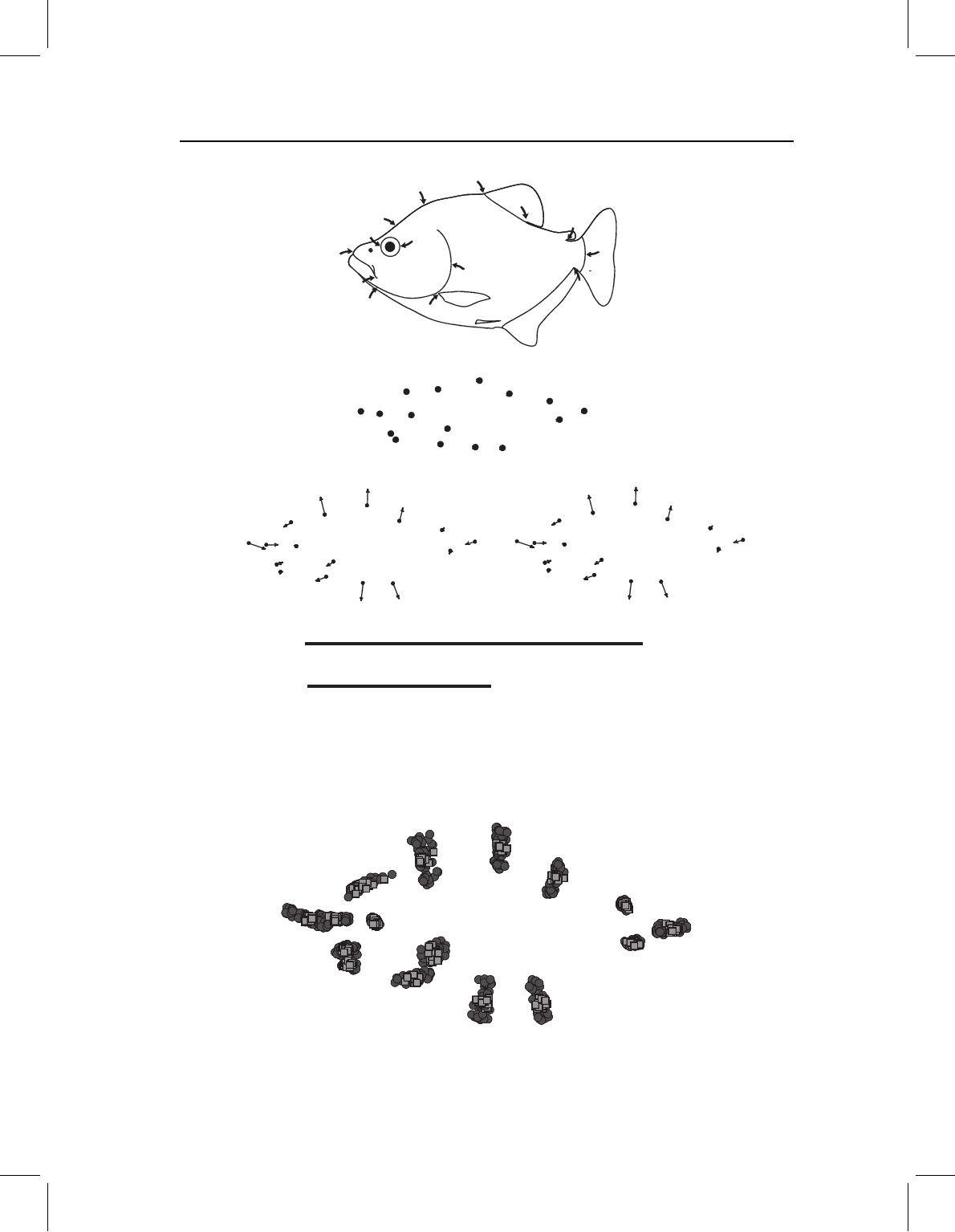
log (head length)
log (body length)
log (head depth)
log (body length)
Figure 13.16 A complex change in three parameters: early morphogenesis; modification of later
morphogenesis; and change in developmental rate and timing (due to increased body size). The
ancestor ontogeny is indicated by circles, the descendent ontogeny by squares.
