Вестхайде В.,Ригер Р.( ред.) Зоология беспозвоночных в двух томах. Том 1: от простейших до моллюсков и артропод
Подождите немного. Документ загружается.

46
Protozoa
тернах эндоплазматической сети или аппарата Голь-
джи и выводятся путем экзоцитоза в определённые
для них места на наружной стороне клеточной мем-
браны. Одинаковое строение «страменопил» и один
и тот же способ их формирования в онтогенезе оце-
нивают как синапоморфный признак, объединяющий
различные таксоны. Как другой общий признак выс-
тупают так называемые «переходные спирали», ко-
торые в качестве осмиофильных протеиновых комп-
лексов маркируют переход от кинетосом к стержням
жгутиков.
Исходным пунктом радиации были гетеротроф-
ные жгутиковые, у которых страменопили развива-
ются не на жгутике, а на заднем конце тела клетки
(Somatonemata). От них можно вывести как гетерот-
рофные, так и автотрофные таксоны.
9.3.1.
Proteromonadea
Космополитические организмы, эндобионты
пищеварительного тракта амфибий, рептилий и мле-
копитающих. Распространение осуществляется ци-
стами, которые выводятся с экскрементами. Патоген-
ное действие неизвестно. Два рода с одной (Protero-
monas) или двумя парами (Keratomorpha) гетероди-
намных жгутиков (илл. 51), расположенных на пере-
днем конце продолговатых одноядерных клеток 10-
30 мкм длиной. Утолщенное основание более длин-
ного переднего жгутика Proteromonas возле аксоне-
мы содержит пучок рыхло собранных микрофибрилл
диктиосома
ядро
митохондрия
неизвестной химической природы, который особым
соединительным комплексом связан с базальной ча-
стью «рулевого» жгутика (Schleppgeissel). Мастиго-
немы отсутствуют. Как и у хризомонад, область пе-
рехода от кинетосом к стержню жгутика имеет с пе-
реходную спираль, связанную с поверхностью един-
ственной митохондрии через лентовидный ризопласт
из микротубулярных и филаментозных элементов. На
своем протяжении ризопласт пересекает кольцевид-
ную диктиосому, а также проходит сквозь клеточное
ядро.
Другие составные части цитоскелета — мик-
ротрубочки, которые по одной или группами подсти-
лают поверхность клетки и придают ей типичную
продольную или косую исчерченносты Задний отдел
клетки Proteromonas несёт волосовидные придатки
(соматонемы). Отчётливо выраженный пелликуляр-
ный цитоскелет состоит из рядов микротрубочек; от-
сутствует, например, у хризомонад; рассматривают
как исходный признак.
*
Proteromonas
lacertae-viridis,
10-30
мкм,
в
кишеч-
ном тракте ящериц.
9.3.2.
Opalinea
Обитают в холоднокровных позвоночных,
прежде всего Anura, а также в хвостатых амфибиях
и в рыбах, очень редко в рептилиях, исключительно
как непатогенные внутренние комменсалы в задних
отделах пищеварительного тракта. Распространение
посредством цист.
Около 400 видов, собранных лишь в четыре
рода. Клетки с несколькими или многочисленными
ядрами, частью уплощённые (Opalina, Zelleria), иног-
да до 3 мм. Раньше относились к Ciliata, поскольку
вся поверхность покрыта тысячами относительно ко-
ротких ресничковидных жгутиков (илл. 52); у них,
однако, нет ядерного дуализма, альвеол и типичной
инфрацилиатуры. Сходство в строении кортекса де-
лает более правдоподобным тесное родство с Prote-
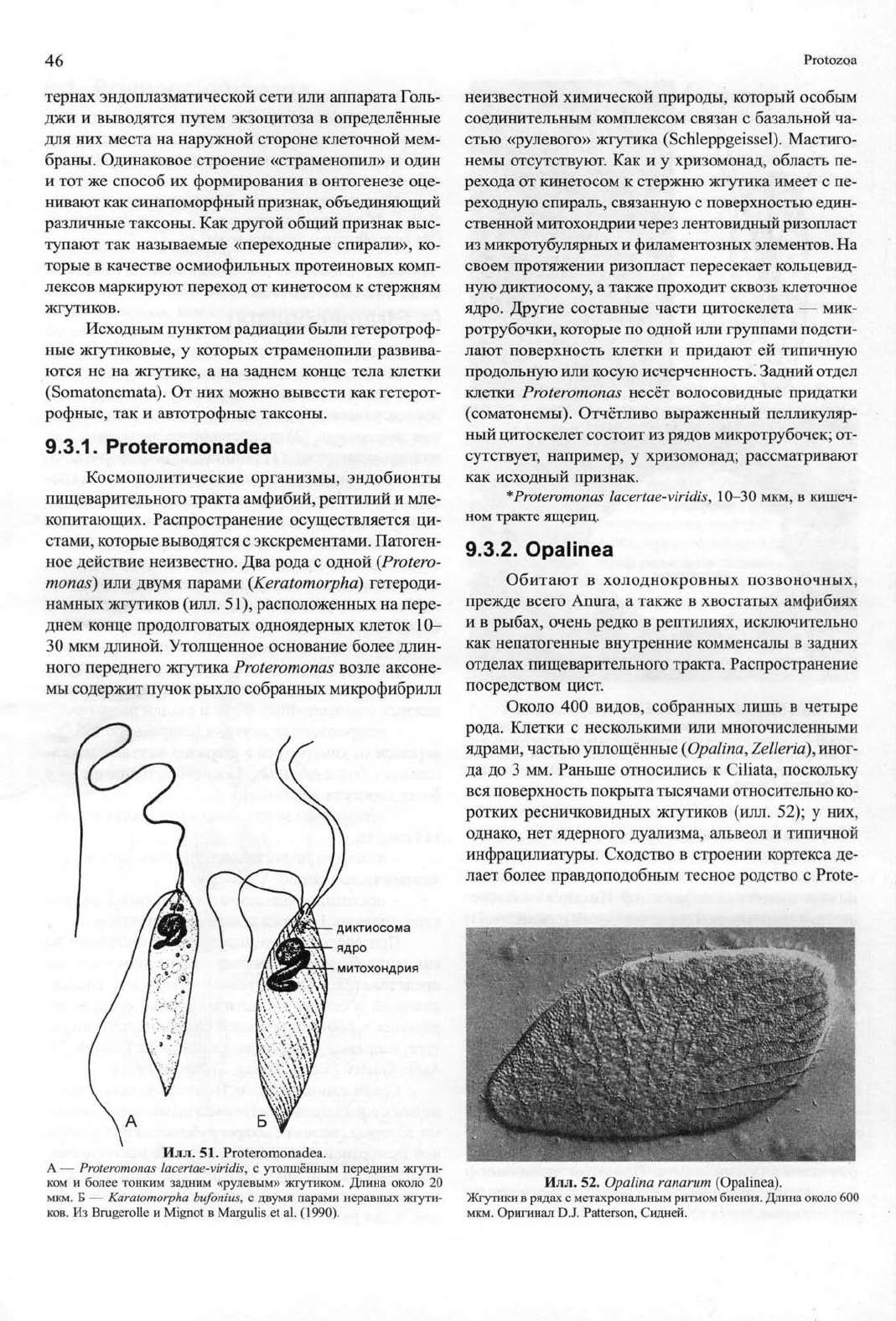
Илл. 51. Proteromonadea.
