Смирнов В.М. Физиология человека: Учебник
Подождите немного. Документ загружается.


ческих волокон блуждающего нерва, анасто-
мозирующих с веточками звездчатого ган-
глия. Усиление сокращений желудка и
кишки при раздражении симпатического
нерва осуществляется с помощью серотонин-
ергических нервных волокон, идущих в со-
ставе симпатического ствола.
На основании приведенных фактов можно
заключить, что один и тот же нерв с помо-
щью одного и того же выделяемого им меди-
атора, действующего на одни и те же рецеп-
торы, не может оказывать двоякого влияния
на деятельность органа.
3.1.3.
РЕФЛЕКТОРНЫЙ ПРИНЦИП НЕРВНОЙ
РЕГУЛЯЦИИ
Рефлекс — ответная реакция организма на
раздражение сенсорных рецепторов, осу-
ществляемая с помощью нервной системы.
Каждый рефлекс осуществляется посредст-
вом рефлекторной дуги.
А. Рефлекторная дуга — это совокупность
структур, при помощи которых осуществля-
ется рефлекс. Схематично рефлекторную
дугу вегетативного и соматического рефлек-
сов можно представить состоящей из пяти
звеньев (рис. 3.1).
1.
Рецептор предназначен для восприятия
изменений внешней или внутренней среды
организма, что достигается посредством
трансформации энергии раздражения в нерв-
ный импульс. Совокупность рецепторов, раз-
дражение которых вызывает рефлекс, называ-
ют рефлексогенной зоной. Конкретные меха-
низмы восприятия раздражителя (света,
звука, изменения химического состава крови)
различны у каждого вида рецепторов, но во
всех случаях обеспечивается посылка нервно-
го импульса в центральную нервную систему.
2.
Афферентный
путь, передающий сигнал
в ЦНС. Для соматической нервной системы
это афферентный нейрон с его отростками, те-
ло его расположено в спинномозговых гангли-
ях или ганглиях черепных нервов. Импульс от
рецептора поступает на дендрит афферентного
нейрона, а по его аксону
—
в ЦНС.
3.
Вставочные нейроны ЦНС. В составе ве-
гетативной нервной системы вставочные ней-
роны могут находиться вне ЦНС — интра-
и(или) экстраорганно. Их назначение
—
обес-
печение связи с другими отделами ЦНС,
переработка и передача импульсов к эффек-
торному нейрону (рис. 3.2).
4.
Эффекторный нейрон. Для соматичес-
кой нервной системы это мотонейрон. Его
назначение — вместе с другими нейронами
ЦНС переработать информацию, сформиро-
Рис. 3.1. Рефлекторная дуга: А — соматического,
Б
—
вегетативного рефлексов.
1
—
рецептор; 2
—
афферентное звено; 3
—
центральное
звено; 4
—
эфферентное звено; 5
—
эффектор.
вать ответ в виде нервных импульсов, посы-
лаемых к 5-му звену рефлекторной дуги —
эффектору (рабочему органу). В простейшем
случае рефлекторная дуга может быть двух-
нейронной — без вставочного нейрона.
Б.
Классификацию рефлексов проводят по
нескольким критериям. В частности, по сро-
кам появления рефлексов в онтогенезе их
делят на две группы: врожденные (безуслов-
ные) и приобретенные (условные). Врожден-
ные (безусловные) рефлексы делят также на
несколько групп.
1.
В
зависимости
от
числа синапсов
в цент-
ральной части рефлекторной дуги различают
моно-
и полисинаптические рефлексы. Приме-
Кора большого
мозга
Промежуточный
мозг
Средний
мозг
Продолговатый
мозг
Спинной
мозг
Рис. 3.2. Структура дуги безусловного рефлекса
(по Асратяну).
1—5
—
ветви центральной части дуги на разных уровнях;
А —
афферентная клетка; Б
—
мотонейрон.
31

ром моносинаптического рефлекса может
служить рефлекс на растяжение четырехгла-
вой мышцы
—
коленный разгибательный реф-
лекс,
возникающий при ударе по сухожилию
ниже коленной чашечки. Большинство реф-
лексов являются полисинаптическими, в их
осуществлении участвует несколько последо-
вательно включенных нейронов ЦНС. От
числа нейронов, участвующих в осуществле-
нии рефлекса, зависит время рефлекса
—
дли-
тельность интервала от начала стимуляции до
окончания ответной реакции. Оно включает:
1) время трансформации энергии раздраже-
ния в распространяющийся импульс, что осу-
ществляется с помощью рецептора; 2) время
проведения возбуждения в афферентном
пути, в центральной части рефлекторной дуги
и в эфферентном пути; 3) время активации
эффектора и его ответной реакции.
2.
По биологическому значению рефлексы
делят на пищедобывательные, половые, за-
щитные (оборонительные), исследователь-
ские,
родительские. Примером исследова-
тельского рефлекса является ориентировоч-
ный рефлекс (рефлекс «что такое?») — пово-
рот головы, глаз, туловища в сторону внезап-
но появившегося раздражителя.
3.
По рецепторам, раздражение которых
вызывает ответную реакцию, различают экс-
тероцептивные, интероцептивные и про-
приоцептивные рефлексы. Они используются
в клинической практике для оценки состоя-
ния центральной нервной системы (ЦНС).
4.
По локализации рефлекторной дуги раз-
личают центральные рефлексы (дуга которых
проходит через ЦНС) и периферические реф-
лексы, дуга которых замыкается вне ЦНС.
Последними могут быть только вегетативные
рефлексы, они подразделяются на интраор-
ганные и экстраорганные (в последнем случае
рефлекс осуществляется с участием нейронов
экстраорганных ганглиев). Выделяют также
межорганные рефлексы, которые могут быть
периферическими и центральными. Напри-
мер,
механическое раздражение желудка
может затормозить сердечную деятельность.
5. В зависимости от отдела нервной сис-
темы выделяют соматические и вегетатив-
ные рефлексы.
3.1.4. РАЗВИТИЕ КОНЦЕПЦИИ РЕФЛЕКСА.
НЕРВИЗМ И НЕРВНЫЙ ЦЕНТР
А. Основные положения рефлекторного прин-
ципа деятельности ЦНС были разработаны
на протяжении примерно двух с половиной
веков. Можно назвать пять основных этапов
развития этой концепции.