Table 13.5 Allometric coefficients b and k and their standard errors for
S. gouldingi and S. manueli (see Figure 13.1 for the definition of the variables)
Variable S. gouldingi S. manueli
b (std error) k (std error) b (std error) k (std error)
v2 −0.94 (0.13) 0.81 (0.03) −1.81 (0.09) 1.03 (0.02)
v3 −0.61 (0.05) 0.89 (0.01) −0.47 (0.06) 0.87 (0.01)
v4 −1.16 (0.07) 0.85 (0.01) −1.27 (0.07) 0.90 (0.02)
v5 −2.64 (0.13) 1.23 (0.03) −1.45 (0.11) 0.96 (0.03)
v6 −1.40 (0.04) 1.04 (0.01) −1.14 (0.05) 1.00 (0.01)
v7 −1.51 (0.06) 1.00 (0.01) −1.80 (0.05) 1.09 (0.01)
v8 −1.38 (0.09) 0.93 (0.02) −1.00 (0.11) 0.85 (0.03)
v9 −1.97 (0.08) 1.10 (0.02) −1.81 (0.08) 1.06 (0.02)
v10 −1.76 (0.04) 1.22 (0.01) −1.56 (0.04) 1.17 (0.01)
v11 −1.93 (0.04) 1.20 (0.01) −1.59 (0.04) 1.13 (0.01)
v12 −1.60 (0.04) 1.14 (0.01) −1.32 (0.04) 1.08 (0.01)
v13 −2.38 (0.09) 1.19 (0.02) −2.27 (0.07) 1.15 (0.02)
v14 −2.23 (0.07) 1.12 (0.02) −2.10 (0.09) 1.08 (0.02)
v15 −1.68 (0.04) 1.21 (0.01) −1.41 (0.05) 1.15 (0.01)
v16 −1.78 (0.04) 1.23 (0.01) −1.52 (0.05) 1.17 (0.01)
v17 −1.55 (0.03) 1.17 (0.01) −1.24 (0.05) 1.10 (0.01)
v18 −2.57 (0.08) 1.17 (0.02) −1.98 (0.10) 1.05 (0.02)
v19 −1.94 (0.05) 1.09 (0.01) −1.62 (0.06) 1.02 (0.02)
v20 −1.69 (0.05) 1.10 (0.01) −1.40 (0.04) 1.04 (0.01)
v21 −1.99 (0.04) 1.23 (0.01) −1.74 (0.05) 1.17 (0.01)
v22 −1.83 (0.04) 1.22 (0.01) −1.56 (0.05) 1.16 (0.01)
v23 −1.47 (0.04) 1.07 (0.01) −1.30 (0.05) 1.03 (0.01)
v24 −1.67 (0.08) 0.94 (0.02) −2.70 (0.13) 1.13 (0.03)
v25 −2.69 (0.07) 1.12 (0.01) −2.57 (0.06) 1.09 (0.01)
v26 −1.56 (0.03) 1.13 (0.01) −1.37 (0.04) 1.09 (0.01)
v27 −1.82 (0.11) 0.90 (0.02) −3.41 (0.16) 1.23 (0.04)
v28 −1.23 (0.09) 0.81 (0.02) −1.89 (0.10) 0.97 (0.02)
v29 −1.16 (0.08) 0.75 (0.02) −1.50 (0.08) 0.82 (0.02)