А — Proteromonas lacertae-viridis, с утолщённым передним жгути-
ком и более тонким задним «рулевым» жгутиком. Длина около 20
мкм.
Б — Karatomorpha bufonius, с двумя парами неравных жгути-
ков.
Из Brugerolle и Mignot в Margulis et al. (1990).
Илл.
52. Opalina ranarum (Opalinea).
Жгутики в рядах с метахрональным ритмом биения. Длина около 600
мкм.
Оригинал D.J. Patterson, Сидней.
http://jurassic.ru/

Protozoa
47
Илл.
S3.
Opalina ranarum
(Opalinea).
Жизненный
цикл.
С
изменениями
из
Grell
(1980).
romonadea, хотя на жгутиках отсутствуют, возмож-
но,
вторично, мастигонемы.
Жгутики расположены в частых косо идущих
рядах, в их биении заметны метахрональные волны.
Между отдельными рядами находится несколько
складок, поддерживаемых многочисленными микро-
трубочками. Нет клеточного рта для приёма оформ-
ленной пищи: питание только посредством микро-
пиноцитоза. В отличие от Ciliata, которые делятся,
главным образом, поперечно ходу рядов ресничек, у
Opalinea плоскость деления проходит между рядами
жгутиков (интеркинетально), то есть наискось к про-
дольной оси клетки.
Жизненный цикл весьма сложный (илл. 53).
Трофические формы почти всегда находятся у взрос-
лых хозяев.
*
Opalina ranarum
размножается к периоду спарива-
ния
лягушек путём быстро следующих друг за другом де-
лений
без вставочных фаз роста. Образующиеся в резуль-
тате
мелкие клетки имеют лишь немного ядер и немного-
численные остатки первоначально присутствующих рядов
жгутиков.
Возможно, быстрая фаза
размножения
вызыва-
ется
и контролируется гормонами хозяина. Микроформы
инцистируются
лишь с
тремя-шестью
ядрами и выводят-
ся
из
хозяина
с
содержимым
кишечника.
Цисты могут
про-
водить
в воде несколько недель,
проглатываются
головас-
тиками
с пищей и проникают в их
кишечный
тракт.
После
этого
последовательность делений, сопряжённая с
мейо-
тическими
делениями ядра, приводит к
образованию
тон-
ких
микрогамет
и крупных
макрогамет.
Диплоидная
зиго-
та
инцистируется
и выносится в воду. Далее зигота вновь
проглатывается
повзрослевшим
головастиком,
попадает в
его
желудок и
кишечник,
после чего образуется новое по-
коление
гамет или происходит развитие крупных трофи-
ческих
форм.
Генерация цист может также возникнуть
бес-
полым
путем. Различные возможности развития на этой
стадии
жизненного цикла обеспечивают очень эффектив-
ное заражение головастиков.
—
*Protoopalina
intestinalis,
330 мкм,
у
видовBombina.
—
Cepedea
obovoidea,
310 мкм,
у видов
Bufo.
9.3.3.
Chrysomonadea
Около 1000 фотоавтотрофных или гетеротроф-
ных видов, обитающих, главным образом, в пресных
водах. Клетки некрупные (5-20 мкм). В типичном
случае несут два анизоконтных гетеродинамных жгу-
тика на переднем конце тела; более длинный жгутик
направлен вперёд и служит двигателем, снабжён дву-
мя рядами жёстких трёхчленных мастигонем (плев-
ронематический жгутик) (илл. 11
Б).
Второй жгутик,
если он есть, гладкий и значительно более короткий;
проходит вдоль клетки назад и имеет базальное утол-
щение, которое расположено напротив слабовогну-
того впячивания на переднем конце клетки. Здесь,
внутри хлоропласта, обычно имеется красный гла-
зок из липидной гранулы (илл. 9).
Имеется только одно ядро, оно связано с базаль-
ным телом плевронематического жгутика через по-
перечно-исчерченную корешковую структуру (ризо-
пласт).
По сторонам от ядра находятся одна или две
пульсирующие вакуоли, а также одна или несколько
диктиосом, которые, помимо прочего, служат места-
ми образования мастигонем. Обычно клетка голая,
но у некоторых родов (Synura, Mallomonas, Paraphy-
somonas) на поверхности клетки имеются также
изящные чешуйки из кремнезёма, которые создают-
ся в вакуолях возле хлоропластов. Некоторые фор-
мы,
такие как Dinobryon (илл. 54), имеют «домики»
(loricae). В качестве экструсом известны дискоболо-
цисты. У некоторых родов (например, Dinobryon)
имеется половой процесс (изогамия): при этом две
одиночных клетки без образования гамет сливают-
ся.
Получившаяся зигота окружается окремнённой
оболочкой цисты, образующейся внутри протоплас-
та (эндогенно), и переходит в покоящуюся стадию.
Эндогенные покоящиеся цисты могут образовывать-
ся также бесполым путём.
Хлоропласты фотосинтезирующих представи-
телей одиночные или парные, имеют характерную
золотисто-жёлтую или коричневатую окраску из-за
фукоксантина, сопровождающего хлорофиллы а и с.
Запасными веществами служат липиды и полисаха-
рид хризоламинарин, откладывающиеся в вакуолях
снаружи от хлоропластов. Эти хлоропласты окружа-
ют клеточное ядро и морфологически связаны с ним
общей мембранной оболочкой из эндоплазматичес-
кого ретикулюма. Вместе с ней хлоропласты окру-
жены тремя мембранными системами (в области кон-
http://jurassic.ru/

48
Protozoa
такта
с
ядром
они
отграничены внутренней,
а в ос-
тальных их частях — наружной ядерной мембраной),
оставшимися, по-видимому,
от
деградировавшей
эукариотной клетки. Внутри хлоропластов находят-
ся стопки
из
трёх тилакоидов,
по
периферии окру-
женные опоясывающей ламеллой, также
из
трёх
ти-
лакоидов (илл.
17).
Наряду с монадным типом (одноклеточные или
их колониальные объединения) имеются
и
многочис-
ленные переходы
к
более высоким ступеням органи-
зации. Очевидна тенденция
к
становлению ризопод-
ного плана строения,
что
сопровождается редукци-
ей пластид
и
жгутиков,
и
формированием различных
типов псевдоподий, таким образом, вторично снова
приводя
к
«животным» формам. Среди них, наряду
с одноклеточными (такими
как
Chrysamoeba),
встречаются также
и
плазмоидальные агрегаты, раз-
мер которых исчисляется миллиметрами (Chrysarach-
niori).
Хризомонады
—
важнейший компонент нано-
планктона. Значение фотоавтотрофных представите-
лей состоит
в их
функции первичных продуцентов.
Массовое размножение, которое может происходить
в холодное время года, иногда создаёт проблемы для
рыбоводства из-за выделения продуктов обмена
ве-
ществ (кетонов
и
альдегидов). Хризомонады, отми-
рающие
в
начале тёплого периода, портят питьевую
Илл. 54.