Первый этап
—
заложены основы понима-
ния рефлекторного принципа деятельности
ЦНС французским естествоиспытателем и
математиком Р.Декартом (XVII в.). Р.Декарт
считал, что «все вещи и явления можно объ-
яснить естественнонаучным путем». Эта ис-
ходная позиция позволила Р.Декарту сфор-
мулировать два важных положения рефлек-
торной теории: 1) деятельность организма
при внешнем воздействии является отражен-
ной (впоследствии ее стали называть рефлек-
торной: лат. reflexus — отраженный), 2) от-
ветная реакция на раздражение осуществля-
ется при помощи нервной системы. По Р.Де-
карту нервы — это трубочки, по которым с
огромной скоростью движутся животные
духи, материальные частицы неизвестной
природы, по нервам они попадают в мышцу
и мышца раздувается (сокращается).
Второй этап — экспериментальное обо-
снование материалистических представлений
о рефлексе (XVII—XIX вв.). В частности,
было установлено, что рефлекторная реакция
может осуществляться на одном метамере ля-
гушки (сегмент спинного мозга, связанный с
«кусочком тела»). Выявлено, что стимуль
могут быть не только внешними, но и внут
ренними, установлена роль задних (чувстви
тельных) и передних (двигательных) кореш
ков спинного мозга (закон Белла—Мажан
ди).
Весьма активно сегментарные рефлексь
изучал Ч.Шеррингтон (конец XVIII в. — на
чало XIX в.).
Третий этап — победа материалистичес
ких представлений о психической деятель
ности (И.М.Сеченов, 60-е годы XIX в.). На
блюдая за развитием детей, И.М.Сечено
пришел к заключению, что в основе форми
рования психической деятельности лежи
принцип рефлекса. Это положение он вырг
зил следующей фразой: «Все акты сознател]
ной и бессознательной жизни по способ
происхождения суть рефлексы». Таким обр!
зом, И.М.Сеченов стал на путь детерминиз\
в вопросах психической деятельности чел»
века. При изучении рефлексов он обоснов.
приспособительный характер изменчивое
рефлекса, открыл торможение рефлекс»
(1863,
центральное торможение), суммаци
возбуждения в ЦНС (1868).
Четвертый этап — разработаны OCHOI
учения о высшей нервной деятельное
(И.П.Павлов, начало XX в.). И.П.Павлов с
крыл условные рефлексы и использовал
как объективный метод изучения психич*
кой деятельности (высшей нервной деяте.1
ности, по И.П.Павлову). Он сформулироЕ
три принципа рефлекторной теории: 1) пру
32

цип детерминизма (принцип причинности),
согласно которому любая рефлекторная ре-
акция причинно обусловлена; 2) принцип
структурности, суть которого заключается в
том, что каждая рефлекторная реакция осу-
ществляется с помощью определенных
структур, и чем больше структурных элемен-
тов участвует в осуществлении этой реакции,
тем она совершеннее; 3) принцип единства
процессов
анализа и синтеза в составе рефлек-
торной реакции: нервная система анализиру-
ет (различает) с помощью рецепторов все
действующие внешние и внутренние раздра-
жители и на основании этого анализа форми-
рует целостную ответную реакцию (синтез).
Пятый этап — создано учение о функци-
ональных системах (П.К.Анохин, середина
XX в.).
Б.
Нервизм — это концепция, признаю-
щая ведущую роль нервной системы в регу-
ляции функций всех органов и тканей орга-
низма (физиологический нервизм). Концеп-
ция нервизма прошла весьма длинный исто-
рический путь развития.
Почву для этой концепции подготовил Р.Де-
карт (1596—1650), выдвинувший идею о рефлек-
торном принципе деятельности нервной системы.
Ф.Гофман (1660—1742) сформулировал гипотезу о
влиянии нервов на «все перемены в здоровом и
больном состоянии». Согласно У.Куллену (1712—
1790),
все процессы в здоровом и больном орга-
низме регулирует «нервный принцип», который
проявляет свое действие через головной мозг при
посредстве нервов — проводников нервной дея-
тельности. По мнению Е.О.Мухина (1817), «все
человеческое тело вообще можно, отвлекаясь,
рассматривать как построенное из нервов, ибо ос-
тальные части тела, видимо, существуют вследст-
вие нервов как управляющих их способностями».
Велики заслуги в развитии концепции нервиз-
ма И.М.Сеченова (1829—1905), С.П.Боткина
(1832—1889). Согласно С.П.Боткину, организм
—
это целостная система, деятельность которой на-
правляется и регулируется нервной системой. Он
рассматривал различные заболевания как следст-
вие нарушения нормальных нервных соотноше-
ний —
клинический
нервизм.
Нарушение «регуля-
торных нервных аппаратов» может явиться при-
чиной ряда болезней человека, что убедительно
подтверждено клиническими наблюдениями.
Выдающийся вклад в развитие концепции фи-
зиологического нервизма сделал И.П.Павлов
(1849—1936). И.П.Павлов обосновал представле-
ние о трофическом влиянии нервной системы на
органы и ткани, сформулировал принципы реф-
лекторной теории, доказал важную роль нервной
системы в регуляции секреции желез желудочно-
кишечного тракта, открыл условные рефлексы и с
их помощью разработал основы учения о высшей
нервной деятельности.
В.
Нервный центр — это совокупность
нейронов, расположенных на различных
уровнях ЦНС, достаточных для регуляции
функции органа согласно потребностям орга-
низма. Например, нейроны дыхательного
центра располагаются и в спинном мозге, и в
продолговатом мозге, и в мосту. Однако сре-
ди нескольких групп клеток, расположенных
на различных уровнях ЦНС, обычно имеется
главная часть центра. Главная часть дыха-
тельного центра находится в продолговатом
мозге и включает инспираторные и экспира-
торные нейроны.
Нервный центр реализует свое влияние на
эффекторы либо непосредственно с помо-
щью эфферентных импульсов соматической
и вегетативной нервной системы, либо с по-
мощью активации выработки соответствую-
щих гормонов. Кроме нервной регуляции, в
организме существуют гуморальный и мио-
генный механизмы регуляции. Гуморальный
механизм регуляции функций органов и тка-
ней организма осуществляется с помощью
гормонов, медиаторов, метаболитов и ткане-
вых гормонов.
3.2.
ХАРАКТЕРИСТИКА ГОРМОНАЛЬНОЙ
РЕГУЛЯЦИИ
У рефлекторной реакции может быть гормо-
нальное звено, что характерно для регуляции
работы внутренних органов (вегетативных
функций) в отличие от соматических, реф-
лекторная регуляция которых осуществляет-
ся только нервным путем (деятельность
опорно-двигательного аппарата). Если вклю-
чается гормональное звено, то это осущест-
вляется за счет дополнительной выработки
биологически активных веществ. Например,
при действии на экстерорецепторы сильных
раздражителей (холод, жара, болевой раздра-
житель) возникает мощный поток афферент-
ных импульсов, поступающих в ЦНС, при
этом в кровь выбрасывается дополнительное
количество адреналина и гормонов коры над-
почечников, играющих адаптивную (защит-
ную) роль.