v30 −0.64 (0.04) 0.90 (0.01) −0.51 (0.04) 0.89 (0.01)
chap-13 4/6/2004 17: 28 page 349
THE RELATIONSHIP BETWEEN ONTOGENY AND PHYLOGENY 349
(B)
(A)
1.12
.81
1.14
1.23
.75
.90
1.04
1.22
1.21
1.23
1.13
1.10
.89
1.00
1.23
1.00
.81
.85
.93
1.19
1.17
1.07
.90
.94
1.09
1.12
1.10
1.22
1.17
1.20
1.10
1.08
1.17
.8 2
1.13
.88
1.00
1.16
1. 16
1.18
1.09
1.04
1.09
.96
1.03
.97
.89
.87
.85
1.16
1.05
1.03
1.24
1.14
1.18
1.11
1.00
1.07
1.08
1.02
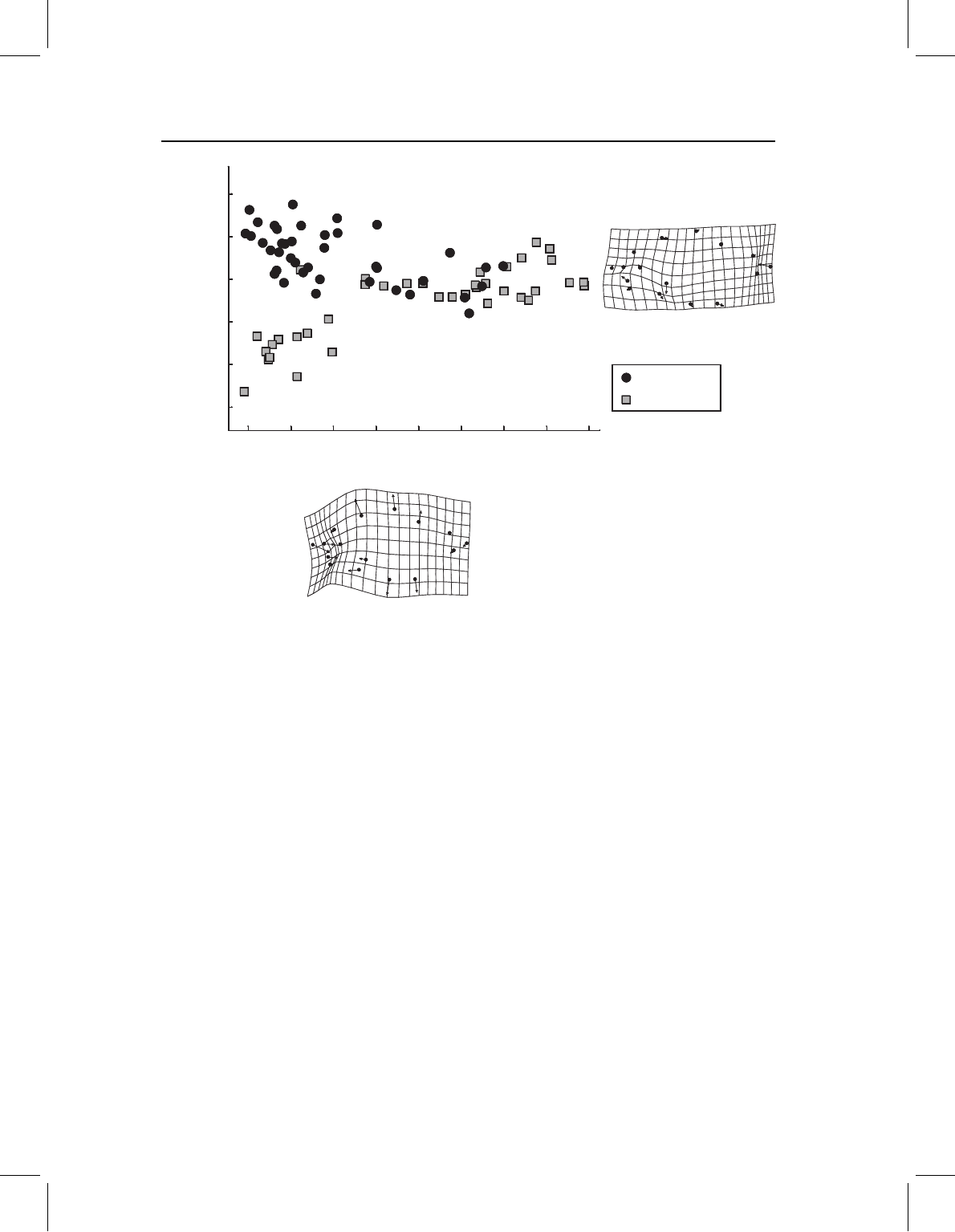
Figure 13.17 Allometric coefficients of S. gouldingi and S. manueli.
differ (Figure 13.17). Statistically, they differ in k for nearly all variables; the exceptions
are two in the head (v3 and v30), two anterior postcranial measurement (v9 and v13)
and the depth measurement of the caudal peduncle (v25). The angle between the vectors is
small (6.27
◦
), but is nonetheless significantly greater than 0.0
◦
(p < 0.05). Considering that
an angle of only just over 12.74
◦
indicates greater dissimilarity than expected by chance,
6.27
◦
does not seem so modest. There is some evidence, albeit slight, for an interaction
between modifications of early and late ontogeny that reduce disparity over ontogeny;
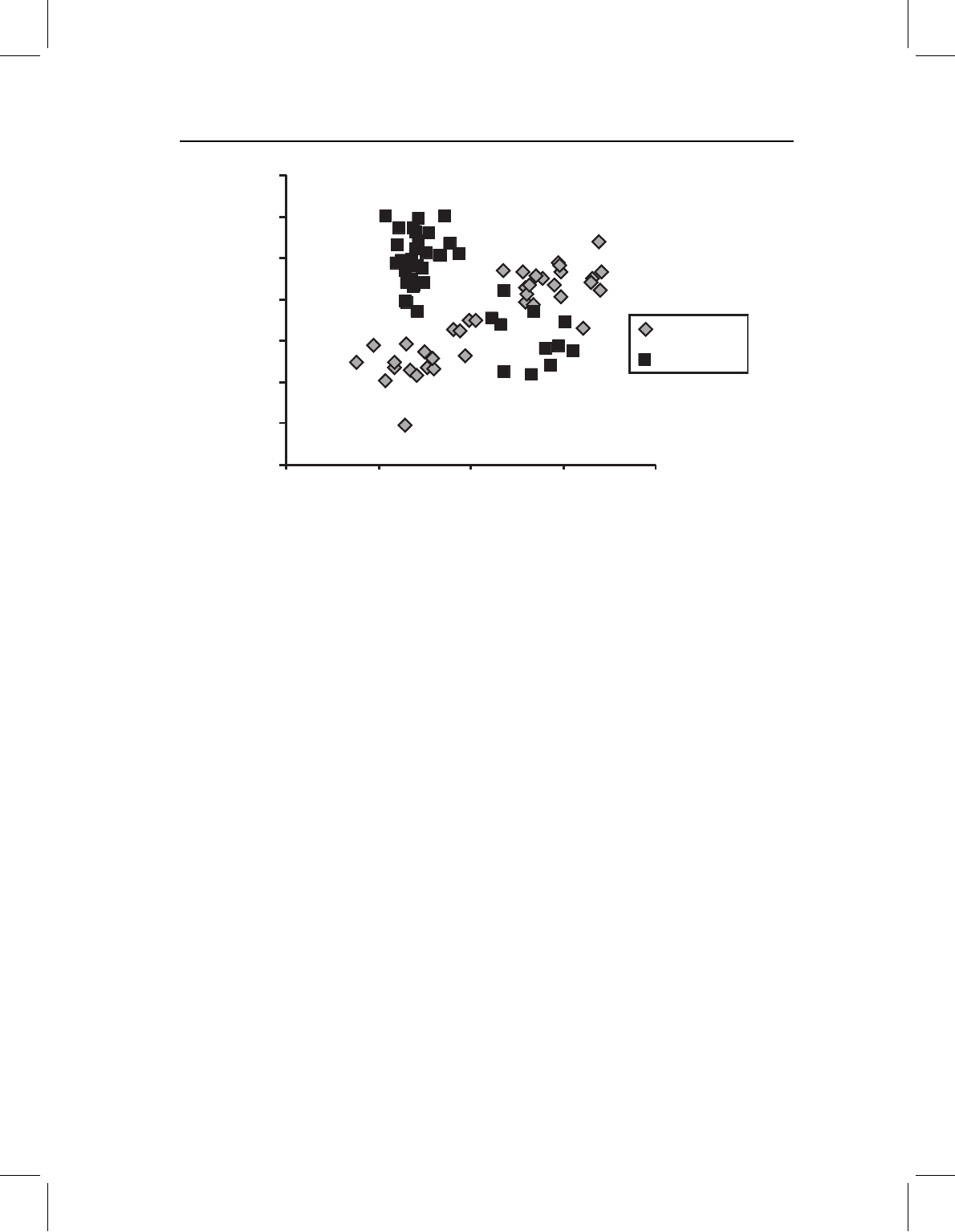
this pattern of counterbalancing modifications is suggested by the plot of PC2 on PC1
(Figure 13.18). Scores on PC1 increase with age, it also looks as though they are more
differentiated on PC2 earlier rather than later, and that they gradually reach a more similar
form. Yet the scale of PC2 is tremendously exaggerated in the plot; that axis accounts for
only 0.4% of the variance (PC1 accounts for 98.9%). So it is not clear whether this really
is a case of counterbalancing, an issue we will return to in the analysis of these species
using a geometric approach.
