Вестхайде В.,Ригер Р.( ред.) Зоология беспозвоночных в двух томах. Том 1: от простейших до моллюсков и артропод
Подождите немного. Документ загружается.


136
Mesozoa
Илл.
180. Бесполое размножение
Dicyemidae (стадия нематогена).
А — аксобласт претерпевает неравномерное
деление, более мелкая клетка продолжает
делиться и формирует эпидермис. Б — ак-
собласт ещё раз делится неравномерно, бо-
лее мелкая клетка погружается в исходный
аксобласт (теперь уже аксиальную клетку) и
превращается в новый аксобласт. В — но-
вый аксобласт продолжает деление, и весь
процесс повторяется внутри аксиальной
клетки. По Lapan, Morowitz (1972), с изме-
нениями.
Размножение и развитие
Бесполое размножение в нематогене осуществ-
ляется аксобластами, которые содержатся в аксиаль-
ной клетке (илл. 179-181). Название группы дослов-
но означает «с двумя зародышами» (греч. di, суета =
два, зародыш). Молодой нематоген покидает мате-
ринскую особь через стенку тела. Повышение плот-
ности популяции дициемид в почках хозяина в тече-
ние нескольких часов вызывает (с помощью неуста-
новленного химического фактора) развитие стадии
ромбогена (илл. 180) с гермафродитными гонадами,
в которых происходит оплодотворение*. Из зигот пу-
тём спирального дробления развиваются инфузори-
шероховатый
эндоплазматический
эмбрион ретикулюм ядро
I
. //; /
соматической
лизосома
ri
.|. \ Ли / .
клетки
митохондрия
микровиллярные
складки
ленточная
десмосома
аксиальная
клетка
ресничка
аксобласт
Илл.
181. Dicyemidae.
Полусхематичный поперечный срез через нематоген. По данным раз-
ных авторов, из Starch и Welsch (1990), с изменениями.
формные личинки (илл. 182). Они билатерально-
симметричны, состоят из 37 (иногда 39) клеток и по
сложности организации превосходят материнских
особей. В составе тела личинки есть две крупные
клетки с сильно светопреломляющими включения-
ми,
содержащими инозитол-6-фосфат. Внутри личин-
ки ресничная полость с медианной наружной порой
(урночка) и четырьмя большими урночковыми клет-
ками. Личинки покидают хозяина через выделитель-
ные поры и мантийную полость.
Инфузориформные личинки инфицируют ново-
го хозяина, проникая сначала в жаберный отдел ман-
тийной полости, а затем в печень. Позже мелкие не-
матогены-основатели с тремя аксиальными клет-
ками появляются в выделительных органах.
Dicyema typus и D. clausilianum обитают в почках
головоногих моллюсков
Octopus
vulgaris,
Eledone moschata
и Sepia
officinalis.
— Kantharella
antarctica
из почек Pa-
raeledone turqueti
является хозяином микроспоридий. От-
личается непостоянным числом соматических и половых
клеток.
1.2.
Heterocyemidae
Данный таксон включает только два редких
вида Сопосуета polymorpha и Microcyema vespa, ко-
торые также обитают в выделительных органах го-
ловоногих моллюсков (Sepia, Octopus). Они значи-
тельно отличаются от Dicyemidae, что отражено в
названиях родов (греч. conos, heteros = конус, иначе;
* По другой, широко распространённой трактовке это
не гонады, а сильно упрощённые особи полового поколения
(инфузоригены), состоящие из одной-единственной сомати-
ческой клетки, мужских и женских половых клеток. Инфузо-
риген не покидает вакуоли внутри аксиальной клетки. Мужс-
кие и женские половые клетки инфузоригена соединяются, в
результате чего образуются зиготы, далее развивающиеся в
инфузориформных личинок
(прим.
ред.)
http://jurassic.ru/
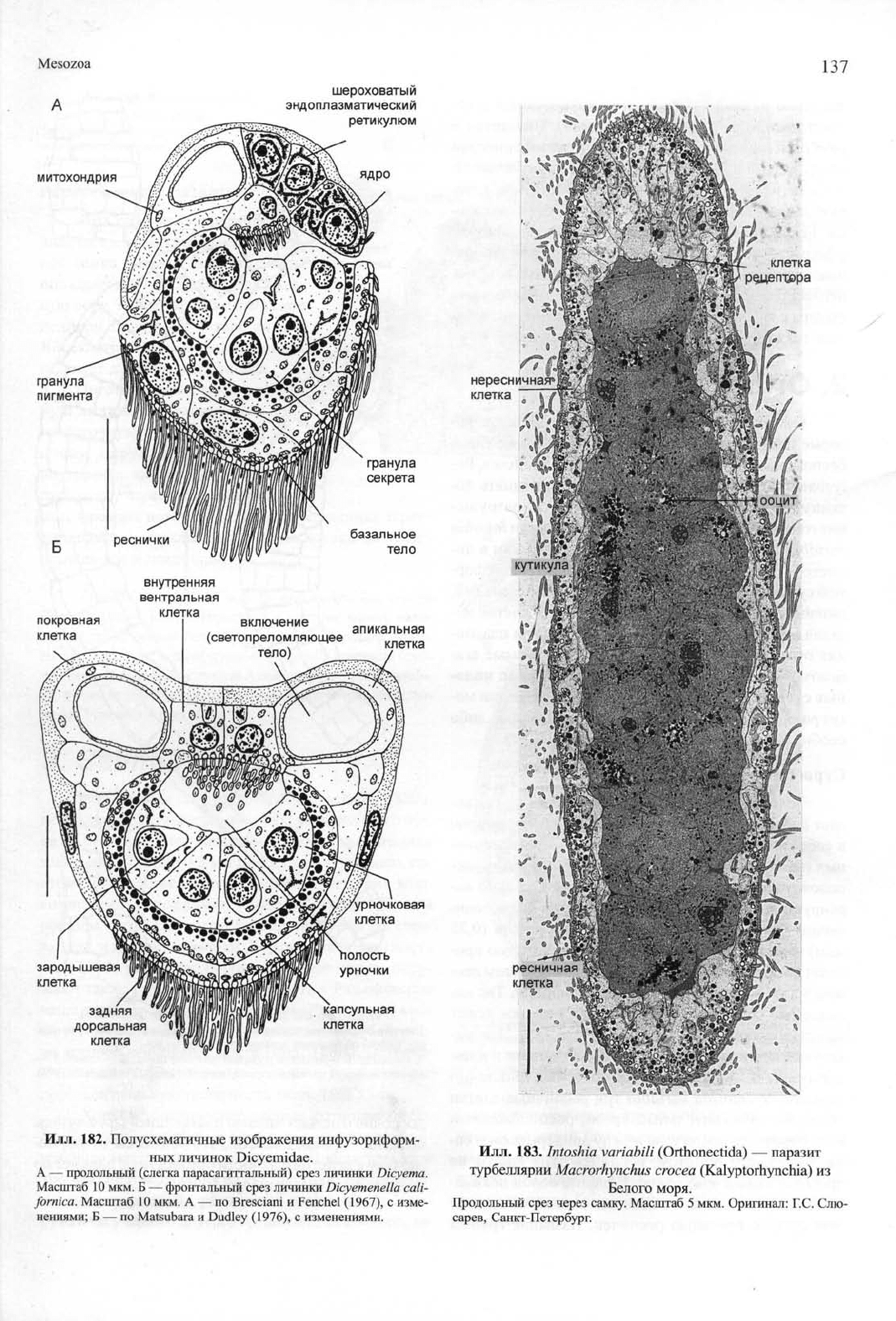
Илл.