Гормоны — биологически активные веще-
ства, вырабатываемые эндокринными желе-
зами или специализированными клетками,
находящимися в различных органах (напри-
мер,
в поджелудочной железе, желудочно-ки-
шечном тракте). Гормоны вырабатываются
также нервными клетками — это нейрогор-
моны, например гормоны гипоталамуса (ли-
берины и статины), регулирующие функцию
гипофиза (схема 3.1). Биологически актив-
ные вещества вырабатываются также неспе-
2-5484
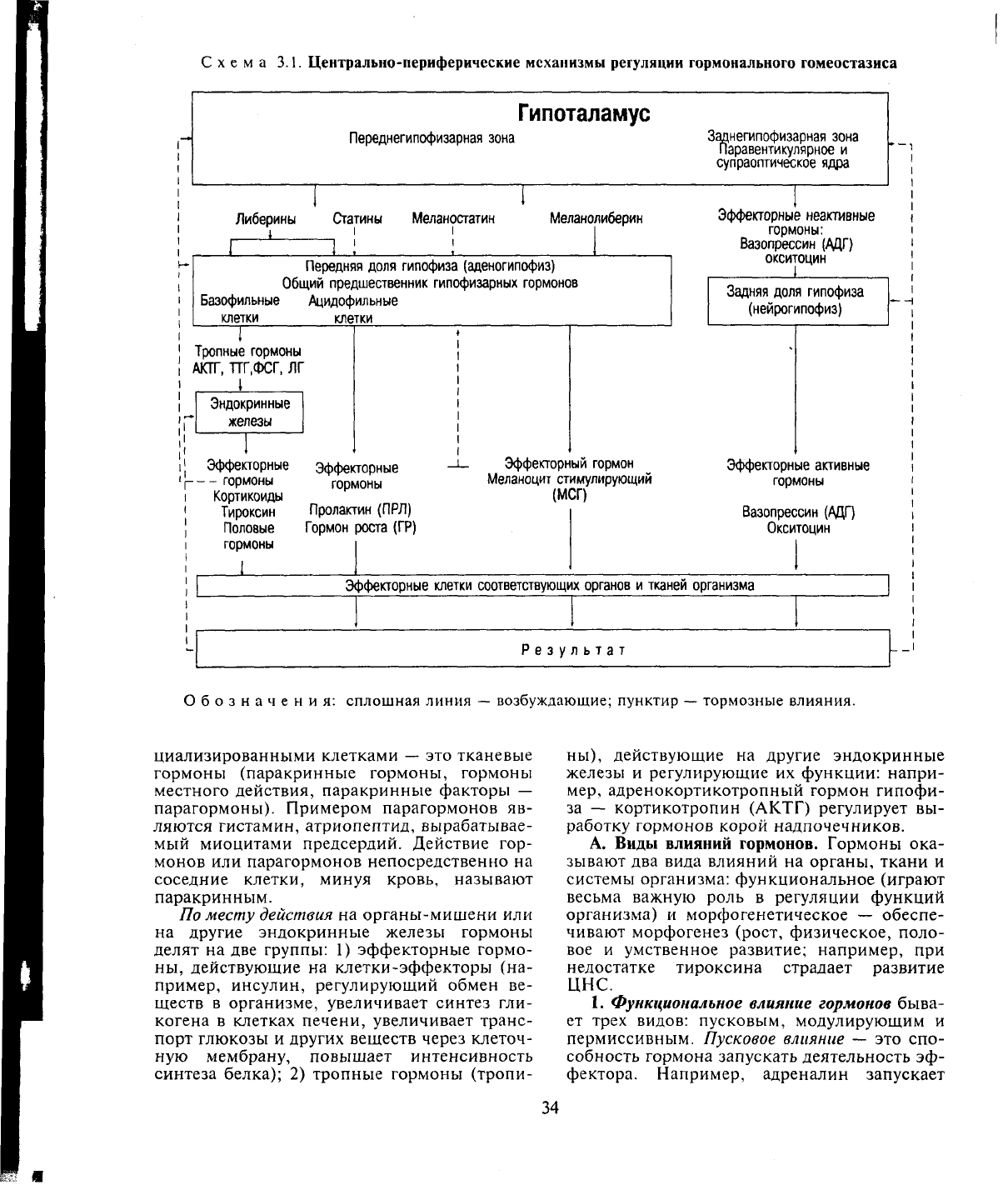
Схема 3.1. Центрально-периферические механизмы регуляции гормонального гомеостазиса
Гипоталамус
Переднегипофизарная зона
Заднегипофизарная зона
Паравентикулярное
и
супраоптическое ядра
Либерины Статины Меланостатин
Меланолиберин
Передняя доля гипофиза (аденогипофиз)
Общий предшественник гипофизарных гормонов
Базофильные Ацидофильные
клетки клетки
_
Тропные гормоны
АЮТ, ТТГ.ФСГ,
ЛГ
1
Эндокринные
железы
Эффекторные
--
гормоны
Кортикоиды
Тироксин
Половые
гормоны
Эффекторные
гормоны
Пролактин (ПРЛ)
Гормон роста (ГР)
Эффекторный гормон
Меланоцит стимулирующий
(МСГ)
Эффекторные неактивные
гормоны:
Вазопрессин (АДГ)
окситоцин
Задняя доля гипофиза
(нейрогипофиз)
Эффекторные активные
гормоны
Вазопрессин (АДГ)
Окситоцин
Эффекторные клетки соответствующих органов
и
тканей организма
Результат
Обозначения: сплошная линия
—
возбуждающие; пунктир
—
тормозные влияния.
циализированными клетками
—
это тканевые
гормоны (паракринные гормоны, гормоны
местного действия, паракринные факторы —
парагормоны). Примером парагормонов яв-
ляются гистамин, атриопептид, вырабатывае-
мый миоцитами предсердий. Действие гор-
монов или парагормонов непосредственно на
соседние клетки, минуя кровь, называют
паракринным.
По месту действия на органы-мишени или
на другие эндокринные железы гормоны
делят на две группы: 1) эффекторные гормо-
ны,
действующие на клетки-эффекторы (на-
пример, инсулин, регулирующий обмен ве-
ществ в организме, увеличивает синтез гли-
когена в клетках печени, увеличивает транс-
порт глюкозы и других веществ через клеточ-
ную мембрану, повышает интенсивность
синтеза белка); 2) тропные гормоны (тропи-
ны),
действующие на другие эндокринные
железы и регулирующие их функции: напри-
мер,
адренокортикотропный гормон гипофи-
за — кортикотропин (АКТГ) регулирует вы-
работку гормонов корой надпочечников.