chap-13 4/6/2004 17: 28 page 350
350 GEOMETRIC MORPHOMETRICS FOR BIOLOGISTS
⫺0.8
⫺0.6
⫺0.4
0
⫺10 0 5 10
PC1
0.6
0.4
0.2
⫺0.2
⫺5
PC2
S. gouldingi
S. manueli
Figure 13.18 PCA of the pooled ontogenetic data of S. gouldingi and S. manueli. The scale of
the Y-axis (PC2) is greatly exaggerated to make the separation between species more visible; older
specimens are to the right of the plot. The two species appear to differ, albeit subtly, in shape at the
outset of the measured stage and in their ontogenies. The modification of juvenile growth appears to
counterbalance the change in larval morphogenesis such that adults are more similar than juveniles.
Exploring evolutionary patterns of evolving ontogenies: geometric
morphometric analyses
Geometric studies of ontogenetic allometry are formally similar to traditional morphome-
tric ones, but there are important differences in the meanings of the parameters. Revisiting
the regression of geometric shape on size (covered in Chapter 10), the general bivariate
linear regression model is:
Y
i
= mX
i
+b
i
+ε
i
(13.4)
where Y is a shape variable, X is body size (measured by centroid size), b is the Y-intercept
and ε is the error term. Because we invariably analyze shape data multivariately, the model
we actually use is:
{Y
1
, Y
2
, Y
3
, ... Y
P
}={m
1
, m
2
, m
3
, ...m
P
}X +{b
1
, b
2
, b
3
, ...b
P
}+{ε
1
, ε
2
, ε
3
, ...ε
P
}
(13.5)
where {Y
1
, Y
2
, Y
3
...Y
P
} is the vector of shape variables (i.e. landmark coordinates
obtained by GLS, or partial warp scores including the uniform component), X is cen-
troid size, and {m
1
, m
2
, m
3
, ...m
P
}, {b
1
, b
2
, b
3
, ...b
P
} and {ε
1
, ε
2
, ε
3
, ...ε
P
} are vectors
of slope coefficients, intercepts and residuals, respectively.