182. Полусхематичные изображения инфузориформ-
ных личинок Dicyemidae.
А — продольный (слегка парасагиттальный) срез личинки Dicyema.
Масштаб 10 мкм. Б — фронтальный срез личинки Dicyemenella cali-
fornica. Масштаб 10 мкм. А — по Bresciani и Fenchel (1967), с изме-
нениями; Б — по Matsubara и Dudley (1976), с изменениями.
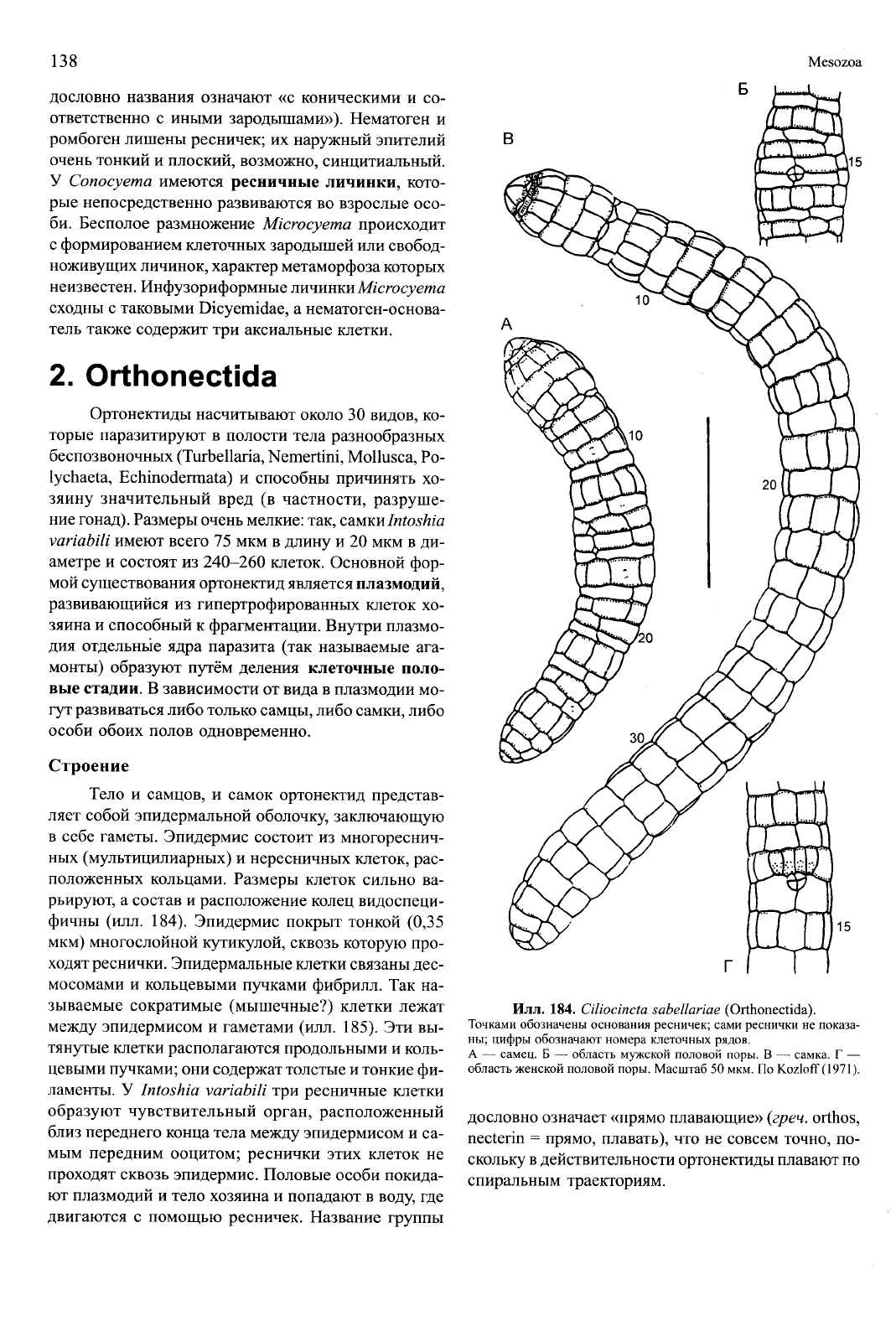
Илл.
183. Intoshia variabili (Orthonectida) — паразит
турбеллярии Macrorhynchus crocea (Kalyptorhynchia) из
Белого моря.
Продольный срез через самку. Масштаб 5 мкм. Оригинал: Г.С. Слю-
сарев, Санкт-Петербург.
http://jurassic.ru/
138
Mesozoa
Илл. 184.
Ciliocincta sabellariae
(Orthonectida).
Точками обозначены основания ресничек; сами реснички не показа-
ны;
цифры обозначают номера клеточных рядов.
А — самец. Б — область мужской половой поры. В — самка. Г —
область женской половой поры. Масштаб 50 мкм. По Kozloff(1971).
дословно означает «прямо плавающие» (греч. orthos,
necterin = прямо, плавать), что не совсем точно, по-
скольку в действительности ортонектиды плавают по
спиральным траекториям.
дословно названия означают «с коническими и со-
ответственно с иными зародышами»). Нематоген и
ромбоген лишены ресничек; их наружный эпителий
очень тонкий и плоский, возможно, синцитиальный.
У Сопосуета имеются ресничные личинки, кото-
рые непосредственно развиваются во взрослые осо-
би.
Бесполое размножение Microcyema происходит
с формированием клеточных зародышей или свобод-
ноживущих личинок, характер метаморфоза которых
неизвестен. Инфузориформные личинки Microcyema
сходны с таковыми Dicyemidae, а нематоген-основа-
тель также содержит три аксиальные клетки.
2.
Orthonectida
Ортонектиды насчитывают около 30 видов, ко-
торые паразитируют в полости тела разнообразных
беспозвоночных (Turbellaria, Nemertini, Mollusca, Ро-
lychaeta, Echinodermata) и способны причинять хо-
зяину значительный вред (в частности, разруше-
ние гонад). Размеры очень мелкие: так, самкиIntoshia
variabili имеют всего 75 мкм в длину и 20 мкм в ди-
аметре и состоят из 240-260 клеток. Основной фор-
мой существования ортонектид является плазмодий,
развивающийся из гипертрофированных клеток хо-
зяина и способный к фрагментации. Внутри плазмо-
дия отдельные ядра паразита (так называемые ага-
монты) образуют путём деления клеточные поло-
вые стадии. В зависимости от вида в плазмодии мо-
гут развиваться либо только самцы, либо самки, либо
особи обоих полов одновременно.