Chrysomonadea.
А
—
Dinobryon
sp.,
длина отдельных клеток около
30 мкм. Б —
Pedinella sp.; длина около
10
мкм.
А —
оригинал
W.
Herth, Гейдель-
берг;
Б — по
Swale (1969).
воду
в
резервуарах, образуя дурнопахнущие продук-
ты разложения.
9.3.3.1.
Chrysomonadida
Свободноплавающие
или
сидячие монадные
формы (Ochromonas (илл.
9),
Dinobryon, Mallomonas,
Synura),
а
также формы
с
ризоподной организацией
(Chrysamoeba, Chrysarachniori).
9.3.3.2.
Pedinellidida
Сидячие, обычно
с
хлоропластами. Наряду
с
едва заметным жгутиком, направленным назад, на пе-
реднем конце клетки имеется длинный жгутик,
ок-
ружённый воротничком
из
упругих отростков клет-
ки (илл. 54). Строение Pedinella spp.
в
некоторых от-
ношениях сильно напоминает строение хоанофлагел-
лят; по-видимому,
это
конвергенция.
9.3.3.3.
Silicoflagellida
Таксон беден видами. Из-за кремнезёмного ске-
лета имеют большое значение для палеонтологии: ис-
копаемые остатки известны с мела, но время наиболь-
шего разнообразия приходится
на
третичный пери-
од.
Современные формы только
с
одним жгутиком
и
многочисленными тонкими псевдоподиями (илл. 55).
Скелетные элементы преимущественно внеклеточ-
ные,
звездообразные. Отнесение
к
хризомонадам ос-
новано на
том,
что, помимо наличия кремнезёма, име-
ются совпадающие детали строения хлоропластов;
однако вопрос остаётся очень спорным. Только один
современный
род с
двумя видами.
Dictyocha (Distephanus) fibula,
с псевдоподиями, бо-
лее 100 мкм,
в
холодных морях до 20°С.
9.3.4.
Heteromonadea
Билатерально-симметричные; напоминают хри-
зомонад наличием гетероконтных жгутиков
и
очень
сходными ультраструктурными признаками. Наряду
с длинным плевронематическим моторным жгути-
Илл. 55. Distephanus speculum
(Silicoflagellida).
Диаметр
40
мкм.
По
данным разных авторов.
http://jurassic.ru/
Protozoa
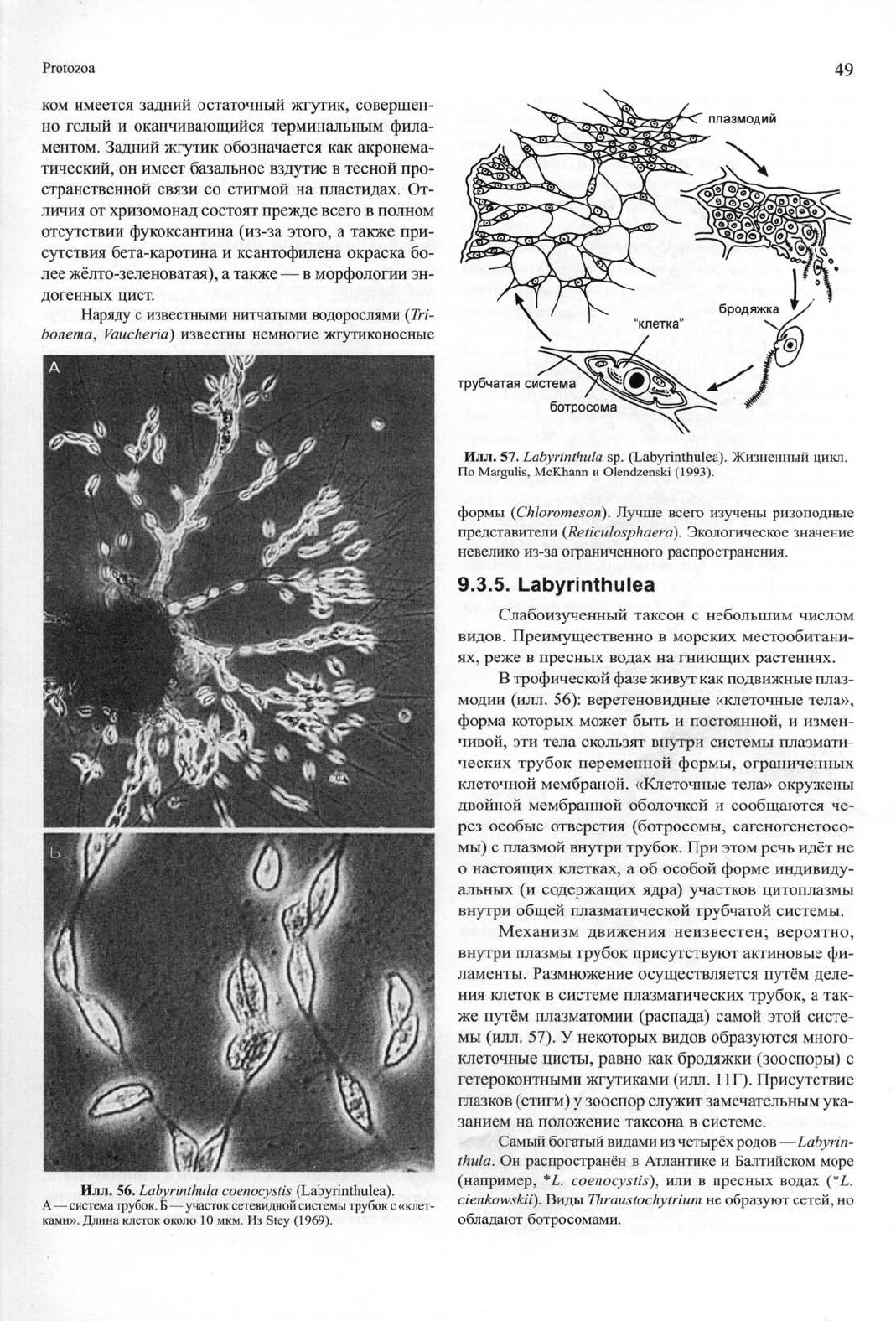
Илл.
56. Labyrinthula coenocystis
(Labyrinthulea).
А — система трубок.
Б
— участок сетевидной системы трубок с «клет-
ками». Длина клеток около 10 мкм. Из Stey (1969).
49
Илл.
57. Labyrinthula
sp. (Labyrinthulea). Жизненный цикл.
По Margulis, McKhann
и
Olendzenski (1993).
формы (Chloromeson). Лучше всего изучены ризоподные
представители (Reticulosphaera). Экологическое значение
невелико из-за ограниченного распространения.
9.3.5.
Labyrinthulea
Слабоизученный таксон
с
небольшим числом
видов. Преимущественно
в
морских местообитани-
ях, реже
в
пресных водах на гниющих растениях.