One consequence of analyzing allometry using geometric data instead of traditional data
is that the allometric coefficients are no longer meaningful in terms of a specific growth

chap-13 4/6/2004 17: 28 page 351
THE RELATIONSHIP BETWEEN ONTOGENY AND PHYLOGENY 351
model – either a power law (Equation 13.1) or a sigmoidal curve of size or shape relative
to time. Instead, they indicate that shape changes with size. The underlying causes of those
changes are the same ones that account for allometric coefficients of traditional measure-
ments, but we cannot treat the coefficients as if they were rates of growth of individually
meaningful variables, nor can we use them to estimate time-lags between growth curves of
organs. Unlike the allometric coefficients of traditional measurements, those of geometric
shape variables have no individual biological meaning. For that reason, comparisons of
ontogenetic allometries using geometric variables are rarely (if ever) done bivariately. We
would not plot one shape variable at a time on size, except to check for linearity. In a geo-
metric analysis we are not comparing coefficients measurement-by-measurement; rather,
we are comparing whole sets of coefficients describing the ontogeny of an entire landmark
configuration. However, the difference in the meanings of the coefficients does not impede
our ability to recognize the patterns discussed above in the context of traditional measure-
ments. None of these patterns were defined in terms of particular coefficients; hence they
are not functions of a particular measurement scheme. We can thus examine the evidence
for them in geometric as well as traditional data.
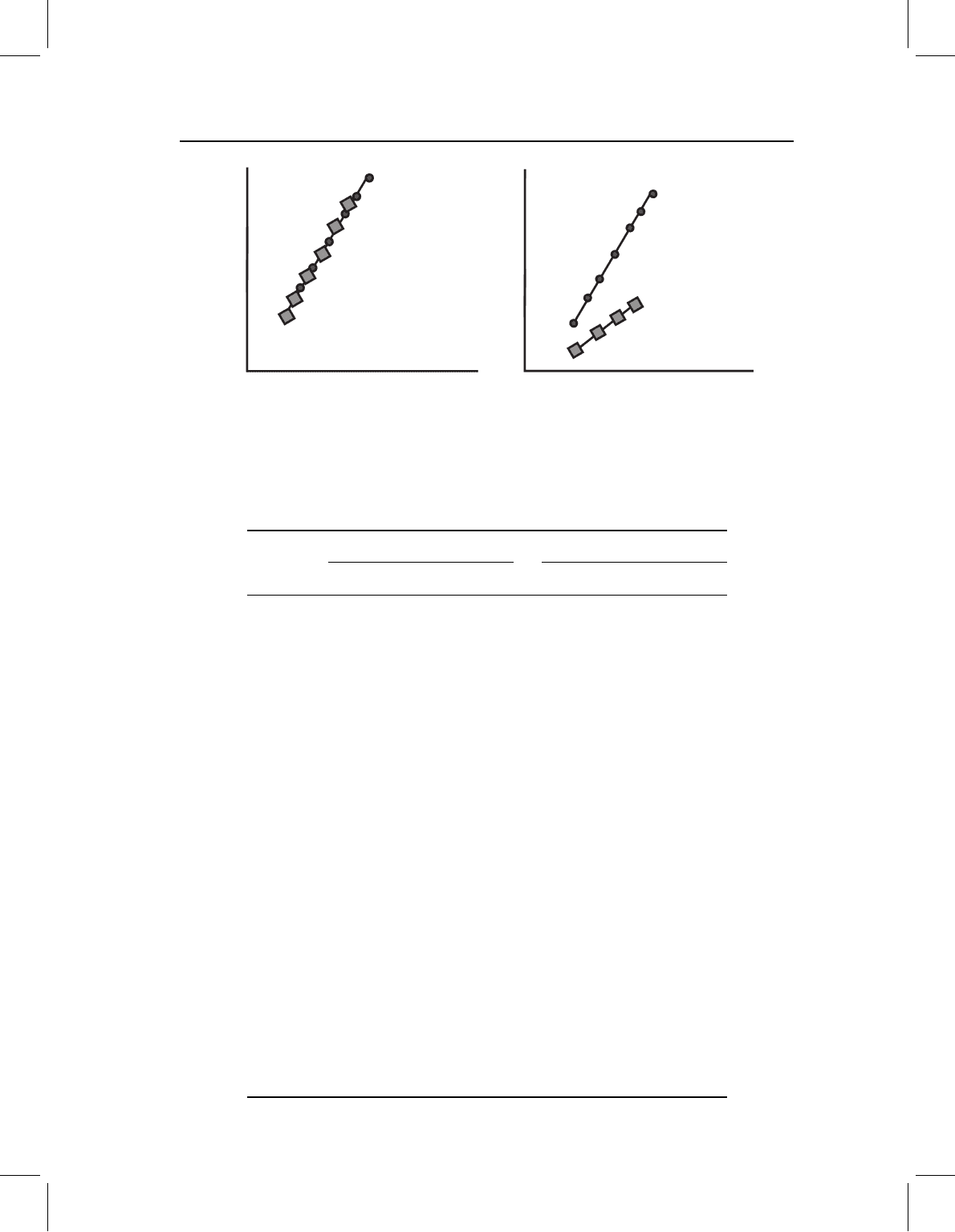
Channeling
To depict the pattern of channeling, we can consider a hypothetical case in which species
have the same shape at the outset of development and follow the same ontogeny of shape,
but differ in the overall rate or timing of development. Graphical evidence of channeling is
shown in Figure 13.19, where we see that the coordinates of the juveniles of the two species
are the same (Figure 13.19A), as are the two ontogenies of shape (Figure 13.19B), but the
trajectories differ in length (Figure 13.19C). Perhaps the most compelling visual evidence
is shown in Figure 13.20 – the descendant adult morphology lies at a subadult position on
the ancestral ontogeny. We can see the coordinates for the descendant’s landmarks in an
intermediate position along the ancestral ontogeny.