Строение
Тело и самцов, и самок ортонектид представ-
ляет собой эпидермальной оболочку, заключающую
в себе гаметы. Эпидермис состоит из многореснич-
ных (мультицилиарных) и нересничных клеток, рас-
положенных кольцами. Размеры клеток сильно ва-
рьируют, а состав и расположение колец видоспеци-
фичны (илл. 184). Эпидермис покрыт тонкой (0,35
мкм) многослойной кутикулой, сквозь которую про-
ходят реснички. Эпидермальные клетки связаны дес-
мосомами и кольцевыми пучками фибрилл. Так на-
зываемые сократимые (мышечные?) клетки лежат
между эпидермисом и гаметами (илл. 185). Эти вы-
тянутые клетки располагаются продольными и коль-
цевыми пучками; они содержат толстые и тонкие фи-
ламенты. У Intoshia variabili три ресничные клетки
образуют чувствительный орган, расположенный
близ переднего конца тела между эпидермисом и са-
мым передним ооцитом; реснички этих клеток не
проходят сквозь эпидермис. Половые особи покида-
ют плазмодий и тело хозяина и попадают в воду, где
двигаются с помощью ресничек. Название группы
http://jurassic.ru/

Mesozoa
139
ресничные клетки
Илл.
185.
Intoshia variabili
(Orthonectida).
Трёхмерная схема строения средней части
тела самки.
Из
Slyusarev (1994).
Размножение
и
развитие
При копуляции самец присое-
диняется
к
значительно более круп-
ной самке. Мелкие сперматозоиды
попадают внутрь самки через поло-
вую пору.
У
некоторых видов самец
целиком проникает
в
тело самки.
Яйцеклетка
в
ходе оплодотворения
отделяет
два
полярных тельца.
Зигота развивается
в
реснич-
ную личинку, которая имеет мно-
гоклеточную эпидермальную обо-
лочку, окружающую несколько
внутренних клеток. Личинка поки-
дает самку через разрыв эпидерми-
са
и
заражает нового хозяина. Затем личинка теряет
эпидермис,
а
каждая внутренняя клетка начинает
развиваться
в
плазмодий.
Intoshia variabili,
75
мкм,
в
Macrorhynchus crocea
(Plathelminthes, Kalyptorhynchia);
в
хозяине может нахо-
диться
до 30
особей (илл.
183,
185).
—
Rhopalura ophio-
сотае,
в
полости тела офиуры
Amphipholis squamata
(суб-
тропическая часть Восточной Атлантики).
—
Rh.
granulo-
sa, в
гонаде двустворчатого моллюска
Heteranomia
squa-
mula
(Западная Атлантика).
3. Salinella salve
Вид Salinella salve (илл. 186) был описан
в
1889
г.
по экземплярам, выделенным из грунта солёного озе-
ра
в
Аргентине. Этот питающийся детритом организм
имеет длину около
0,2 мм и,
согласно описанию,
со-
стоит
из
единственного слоя клеток, несущих мно-
гочисленные реснички
с
обеих сторон. Таким
образом, один слой клеток функционирует
и
как стен-
ка тела,
и
как стенка кишки. Дорсальная поверхность
тела несёт ригидные цирры; длинные цирры окру-
жают также
рот и
анальное отверстие. Размножение
чаще всего происходит поперечным делением; кро-
ме того, может происходить конъюгация особей, пос-
ле которой образуется циста
(илл.
186В). Встреча-
ются также одноклеточные личинки, проявляющие
дорсовентральную полярность (илл. 186Г).
Хотя светооптические данные
в
отношении
Sa-
linella очень подробны
и
опираются
на
изучение
об-
ширного материала
из
лабораторной культуры,
они
требуют подтверждения на ультраструктурном уров-
не.
«Двусторонний» ресничный эпителий
не
обна-
ружен ни
у
одного другого организма; более того,
та-
кутикула
нересничные
клетки
сократимые
клетки
ооцит
Илл.
186.
Salinella
salve.
А
—
сагиттальный срез; длина животного около
200
мкм.
Б — вид с
вентральной стороны.
В —
образование цисты после конъюгации двух
особей; диаметр около
120
мкм.
Г
—
одноклеточная ювенильная ста-
дия,
длина
23 мкм. По
Frenzel (1892),
с
изменениями.
кую структуру очень трудно объяснить
с
цитологи-
ческих позиций (двусторонние ленточные десмосо-
мы,
положение ресничных корешков, полярность
клетки?). Таким образом, обсуждать филогенетичес-
кое положение Salinella пока
не
представляется воз-
можным.
http://jurassic.ru/
IV. Двуслойные
Eumetazoa,
или Diploblastica
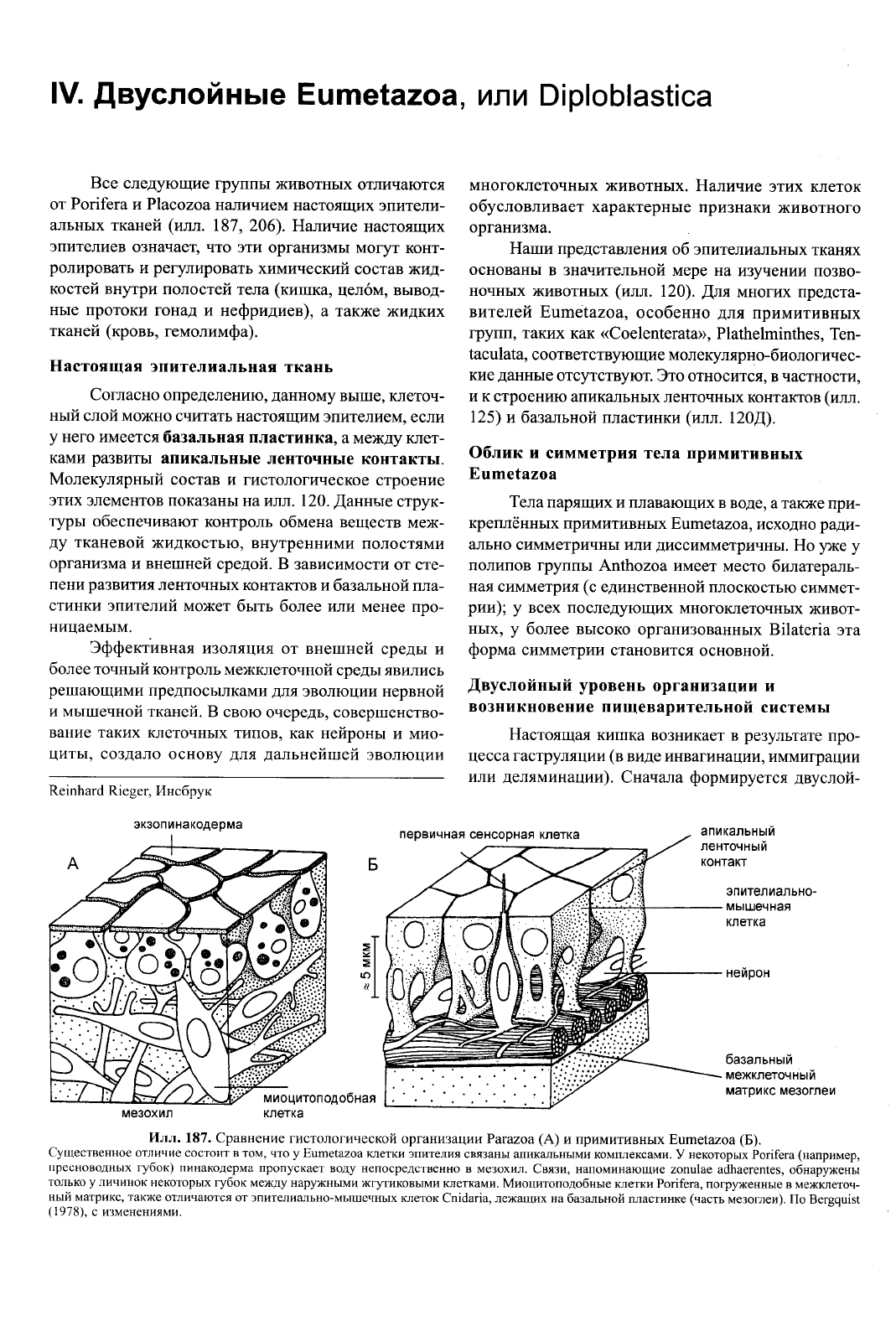
Все следующие группы животных отличаются
от Porifera и Placozoa наличием настоящих эпители-
альных тканей (илл. 187, 206). Наличие настоящих
эпителиев означает, что эти организмы могут конт-
ролировать и регулировать химический состав жид-
костей внутри полостей тела (кишка, целом, вывод-
ные протоки гонад и нефридиев), а также жидких
тканей (кровь, гемолимфа).