В трофической фазе живут как подвижные плаз-
модии (илл. 56): веретеновидные «клеточные тела»,
форма которых может быть
и
постоянной,
и
измен-
чивой, эти тела скользят внутри системы плазмати-
ческих трубок переменной формы, ограниченных
клеточной мембраной. «Клеточные тела» окружены
двойной мембранной оболочкой
и
сообщаются че-
рез особые отверстия (ботросомы, сагеногенетосо-
мы) с плазмой внутри трубок. При этом речь идёт не
о настоящих клетках,
а
об особой форме индивиду-
альных
(и
содержащих ядра) участков цитоплазмы
внутри общей плазматической трубчатой системы.
Механизм движения неизвестен; вероятно,
внутри плазмы трубок присутствуют актиновые фи-
ламенты. Размножение осуществляется путём деле-
ния клеток
в
системе плазматических трубок, а так-
же путём плазматомии (распада) самой этой систе-
мы (илл. 57). У некоторых видов образуются много-
клеточные цисты, равно как бродяжки (зооспоры)
с
гетероконтными жгутиками (илл.
11
Г).
Присутствие
глазков (стигм) у зооспор служит замечательным ука-
занием на положение таксона
в
системе.
Самый богатый видами из четырёх родов —Labyrin-
thula. Он распространён в Атлантике
и
Балтийском море
(например, *L. coenocystis), или
в
пресных водах
(*L.
cienkowskii). Виды Thraustochytrium не образуют сетей, но
обладают ботросомами.
ком имеется задний остаточный жгутик, совершен-
но голый
и
оканчивающийся терминальным фила-
ментом. Задний жгутик обозначается как акронема-
тический, он имеет базальное вздутие в тесной про-
странственной связи
со
стигмой
на
пластидах. От-
личия от хризомонад состоят прежде всего в полном
отсутствии фукоксантина (из-за этого,
а
также при-
сутствия бета-каротина и ксантофилена окраска бо-
лее жёлто-зеленоватая), а также — в морфологии эн-
догенных цист.
Наряду
с
известными нитчатыми водорослями (7W-
bonema, Vaucheria) известны немногие жгутиконосные
http://jurassic.ru/

50
Protozoa
корешковые элементы
ядро
мукоцисты
Илл.
58.
Chattonella
sp.
(Raphidomonadea).
Схема организации.
Длина
50
мкм.
По
Margulis, McKhann
и
Olendzenski (1993).
9.3.6.
Raphidomonadea
Всего десять фотоавтотрофных видов, имею-
щих исключительно монадную организацию, обыч-
но уплощённых дорсо-вентрально. Клетки
без
стен-
ки
и
наружной плотной оболочки достигают необыч-
но большого размера
— до 90 мкм.
Форма клетки
относительно постоянная благодаря упругой пелли-
куле (илл. 58).
В
клетке имеются мукоцисты,
а
также
варианты веретеновидных трихоцист, функциональ-
ное значение которых неясно. Гетероконтные жгу-
тики начинаются
в
вентральной борозде на переднем
конце; кинетосомы связаны
с
поверхностью клетки.
Более длинный жгутик направлен назад, он гладкий,
не имеет базального утолщения
и, как
правило,
не-
активен. Передний, более короткий
и
очень быстро
двигающийся моторный жгутик с упругими тубуляр-
ными мастигонемами. Глазок отсутствует. Характер-
но кольцо
из
элементов аппаратов Гольджи, окружа-
ющее апикальную часть крупного ядра.
У
пресно-
водных родов (как минимум,
у
Vacuolaria) пузырьки
аппарата Гольджи, непрерывно увеличиваясь
в
раз-
мерах, превращаются
в
сократительные вакуоли.
Пластиды многочисленные, светло-зелёные,
с
хлорофиллами
а я с,
а также
с
различными дополни-
тельными пигментами (каротин, ксантофилл
или
фукоксантин).
В
пластидах имеются стопки
из
трёх
тилакоидов
и, как
правило, опоясывающая ламелла.
Запасные вещества
в
виде липидных капель.
9.3.7.
Plasmodiophorea
Облигатные внутриклеточные паразиты
в
кор-
нях растений,
в
которых
они
образуют плазмодии.
Плодовые тела
й
образующиеся
в них
споры попа-
дают
во
внешнюю среду
при
разрушении растения.
Плазмодии следующего поколения образуются
в
ре-
зультате полового слияния свободных зооспор,
ко-
торые имеют гетероконтные жгутики без мастигонем.
При проникновении через клеточную стенку
у них
вступают в действие специальные органеллы («жало»
и «ствол» как особые производные эндоплазматичес-
кой сети).
Как у
одиночных клеток,
так и у
плазмо-
диев
во
время деления ядер наблюдается характер-
ный внутриядерный крест, который образуется
из
ядрышек, растягиваемых
к
полюсам, или,
на
стадии
метафазы,
из
упорядоченных хромосом.
*Plasmodiophora brassicae, возбудитель килы
у
раз-
личных сортов капусты.
—
*Виды Spongospora вызывают
порошистую паршу картофеля.
9.3.8.
Bicosoecidea
Около
40
видов одноклеточных (размером око-
ло
5
мкм)
или
колониальных организмов, живущих
на различных субстратах (Bicosoeca)
или
свободно
плавающих
в
морском
или
пресноводном планкто-
не.
Они представляют собой значительную часть ге-
теротрофного нанопланктона
и
важных консументов
водных бактерий. Живут
в
вазовидных домиках
(loricae), которые
—
в
отличие от хоанофлагеллят или
хризомонад
—
состоят
из
хитина. Одноядерные,
с
гетероконтными жгутиками. Жгутик, отходящий
на-
зад,
гладкий
и
расположен
в
желобке вдоль поверх-
ности клетки, служит
для
прикрепления внутри
до-
мика. Второй, более длинный жгутик, направлен впе-
рёд,
с
одним или двумя рядами упругих мастигонем,
которые создают
ток
воды, приносящий бактерий
и
сходные
с
ними пищевые частицы.
Bicosoeca (Bicoeca) socialis, 10 мкм, обычно образу-
ют колонии.
10. Alveolata
Данные
по
различным последовательностям
рРНК показывают,
что три
крупные
и
непохожие
группы традиционной системы (Dinoflagellata,
Apicomplexa
и
Ciliophora) образуют монофилети-
ческий таксон.
В
свете этих результатов молекуляр-
http://jurassic.ru/
Protozoa
51
но-биологических исследований некоторые из тех
структурных комплексов, которые раньше трактова-
лись как возникшие независимо друг от друга, сей-
час представляются гомологичными; таковы амфи-
есмы жгутиковых динофлагеллят, внутренние мем-
бранные комплексы безжгутиковых Apicomplexa,
а также альвеолы ресничных инфузорий. Кроме
того,
парасомальные мешочки Ciliophora, пузулы ди-
нофлагеллят и микропоры Apicomplexa могут быть
гомологичными структурами, выполняющими фун-
кцию обмена веществами с внешней средой. Пред-
полагаемым предком всех трёх таксонов был двужгу-
тиковый одноклеточный организм, живший пример-
но 900 млн. лет назад и имевший пелликулярную си-
стему вакуолей. Жгутики этого предка были, по-
видимому, гетероконтными и частично с мастигоне-
мами. Это состояние сохранилось только у динофла-
геллят.
10.1.