The graphical evidence is corroborated by statistical analysis. In a statistical test of
channeling, we would not expect to find a significant difference between the two ontogenies
of shape or in the shape at the youngest comparable phase but we would anticipate a
difference in the length of the ontogenetic vector (the parameter that measures the total
amount of change undergone in each ontogeny over the observed phase). Carrying out these
tests for the hypothetical species depicted in Figure 13.18, we would measure the similarity
between ontogenies of shape by the angle between the (normalized) vector of allometric
coefficients {m
1
, m
2
, m
3
, ...m
P
}. We find no significant difference between them; the
tiny angle of only 1.8
◦
is not significant compared to the ranges that can be obtained by
resampling within them (4.2
◦
, 7.5
◦
). We also find no significant interspecific difference
between shapes at the outset of the measured phase of development using Goodall’s F-test
(p > 0.999); and the magnitude of the difference between them is a Procrustes distance of
0.0. We do, however, find a significant difference in the length of their ontogenetic vectors
as estimated by the Procrustes distance between youngest and oldest comparable stages; for
the ancestral species that distance is 0.1999 (0.1961–0.2030), whereas for the descendant
it is only 0.109 (0.1055–0.1129). Thus, the descendant’s ontogeny is a truncated version
of the ancestor’s.
chap-13 4/6/2004 17: 28 page 352
352 GEOMETRIC MORPHOMETRICS FOR BIOLOGISTS
4
1
2
3
5
6
7
8
11
12
13
14
15
16
(A)
(B)
(C)
Figure 13.19 Channeling, depicted geometrically (see Figure 13.1 for the same pattern depicted
for traditional measurement data). (A) Superimposed coordinates of juvenile shapes; (B) ontogenies
of shape; (C) lengths of ontogenetic vectors of shape. The two species have the same shape at the
outset of the measured phase, follow the same ontogeny of shape, but differ in the length of their
ontogenetic vectors; the descendant has a truncated version of the ancestral ontogeny.
Figure 13.20 Superimposed coordinates for showing the ontogenetic transformation of ancestral
shape (black circles) and the descendant adult shape (gray squares). The descendant adult shape is
at an intermediate position along the ancestral ontogeny.

chap-13 4/6/2004 17: 28 page 353
THE RELATIONSHIP BETWEEN ONTOGENY AND PHYLOGENY 353
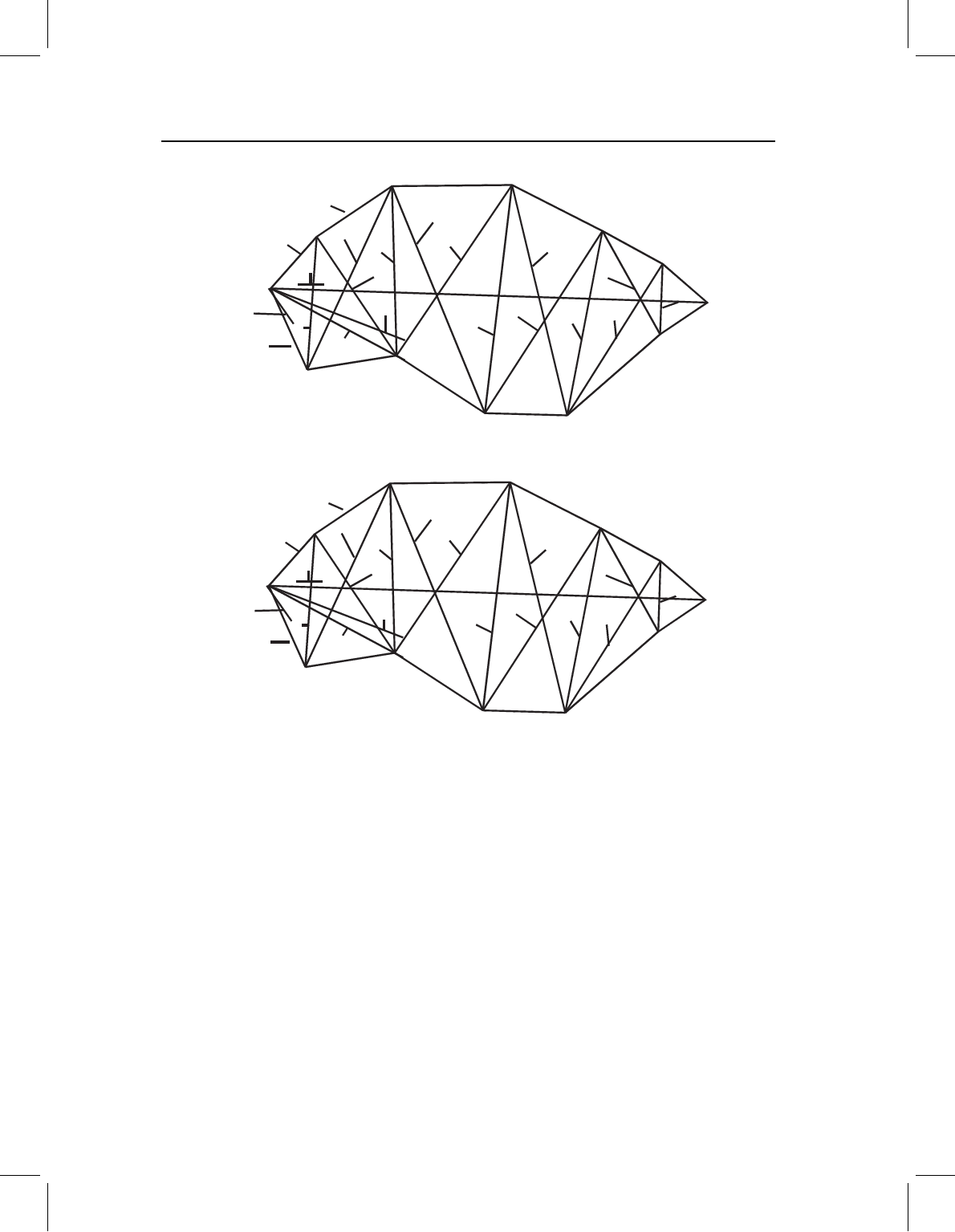
Changes in the ontogenetic trajectory
Changes confined to early morphogenesis
If changes occur solely in early morphogenesis, we would expect that species would be
shaped differently at the youngest comparable stage (Figure 13.21A) but would subse-
quently follow the same ontogeny of form (Figure 13.21B), and to the same extent (Figure
13.21C). To test the hypothesis that only early development is labile, we can show that
there is a significant difference in shape at the outset of the measured phase, but that