А. Виды влияний гормонов. Гормоны ока-
зывают два вида влияний на органы, ткани и
системы организма: функциональное (играют
весьма важную роль в регуляции функций
организма) и морфогенетическое — обеспе-
чивают морфогенез (рост, физическое, поло-
вое и умственное развитие; например, при
недостатке тироксина страдает развитие
цнс.
1.
Функциональное
влияние гормонов быва-
ет трех видов: пусковым, модулирующим и
пермиссивным. Пусковое влияние — это спо-
собность гормона запускать деятельность эф-
фектора. Например, адреналин запускает
34

распад гликогена в печени и выход глюкозы в
кровь, вазопрессин (антидиуретический гор-
мон — АДГ) включает реабсорбцию воды из
собирательных трубок нефрона в интерсти-
ций почки. Модулирующее влияние — измене-
ние интенсивности протекания биохимичес-
ких процессов в органах и тканях. Например,
активация тироксином окислительных про-
цессов, которые могут проходить и без него;
стимуляция адреналином деятельности серд-
ца, которая проходит и без адреналина. Мо-
дулирующим влиянием гормонов является
также изменение чувствительности ткани к
действию других гормонов. Например, фол-
ликулин усиливает действие прогестерона на
слизистую оболочку матки, тиреоидные гор-
моны усиливают эффекты катехоламинов.
Пермиссивное влияние — способность одного
гормона обеспечивать реализацию эффекта
другого гормона. Например, инсулин необхо-
дим для проявления действия соматотропно-
го гормона, фоллитропин — для реализации
эффекта лютропина.
2.
Морфогенетическое влияние гормонов
(на рост, физическое и половое развитие) по-
дробно изучается другими дисциплинами
(гистология, биохимия) и лишь частично
—
в
курсе, физиологии (см. раздел 10). Оба вида
влияний гормонов (морфогенетическое и
функциональное) реализуются с помощью
метаболических процессов, запускаемых по-
средством клеточных ферментных систем.
Б.
Механизм действия различных гормо-
нов.
Известны положения Эрлиха
(1911,
1913):
«вещества не действуют, не будучи
связанными» и «замок и ключ». Роль замка
выполняют рецепторы, расположенные на
клетках, роль ключа
—
лиганд (гормон, меди-
атор,
фармакологический препарат). Для
всех гормонов на клетках-мишенях имеется
рецептор, выполняющий функцию распозна-
вания гормона (рецептор узнавания). Влия-
ние любого гормона на клетку-мишень начи-
нается со взаимодействия с рецептором узна-
вания. Далее реализация эффекта зависит от
природы гормона. Имеется два основных ва-
рианта. Если клеточная мембрана непрони-
цаема для гормона, то эффекторные рецепто-
ры расположены на ее поверхности (белково-
полипептидные и аминокислотные гормоны
гипоталамуса, гипофиза, поджелудочной же-
лезы, мозгового слоя надпочечников водо-
растворимы, но нерастворимы в липидах).
Если клеточная мембрана проницаема для
гормона, то эффекторные рецепторы для
этих гормонов расположены в основном в
цитоплазме (стероидные гормоны: кортикои-
ды и половые гормоны — растворимы в ли-
пидах). Гормоны щитовидной железы могут
проникать в клетку, но только в комплексе
со специальным белком плазмы крови, по-
средством эндоцитоза. Их рецепторы распо-
ложены как на поверхности клеточной мем-
браны, так и в цитоплазме.
Гормонрецепторный комплекс белково-
пептидных гормонов и производных амино-
кислот активирует мембранные ферменты,
что ведет к образованию вторых посредни-
ков,
реализующих свое действие в цитоплаз-
ме и ядре клетки. Основными из них являют-
ся:
1) аденилатциклаза — циклический аде-
нозинмонофосфат (цАМФ); 2) гуанилатцик-
лаза — циклический гуанозинмонофосфат
(цГМФ); 3) фосфолипаза С — инозитолтри-
фосфат (ИФ
3
); 4) ионизированный каль-
ций
—
кальмодулин (белок немышечных кле-
ток) или тропонин-С (в мышечных клетках).
Гормонрецепторный комплекс, образуемый в
цитоплазме стероидными гормонами и гор-
монами щитовидной железы, проникающи-
ми в клетку, поступает в клеточное ядро и
другие органеллы клетки, где происходит со-
единение его с другим рецептором, после
чего реализуется влияние данного гормона на
клетку. Своеобразными вторыми посредни-
ками этих гормонов являются различные
виды РНК. Часть эффектов стероидных гор-
монов, как и гормонов щитовидной железы,
реализуется посредством рецепторов, распо-
ложенных на клеточной мембране, и вторых
посредников, как и у белково-пептидных
гормонов.
Прекращение действия гормонов осущест-
вляется с помощью тканевых ферментов и
ферментов самих эндокринных желез, пече-
ни,
почек. Многие продукты расщепления
гормонов также активны и вызывают иногда
сходные эффекты. Продукты распада гормо-
нов выводятся главным образом почками, а
также железами — слюнными, желудочно-
кишечного тракта, потовыми и с желчью.
В.
Регуляция выработки гормонов. В желе-
зах внутренней секреции иннервированы,
как правило, только сосуды. Выработка гор-
монов регулируется метаболитами, гормона-
ми гипофиза и другими гормонами. Напри-
мер,
ангиотензин-П стимулирует секрецию
альдостерона. Соматостатин, кроме гипофи-
за, вырабатывается в поджелудочной железе,
где он подавляет секрецию инсулина и глю-
кагона. Функция гипофиза регулируется в
свою очередь гормонами гипоталамуса —
нейрогормонами. Для некоторых эндокрин-
ных желез основным механизмом является
местная саморегуляция. Так, секреция инсу-
лина и глюкагона клетками островков Лан-
г-

герганса регулируется уровнем глюкозы в
крови. Если концентрация глюкозы в крови
высока, то по принципу обратной отрица-
тельной связи стимулируется выработка ин-
сулина, который снижает концентрацию
глюкозы в крови с помощью увеличения ути-
лизации ее клетками организма и увеличения
отложения в виде гликогена в клетках пече-
ни,
в результате чего снижается (нормализу-
ется) концентрация глюкозы в крови. В слу-
чае снижения концентрации глюкозы в кро-
ви выработка инсулина уменьшается, выра-
ботка глюкагона клетками островков Лангер-
ганса возрастает (глюкагон увеличивает пре-
образование гликогена печени в глюкозу и
выход ее в кровь). Секреция кальцийрегули-
рующих гормонов (паратирин и кальцито-
нин) также регулируется по принципу обрат-
ной отрицательной связи — концентрацией
кальция в крови. Общую схему регуляции
выработки гормонов см. на рис. 3.3.
