Вестхайде В.,Ригер Р.( ред.) Зоология беспозвоночных в двух томах. Том 1: от простейших до моллюсков и артропод
Подождите немного. Документ загружается.

96
Metazoa
основание реснички
микроворсинки
с
гл
и
ко
кал иксом
"ножка"
zonula
adhaerens
передний (ростральный)
корешок
дополнительная
центриоль
септальная десмосома
диктиосома
задний (каудальный)
корешок
ядро
1 мкм
базиэпителиальные
аксоны
базальная
ламина
\фиброретику-
лярная
ламина
базальный
матрикс
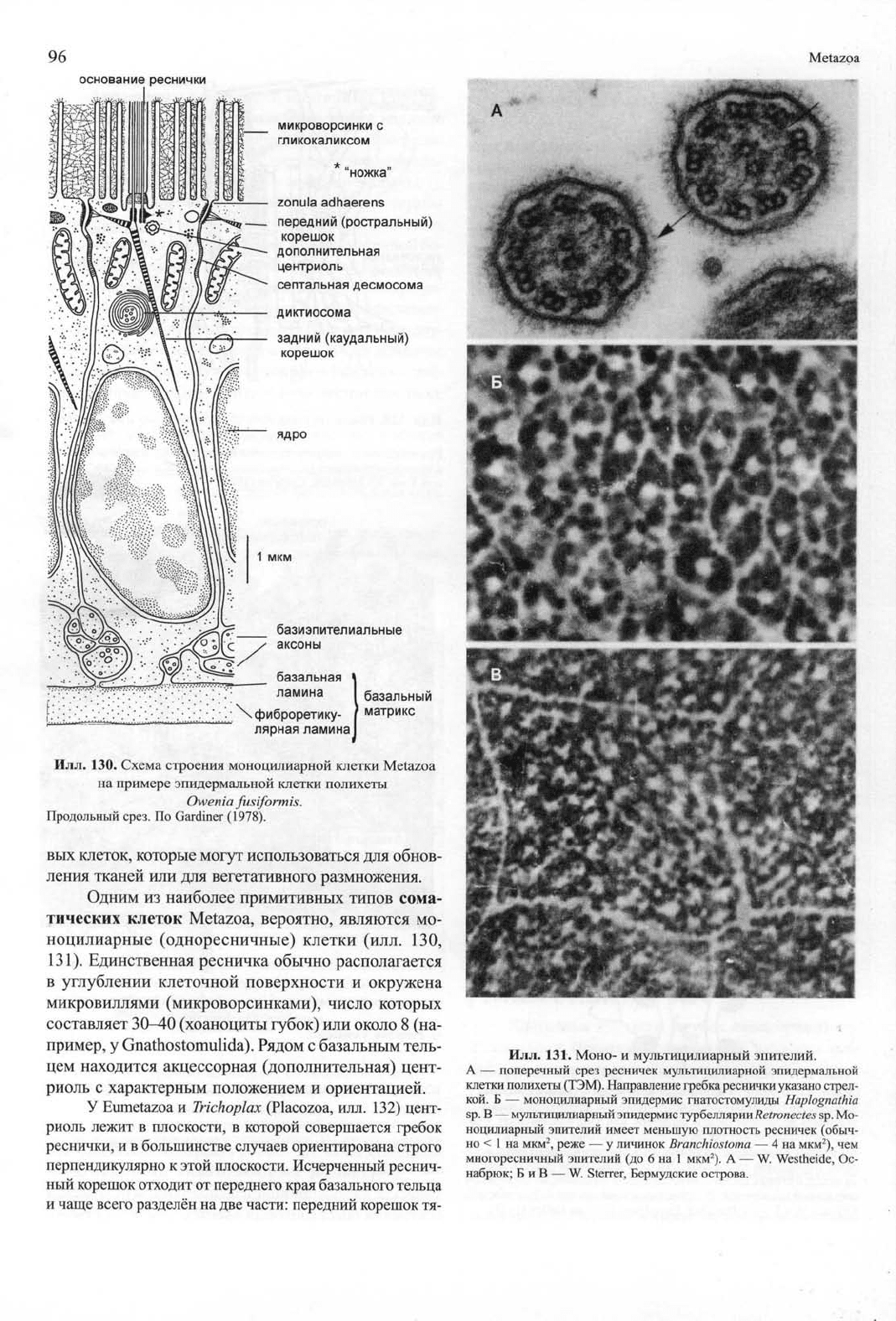
Илл.
130. Схема строения моноцилиарной клетки Metazoa
на примере эпидермальной клетки полихеты
Owenia fusiformis.
Продольный срез. По Gardiner
(1978).
вых клеток, которые могут использоваться для обнов-
ления тканей или для вегетативного размножения.
Одним из наиболее примитивных типов сома-
тических клеток Metazoa, вероятно, являются мо-
ноцилиарные (одноресничные) клетки (илл. 130,
131).
Единственная ресничка обычно располагается
в углублении клеточной поверхности и окружена
микровиллями (микроворсинками), число которых
составляет ЗО^Ю (хоаноциты губок) или около 8 (на-
пример, у Gnathostomulida). Рядом с базальным тель-
цем находится акцессорная (дополнительная) цент-
риоль с характерным положением и ориентацией.
У Eumetazoa и
Trichoplax
(Placozoa, илл. 132) цент-
риоль лежит в плоскости, в которой совершается гребок
реснички, и в большинстве случаев ориентирована строго
перпендикулярно к этой плоскости. Исчерченный реснич-
ный корешок отходит от переднего края базального тельца
и чаще всего разделён на две части: передний корешок тя-
Илл.
131. Моно- и мультицилиарный эпителий.
А — поперечный срез ресничек мультицилиарной эпидермальной
клетки полихеты (ТЭМ). Направление гребка реснички указано стрел-
кой.