Настоящая эпителиальная ткань
Согласно определению, данному выше, клеточ-
ный слой можно считать настоящим эпителием, если
у него имеется базальная пластинка, а между клет-
ками развиты апикальные ленточные контакты.
Молекулярный состав и гистологическое строение
этих элементов показаны на илл. 120. Данные струк-
туры обеспечивают контроль обмена веществ меж-
ду тканевой жидкостью, внутренними полостями
организма и внешней средой. В зависимости от сте-
пени развития ленточных контактов и базальной пла-
стинки эпителий может быть более или менее про-
ницаемым.
Эффективная изоляция от внешней среды и
более точный контроль межклеточной среды явились
решающими предпосылками для эволюции нервной
и мышечной тканей. В свою очередь, совершенство-
вание таких клеточных типов, как нейроны и мио-
циты, создало основу для дальнейшей эволюции
Reinhard Rieger, Инсбрук
многоклеточных животных. Наличие этих клеток
обусловливает характерные признаки животного
организма.
Наши представления об эпителиальных тканях
основаны в значительной мере на изучении позво-
ночных животных (илл. 120). Для многих предста-
вителей Eumetazoa, особенно для примитивных
групп, таких как «Coelenterata», Plathelminth.es, Теп-
taculata, соответствующие молекулярно-биологичес-
кие данные отсутствуют. Это относится, в частности,
и к строению апикальных ленточных контактов (илл.
125) и базальной пластинки (илл. 120Д).
Облик и симметрия тела примитивных
Eumetazoa
Тела парящих и плавающих в воде, а также при-
креплённых примитивных Eumetazoa, исходно ради-
ально симметричны или диссимметричны. Но уже у
полипов группы Anthozoa имеет место билатераль-
ная симметрия (с единственной плоскостью симмет-
рии);
у всех последующих многоклеточных живот-
ных, у более высоко организованных Bilateria эта
форма симметрии становится основной.
Двуслойный уровень организации и
возникновение пищеварительной системы
Настоящая кишка возникает в результате про-
цесса гаструляции (в виде инвагинации, иммиграции
или деляминации). Сначала формируется двуслой-
экзопинакодерма
первичная сенсорная клетка
миоцитоподобная
клетка
апикальный
ленточный
контакт
эпителиально-
мышечная
клетка
нейрон
базальный
межклеточный
матрикс
мезоглеи
Илл.
187. Сравнение гистологической организации Parazoa (А) и примитивных Eumetazoa (Б).
Существенное отличие состоит в том, что у Eumetazoa клетки эпителия связаны апикальными комплексами. У некоторых Porifera (например,
пресноводных губок) пинакодерма пропускает воду непосредственно в мезохил. Связи, напоминающие zonulae adhaerentes, обнаружены
только у личинок некоторых губок между наружными жгутиковыми клетками. Миоцитоподобные клетки Porifera, погруженные в межклеточ-
ный матрикс, также отличаются от эпителиально-мышечных клеток Cnidaria, лежащих на базальной пластинке (часть мезоглеи). По Bergquist
(1978),
с изменениями.
http://jurassic.ru/

Двуслойные Eumetazoa
141
ный зародыш (гаструла) с первичными зародышевы-
ми листками — наружной эктодермой и внутренней
энтодермой. У «Coelenterata» все типы клеток про-
исходят из этих двух слоев. Из эктодермы образует-
ся эпидермис, эпителий ротовой трубки, мигрирую-
щие в мезоглею клетки и, возможно, также нервная
система. Все гастродермальные структуры образуют-
ся из энтодермы. Eumetazoa, находящиеся на этом
двуслойном уровне организации, относятся к Diplo-
blastica. Полость между двумя зародышевыми лист-
ками — бластоцель — соответствует первичной по-
лости тела Eumetazoa. У личинок и взрослых «Сое-
lenterata» она заполняется внеклеточной мезоглеей.
Иммигрирующие в мезоглею клетки называют эк-
томезенхимой, так как они происходят из наружно-
го первичного зародышевого листка.
Возникновение третьего зародышевого листка —
мезодермы — как производного энтодермы имеет место
только
у
Bilateria, которых поэтому обозначают
как
Triplo-
blastica. Вторичные микромеры у Ctenophora также при-
нято называть «мезодермальными», поскольку из них об-
разуются погружённые в мезоглею мускулатура и соеди-
нительная
ткань;
однако правомерность
этого
термина при-
менительно к гребневикам вызывает сомнения.
Уже у самых примитивных ныне живущих Eu-
metazoa («Coelenterata») имеется гастроваскулярная
полость, ограниченная эпителизованным гастродер-
мисом и связанная с внешней средой одним отвер-
стием, которое функционирует и как рот, и как анус.
На базе этой пищеварительной полости в дальней-
шем развивается пищеварительный тракт (кишка) Bi-
lateria, почти всегда имеющий отдельные ротовое и
анальное отверстия (илл. 189).
Интересно, что обе современные группы «Coelente-
rata» конвергентным путем приобрели сложные клетки,
служащие
для ловли
добычи — нематоциты
у
Cnidaria (илл.
208,
210) и коллобласты у Ctenophora (илл. 260). Благода-
ря этому они могут использовать значительно более круп-
ные
пищевые объекты (одноклеточных, мелких ракообраз-
ных, личинок Metazoa), чем губки, хоаноциты которых
улавливают преимущественно органические частицы и
бактерий размером
в
несколько микрометров. Большинство
«Coelenterata» добывают пищу пассивно, захватывая толь-
ко те объекты, которые подносятся к их щупальцам тока-
ми воды. Добыча заглатывается целиком.
Возникновение замкнутой пищеварительной
полости — гастроваскулярной системы — привело
к развитию внеклеточных механизмов пищеварения
Eumetazoa
•&
о
о.
а.
о
О
! «Coelenterata»
8
7 ?
4
_1_
2
Илл. 188. Общая схема родственных связей Metazoa.