3.3.
РЕГУЛЯЦИЯ С ПОМОЩЬЮ
МЕТАБОЛИТОВ И ТКАНЕВЫХ ГОРМОНОВ.
МИОГЕННЫЙ МЕХАНИЗМ РЕГУЛЯЦИИ
А.
Метаболиты — продукты, образующиеся
в организме в процессе обмена веществ как
результат различных биохимических реак-
ций. Это аминокислоты, нуклеотиды, кофер-
менты, угольная, молочная, пировиноград-
ная,
адениловая кислоты, ионный сдвиг, из-
менения рН. Регуляция с помощью метабо-
литов на ранних этапах филогенеза была
единственной. Метаболиты одной клетки не-
посредственно влияли на другую — сосед-
нюю клетку или группу клеток, которые в
свою очередь таким же способом действова-
ли на следующие клетки (контактная регуля-
ция).
С появлением гемолимфы и сосудистой
системы метаболиты стали передаваться к
другим клеткам организма с движущейся ге-
молимфой на большие расстояния, причем
осуществляться это стало быстрее. Затем по-
явилась нервная система как регулирующая
система, а еще позже — эндокринные желе-
зы.
Метаболиты хотя и действуют в основном
как местные регуляторы, но могут влиять и
на другие органы и ткани, на активность
нервных центров. Например, накопление
угольной кислоты в крови ведет к возбужде-
нию дыхательного центра и усилению дыха-
ния. Примером местной гуморальной регуля-
ции может служить гиперемия интенсивно
работающей скелетной мышцы: накапливаю-
щиеся метаболиты обеспечивают расширение
кровеносных сосудов, что увеличивает до-
ставку кислорода и питательных веществ к
мышце. Подобные регуляторные влияния ме-
таболитов происходят и в других активно ра-
ботающих органах и тканях организма.
Б. Тканевые гормоны: биогенные амины
(гистамин, серотонин), простагландины, ки-
нины (калликреин, брадикинин), атриопеп-
тид и др. занимают промежуточное положе-
ние между гормонами и метаболитами как
гуморальные факторы регуляции. Эти веще-
ства оказывают свое регулирующее влияние
на клетки тканей посредством изменения их
биофизических свойств (проницаемости
мембран, их возбудимости), изменения ин-
тенсивности обменных процессов, чувстви-
тельности клеточных рецепторов, образова-
ния вторых посредников. В результате этого
изменяется чувствительность клеток к нерв-
ным и гуморальным влияниям. Поэтому тка-
невые гормоны называют модуляторами ре-
гуляторных сигналов: они оказывают моду-
лирующее влияние. Тканевые гормоны обра-
зуются неспециализированными клетками,
но действуют они посредством специализи-
рованных клеточных рецепторов: например,
для гистамина обнаружено два вида рецепто-
ров — Н, и Н
2
. Поскольку тканевые гормоны
влияют на проницаемость клеточных мем-
бран, они регулируют поступление в клетку и
выход из клетки различных веществ и ионов,
определяющих мембранный потенциал, а
значит
—
и развитие потенциала действия.
В.
Миогенный механизм регуляции. С раз-
витием мышечной системы в процессе эво-
люции миогенный механизм регуляции
функций постепенно становится все более
заметным. Организм человека примерно на
50 % состоит из мышц по своей массе. Это
скелетная мускулатура (40 % массы тела),
мышца сердца, гладкие мышцы кровеносных
и лимфатических сосудов, стенки желудоч-
но-кишечного тракта, желчного, мочевого
пузырей и других внутренних органов.
Сущность
миогенного механизма регуляции
состоит в том, что предварительное умеренное
растяжение скелетной или сердечной мышц
увеличивает силу их сокращений. Сократи-
тельная активность гладкой мышцы также за-
висит от степени наполнения полого мышеч-
ного органа, а значит, и его растяжения. При
увеличении наполнения органа тонус гладкой
мышцы сначала возрастает, а затем возвраща-
ется к исходному уровню (пластичность глад-
кой мышцы), что обеспечивает регуляцию то-
нуса сосудов и наполнение внутренних полых
органов без существенного повышения давле-
ния в них (до определенной величины).
Кроме того, большинство гладких мышц об-
ладает автоматией, они постоянно находятся
36

в некоторой степени сокращения под влияни-
ем импульсов, возникающих в них самих (на-
пример, мышцы кишечника, кровеносных со-
судов). Импульсы, поступающие к ним по ве-
гетативным нервам, оказывают модулирую-
щее влияние — увеличивают или уменьшают
тонус гладких мышечных волокон.
3.4.
ЕДИНСТВО И ОСОБЕННОСТИ
РЕГУЛЯТОРНЫХ МЕХАНИЗМОВ.
ФУНКЦИИ ГЕМАТОЭНЦЕФАЛИЧЕСКОГО
БАРЬЕРА
А. Единство регуляторных механизмов заклю-
чается в их взаимодействии. Так, например,
увеличение содержания углекислого газа в
крови возбуждает хеморецепторы аортальной
и синокаротидной рефлексогенных зон, при
этом увеличивается поток импульсов по со-
ответствующим нервам в ЦНС, а оттуда — к
дыхательной мускулатуре, что ведет к учаще-
нию и углублению дыхания. Углекислый газ
действует на дыхательный центр и непосред-
ственно, что также вызывает усиление дыха-
ния. При действии холодного воздуха на тер-
морецепторы кожи увеличивается поток аф-
ферентных импульсов в ЦНС; это ведет к вы-
бросу гормонов, увеличивающих интенсив-
ность обмена веществ, и к увеличению теп-
лопродукции.
Б.
Особенности нервного и гуморального
механизмов регуляции функций организма.
1.
Нервная система в отличие от гумораль-
ного механизма регуляции организует ответ-
ные реакции на изменения внешней среды
организма. Пусковым звеном в нейрогумо-
ральной регуляции при изменении внутрен-
ней среды также нередко является нервная
система.
2.
У нервного и гуморального механизмов
регуляции функций различные способы свя-
зи:
у нервной системы — нервный импульс
как универсальный сигнал, а у гуморального
механизма связь с регулируемым органом
или тканью осуществляется с помощью раз-
личных химических веществ. Таковыми яв-
ляются гормоны, медиаторы, метаболиты и
так называемые тканевые гормоны (парагор-
моны).