Б — моноцилиарный эпидермис гнатостомулиды Haplognathia
sp.
В — мультицилиарный эпидермис турбеллярии Retronectes sp. Мо-
ноцилиарный эпителий имеет меньшую плотность ресничек (обыч-
но < 1 на мкм
2
, реже — у личинок Branchiostoma — 4 на мкм
2
), чем
многоресничный эпителий (до 6 на 1 мкм
2
). А — W. Westheide, Ос-
набрюк; Б и В — W. Sterrer, Бермудские острова.
http://jurassic.ru/
Metazoa
97
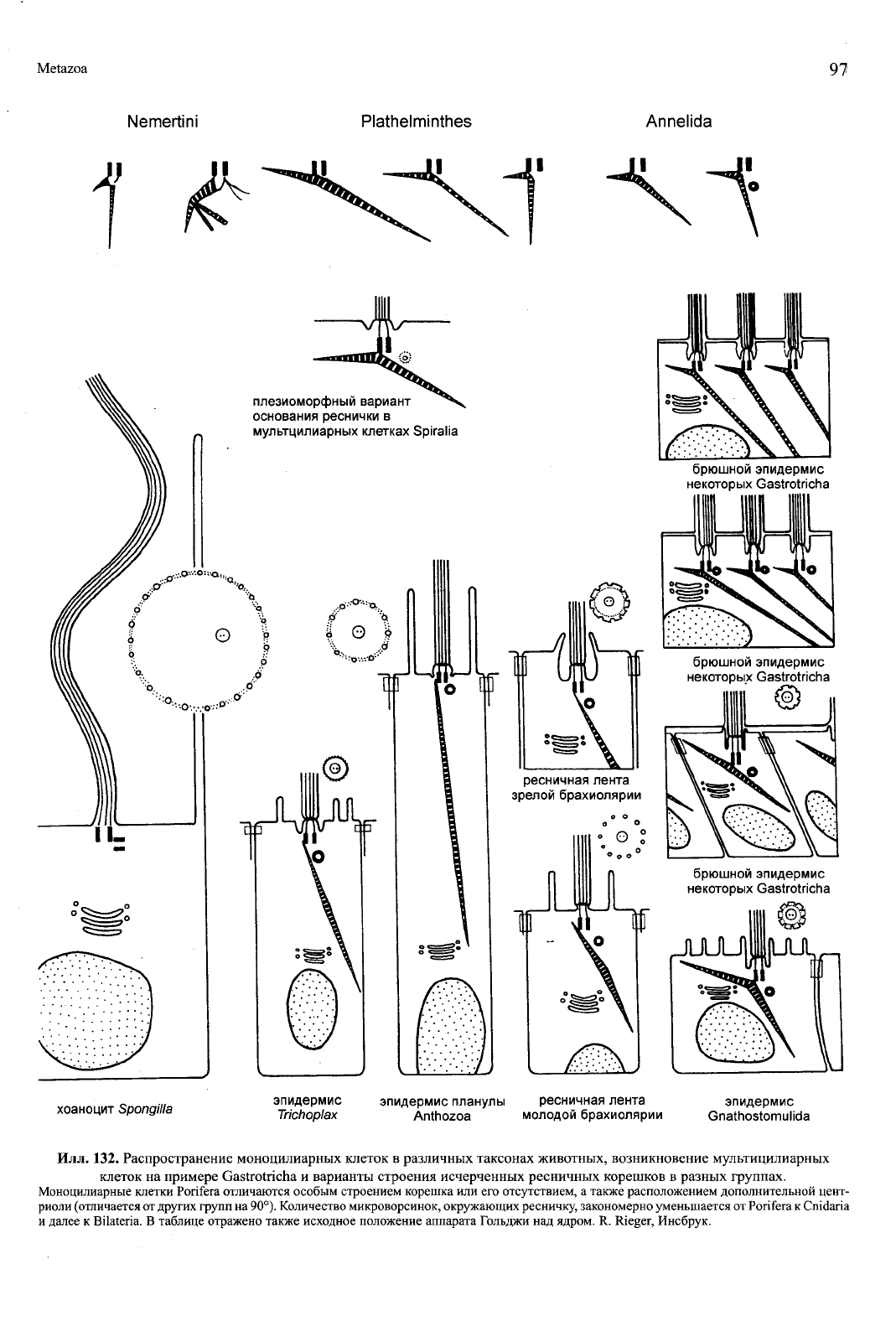
Илл.
132. Распространение моноцилиарных клеток в различных таксонах животных, возникновение мультицилиарных
клеток на примере Gastrotricha и варианты строения исчерченных ресничных корешков в разных группах.
Моноцилиарные клетки Porifera отличаются особым строением корешка или его отсутствием, а также расположением дополнительной цент-
риоли (отличается от других групп на 90°). Количество микроворсинок, окружающих ресничку, закономерно уменьшается от Porifera к Cnidaria
и далее к Bilateria. В таблице отражено также исходное положение аппарата Гольджи над ядром. R. Rieger, Инсбрук.
http://jurassic.ru/

98
Metazoa
нется приблизительно горизонтально под клеточной мем-
браной, а задний направлен в глубь клетки. В примитив-
ном варианте корешок, вероятно, достигал ядра клетки.
Между ядром и основанием реснички лежит аппарат Голь-
джи.
Базальное тело имеет вырост — «ножку» (basal foot,
см.
илл. 130) на стороне, соответствующей направлению
гребка. Взаимную ориентацию ресничек в большинстве
случаев можно установить по поперечным срезам (илл.
131
А).
Строение сенсорных клеток (так наз. воротничко-
вых рецепторов, илл. 196А) и терминальных клеток про-
тонефридиев (илл. 280) может отклоняться от общей схе-
мы;
в частности, может исчезать ресничный корешок или
дополнительная центриоль.