any differences in later development are neither significant nor large. For the hypothet-
ical species shown in Figure 13.21, the difference between their shapes at the transition
(B)
(C)
4
1
2
3
5
6
7
8
11
12
13
14
15
16
(A)
Figure 13.21 Change confined to early morphogenesis, depicted geometrically (see Figure 13.2
for the same pattern depicted for traditional measurement data). (A) Superimposed coordinates of
juvenile shapes; (B) ontogenies of shape; (C) lengths of ontogenetic vectors of shape. The two species
differ in shape at the outset of the measured phase, but subsequently follow the same ontogeny of
shape and do not differ in the length of their ontogenetic vectors.

chap-13 4/6/2004 17: 28 page 354
354 GEOMETRIC MORPHOMETRICS FOR BIOLOGISTS
from larval to juvenile phases is highly significant (p < 0.0001) and the Procrustes distance
between their means is large: 0.1247 (0.1196–0.1317). The contrast may seem particularly
striking when we look at the superimposed coordinates and find so little overlap between
species in several of them (Figure 13.21A). As anticipated, there is no significant difference
in their ontogenies of form; the angle between the two vectors is a tiny 1.9
◦
(compared to
the within-species angles of 4.0
◦
and 3.7
◦
). Also as anticipated, the lengths of the ontoge-
netic vectors are the same for both species: 0.1999 (0.1961–0.2030) for the ancestor, and
0.2040 (0.1978–0.2095) for the descendant.
Changes in the spatiotemporal organization of development from the outset
to the end of allometric growth
Should species differ in the spatiotemporal organization of morphogenesis, their ontoge-
netic vectors of shape will differ. It says nothing about shape at the youngest comparable
stage, so the only expectation is that the ontogenies of shape will differ. Rather than dis-
cussing this simple case any further, we will consider the more complex hypotheses that
include it.
Changes in the spatiotemporal organization of development, confined to
late development
Should all change be confined to late morphogenesis, we would expect to see no difference
between the species at the youngest comparable developmental stage (Figure 13.22A),
and a visible difference in their ontogenies of shape (Figure 13.22B). This hypothesis says
nothing about the length of the trajectory of late development; however, changes in length
are due to differences in rate/timing and not to differences in morphogenesis, so we have
drawn trajectories of the same length (Figure 13.22C). For this hypothetical case, juvenile
shapes do not differ (p > 0.999, Procrustes distance =0.0) and neither do the lengths of
the ontogenetic vector: that of the ancestor is 0.1999 (0.1961–0.2030) and that of the
descendant is 0.2011 (0.1977–0.2046). However, as anticipated, the ontogenies of shape
differ significantly; the angle between the two vectors is 32
◦
compared to the range of
within-species angles that can be obtained by resampling (3.7
◦
for both).