Некоторые медиаторы, например ка-
техоламины, попадая в кровь, могут действо-
вать не только в месте их выделения нервны-
ми окончаниями, но и на другие органы и
ткани организма, т.е. выступать в роли гумо-
ральных факторов, участвующих в регуляции
функций других органов организма.
3.
У нервного и гуморального механизмов
регуляции функций организма различная
точность связи. Химические вещества, попа-
дая в кровь, разносятся по всему организму и
действуют нередко на многие органы и тка-
ни — системный (генерализованный) харак-
тер влияния. Например, адреналин, тирок-
син (см. раздел 10), попадая в кровь, разно-
сятся по всему организму и действуют на
клетки всех органов и тканей организма.
Нервная система может оказывать точное,
локальное влияние на отдельный орган или
даже на группу клеток этого органа. Так,
нервная система может вызывать сокраще-
ния мышц указательного или другого пальца
руки, не вызывая сокращения мышц всей ко-
нечности или других пальцев. Следует, одна-
ко,
заметить, что и у гуморального механизма
нередко имеется точный адресат воздейст-
вий. Кортикотропин хотя и разносится с кро-
вью по всему организму, но действует только
на кору надпочечников. Тиреотропин (ТТГ)
регулирует функцию щитовидной железы.
В свою очередь и нервная система может
оказывать генерализованное влияние. На-
пример, возбуждение симпатической нерв-
ной системы в экстремальных условиях ведет
к мобилизации ресурсов всего организма для
достижения цели (стимулируется деятель-
ность сердечно-сосудистой, дыхательной и
эндокринной систем).
4.
У нервного и гуморального механизмов
регуляции различная скорость связи: относи-
тельно медленно с током крови распростра-
няются химические вещества (самая большая
средняя скорость в аорте — 0,25 м/с, а самая
маленькая — в капиллярах: 0,3—0,5 мм/с).
Частица крови проходит один раз через весь
организм (большой и малый круг кровообра-
щения) за 22 с. Нервный импульс распро-
страняется со скоростью до 120 м/с.
5.
Гормональные механизмы регуляции
подчиняются нервной системе, которая пере-
дает свое влияние на эндокринные железы
непосредственно или с помощью нейропеп-
тидов и своих медиаторов (посредников), вы-
деляемых нервными окончаниями и действу-
ющих на специальные, чувствительные к ме-
диаторам структуры — рецепторы.
6. У гуморального механизма регуляции
нередко наблюдается противоположное вли-
яние биологически активных веществ на
один и тот же орган в зависимости от точки
приложения действия этого химического ве-
щества. Так, например, угольная кислота,
действуя прямо на кровеносные сосуды, вы-
зывает их расширение, а посредством воз-
буждения центра кровообращения — суже-
ние.
Адреналин при непосредственном дей-
ствии на сердце стимулирует его работу, а
при введении в ликвор, возбуждая центры
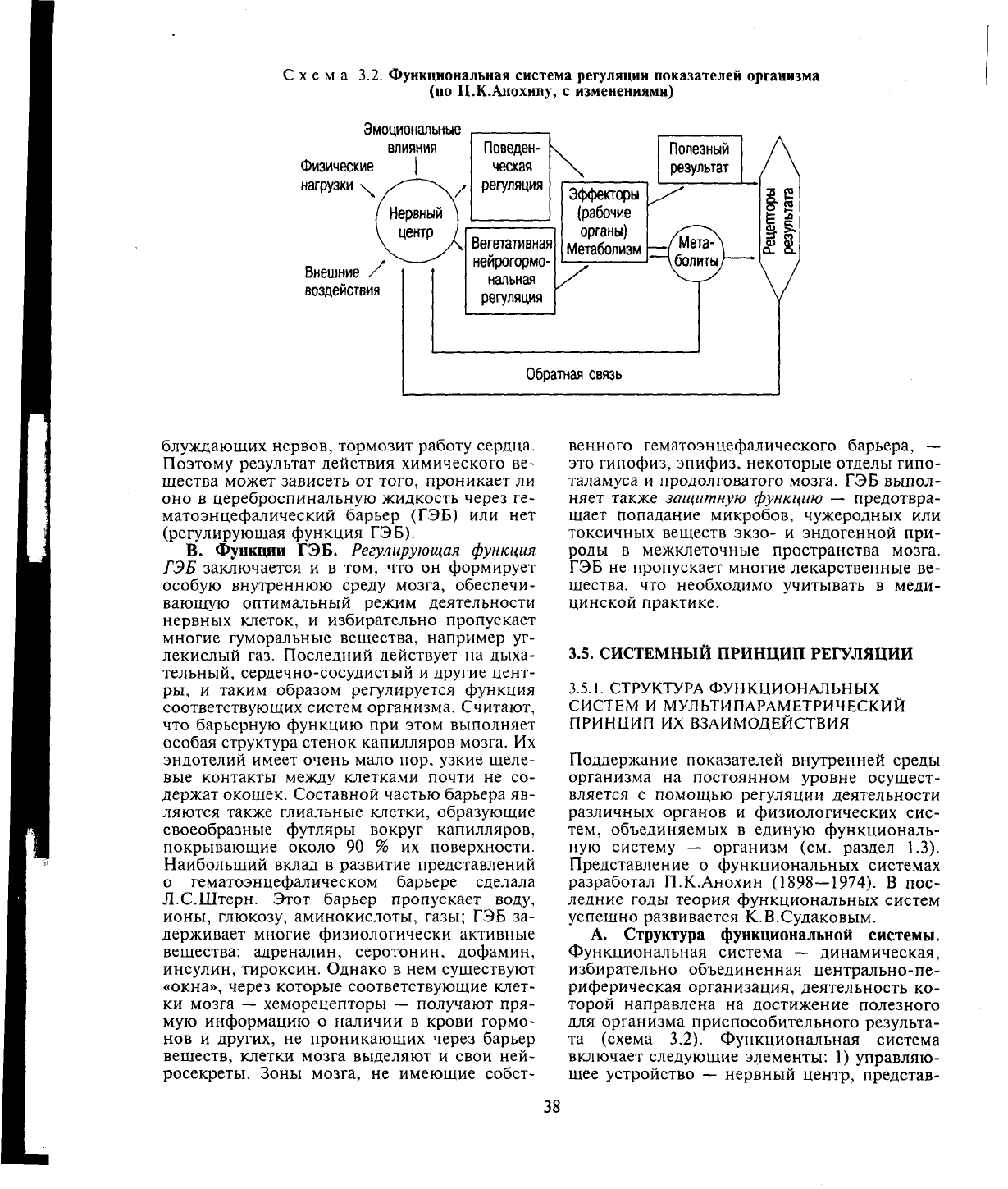
Схема 3.2. Функциональная система регуляции показателей организма
(по П.К.Анохину, с изменениями)
Эмоциональные
влияния
Физические
нагрузки
\^
Внешние
воздействия
Поведен-
ческая
регуляция
Вегетативная
нейрогормо-
нальная
регуляция
Эффекторы
(рабочие
органы)
Метаболизм
Полезный
результат
8-g
it
Обратная связь
блуждающих нервов, тормозит работу сердца.