У личинок несинцитиальных губок (Cellularia) вен-
чик микровиллей вокруг реснички отсутствует; напротив,
у хоаноцитов взрослых губок микровилли очень длинные.
Дополнительная центриоль у губок, если она есть, «повёр-
нута» на 90° по сравнению с Eumetazoa и
Trichoplax
(илл.
132).
Исчерченный корешок может отсутствовать, а иног-
да вместо него имеется система микротрубочек.
Мультицилиарный (многоресничный) эпителий
возникал на основе моноцилианого неоднократно в
пределах Metazoa (илл. 131, 132). Следует заметить,
что между этими двумя состояниями нет плавного
перехода: в большинстве случаев имеется либо одна
ресничка на клетку, либо десять или больше, а, на-
пример, клетки с двумя ресничками встречаются
крайне редко. Исключение составляют только сен-
сорные клетки, в которых число ресничек может ва-
рьировать от одной до нескольких. Мультицилиар-
ные эпителиальные клетки почти всегда утрачивают
дополнительные центриоли (исключения из этого
правила есть среди Cnidaria, Gastrotricha и личинок
кольчатых червей).
Только моноцилинарые клетки имеются у Placozoa,
Porifera (хотя недавно у личинки Hexactinellida были най-
дены мультицилиарные клетки), Gnathostomulida, а также,
по-видимому, у всех Phoronida, Brachiopoda, Pterobranchia,
Echinodermata и Acrania. Эпителии Cnidaria также преиму-
щественно моноцилиарные. Среди Gastrotricha и полихет
группы Oweniida известны формы с моно- и мультицили-
арным эпителием. Одновременное присутствие моно- и
мультицилиарных клеток у одного вида отмечается редко,
например у
Renilla
(Anthozoa) и у торнарий (личинок
Enteropneusta).
На эмбриологическом материале показано, что
одноресничная клетка является исходной стадией
развития клеток соединительной ткани, миоцитов и
нейронов Eumetazoa.
Metazoa — диплоидные организмы, мейоз у
которых происходит при образовании гамет. Харак-
терно, что из мужского гоноцита при мейозе образу-
ются четыре сперматозоида, а из женского — только
одна яйцеклетка и три полярных тельца. У Porifera
женские гаметы образуются из хоаноцитов или ар-
хеоцитов, а мужские, по-видимому, только из хоано-
цитов. Половые клетки Eumetazoa исходно возника-
ют в энтодерме или в производной от неё энтомезо-
дерме; только у некоторых Hydrozoa гаметы, возмож-
но,
имеют эктодермальное происхождение. Для дру-
гих представителей Hydrozoa было показано, что
яйцеклетки у них формируются в гастродермисе и
лишь затем перемещаются в эпидермис.
У губок в примитивном случае гоноциты сво-
бодно лежат в мезохиле или в эпителиальной ткани
и не связаны с какими-либо вспомогательными клет-
ками. Однако уже в пределах Porifera и примитив-
ных Bilateria (например, Plathelminthes) многие фор-
мы имеют специализированные соматические клет-
ки (или модифицированные половые клетки), кото-
рые обеспечивают питание гоноцитов и/или отгра-
ничивают их от соматических тканей. У более высо-
коорганизованных форм такие клетки могут участво-
вать в определении осей тела эмбриона (например, в
развитии насекомых — см. ниже).
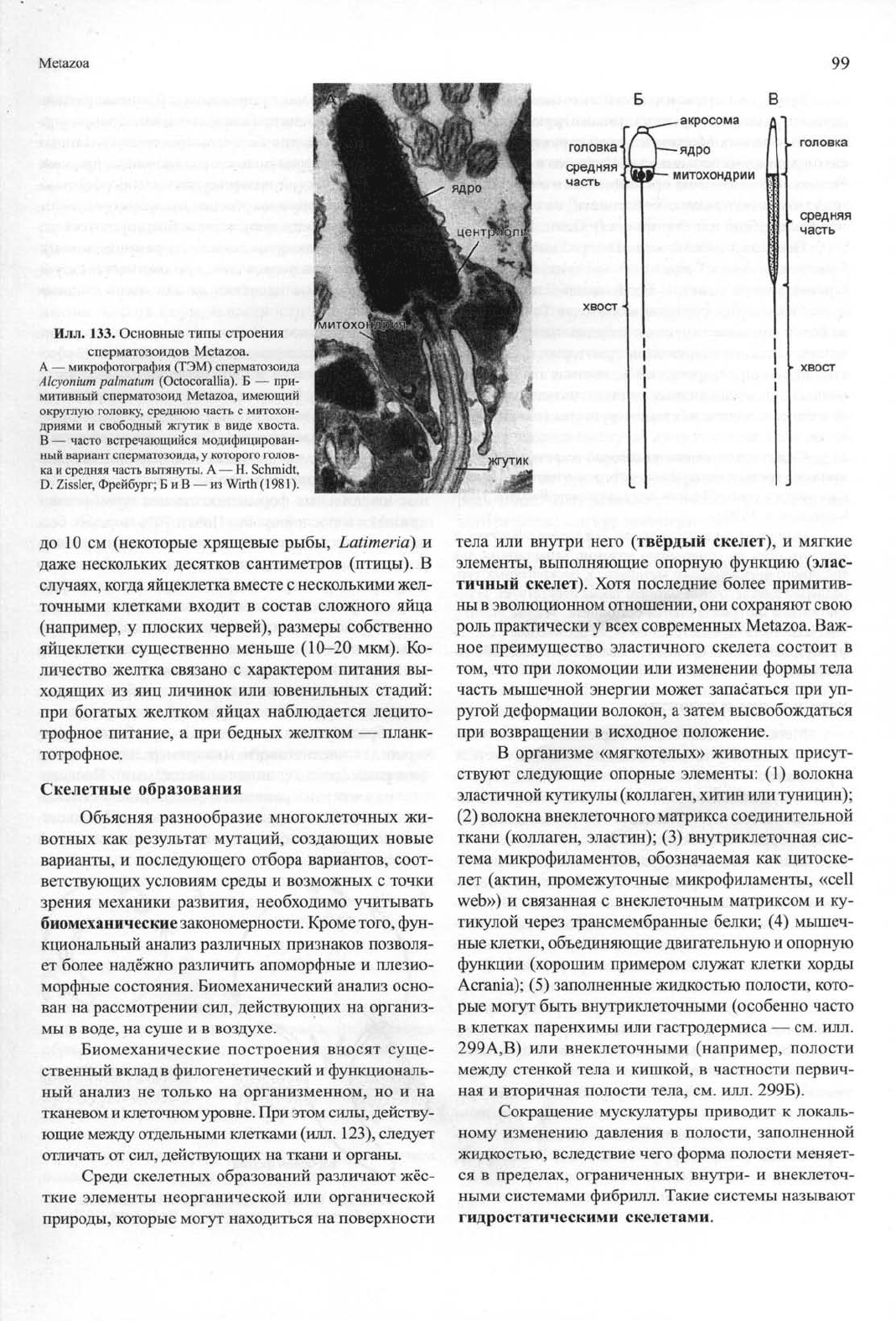
Строение мужских гамет (спермиев, или спер-
матозоидов) всех Metazoa можно вывести из одного
исходного типа (илл. 133). Примитивный спермий
имеет округлую головку, в которой находятся ядро и
расположенная перед ним акросома. Позади голов-
ки имеется небольшая средняя часть с митохондри-
ей,
а между ядром и митохондрией лежит базальное
тело жгутика, который формирует хвост спермия. В
примитивном состоянии имеется и дополнительная
центриоль, что соответствует исходному типу строе-
ния моноцилиарной клетки.