Dinoflagellata
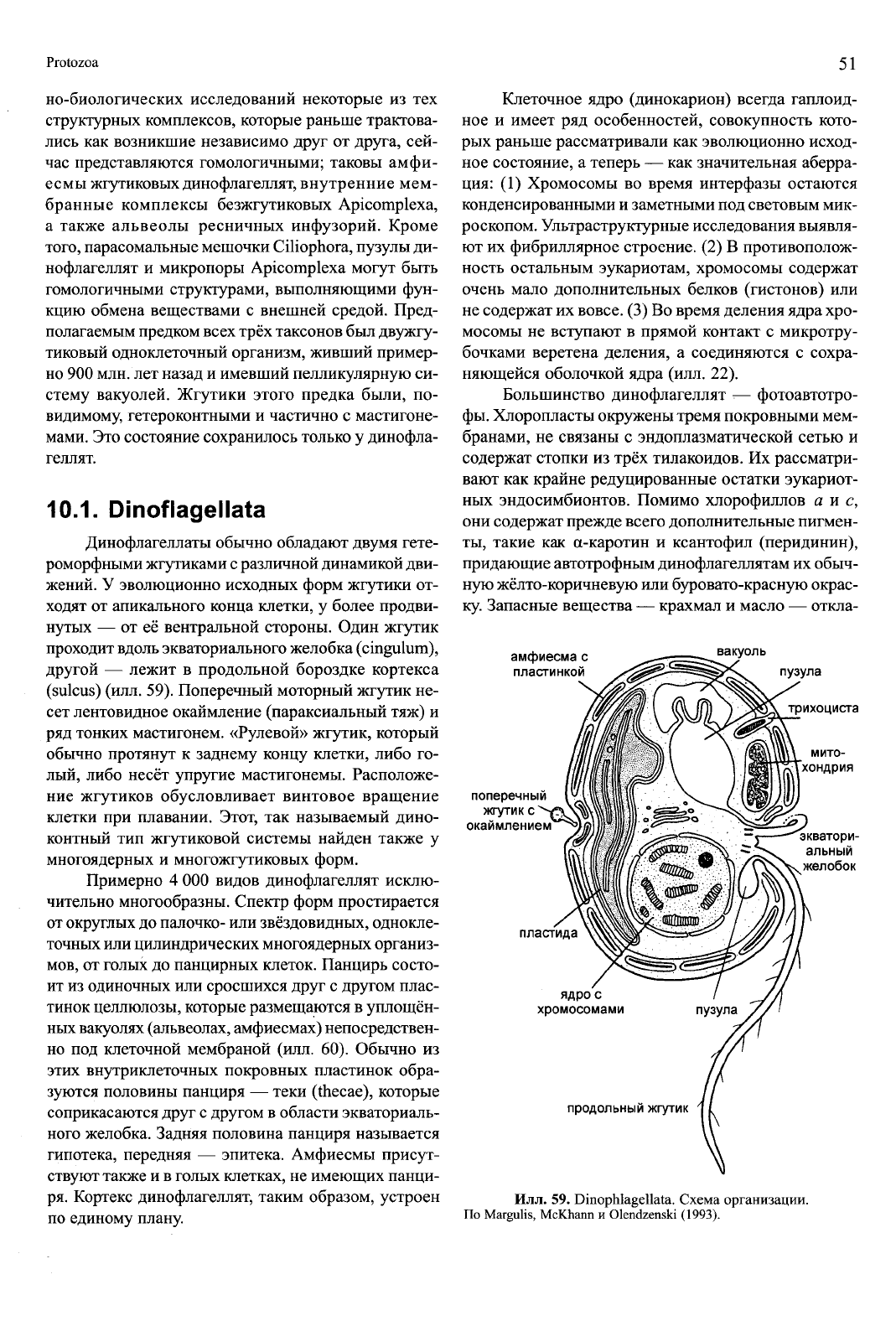
Динофлагеллаты обычно обладают двумя гете-
роморфными жгутиками с различной динамикой дви-
жений. У эволюционно исходных форм жгутики от-
ходят от апикального конца клетки, у более продви-
нутых — от её вентральной стороны. Один жгутик
проходит вдоль экваториального желобка (cingulum),
другой — лежит в продольной бороздке кортекса
(sulcus) (илл. 59). Поперечный моторный жгутик не-
сет лентовидное окаймление (параксиальный тяж) и
ряд тонких мастигонем. «Рулевой» жгутик, который
обычно протянут к заднему концу клетки, либо го-
лый, либо несёт упругие мастигонемы. Расположе-
ние жгутиков обусловливает винтовое вращение
клетки при плавании. Этот, так называемый дино-
контный тип жгутиковой системы найден также у
многоядерных и многожгутиковых форм.
Примерно 4
ООО
видов динофлагеллят исклю-
чительно многообразны. Спектр форм простирается
от округлых до палочко- или звёздовидных, однокле-
точных или цилиндрических многоядерных организ-
мов,
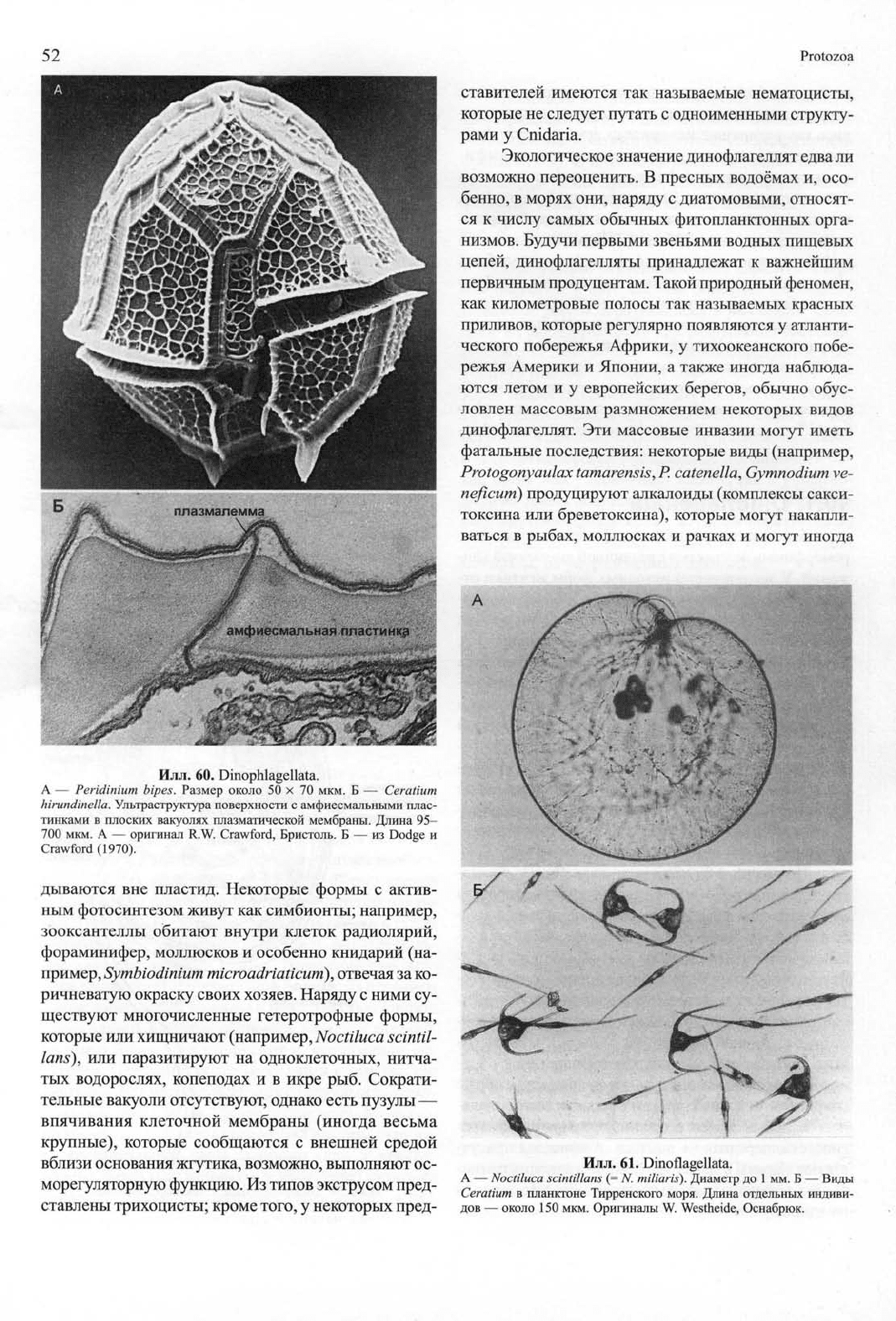
от голых до панцирных клеток. Панцирь состо-
ит из одиночных или сросшихся друг с другом плас-
тинок целлюлозы, которые размещаются в уплощён-
ных вакуолях (альвеолах, амфиесмах) непосредствен-
но под клеточной мембраной (илл. 60). Обычно из
этих внутриклеточных покровных пластинок обра-
зуются половины панциря — теки (thecae), которые
соприкасаются друг с другом в области экваториаль-
ного желобка. Задняя половина панциря называется
гипотека, передняя — эпитека. Амфиесмы присут-
ствуют также и в голых клетках, не имеющих панци-
ря.
Кортекс динофлагеллят, таким образом, устроен
по единому плану.
Клеточное ядро (динокарион) всегда гаплоид-
ное и имеет ряд особенностей, совокупность кото-
рых раньше рассматривали как эволюционно исход-
ное состояние, а теперь — как значительная аберра-
ция: (1) Хромосомы во время интерфазы остаются
конденсированными и заметными под световым мик-
роскопом. Ультраструктурные исследования выявля-
ют их фибриллярное строение. (2) В противополож-
ность остальным эукариотам, хромосомы содержат
очень мало дополнительных белков (гистонов) или
не содержат их вовсе. (3) Во время деления ядра хро-
мосомы не вступают в прямой контакт с микротру-
бочками веретена деления, а соединяются с сохра-
няющейся оболочкой ядра (илл. 22).
Большинство динофлагеллят -.— фотоавтотро-
фы.
Хлоропласты окружены тремя покровными мем-
бранами, не связаны с эндоплазматической сетью и
содержат стопки из трёх тилакоидов. Их рассматри-
вают как крайне редуцированные остатки эукариот-
ных эндосимбионтов. Помимо хлорофиллов а и с,
они содержат прежде всего дополнительные пигмен-
ты,
такие как а-каротин и ксантофил (перидинин),
придающие автотрофным динофлагеллятам их обыч-
ную жёлто-коричневую или буровато-красную окрас-
ку. Запасные вещества — крахмал и масло — откла-
Илл. 59. Dinophlagellata. Схема организации.
По Margulis, McKhann и Olendzenski (1993).