Апоморфии: [1] Диплоидные организмы, мейоз у которых происходит при образовании гамет; ооциты с полярными телами; сперматозоиды
подразделены на головку (акросома и ядро), среднюю часть (основание жгутика, дистальная центриоль и митохондрия) и хвост (аксонема
жгутика). Имеется межклеточный матрикс (ЕСМ), состоящий из различных фибриллярных и связующих белков, прежде всего коллагенов. [2]
Тело образовано пинакодермой (наружный и внутренний покровные слои), хоанодермой, состоящей из хоаноцитов, и промежуточной соеди-
нительной тканью — мезохилом. Фильтрующие колонии с входными отверстиями (остиями), приводящими и выводящими каналами, жгути-
ковыми камерами и устьем. [3] Наличие настоящих эпителиев с апикальным клеточным комплексом. Наличие железистых клеток. [4] Покро-
вы образованы моноцилиарными клетками и дифференцированы на дорсальный и вентральный слои; последний служит для питания. Между
двумя слоями имеются звёздчатые волокнистые клетки, образующие сеть. [5] Два зародышевых листка (экто- и энтодерма), из которых обра-
зуются настоящие эпителии — наружный эпидермис и выстилающий внутреннюю пищеварительную полость гастродермис, соответственно.
В пищеварительную полость ведёт отверстие, функционирующее как рот и анус. Имеются сенсорные клетки и нейроны, часто образующие
базиэпителиальное нервное сплетение. Между сенсорными, нервными, мышечными клетками есть характерные контакты — щелевые элект-
ротонические синапсы (gap junctions), образованные белками коннексинами. Эпителиально-мышечные или миоэпителиальные клетки. Гона-
ды формируются в гастродермисе. [6] Книды (стрекательные капсулы), образующиеся в особых клетках — книдоцитах. Бесполое размноже-
ние у полипов. [7] (?) Акросома с акросомальным пузырьком и ассоциированными субакросомными структурами. [8] Парные щупальца.
Коллобласты. Двулучевая симметрия. Сложный апикальный комплекс со статоцистом. [9] Трёхслойное строение: наличие третьего зароды-
шевого листка — энтомезодермы. Билатеральная симметрия с передне-задней полярностью. Образование головного мозга на переднем конце
тела. Специальные органы выделения в виде фильтрующих почек (прото- или метанефридиев). Большинство авторов полагают, что прими-
тивные Bilateria имели только протонефридии; однако возможно, что у предковых форм уже существовали оба типа выделительных орга-
нов — протонефридии у мелких (порядка нескольких мм) псевдоцеломических личинок, а метанефридии у целомических взрослых особей.
http://jurassic.ru/
142 Двуслойные Eumetazoa
В
фагоциты
гастроваскулярная
полость
мезентерии
кишка
пластинка
нейрон
стволовая
клетка
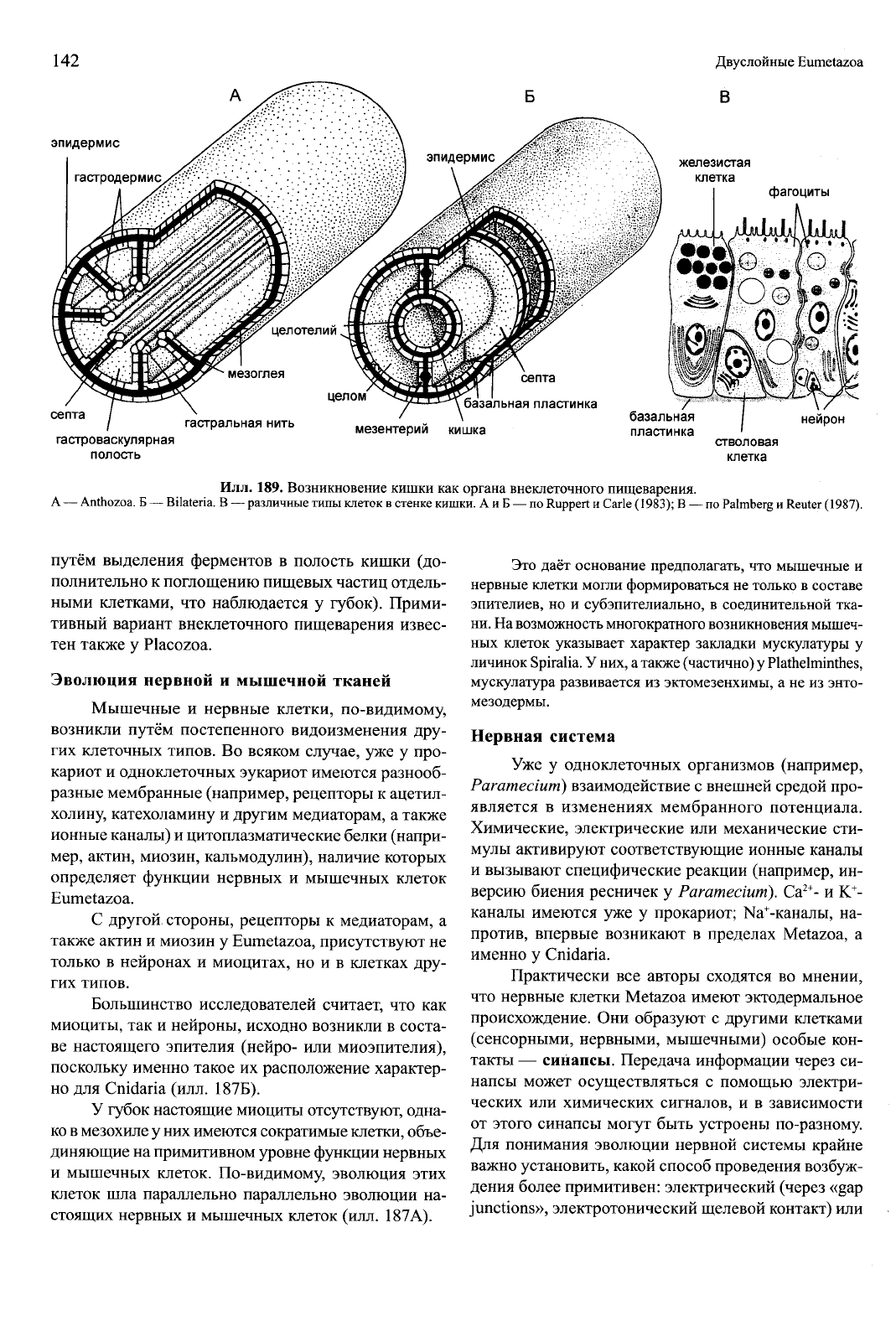
Илл.
189. Возникновение кишки как органа внеклеточного пищеварения.
А — Anthozoa. Б — Bilateria. В — различные типы клеток в стенке кишки. А и Б — по Ruppert и Carle (1983); В — по Palmberg и Reuter (1987).
путём выделения ферментов в полость кишки (до-
полнительно к поглощению пищевых частиц отдель-
ными клетками, что наблюдается у губок). Прими-
тивный вариант внеклеточного пищеварения извес-
тен также у Placozoa.