На этой основе могут возникать очень разно-
образные модификации, которые особенно характер-
ны для групп, имеющих внутреннее оплодотворение
(илл. 133В). По морфологическому разнообразию
спермин Metazoa превосходят все прочие типы кле-
ток. Группы спермиев часто бывают упакованы в
разного рода внеклеточные оболочки — спермато-
форы.
Женские гаметы (яйцеклетки) Metazoa разли-
чаются по запасам желтка и характеру его распреде-
ления внутри клетки, а также по полярности. Широ-
ко распространён вариант, при котором желток от-
кладывается в цитоплазме самого ооцита; такие яйца
называются эндолецитальными. В других случаях
ооцит содержит немного желтка или вообще лишён
его,
а основной запас желтка накапливается в осо-
бых желточных клетках — вителлоцитах (экзолеци-
тальные яйца, например, у Plathelminthes). Форми-
рование желтка может происходить только в самом
ооците (аутосинтетически) или же — частично или
полностью — вне его (гетеросинтетически).
Размеры эндолецитальных яиц варьируют от 50
мкм (у некоторых Porifera, Cnidaria, Tentaculata и от-
дельных представителей Spiralia и Nemathelminthes)
http://jurassic.ru/
Metazoa
99
Илл.
133. Основные типы строения
сперматозоидов Metazoa.
А — микрофотография (ТЭМ) сперматозоида
Alcyonium palmatum (Octocorallia). Б — при-
митивный сперматозоид Metazoa, имеющий
округлую головку, среднюю часть с митохон-
дриями и свободный жгутик в виде хвоста.
В — часто встречающийся модифицирован-
ный вариант сперматозоида, у которого голов-
ка и средняя часть вытянуты. А — Н. Schmidt,
D.
Zissler, Фрейбург; Б и В — из Wirth (1981).
В
средняя
часть
хвост
акросома
ядро
митохондрии
средняя
часть
хвост
до 10 см (некоторые хрящевые рыбы, Latimeria) и
даже нескольких десятков сантиметров (птицы). В
случаях, когда яйцеклетка вместе с несколькими жел-
точными клетками входит в состав сложного яйца
(например, у плоских червей), размеры собственно
яйцеклетки существенно меньше (10-20 мкм). Ко-
личество желтка связано с характером питания вы-
ходящих из яиц личинок или ювенильных стадий:
при богатых желтком яйцах наблюдается лецито-
трофное питание, а при бедных желтком — планк-
тотрофное.
Скелетные образования
Объясняя разнообразие многоклеточных жи-
вотных как результат мутаций, создающих новые
варианты, и последующего отбора вариантов, соот-
ветствующих условиям среды и возможных с точки
зрения механики развития, необходимо учитывать
биомеханические закономерности. Кроме того, фун-
кциональный анализ различных признаков позволя-
ет более надёжно различить апоморфные и плезио-
морфные состояния. Биомеханический анализ осно-
ван на рассмотрении сил, действующих на организ-
мы в воде, на суше и в воздухе.
Биомеханические построения вносят суще-
ственный вклад в филогенетический и функциональ-
ный анализ не только на организменном, но и на
тканевом и клеточном уровне. При этом силы, действу-
ющие между отдельными клетками (илл. 123), следует
отличать от сил, действующих на ткани и органы.
Среди скелетных образований различают жёс-
ткие элементы неорганической или органической
природы, которые могут находиться на поверхности
тела или внутри него (твёрдый скелет), и мягкие
элементы, выполняющие опорную функцию (элас-
тичный скелет). Хотя последние более примитив-
ны в эволюционном отношении, они сохраняют свою
роль практически у всех современных Metazoa. Важ-
ное преимущество эластичного скелета состоит в
том, что при локомоции или изменении формы тела
часть мышечной энергии может запасаться при уп-
ругой деформации волокон, а затем высвобождаться
при возвращении в исходное положение.