Complex changes in multiple parameters and stages
It is difficult to construct hypothetical ontogenies to demonstrate interactions among mul-
tiple parameters, because that requires modeling perturbations of the ontogeny of entire
landmark configurations. Also, the most interesting distinctions among the possibilities
lies in their conflicting predictions about the ontogenetic dynamics of disparity. In par-
ticular, we would wish to distinguish between amplification (which predicts increasing
disparity over ontogenetic time) and counterbalancing (which predicts decreasing dispar-
ity over ontogenetic time). The contrasting patterns are not self-evident in the regression
parameters; distinguishing them requires measuring disparity. Rather than considering
another hypothetical case we will return to the analysis of an empirical case, S. gouldingi
and S. manueli, because our earlier analysis suggested that these species might exemplify
counterbalancing.

chap-13 4/6/2004 17: 28 page 355
THE RELATIONSHIP BETWEEN ONTOGENY AND PHYLOGENY 355
4
1
2
3
5
6
7
8
11
12
13
14
15
16
(A)
(B)
(C)
Figure 13.22 Change in the spatiotemporal pattern of development confined to late development,
depicted geometrically. (A) Superimposed coordinates of juvenile shapes; (B) ontogenies of shape;
(C) lengths of ontogenetic vectors of shape. The two species have the same shape at the outset of
the measured phase, but subsequently follow different ontogenies of shape; they do not differ in the
length of their ontogenetic vectors.
An empirical case: comparing ontogenies of S. gouldingi and S. manueli
Comparing these two species using geometric methods provides compelling graphical
evidence that the species differ in morphology at the transition from larval to juvenile
development (Figure 13.23A), in juvenile morphogenesis (Figure 13.23B), and in length of
their ontogenetic trajectories (Figure 13.23C). The difference in shape at the transition from
larval to juvenile development is statistically significant (p< 0.0001). Moreover, this differ-
ence is not just significant, it is large; the Procrustes distance between mean shapes at that
stage is 0.080 (0.077–0.084). Ontogenies of shape are visibly different and the difference
is statistically significant; the angle between their ontogenetic vectors is 34.9
◦
(compared
to the within-species ranges of 11.0
◦
and 16.6
◦
for S. gouldingi and S. manueli, respec-
tively). The ontogenetic trajectories also differ significantly in length; that of S. gouldingi

chap-13 4/6/2004 17: 28 page 356
356 GEOMETRIC MORPHOMETRICS FOR BIOLOGISTS
4
1
2
3
5
6
7
8
11
12
13
14
15
16
(A)
(B)
(C)
Figure 13.23 Ontogenies of S. gouldingi and S. manueli, depicted geometrically. (A) Superimposed
coordinates of juvenile shapes; (B) ontogenies of shape; (C) lengths of ontogenetic vectors of shape.
The two species differ in shape at the outset of the measured phase (the transition from larval to
juvenile phases), subsequently follow different ontogenies of shape and differ in the length of their
ontogenetic vectors.
is 0.2095 (0.2065–0.2128) and that of S. manueli is 0.1864 (0.01823–0.1906). Therefore,
these two species differ in all three parameters. However, the adults are far less different
than the young juveniles. The Procrustes distance between mean adult shapes is 0.051
(0.047–0.054) – a substantial drop from 0.080. That decrease can be seen in the analysis
of the pooled ontogenies of shape by PCA (Figure 13.24); with increasing age, scores on
PC1 increase, and the two species more closely resemble each other. We can now clearly
see the pattern hinted at in the analysis of traditional morphometric data. The evidence
based on geometric data is stronger, because now there are two distinct eigenvalues, and
PC2 explains 9.5% rather than 0.4% of the variance. Taken together, these results all
demonstrate that the modifications of juvenile development counterbalance those of larval
development, thereby stabilizing adult morphology.
chap-13 4/6/2004 17: 28 page 357
THE RELATIONSHIP BETWEEN ONTOGENY AND PHYLOGENY 357
0
0
PC1
PC2
0.04
0.02
⫺0.02
⫺0.04
⫺0.06
⫺0.06 ⫺0.04 ⫺0.02 0.02 0.04 0.06 0.08 0.1
S. gouldingi
S. manueli
Figure 13.24 PCA of the pooled ontogenetic data of S. gouldingi and S. manueli. The two species
differ in shape at the outset of the measured stage (the transition from larval to juvenile phases) and
also in their ontogenies; older specimens are to the right of the plot. The modifications of juvenile
growth appear to counterbalance the change in larval morphogenesis such that the adults are more
similar than the juveniles.
Open questions
Having characterized a variety of evolutionary transformations of ontogeny, obvious
questions are:
1. Which is (are) most likely?
2. Which occur(s) most often?
To answer the first question, we can derive expectations from developmental theory or
functional morphology, or we can even turn to the empirical literature for insights into what
expectations are biologically reasonable. Developmental biology offers few general theo-
ries, but there is a general set of principles that can be useful for framing hypotheses. One
such general principle is that development is likely to be conservative when modifications
disrupt the sequence of epigenetic interactions on which later development depends. Hall
(1992) discusses the importance of epigenetic cascades, and these may resist modification
either because modifications are lethal or because the cascades are internally stabilized.
Stabilizing selection may play an important role in both cases, not only by eliminating
deviants but also by building internal mechanisms that resist such disruptions (internal