Поэтому результат действия химического ве-
щества может зависеть от того, проникает ли
оно в цереброспинальную жидкость через ге-
матоэнцефалический барьер (ГЭБ) или нет
(регулирующая функция ГЭБ).
В.
Функции ГЭБ. Регулирующая функция
ГЭБ заключается и в том, что он формирует
особую внутреннюю среду мозга, обеспечи-
вающую оптимальный режим деятельности
нервных клеток, и избирательно пропускает
многие гуморальные вещества, например уг-
лекислый газ. Последний действует на дыха-
тельный, сердечно-сосудистый и другие цент-
ры,
и таким образом регулируется функция
соответствующих систем организма. Считают,
что барьерную функцию при этом выполняет
особая структура стенок капилляров мозга. Их
эндотелий имеет очень мало пор, узкие щеле-
вые контакты между клетками почти не со-
держат окошек. Составной частью барьера яв-
ляются также глиальные клетки, образующие
своеобразные футляры вокруг капилляров,
покрывающие около 90 % их поверхности.
Наибольший вклад в развитие представлений
о гематоэнцефалическом барьере сделала
Л.С.Штерн. Этот барьер пропускает воду,
ионы, глюкозу, аминокислоты, газы; ГЭБ за-
держивает многие физиологически активные
вещества: адреналин, серотонин, дофамин,
инсулин, тироксин. Однако в нем существуют
«окна», через которые соответствующие клет-
ки мозга — хеморецепторы — получают пря-
мую информацию о наличии в крови гормо-
нов и других, не проникающих через барьер
веществ, клетки мозга выделяют и свои ней-
росекреты. Зоны мозга, не имеющие собст-
венного гематоэнцефалического барьера, —
это гипофиз, эпифиз, некоторые отделы гипо-
таламуса и продолговатого мозга. ГЭБ выпол-
няет также защитную функцию — предотвра-
щает попадание микробов, чужеродных или
токсичных веществ экзо- и эндогенной при-
роды в межклеточные пространства мозга.
ГЭБ не пропускает многие лекарственные ве-
щества, что необходимо учитывать в меди-
цинской практике.
3.5.
СИСТЕМНЫЙ ПРИНЦИП РЕГУЛЯЦИИ
3.5.1.
СТРУКТУРА ФУНКЦИОНАЛЬНЫХ
СИСТЕМ И МУЛЬТИПАРАМЕТРИЧЕСКИЙ
ПРИНЦИП ИХ ВЗАИМОДЕЙСТВИЯ
Поддержание показателей внутренней среды
организма на постоянном уровне осущест-
вляется с помощью регуляции деятельности
различных органов и физиологических сис-
тем, объединяемых в единую функциональ-
ную систему — организм (см. раздел 1.3).
Представление о функциональных системах
разработал П.К.Анохин (1898—1974). В пос-
ледние годы теория функциональных систем
успешно развивается К.В.Судаковым.
А. Структура функциональной системы.
Функциональная система — динамическая,
избирательно объединенная центрально-пе-
риферическая организация, деятельность ко-
торой направлена на достижение полезного
для организма приспособительного результа-
та (схема 3.2). Функциональная система
включает следующие элементы: 1) управляю-
щее устройство — нервный центр, представ-
38

ляющий объединение ядер различных уров-
ней ЦНС; 2) его выходные каналы (нервы и
гормоны); 3) исполнительные органы — эф-
фекторы, обеспечивающие в ходе физиологи-
ческой деятельности поддержание регулируе-
мого процесса показателя на некотором оп-
тимальном уровне (полезный результат дея-
тельности функциональной системы); 4) ре-
цепторы результата (сенсорные рецепторы) —
датчики, воспринимающие информацию о
параметрах отклонения регулируемого про-
цесса показателя от оптимального уровня;
5) канал обратной связи (входные каналы),
информирующие нервный центр с помощью
импульсаций от рецепторов результата или
же на основе изменений химического состава
тех или иных жидкостей организма о доста-
точности или недостаточности эффекторных
усилий по поддержанию регулируемого про-
цесса показателя на оптимальном уровне.
Афферентные импульсы от рецепторов ре-
зультата по каналам обратной связи поступа-
ют в нервный центр, регулирующий тот или
иной показатель. Например, при увеличении
артериального давления крови в большей
степени начинают раздражаться барорецеп-
торы рефлексогенных сосудистых зон, в ре-
зультате чего увеличивается поток импульсов
в ЦНС — в центр кровообращения. Взаимо-
действие нейронов этого центра и изменения
интенсивности эфферентной импульсаций
ведут к ослаблению деятельности сердца и
расширению кровеносных сосудов. Артери-
альное давление крови снижается. Возможны
колебания величины артериального давле-
ния, но оно после ряда колебаний возвраща-
ется к нормальной величине. Если же опи-
санного механизма оказалось недостаточно и
давление остается повышенным, включаются
дополнительные регуляторные механизмы: в
частности возрастает переход жидкости из
кровеносного русла в межклеточное про-
странство (интерстиций), включается эндо-
кринная система, больше воды выводится из
организма (подробнее см. раздел 13.10).
При изменении интенсивности работы
эффектора изменяется интенсивность мета-
болизма, что также играет важную роль в ре-
гуляции деятельности органов той или иной
функциональной системы. Например, при
усилении сокращений мышцы увеличивается
интенсивность обмена веществ, в кровь вы-
деляется значительно больше метаболитов.
Последние действуют, во-первых, непосредст-
венно на орган-эффектор (в данном случае
это приводит к расширению кровеносных со-
судов и к улучшению кровоснабжения орга-
на, что весьма важно). Во-вторых, метаболи-
ты,
попадая в кровь, а с кровью — в ЦНС,
действуют также и на соответствующие цент-
ры,
изменение активности которых оказыва-
ет корригирующее влияние на органы и
ткани организма. В-третьих, метаболиты
действуют также на рецепторы рабочего ор-
гана (или органов) — рецепторы результата,
что тоже отражается на активности рецепто-
ров и, естественно, на импульсаций в аффе-
рентных путях, проводящих импульсы в
ЦНС по принципу обратной связи.