В организме «мягкотелых» животных присут-
ствуют следующие опорные элементы: (1) волокна
эластичной кутикулы (коллаген, хитин или туницин);
(2) волокна внеклеточного матрикса соединительной
ткани (коллаген, эластин); (3) внутриклеточная сис-
тема микрофиламентов, обозначаемая как цитоске-
лет (актин, промежуточные микрофиламенты, «cell
web») и связанная с внеклеточным матриксом и ку-
тикулой через трансмембранные белки; (4) мышеч-
ные клетки, объединяющие двигательную и опорную
функции (хорошим примером служат клетки хорды
Acrania); (5) заполненные жидкостью полости, кото-
рые могут быть внутриклеточными (особенно часто
в клетках паренхимы или гастродермиса — см. илл.
299А,В) или внеклеточными (например, полости
между стенкой тела и кишкой, в частности первич-
ная и вторичная полости тела, см. илл. 299Б).
Сокращение мускулатуры приводит к локаль-
ному изменению давления в полости, заполненной
жидкостью, вследствие чего форма полости меняет-
ся в пределах, ограниченных внутри- и внеклеточ-
ными системами фибрилл. Такие системы называют
гидростатическими скелетами.
http://jurassic.ru/

100
Metazoa
Примером органического экзоскелета могут
служить различные кутикулярные структуры. Сре-
ди примитивных Metazoa к таким структурам отно-
сится перидерма большинства Hydrozoa и некоторых
Octocorallia. Элементы органического эндоскелета
могут быть построены, в частности, из спонгина (в
мезохиле губок) или горгонина (у Octocorallia).
Неорганический, или минеральный, скелет
может состоять из отдельных мелких склеритов
(кремнезёмные спикулы Porifera, известковые спи-
кулы Octocorallia, спикулы в оболочке Tunicata) или
из более или менее крупных твёрдых элементов (на-
пример, известковые скелеты некоторых губок, пла-
стинки панциря морских ежей, костная ткань позво-
ночных). Отдельные спикулы также могут соединять-
ся в более крупные жёсткие структуры (например, у
Hexactinellida).
Скелеты, состоящие из спикул, встречаются среди
наиболее древних ископаемых остатков животных, имею-
щих возраст около 550 млн. лет (например, Porifera, fHal-
kieriida, илл. 487В).
Особое положение занимает
Trichoplax
adhaerens,
у
которого пока не обнаружено волокон, характерных для
типичного внеклеточного матрикса. У бескишечных тур-
беллярий внеклеточный матрикс также отсутствует, за ис-
ключением материала стенки статоциста. Усилие мышц у
этих животных передаётся либо через десмосомы и систе-
му
внутриклеточных промежуточных
филаментов,
либо
че-
рез ресничные корешки.
Размножение и развитие
Metazoa — диплоидные организмы. Это озна-
чает, что мейоз у них происходит только в клетках
зародышевого пути, непосредственно перед образо-
ванием мужских и женских гамет. В жизненном цик-
ле Metazoa бесполое размножение (почкование, де-
ление, образование бесполых бродяжек, архитомия,
паратомия) может чередоваться с половым. Способ-
ность к бесполому размножению более характерна
для примитивных представителей Metazoa. Чередо-
вание одного или нескольких поколений, размножа-
ющихся бесполым путем, с половым поколением
называется метагенезом (например, у Cnidaria).
Развитие потомства из неоплодотворённых яй-
цеклеток — партеногенез — широко распростране-
но в животном мире, в том числе и среди позвоноч-
ных (костистые рыбы, рептилии). Известны различ-
ные механизмы модификации мейоза при партено-
генезе, вплоть до полной его утраты; последний ва-
риант обозначается как апомиксис. Смена двуполо-
го и партеногенетического поколений называется ге-
терогонией (например, у «Rotatoria», Anomopoda,
Aphidina).
Колониальная организация и бесполое размно-
жение свойственны прежде всего примитивным фор-
мам. Напротив, для более высокоорганизованных
Metazoa характерно половое размножение, приводя-
щее к генетической индивидуальности особей в по-
пуляции. Партеногенез обеспечивает быстрое увели-
чение численности популяции в благоприятных ус-
ловиях. Как и в случае бесполого размножения пу-
тём деления или почкования, при партеногенезе об-
разуются особи с идентичным или очень сходным
генотипом.
В жизненном цикле многих животных встре-
чается особая ювенильная форма (личинка), кото-
рая морфологически и биологически отличается от
взрослой особи. Превращение личинки во взрослую
форму может сопровождаться кардинальной пере-
стройкой (метаморфозом). Этот вариант обознача-
ется как непрямое развитие и противопоставляет-
ся прямому развитию, при котором последователь-
ные ювенильные формы постепенно приобретают
признаки взрослой особи. Почти 70% морских бес-
позвоночных имеют жизненный цикл, в котором мик-
роскопическая планктонная личинка даёт начало
макроскопической, часто бентосной взрослой фор-
ме (двухфазный пелаго-бентосный цикл, см. илл.
134).
Прямое развитие обычно считают примитив-
ным состоянием для Metazoa, хотя некоторые авто-
ры придерживаются другого мнения.
У некоторых животных постэмбриональное
развитие сопровождается линькой, т.е., периодичес-
ким сбрасыванием кутикулы. Это особенно харак-
терно для членистоногих (например, науплиус или
зоэа ракообразных, личинки насекомых). Последо-
вательные стадии развития в этом случае часто обо-
значаются как личинки разного возраста. Число сег-
ментов тела у последовательных личиночных стадий
Илл.
134.
Схема двухфазного жизненного цикла.