Эволюция нервной и мышечной тканей
Мышечные и нервные клетки, по-видимому,
возникли путём постепенного видоизменения дру-
гих клеточных типов. Во всяком случае, уже у про-
кариот и одноклеточных эукариот имеются разнооб-
разные мембранные (например, рецепторы к ацетил-
холину, катехоламину и другим медиаторам, а также
ионные каналы) и цитоплазматические белки (напри-
мер,
актин, миозин, кальмодулин), наличие которых
определяет функции нервных и мышечных клеток
Eumetazoa.
С другой стороны, рецепторы к медиаторам, а
также актин и миозин у Eumetazoa, присутствуют не
только в нейронах и миоцитах, но и в клетках дру-
гих типов.
Большинство исследователей считает, что как
миоциты, так и нейроны, исходно возникли в соста-
ве настоящего эпителия (нейро- или миоэпителия),
поскольку именно такое их расположение характер-
но для Cnidaria (илл. 187Б).
У губок настоящие миоциты отсутствуют, одна-
ко в мезохиле у них имеются сократимые клетки, объе-
диняющие на примитивном уровне функции нервных
и мышечных клеток. По-видимому, эволюция этих
клеток шла параллельно параллельно эволюции на-
стоящих нервных и мышечных клеток (илл. 187А).
Это даёт основание предполагать, что мышечные и
нервные клетки могли формироваться не только в составе
эпителиев, но и субэпителиально, в соединительной тка-
ни.
На возможность многократного возникновения мышеч-
ных клеток указывает характер закладки мускулатуры у
личинок Spiralia. У них,
а
также (частично) у Plathelminthes,
мускулатура развивается из эктомезенхимы, а не из энто-
мезодермы.
Нервная система
Уже у одноклеточных организмов (например,
Paramecium) взаимодействие с внешней средой про-
является в изменениях мембранного потенциала.
Химические, электрические или механические сти-
мулы активируют соответствующие ионные каналы
и вызывают специфические реакции (например, ин-
версию биения ресничек у Paramecium). Са
2+
- и К
+
-
каналы имеются уже у прокариот; Ка
+
-каналы, на-
против, впервые возникают в пределах Metazoa, а
именно у Cnidaria.
Практически все авторы сходятся во мнении,
что нервные клетки Metazoa имеют эктодермальное
происхождение. Они образуют с другими клетками
(сенсорными, нервными, мышечными) особые кон-
такты — синапсы. Передача информации через си-
напсы может осуществляться с помощью электри-
ческих или химических сигналов, и в зависимости
от этого синапсы могут быть устроены по-разному.
Для понимания эволюции нервной системы крайне
важно установить, какой способ проведения возбуж-
дения более примитивен: электрический (через «gap
junctions», электротонический щелевой контакт) или
эпидермис
гастродермис
http://jurassic.ru/
Двуслойные Eumetazoa
143
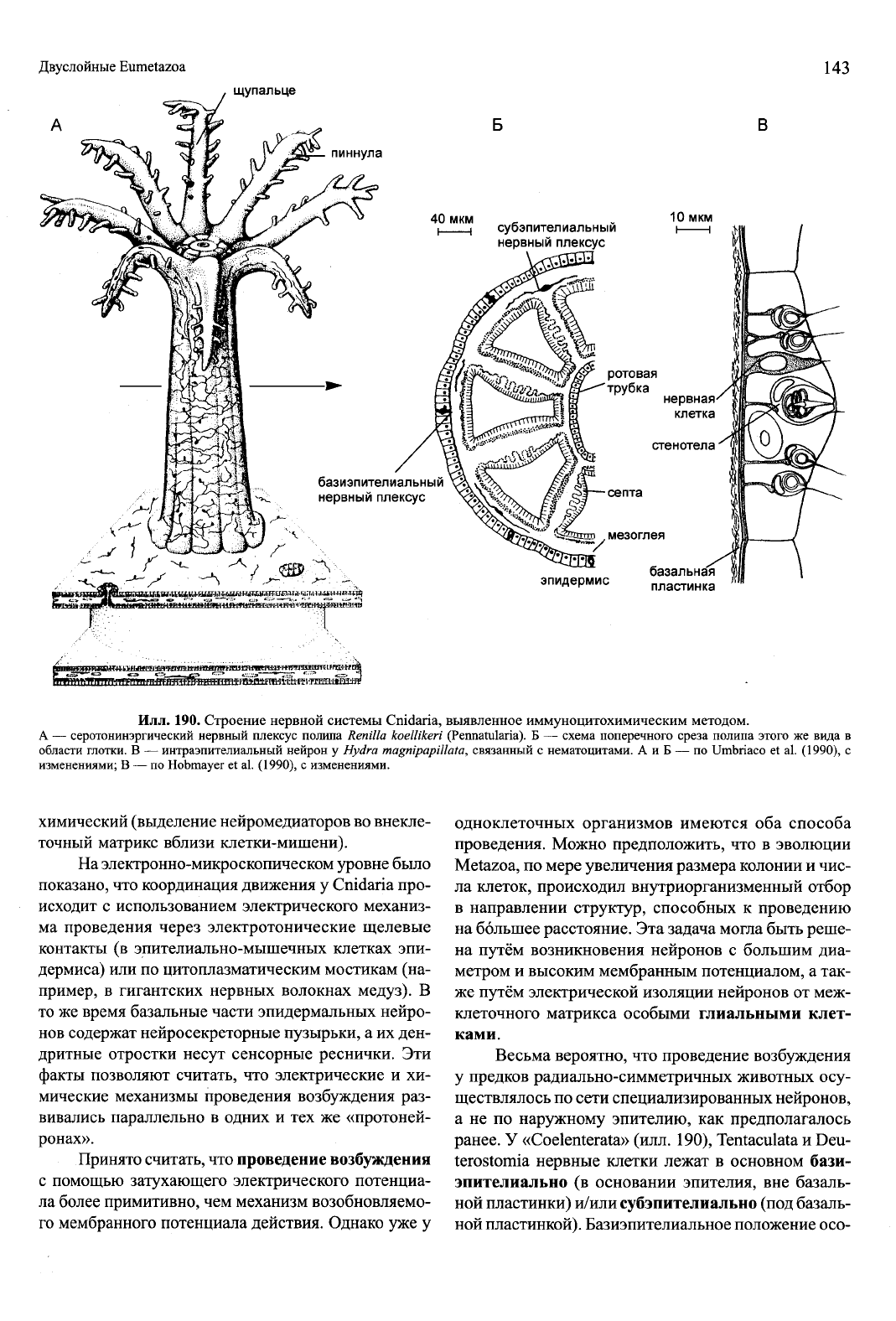
Илл.
190. Строение нервной системы Cnidaria, выявленное иммуноцитохимическим методом.
А — серотонинэргический нервный плексус полипа Renilla koellikeri (Pennatularia). Б — схема поперечного среза полипа этого же вида в
области глотки. В — интраэпителиальный нейрон у Hydra magnipapUlata, связанный с нематоцитами. А и Б — по Umbriaco et al. (1990), с
изменениями; В — по Hobmayer et al. (1990), с изменениями.
химический (выделение нейромедиаторов во внекле-
точный матрикс вблизи клетки-мишени).