Архитектура различных функциональных
систем принципиально одинакова, что назы-
вают изоморфизмом. Вместе с тем функцио-
нальные системы могут отличаться друг от
друга по степени разветвленности как цент-
ральных, так и периферических механизмов.
Необходимо подчеркнуть, что системообра-
зующим фактором, выступающим в качестве
инструмента включения тех или иных орга-
нов,
тканей, механизмов в функциональную
систему, является полезный для жизнедея-
тельности организма приспособительный ре-
зультат — конечный продукт физиологичес-
кой активности функциональной системы.
Ряд гомеостатических функциональных
систем представлен исключительно внутрен-
ними генетически детерминированными ме-
ханизмами вегетативной нервно-гормональ-
ной регуляции и не включает механизмы по-
веденческой соматической регуляции. При-
мером являются функциональные системы,
определяющие оптимальные для обмена ве-
ществ организма кровяное давление, содер-
жание ионов в крови, не изменяющих осмо-
лярность и не вызывающих чувство жажды,
рН внутренней среды организма. Другие го-
меостатические функциональные системы
включают целенаправленное поведение во
внешней среде на базе доминирующих моти-
вационных возбуждений, отражающих сдви-
ги различных показателей метаболизма.
В этом случае системообразующим фактором
является также и мотивация. Примерами
таких функциональных систем являются сис-
темы, обеспечивающие поддержание опти-
мального уровня питательных веществ в ор-
ганизме, осмотического давления и объема
жидкости в организме, температуры внутрен-
ней среды организма. В этом случае опорно-
двигательный аппарат выступает как состав-
ная часть эффектора — рабочего органа. При
этом реагируют и многие внутренние органы,
обеспечивающие усиление сократительной
деятельности скелетной мускулатуры — это
тоже составная часть эффектора. В частнос-
ти,
усиливается деятельность сердца, стиму-
лируется дыхание.

Б.
Мультипараметрический принцип взаи-
модействия различных функциональных сис-
тем — принцип, определяющий обобщенную
деятельность функциональных систем. Отно-
сительная стабильность показателей внут-
ренней среды организма является результа-
том согласованной деятельности многих
функциональных систем. Выяснилось, что
различные константы внутренней среды ор-
ганизма оказываются взаимосвязанными. Это
проявляется в том, что изменение величины
одной константы может привести к измене-
нию параметров других показателей. Напри-
мер,
избыточное поступление воды в орга-
низм сопровождается увеличением объема
циркулирующей крови, повышением артери-
ального давления, снижением осмотического
давления плазмы крови. В функциональной
системе, поддерживающей оптимальный уро-
вень газового состава крови, одновременно
осуществляется взаимодействие рН, РС0
2
и
Р0
2
.
Изменение одного из этих параметров
немедленно приводит к изменению количе-
ственных характеристик других параметров.
На основе принципа мультипараметричес-
кого взаимодействия все функциональные
системы гомеостатического уровня факти-
чески объединяются в единую функциональ-
ную систему гомеостазиса. Отдельные компо-
ненты такой системы ориентированы на под-
держание отдельных показателей внутренней
среды организма. Другие компоненты ориен-
тированы на достижение некоторых поведен-
ческих результатов (поведенческое звено ре-
гуляции) в соответствии с глобальными по-
требностями организма поддержать всю со-
вокупность показателей внутренней среды
организма.
Для достижения любого приспособитель-
ного результата формируется соответствую-
щая функциональная система.
3.5.2. СИСТЕМОГЕНЕЗ
Согласно П.К.Анохину, системогенез
—
изби-
рательное созревание и развитие функциональ-
ных систем в анте- и постнатальном онтоге-
незе. В отличие от понятия «морфогенез»,
предложенного А.Н.Северцевым (развитие
органов в онтогенезе), «системогенез» отра-
жает развитие в онтогенезе различных по
функции и локализации структурных образо-
ваний, которые объединяются в полноцен-
ную функциональную систему, обеспечиваю-
щую новорожденному выживание.
В настоящее время термин «системогенез»
применяется в более широком смысле, при
этом под системогенезом понимают процес-
сы не только онтогенетического созревания
функциональных систем, но и формирование
и преобразование функциональных систем в
ходе жизнедеятельности зрелого организма.
Примеры динамичных перестроек функцио-
нальных систем можно найти, анализируя
активность индивидуумов при формирова-
нии новых навыков. Так, системные меха-
низмы достижения полезных результатов на
начальном этапе формирования навыков и
на этапе автоматизированных навыков раз-
личны прежде всего по объему мышечных
усилий и по уровню их вегетативного обеспе-
чения.
Общие принципы формирования функцио-
нальных систем в онтогенезе по П.К.Анохину.
1.
Системообразующим фактором функ-
циональной системы любого уровня является
полезный для жизнедеятельности организма
приспособительный результат, необходимый
в данный момент. Этому правилу подчиня-
ются процесс созревания различных функци-
ональных систем на разных этапах онтогене-
за и деятельность функциональных систем
зрелого организма. Примерами могут быть
поддержание различных физиологических
показателей (артериального давления, осмо-
тического давления, рН внутренней среды
организма, температуры тела) с помощью ре-
гуляции функций внутренних органов и по-
веденческих реакций; достижение результата
социальной деятельности — в работе, учебе.
В конечном итоге все множество полезных
приспособительных результатов можно объ-
единить в две группы: 1) поддержание посто-
янства внутренней среды организма; 2) до-
стижение результата в социальной деятель-
ности. В системогенезе выделяют два основ-
ных периода
—
антенатальный (внутриутроб-
ный) и постнатальный (внеутробный).
2.
Принцип
гетерохронией закладки и гете-
рохронного созревания компонентов функцио-
нальной системы. В ходе антенатального он-
тогенеза различные структуры организма за-
кладываются в разное время и созревают с
различными темпами. Так, нервный центр
группируется и созревает обычно раньше,
чем закладывается и созревает иннервируе-
мый им субстрат. В онтогенезе созревают в
первую очередь те функциональные системы,
без которых невозможно дальнейшее развитие
организма. Например, у плода развивается
функциональная система, поддерживающая
постоянство газового состава крови с помо-
щью плацентарного кровообращения: кисло-
род поступает из крови матери в кровь плода,
а углекислый газ транспортируется в проти-
воположном направлении — из крови плода
40