А— пелаго-бентосный. Б — пелагический. По Jagersten (1972).
http://jurassic.ru/

Metazoa
101
может постепенно увеличиваться (анаморфоз, на-
пример, у Lithobiomorpha). У других форм выходя-
щая из яйца личинка уже имеет такое же число сег-
ментов, что и взрослое животное (эпиморфоз, на-
пример, у Scolopendromorpha).
С точки зрения эволюционной биологии, боль-
шой интерес представляет возникновение новых ви-
дов путем прогенеза, то есть перехода личинок или
ювенильных стадий к половому размножению. Это
явление, по-видимому, особенно характерно для эво-
люции мелких форм, в частности представителей
интерстициальной фауны. Прогенез представляет
собой частный случай гетерохронии, под которой
понимают как ускорение, так и задержку развития
одних органов по сравнению с другими. Гетерохро-
ния очень часто наблюдается в развитии Metazoa и
имеет большое значение для их эволюции. Сохране-
ние личиночных или ювенильных признаков у бо-
лее поздних стадий, вплоть до половозрелых особей,
обозначается как неотения.
Принято считать, что примитивные Metazoa
были раздельнополыми животными с наружным
оплодотворением. Эти признаки обычно сочетают-
ся с производством большого числа гамет и наличи-
ем разного рода механизмов синхронизации вымета
гамет особями обоих полов (см. Annelida). Однако
уже у Porifera, за немногими исключениями, вымет
яйцеклеток происходит только после внутреннего
оплодотворения.
Напротив, гермафродитизм обычно рассмат-
ривают как вторичное явление, связанное с малыми
размерами тела, сидячим образом жизни и низкой
численностью. Различают одновременный и после-
довательный гермафродитизм. В первом случае в
теле животного одновременно функционируют муж-
ские и женские половые органы (например, у Plathel-
minthes, многих Gastropoda, Hirudinea, Tunicata); во
втором случае происходит смена пола, т.е. в каждый
момент времени в теле животного имеются гоноци-
ты и половые органы только одного пола (например,
у некоторых брюхоногих моллюсков, полихеты
Ophryotrocha puerilis, многих рыб). У большинства
гермафродитов вначале развиваются мужские поло-
вые клетки, а затем женские (протерандрия). Реже,
например у некоторых оболочников, наблюдается
обратная последовательность (протерогиния).
Оплодотворение у Eumetazoa чаще всего про-
исходит перед первым делением созревания ооцита.
Полярные тельца часто маркируют положение ани-
мального полюса зиготы. Оплодотворение обычно
сопровождается радикальным перераспределением
ооплазмы. Этот процесс крайне важен для дальней-
шего развития, так как распределение морфогенети-
ческих «факторов детерминации» в цитоплазме зи-
готы определяет попадание их в тот или иной блас-
томер при дроблении.
Оплодотворённая яйцеклетка (зигота) Metazoa
проявляет отчётливую анимально-вегетативную по-
лярность, что выражается в распределении различ-
ных органелл. Деления дробления начинаются обыч-
но в области анимального полюса и распространя-
ются на более богатый желтком вегетативный полюс.
Первые три деления дробления обычно сле-
дуют единой схеме: первых два деления (приводя-
щие,
соответственно, к стадии 2 и 4 бластомеров)
проходят меридионально от анимального к вегета-
тивному полюсу, а третье — экваториально, перпен-
дикулярно к оси яйца. Отклонения от этой схемы из-
вестны в разных группах животных.
Дробление может быть полным или непол-
ным;
в последнем случае из-за большого количества
желтка в зиготе бластомеры не полностью отделя-
ются друг от друга. Полное дробление, в свою оче-
редь,
может быть равномерным (на бластомеры рав-
ного размера) или неравномерным (на макро- и мик-
ромеры). Вариантами неполного дробления являют-
ся дискоидальное (например, Cephalopoda, Sauropsi-
da, некоторые Teleostei) и поверхностное дробление
(Arthropoda).
В начале дробления митозы протекают одно-
временно во всех бластомерах (синхронная фаза), а
затем синхронность нарушается (асинхронная фаза).
Число синхронных делений неодинаково в разных
таксонах животных.
Яйцеклетка в ходе дробления обычно не рас-
тёт, и, соответственно, размеры отдельных бласто-
меров уменьшаются. Завершение дробления означа-
ет переход к стадии бластулы. При наличии блас-
тоцеля (первичной полости тела) последнюю назы-
вают целобластула, а при его отсутствии — стерро-
бластула. Особые случаи — это амфибластулы изве-
стковых губок, несущие реснички только на анималь-
ном полюсе, дискобластулы и перибластулы, возни-
кающие соответственно в результате дискоидально-
го и поверхностного дробления, а также бластоцис-
ты Eutheria.
В зависимости от положения веретён деления
бластомеров различают радиальное дробление (ве-
ретёна деления расположены параллельно и перпен-
дикулярно к оси яйца) и спиральное дробление (ве-
ретёна наклонены попеременно влево или вправо к
оси яйца). По симметрии развивающегося эмбриона
можно различить двулучевое (Ctenophora) и билате-
ральное (примитивные Chordata) дробление. Особые
формы дробления встречаются у Nemathelminthes. Во
многих группах (например, Mollusca, Arthropoda и
др.) наблюдаются отклонения от исходных вариан-
http://jurassic.ru/

102 Metazoa
тов дробления, часто связанные с повышенным со-
держанием желтка в яйце.
В зависимости от того, на каком этапе разви-
тия можно установить соответствие между отдель-
ными бластомерами и клетками дефинитивных ор-
ганов, многие авторы различают два варианта раз-
вития — детерминированное (мозаичное) и регуля-
торное. Первый вариант чаще наблюдают у форм с
двулучевым, спиральным или билатеральным дроб-
лением. Для зародышей со спиральным (Plathelmin-
thes,
Mollusca, Annelida, илл. 288), билатеральным
(Tunicata) или модифицированным билатеральным
дроблением (Nematoda) удалось проследить «родос-
ловную» потомков каждого отдельного бластомера.