http://jurassic.ru/
52
Илл. 60. Dinophlagellata.
А — Peridinium bipes. Размер около 50 х 70 мкм. Б — Ceratium
hirundinella. Ультраструктура поверхности с амфиесмальными плас-
тинками в плоских вакуолях плазматической мембраны. Длина 95-
700 мкм. А — оригинал R.W. Crawford, Бристоль. Б — из Dodge и
Crawford (1970).
дываются вне пластид. Некоторые формы с актив-
ным фотосинтезом живут как симбионты; например,
зооксантеллы обитают внутри клеток радиолярий,
фораминифер, моллюсков и особенно книдарий (на-
пример, Symbiodinium microadriaticum), отвечая за ко-
ричневатую окраску своих хозяев. Наряду с ними су-
ществуют многочисленные гетеротрофные формы,
которые или хищничают (например, Noctiluca scintil-
lans),
или паразитируют на одноклеточных, нитча-
тых водорослях, копеподах и в икре рыб. Сократи-
тельные вакуоли отсутствуют, однако есть пузулы —
впячивания клеточной мембраны (иногда весьма
крупные), которые сообщаются с внешней средой
вблизи основания жгутика, возможно, выполняют ос-
морегуляторную функцию. Из типов экстру сом пред-
ставлены трихоцисты; кроме того, у некоторых пред-
Protozoa
Илл. 61. Dinoflagellata.
А — Noctiluca scintillans (= N. miliaris). Диаметр до 1 мм. Б — Виды
Ceratium в планктоне Тирренского моря. Длина отдельных индиви-
дов — около 150 мкм. Оригиналы W. Westheide, Оснабрюк.
ставителей имеются так называемые нематоцисты,
которые не следует путать с одноименными структу-
рами у Cnidaria.
Экологическое значение динофлагеллят едва ли
возможно переоценить. В пресных водоёмах и, осо-
бенно, в морях они, наряду с диатомовыми, относят-
ся к числу самых обычных фитопланктонных орга-
низмов. Будучи первыми звеньями водных пищевых
цепей, динофлагелляты принадлежат к важнейшим
первичным продуцентам. Такой природный феномен,
как километровые полосы так называемых красных
приливов, которые регулярно появляются у атланти-
ческого побережья Африки, у тихоокеанского побе-
режья Америки и Японии, а также иногда наблюда-
ются летом и у европейских берегов, обычно обус-
ловлен массовым размножением некоторых видов
динофлагеллят. Эти массовые инвазии могут иметь
фатальные последствия: некоторые виды (например,
Protogonyaulax tamarensis,
P.
catenella, Gymnodium ve-
neficum) продуцируют алкалоиды (комплексы сакси-
токсина или бреветоксина), которые могут накапли-
ваться в рыбах, моллюсках и рачках и могут иногда
http://jurassic.ru/
Protozoa
53
приводить к гибели этих организмов и поедающих
их хищников. При употреблении в пищу мидий или
устриц, накопивших много токсинов в местах мас-
сового размножения динофлагеллят, у человека обыч-
но наступает отравление, сопровождаемое парали-
чом и необратимой остановкой дыхания. Но не все
красные приливы смертоносны: массовое появление
гетеротрофной Noctiluca scintillans вызывает, напри-
мер,
лишь свечение моря, обычное в летние месяцы
в Северном море. Это свечение обусловлено актив-
ностью люциферин-люциферазной системы.