На электронно-микроскопическом уровне было
показано, что координация движения у Cnidaria про-
исходит с использованием электрического механиз-
ма проведения через электротонические щелевые
контакты (в эпителиально-мышечных клетках эпи-
дермиса) или по цитоплазматическим мостикам (на-
пример, в гигантских нервных волокнах медуз). В
то же время базальные части эпидермальных нейро-
нов содержат нейросекреторные пузырьки, а их ден-
дритные отростки несут сенсорные реснички. Эти
факты позволяют считать, что электрические и хи-
мические механизмы проведения возбуждения раз-
вивались параллельно в одних и тех же «протоней-
ронах».
Принято считать, что проведение возбуждения
с помощью затухающего электрического потенциа-
ла более примитивно, чем механизм возобновляемо-
го мембранного потенциала действия. Однако уже у
одноклеточных организмов имеются оба способа
проведения. Можно предположить, что в эволюции
Metazoa, по мере увеличения размера колонии и чис-
ла клеток, происходил внутриорганизменный отбор
в направлении структур, способных к проведению
на большее расстояние. Эта задача могла быть реше-
на путём возникновения нейронов с большим диа-
метром и высоким мембранным потенциалом, а так-
же путём электрической изоляции нейронов от меж-
клеточного матрикса особыми глиальными клет-
ками.
Весьма вероятно, что проведение возбуждения
у предков радиально-симметричных животных осу-
ществлялось по сети специализированных нейронов,
а не по наружному эпителию, как предполагалось
ранее. У «Coelenterata» (илл. 190), Tentaculata и Deu-
terostomia нервные клетки лежат в основном бази-
эпителиально (в основании эпителия, вне базаль-
ной пластинки) и/или субэпителиально (под базаль-
ной пластинкой). Базиэпителиальное положение осо-
http://jurassic.ru/
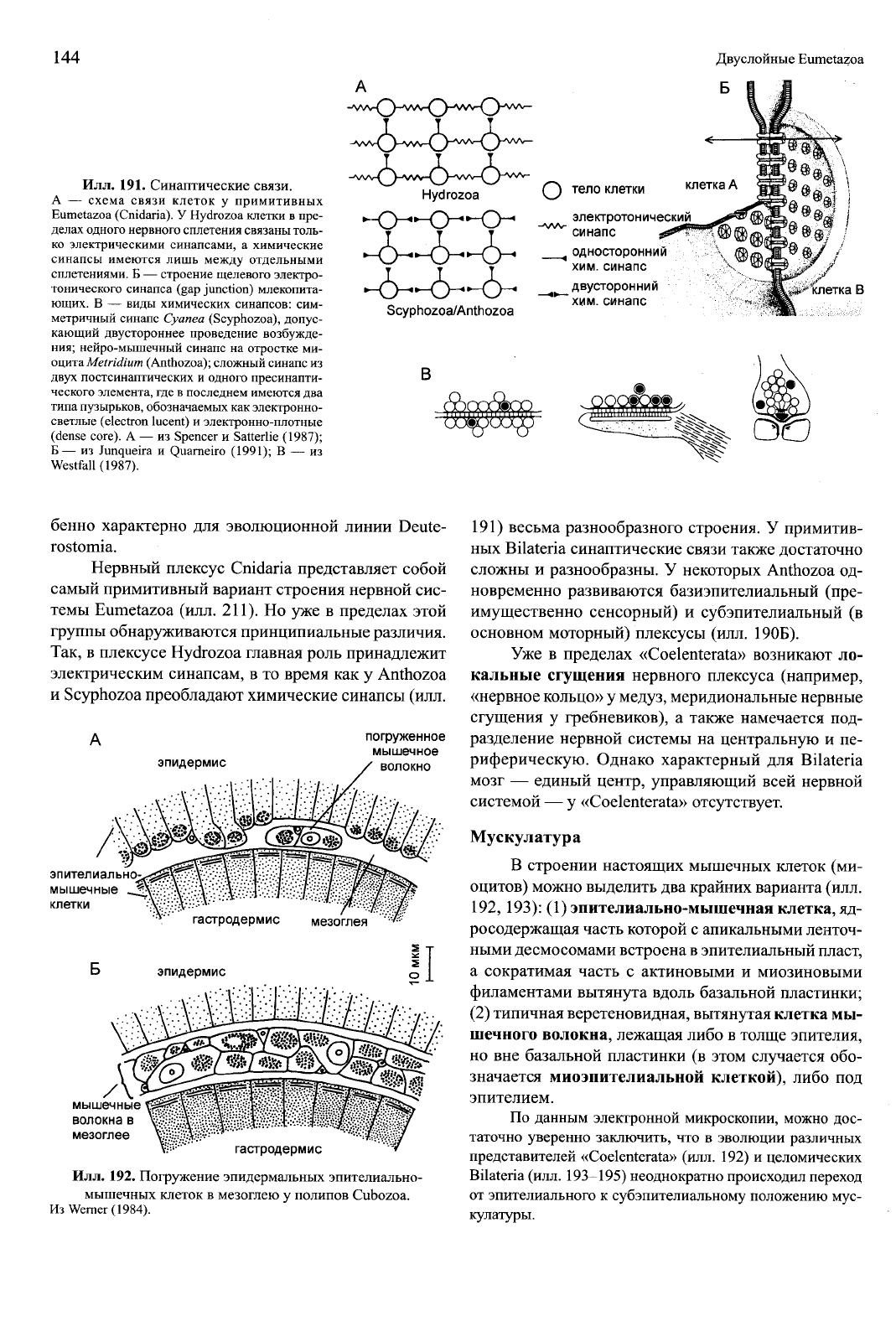
144
Двуслойные Eumetazoa
Илл.
191. Синаптические связи.
А — схема связи клеток у примитивных
Eumetazoa (Cnidaria). У Hydrozoa клетки в пре-
делах одного нервного сплетения связаны толь-
ко электрическими синапсами, а химические
синапсы имеются лишь между отдельными
сплетениями. Б — строение щелевого электро-
тонического синапса (gap junction) млекопита-
ющих. В — виды химических синапсов: сим-
метричный синапс Cyanea (Scyphozoa), допус-
кающий двустороннее проведение возбужде-
ния;
нейро-мышечный синапс на отростке ми-
oaynaMetridium (Anthozoa); сложный синапс из
двух постсинаптических и одного пресинапти-
ческого элемента, где в последнем имеются два
типа пузырьков, обозначаемых как электронно-
светлые (electron lucent) и электронно-плотные
(dense core). А — из Spencer и Satterlie (1987);
Б — из Junqueira и Quarneiro (1991); В — из
Westfall (1987).
Hydrozoa
-6—6—нб-
Scyphozoa/Anthozoa
О
тело
клетки
электротоническии
синапс
!
односторонний
хим.
синапс
двусторонний
"хим.синапс
клетка
В
бенно характерно для эволюционной линии Deute-
rostomia.
Нервный плексус Cnidaria представляет собой
самый примитивный вариант строения нервной сис-
темы Eumetazoa (илл. 211). Но уже в пределах этой
группы обнаруживаются принципиальные различия.
Так, в плексусе Hydrozoa главная роль принадлежит
электрическим синапсам, в то время как у Anthozoa
и Scyphozoa преобладают химические синапсы (илл.
Илл.
192. Погружение эпидермальных эпителиально-
мышечных клеток в мезоглею у полипов Cubozoa.
Из Werner (1984).