Регуляторный тип развития обычно встречается у
форм с радиальным дроблением — Cnidaria, Tenta-
culata и Echinodermata.
Необходимо заметить, однако, что ранняя «детерми-
нированность» судьбы того или иного бластомера может
быть обнаружена
только
экспериментально. Название «мо-
заичный зародыш», разумеется, не означает, что в цитоп-
лазме неоплодотворенного ооцита имеются участки, пря-
мо соответствующие органам взрослого животного.
Интересно, что уже у примитивных Metazoa (неко-
торые известковые губки, различные Hydrozoa) известны
модифицированные процессы развития. У губок вначале
возникает стомобластула со жгутиками, обращенными
внутрь, из которой путем выворачивания (экскурвации)
образуется амфибластула. Интересно, что точно такой же
процесс имеется у колониальной зелёной водоросли
Volvox
(см.
ниже), но не у других Metazoa. Нерегулярное распо-
ложение бластомеров
в
раннем зародыше Hydrozoa (так наз.
«анархия бластомеров») обычно рассматривают как ано-
малию, вызванную неблагоприятными внешними услови-
ями.
Однако можно предположить, что это явление отра-
жает общую лабильность расположения бластомеров. Дру-
гие представители Cnidaria имеют «ложное спиральное»
дробление.
Решающим этапом в развитии Eumetazoa явля-
ется гаструляция, приводящая к формированию
двух первичных зародышевых листков — эктодер-
мы и энтодермы (илл. 135). Этот морфогенетичес-
кий процесс осуществляется благодаря движению
отдельных клеток или целых клеточных пластов.
Различают инвагинационную гаструлу, которая мо-
жет формироваться путем эпиболии (обрастания) или
эмболии (впячивания), и иммиграционную гастру-
лу, в которой эпителий первичной кишки вторично
«собирается» из отдельных клеток. Иммиграция кле-
ток в последнем случае может происходить на од-
ном или нескольких «полюсах» бластулы.
Перемещения отдельных клеток или клеточных
слоев у губок (илл. 138, 158), которые начинаются
после оседания личинки и приводят к так называе-
мому извращению зародышевых листков (наружные
жгутиковые клетки оказываются внутри, а внутрен-
ние — снаружи), сравнивают с процессом гаструля-
ции Eumetazoa (илл. 135). Однако при метаморфозе
амфибластулы выворачивание жгутикового пласта
происходит на полюсе, противоположном тому, где
начинается гаструляция Cnidaria. Проблема гомоло-
гизации гаструляции Eumetazoa и морфогенетичес-
ких событий в развитии Porifera пока остаётся нере-
шённой.
С собственно гаструляцией тесно связано об-
разование третьего зародышевого листка — настоя-
щей мезодермы (энтомезодермы), а также выселе-
ние клеток эктодермального происхождения (экто-
мезенхимы) в пространство между двумя первичны-
ми зародышевыми листками или во внеклеточный
матрикс бластоцеля. Клетки, заселяющие мезоглею
у различных представителей Cnidaria, также имеют
эктодермальную природу. У Ctenophora микромеры
вегетативного полюса дают начало, помимо других
структур, в том числе и мышечным клеткам, погру-
жённым в мезоглею.
Таким образом, гаструляция является началь-
ным этапом формирования крайне стабильных, кон-
сервативных стадий развития, обозначаемых как
филотипические стадии, или зоотипы. Филотипичес-
кие стадии обнаруживают в развитии всех предста-
вителей крупной таксономической группы (напри-
мер,
стадия фарингулы для позвоночных животных).
Симбиоз с прокариотами и одноклеточными
эукариотами
В эволюции Metazoa широко распространено
явление симбиоза с прокариотами, а также однокле-
точными (или нитчатыми многоклеточными) эука-
риотными организмами. Примерами могут служить
симбиозы губок с бактериями и сине-зелёными во-
дорослями, симбиозы актиний и коралловых поли-
пов — с динофлагеллятами, симбиозы нематод и
олигохет, обитающих в микроаэробных и анаэроб-
ных песчаных грунтах, — с хемоавтотрофными бак-
териями. Особые бактерии находятся и в светящих-
ся органах головоногих моллюсков и огнетелок, и в
организме погонофор и других глубоководных мор-
ских организмах (хемоавтотрофы), и в пищеваритель-
ном тракте более высокоорганизованных животных,
включая человека. В случае облигатных симбиотичес-
ких отношений, симбионты либо переносятся вмес-
те с яйцеклеткой, либо каким-то образом (с пищей и
т.п.) попадают в организм молодого животного.
Хорошо известна роль симбиотических связей
в возникновении эукариотической клетки (появление
митохондрий и пластид). Разнообразие вариантов
симбиоза между Metazoa, прокариотами и эукариот-
http://jurassic.ru/
Metazoa
103
ными одноклеточными еще раз подчеркивает значе-
ние этого явления в эволюции организмов.
Систематика
Результаты анализа последовательностей 18S
рРНК, ультраструктурные признаки моноцилиарных
клеток, а также структура и характер развития гамет
и двухфазный жизненный цикл (илл. 188) свидетель-
ствуют о монофилетическом происхождении Metazoa
в целом. Однако главным аргументом в пользу мо-
нофилии Metazoa, по-видимому, может служить вы-
сокая степень сходства молекулярного состава вне-
клеточного матрикса у Porifera и Eumetazoa.