Многие динофлагеллаты могут переносить не-
благоприятные условия среды, образуя покоящиеся
цисты; в виде микрофоссилий они известны как ми-
нимум с силура (400 млн. лет назад).
*Ceratium hirundinella, 700 мкм, с одним апикаль-
ным
и
двумя антиапикальными рогами,
в
море и
в
пресных
водах. Многие другие виды Ceratium (илл.
61 Б)
характер-
ны
для пресноводного и морского планктона. — *Noctiluca
scintillans (= N. miliaris), 1 мм, эпитека редуцирована до
одного щупальца; морской хищник, вызывает свечение
моря.
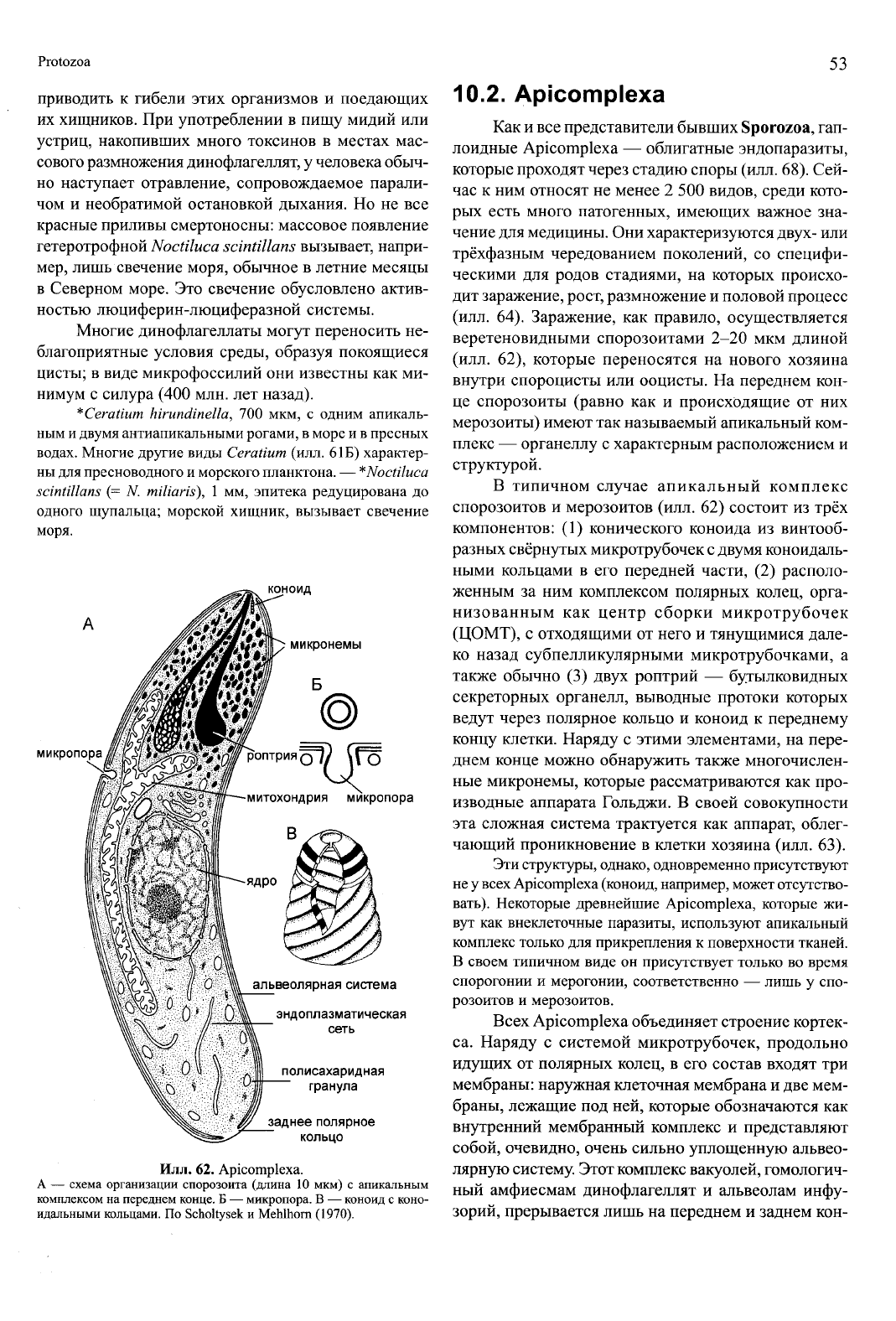
Илл. 62. Apicomplexa.
А — схема организации спорозоита (длина 10 мкм) с апикальным
комплексом на переднем конце. Б — микропора. В — коноид с коно-
идальными кольцами. По Scholtysek и Mehlhorn (1970).
10.2. Apicomplexa
Как и все представители бывших Sporozoa, гап-
лоидные Apicomplexa — облигатные эндопаразиты,
которые проходят через стадию споры (илл. 68). Сей-
час к ним относят не менее 2 500 видов, среди кото-
рых есть много патогенных, имеющих важное зна-
чение для медицины. Они характеризуются двух- или
трёхфазным чередованием поколений, со специфи-
ческими для родов стадиями, на которых происхо-
дит заражение, рост, размножение и половой процесс
(илл. 64). Заражение, как правило, осуществляется
веретеновидными спорозоитами 2-20 мкм длиной
(илл. 62), которые переносятся на нового хозяина
внутри спороцисты или ооцисты. На переднем кон-
це спорозоиты (равно как и происходящие от них
мерозоиты) имеют так называемый апикальный ком-
плекс — органеллу с характерным расположением и
структурой.
В типичном случае апикальный комплекс
спорозоитов и мерозоитов (илл. 62) состоит из трёх
компонентов: (1) конического коноида из винтооб-
разных свёрнутых микротрубочек с двумя коноидаль-
ными кольцами в его передней части, (2) располо-
женным за ним комплексом полярных колец, орга-
низованным как центр сборки микротрубочек
(ЦОМТ), с отходящими от него и тянущимися дале-
ко назад субпелликулярными микротрубочками, а
также обычно (3) двух роптрий — бутылковидных
секреторных органелл, выводные протоки которых
ведут через полярное кольцо и коноид к переднему
концу клетки. Наряду с этими элементами, на пере-
днем конце можно обнаружить также многочислен-
ные микронемы, которые рассматриваются как про-
изводные аппарата Гольджи. В своей совокупности
эта сложная система трактуется как аппарат, облег-
чающий проникновение в клетки хозяина (илл. 63).
Эти структуры, однако, одновременно присутствуют
не у всех Apicomplexa (коноид, например, может отсутство-
вать).
Некоторые древнейшие Apicomplexa, которые жи-
вут как внеклеточные паразиты, используют апикальный
комплекс только для прикрепления к поверхности тканей.
В своем типичном виде он присутствует только во время
спорогонии и мерогонии, соответственно — лишь у спо-
розоитов и мерозоитов.