191) весьма разнообразного строения. У примитив-
ных Bilateria синаптические связи также достаточно
сложны и разнообразны. У некоторых Anthozoa од-
новременно развиваются базиэпителиальный (пре-
имущественно сенсорный) и субэпителиальный (в
основном моторный) плексусы (илл. 190Б).
Уже в пределах «Coelenterata» возникают ло-
кальные сгущения нервного плексуса (например,
«нервное кольцо» у медуз, меридиональные нервные
сгущения у гребневиков), а также намечается под-
разделение нервной системы на центральную и пе-
риферическую. Однако характерный для Bilateria
мозг — единый центр, управляющий всей нервной
системой — у «Coelenterata» отсутствует.
Мускулатура
В строении настоящих мышечных клеток (ми-
оцитов) можно выделить два крайних варианта (илл.
192,
193): (1) эпителиально-мышечная клетка, яд-
росодержащая часть которой с апикальными ленточ-
ными десмосомами встроена в эпителиальный пласт,
а сократимая часть с актиновыми и миозиновыми
филаментами вытянута вдоль базальной пластинки;
(2) типичная веретеновидная, вытянутая клетка мы-
шечного волокна, лежащая либо в толще эпителия,
но вне базальной пластинки (в этом случается обо-
значается миоэпителиальной клеткой), либо под
эпителием.
По данным электронной микроскопии, можно дос-
таточно уверенно заключить, что в эволюции различных
представителей «Coelenterata» (илл. 192) и целомических
Bilateria (илл. 193-195) неоднократно происходил переход
от эпителиального к субэпителиальному положению мус-
кулатуры.
http://jurassic.ru/
Двуслойные Eumetazoa
145
эпителиально-мышечная
клетка
•
i
Mr
•
...
•
1
•
>
1,1,
моноцилиарная
перитонеальная
-о^^
клетка
миоэпителиальная
клетка
"\базальная
пластинка
ядро
соединительная
ткань
ядро
«
5 мкм
субперитонеальное
мышечное
волокно
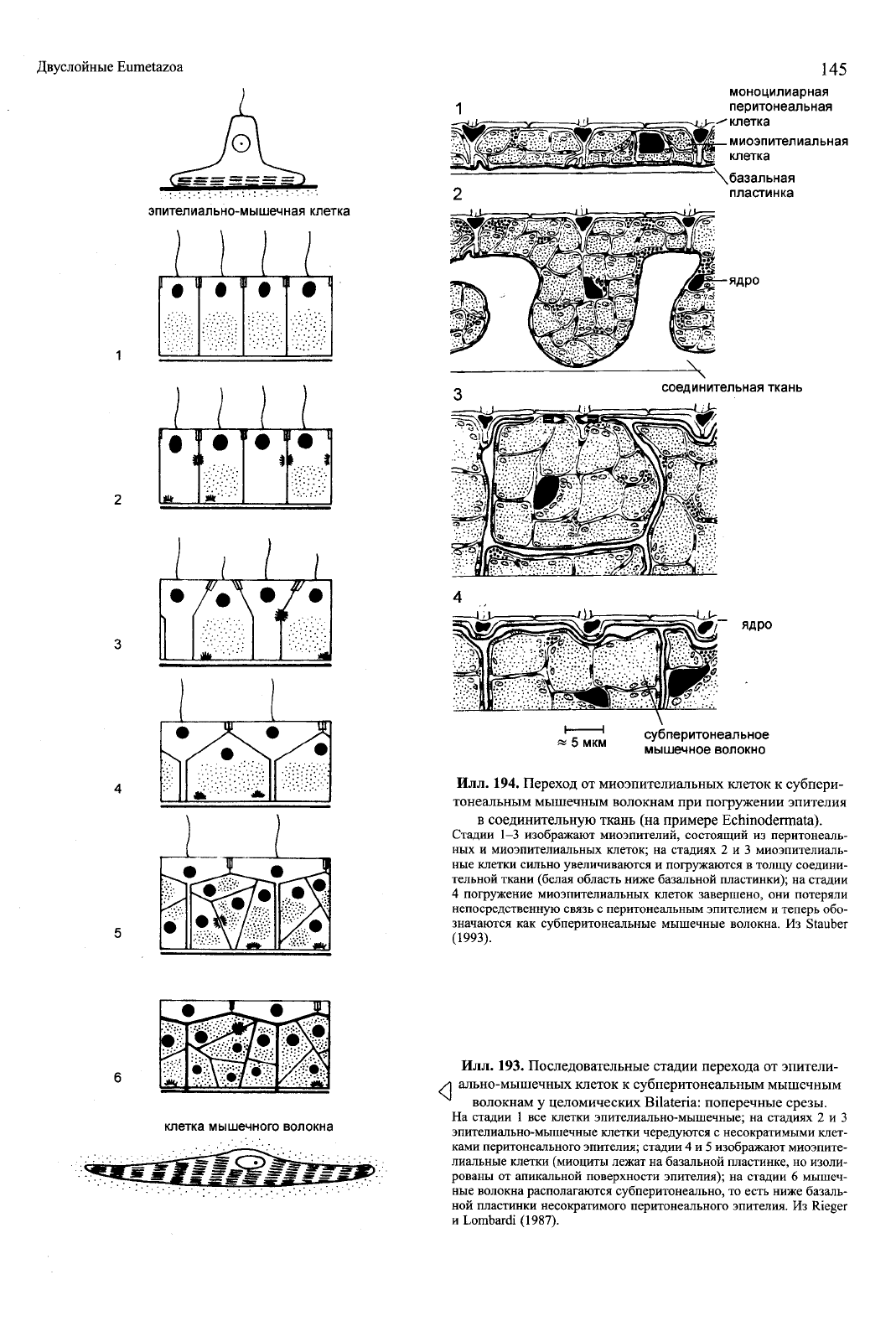
Илл. 194. Переход от миоэпителиальных клеток к субпери-
тонеальным мышечным волокнам при погружении эпителия
в соединительную ткань (на примере Echinodermata).
Стадии 1-3 изображают миоэпителий, состоящий из перитонеаль-
ных и миоэпителиальных клеток; на стадиях 2 и 3 миоэпителиаль-
ные клетки сильно увеличиваются и погружаются в толщу соедини-
тельной ткани (белая область ниже базальной пластинки); на стадии
4 погружение миоэпителиальных клеток завершено, они потеряли
непосредственную связь с перитонеальным эпителием и теперь обо-
значаются как субперитонеальные мышечные волокна. Из Stauber
(1993).
клетка
мышечного волокна
<3
Илл. 193. Последовательные стадии перехода от эпители-
ально-мышечных клеток к субперитонеальным мышечным
волокнам у целомических Bilateria: поперечные срезы.
На стадии 1 все клетки эпителиально-мышечные; на стадиях 2 и 3
эпителиально-мышечные клетки чередуются с несократимыми клет-
ками перитонеального эпителия; стадии 4 и 5 изображают миоэпите-
лиальные клетки (миоциты лежат на базальной пластинке, но изоли-
рованы от апикальной поверхности эпителия); на стадии 6 мышеч-
ные волокна располагаются субперитонеально, то есть ниже базаль-
ной пластинки несократимого перитонеального эпителия. Из Rieger
и Lombardi (1987).
http://jurassic.ru/