Распространённое мнение о том, что сестринс-
кой группой Metazoa являются одноклеточные Choa-
noflagellata, не очень хорошо согласуется с имеющи-
мися данными. Хоаноциты Porifera (рис. 137Б) дей-
ствительно обнаруживают ультраструктурное сход-
ство с клетками Choanoflagellata, но частные гомо-
логии между их отдельными структурами нельзя счи-
тать доказанными.
Parazoa и Placozoa в настоящее время рассмат-
ривают как самые примитивные группы среди ныне
живущих Metazoa (рис. 188) в силу таких признаков,
как отсутствие нервной и мышечной тканей, а также
настоящих эпителиев.
Porifera отчётливо отделяются от других Me-
tazoa как по анатомическим признакам взрослых гу-
бок, так и по характеру развития. Их особое положе-
ние подтверждается также и на ультраструктурном
уровне: жгутики губок в большинстве случаев фик-
http://jurassic.ru/

104
Metazoa
сируются характерно расположенными микротрубоч-
ками, в то время как исчерченные ресничные кореш-
ки найдены только у личинок известковых губок.
Дополнительная центриоль в большинстве случаев
либо отсутствует, либо расположена не так, как у
других Metazoa.
В «эпителии» Placozoa имеются апикальные
клеточные контакты, напоминающие ленточные дес-
мосомы; на основании этого признака Trichoplax ad-
haerens можно считать сестринской группой по от-
ношению ко всем остальным Eumetazoa. Однако
включение Placozoa в Eumetazoa невозможно, по-
скольку у них пока не найдены ни типичный внекле-
точный матрикс, ни мышечные или нервные клетки.
В пользу примитивности Placozoa говорит также то,
что их ядра имеют самое низкое содержание ДНК
среди всех Metazoa.
«Mesozoa», по-видимому, не представляют со-
бой монофилетическую группу. Единственный при-
знак, объединяющий явно различные группы Ortho-
nectida и Dicyemida, это наличие мультицилиарных
покровных клеток, окружающих внутреннее скопле-
ние клеток, в котором формируются гаметы. Поло-
жение «Mesozoa» в пределах Metazoa по-прежнему
составляет предмет дискуссии. Часто их рассматри-
вают как группу Plathelminthes, упрощённых вслед-
ствие паразитического образа жизни. Эту точку зре-
ния подтверждает наличие миоцитоподобных клеток
у Orthonectida, но она плохо согласуется с молеку-
лярно-биологическими и ультраструктурными дан-
ными по Dicyemida. Следует заметить, однако, что
имеются и другие примеры крайнего упрощения
организации у паразитических групп (Myxozoa).
Монофилия Eumetazoa может быть обоснова-
на на гистологическом уровне наличием истинных
эпителиев, а также нервных и мышечных клеток. Её
подтверждает наличие гомологичной пищеваритель-
ной полости, а также процесс гаструляции и образо-
вание первичных зародышевых листков — экто- и
энтодермы. В связи с этим вызывает удивление тот
факт, что анализ последовательностей 28S рРНК не
подтвержает монофилию Eumetazoa, а указывает ско-
рее на отделение Bilateria ещё до разделения осталь-
ных Metazoa на Placozoa, Porifera и «Coelenterata».
http://jurassic.ru/

I.
Parazoa
Губки, единственная группа этой ступени орга-
низации, могут обозначаться как «почти Metazoa».
На основании резко различного гистологического
строения несколько авторов разделяет губки на два
субтаксона, преимущественно клеточных Cellularia
и синцитиальных Symplasma (илл. 144). Обсужда-
ют даже полифилию губок, то есть независимое про-
исхождение обоих таксонов от одноклеточных пред-
ков.
Хотя предполагают, что многоядерная тканевая
организация Symplasma возникает посредством не-
полных клеточных делений и таким образом должна
обозначаться «плазмодиальной», в литературе уко-
ренился термин «синцитий», который предполагает
слияние уже образовавшихся отдельных клеток.
У Porifera внутренняя среда тела не так сильно
отделена от окружающей воды, поскольку у них, в
отличие от Eumetazoa, нет настоящих плотных эпи-
телиев с апикальными опоясывающими межклеточ-
ными контактами.
Reinhard Rieger, Инсбрук
Правда, у некоторых губок из Cellularia выяв-
лены предшественники настоящих мышечных кле-
ток — сократимые клетки с толстыми и тонкими фи-
ламентами, образующие оскулярные сфинктеры.
Однако скорость распространения сокращения этих
клеток ограничена (максимально 1 мм/мин на дис-
танции в несколько сантиметров). Эти «миоциты»
можно рассматривать как «независимые эффектор-
ные клетки» (восприятие раздражения, проведение
возбуждения и эффекторная реакция в одной и той
же клетке). Проведение возбуждения от одной клет-
ки к соседней может осуществляться через десмосо-
моподобные контакты между клетками; при этом на
мембранах последовательно открываются ионные
каналы, в результате чего осуществляется движение
ионов в клетке. Так или иначе, этот процесс может
быть не вполне идентичен аналогичным процессам
у примитивных Eumetazoa (Cnidaria, Ctenophora).
Регистрация в некоторых клетках ацетилхоли-
нэстеразы, катехоламина или серотонина также ещё
не свидетельствует о существовании у губок настоя-
щей нервной системы, поскольку нет указаний на
участие этих агентов в проведении нейрональных
сигналов.
Илл.
136. Модель организации губки
Demospongiae.
Облик личинки губки (А), взрослой губки (Б) и
схемы их поперечных срезов (В — личинка; Г —
взрослая губка, изображена только одна жгути-
ковая камера). Клеточные ядра в покровных сло-
ях клеток показаны чёрным. А, Б — из Riedl
(1970);
В — оригинал S. Weyrer, Инсбрук; Г —
из Bergquist (1978).
http://jurassic.ru/