Всех Apicomplexa объединяет строение кортек-
са. Наряду с системой микротрубочек, продольно
идущих от полярных колец, в его состав входят три
мембраны: наружная клеточная мембрана и две мем-
браны, лежащие под ней, которые обозначаются как
внутренний мембранный комплекс и представляют
собой, очевидно, очень сильно уплощенную альвео-
лярную систему. Этот комплекс вакуолей, гомологич-
ный амфиесмам динофлагеллят и альвеолам инфу-
зорий, прерывается лишь на переднем и заднем кон-
http://jurassic.ru/

цах клетки для процессов экзоцитоза, и на боковых
участках — для эндоцитоза. Микропоры — одиноч-
ные или многочисленные впячивания поверхности
клетки, заметные в латеральной плоскости — слу-
жат для поступления питательных веществ. Недав-
но полученные доказательства наличия фотосинте-
тических пигментов в так называемых полых цилин-
драх спорозоитов (пузырьках с двойной мембраной)
можно трактовать как свидетельство того, что пред-
ки Apicomplexa имели пластиды.
Жизненные циклы Apicomplexa довольно слож-
ные.
В упрощённом виде обобщённый цикл можно
представить следующим образом (илл. 64). Одно-
ядерные спорозоиты появляются в ходе фазы размно-
жения (спорогонии), сопряжённой с мейотическим
редукционным делением. В новом хозяине они вы-
ходят из спороцист, проникают в клетки и растут в
них. В простейшем случае они развиваются в гамон-
тов,
которые многократно делятся, дифференциру-
агамогония
зигота
гамонты
ются в гаметы и, в итоге, объединяются в зиготы (га-
могония). В ходе этого двухфазного чередования по-
колений из зигот снова появляются многочисленные
спорозоиты.
У более эволюционно продвинутых форм меж-
ду спорогонией и гамогонией включаются ещё одна
или несколько фаз размножения. Эти фазы возника-
ют в результате преобразования спорозоитов в тро-
фонты (питающиеся клетки), которые в организмах
своих хозяев вырастают до крупных многоядерных
плазмодиев. Апикальный комплекс у них обычно
сильно деградирует, а дополнительные микропоры
обеспечивают поступление питания в клетку. Из них
при завершении фазы роста формируются шизонты
(меронты), из которых после их множественного де-
ления (шизогонии = мерогонии, илл. 68) образуются
мерозоиты. У них снова появляются апикальные ком-
плексы, благодаря чему мерозоиты могут поражать
другие клетки, и дифференцироваться в трофонтов
что приводит к многократному увеличению числа
паразитов в хозяине. Но мерозоиты неспособны, от-
части из-за смены хозяина, перейти к фазе гамого-
нии. Она обычно происходит как оогамия, причём
макрогамонт превращается прямо в макрогамету;
микрогамонты же, прежде чем образовать микрога-
меты, совершают ещё ряд делений. Отдельные ста-
дии, возникающие в ходе такого развития, можно раз-
личить по особым цитологическим признакам.
Спорогония служит для восстановления гаплоидной
фазы ядра, равно как — с последующими митотическими
делениями — для эффективного освоения потенциальных
новых хозяев. В результате делений во время шизогонии и
гамогонии возрастает число гамет
и
зигот, споро- и ооцист,
поэтому новое заражение становится более вероятным. Из-
за этого Apicomplexa принадлежат
к
числу опаснейших па-
разитов человека и многих животных.
Систематическое деление Apicomplexa до сих
пор вызывает дискуссии. Обычно практикуется раз-
деление на три субтаксона.
Илл. 64. Схема чередования поколений у Apicomplexa.
По данным разных авторов.
http://jurassic.ru/

Protozoa
Илл.
65. Monocystis sp. (Apicomplexa, Gregarinea).
Гамопт
в
жидкости семенного пузырька дождевого червя. Длина около
200 мкм. Нитевидные структуры — это почти зрелые спермин; раз-
личные шарики — стадии сперматогенеза хозяина, созревающие по
периферии цитофора — безъядерной цитоплазматической массы,
расположенной в центре. Оригинал W. Westheide, Оснабрюк.
10.2.1.
Gregarinea
Характерным признаком этой группы, охваты-
вающей около 500 видов (илл. 65), является способ-
ность мужских и женских гамонтов совершать мно-
гократные деления, благодаря чему производится
высокое, но примерно равное число гамет. Гаметы
обоих типов могут иметь различное (анизогамия) или
Илл. 66. Gregarina sp. (Gregarinea).
А — два гамонта, образующие сизигий, длина каждого индивидуума
500 мкм. Б — Gregarina garnhami, складчатость пелликулы. А —
оригинал К. Hausmann, Берлин; Б — из Walker et al. (1979).
55
в значительной мере сходное (изогамия) строение.
Первоначально в эволюции развитие гамет проходи-
ло раздельно и независимо друг от друга; гамонты
же более продвинутых таксонов уже в конце фазы
их роста образуют пары (сизигии) (илл. 66), а позже
инцистируются с образованием общей гамонтоцис-
ты,
в которой образование гамет и зигот протекает
согласованно по времени и месту (гамонтогамия). Из
зигот развиваются прямо спороцисты, которые со-
держат 4—16 гаплоидных спорозоитов.
Трофические стадии, выходящие из спорозои-
тов,
вырастают до крупных (иногда до 10 мм дли-
ной) гамонтов. Многие из них способны к характер-
ному скользящему движению, которое осуществля-
ется благодаря согласованному действию продоль-
ных складок тела, усиленных микрофиламентами
(илл. 66Б).
Тело гамонта обычно расчленено на два отде-
ла — передний протомерит и задний, более крупный
дейтомерит с ядром. Прото- и дейтомерит отделены
друг от друга кольцевой бороздой (илл. 66А). Обыч-
но протомерит несёт на переднем конце особый вы-
рост (эпимерит), образованный из коноида, который
служит для укрепления в эпителиальной клетке. Эпи-
мерит отбрасывается перед образованием сизигия,
когда протомерит задней клетки (сателлит) прикреп-
ляется к дейтомериту передней клетки (примиту).
Грегарины живут — кроме ранних стадий раз-
вития — преимущественно как внеклеточные пара-
зиты в кишечном тракте или в полостях тела анне-
лид, артропод, моллюсков, иглокожих и оболочни-
ков.
Наиболее распространены они у членистоногих.
Правда, патогенными являются лишь немногие виды.
Хозяйственное значение отсутствует.
*Monocystis lumbrici, около 200 мкм, шизонты регу-
лярно встречаются в семенных пузырьках различных ви-
дов дождевых червей (илл. 65). — *Gregarinapolymorpha,
350 мкм, в кишечнике личинок мучного хрущака (илл.
66).
— *Mattesia dispora, 3-12 мкм, в жировом теле мель-
ничной огнёвки Ephestia kuehniella, со стадией шизогонии
в жизненном цикле.
10.2.2.
Coccidea
Ход развития кокцидий существенно отличает-
ся от грегарин. В противоположность микрогамон-
там, макрогамонты не проходят через фазу деления,
а развиваются прямо в ооциты. В типичном случае
из микрогамонтов выходят трёхжгутиковые микро-
гаметы (илл. 67). Зигота преобразуется в зигоцисту
(ооцисту) и во время спорогонии делится на 4-32
(или более) споробластов, которые обычно покрыты
собственной оболочкой (спороциста) и после мито-
тических делений дают по два-восемь (или более)
спорозоитов.
http://jurassic.ru/