Schlick T. Molecular Modeling and Simulation: An Interdisciplinary Guide
Подождите немного. Документ загружается.

160 5. Nucleic Acids Structure Minitutorial
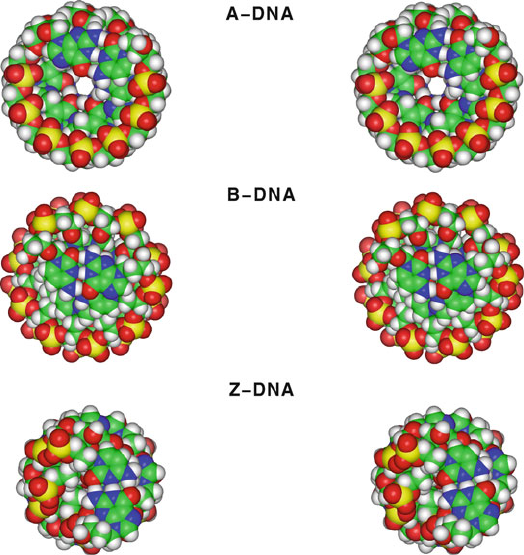
Figure 5.15. Space-filling stereo figures of model A, B, and Z-DNA, as generated by
the polynucleotide building program developed in [1102] based on parameters listed in
Table 5.4. The stereo top-view images are rotated about 8
o
relative to one another, with the
center of images separated by about 6.5 cm, the average distance between two human eyes.
Images are prepared for cross-eyed viewing from about 46–51 cm. See also Figure 5.14
for corresponding side views.
5.4.3 Z-DNA
In contrast to the A and B-DNA models, specific sequences are required for
Z-DNA (e.g., alternating GC). The repeating unit for Z-DNA is a dinucleotide
rather than a mononucleotide. The regular composition of alternating pyrimidines
and purines allows alternating features in geometry, for example in the sugar and
glycosyl orientations. Besides its distinguishing left-handedness, the following
are properties of the slender Z-DNA helix (modeled for GC polymers):
• The bp center is shifted slightly from the helix axis along e1 and e2 (dx ≈
−3
˚
A, dy ≈±2.5
˚
A in Figure 5.11).
• The bps are inclined slightly (η ≈−7
o
).
• The mean helical twist is about 60
o
per dimer, with about 52
o
for the G →C
step and 6
o
for the C → Gstep[1102].
• There are 12 bps per turn.
5.4. Canonical DNA Forms 161
• The sugars adopt C2
-endo conformers at C but C3
-endo forms at G; see
Figure 5.8.
• The glycosyl link is anti for C but syn for G in alternating CG sequences;
see Figure 5.9.
• The major groove bulges out and the minor groove is narrow and deep.
Like the A-form, a stretch of Z-DNA might occur within B-DNA, but direct
experimental evidence is lacking. The negative supercoiling of naturally occurring
DNA (see next chapter) may also promote Z-DNA, or other left-handed, helix
formation.
Biological Significance
The biological role of Z-DNA forms continues to be an active area of research.
It has been recently found that when DNA adopts the Z conformation, an ed-
itor RNA molecule — the double-stranded RNA enzyme adenosine deaminase
(ADAR1) — can bind to left-handed DNA and alter the base in a codon (for
example from adenine to guanine) and thereby produce alternative forms of the
translated proteins. The Z-DNA binding domain of ADAR1, namely Zα,wasre-
cently co-crystallized with a 6-bp DNA fragment [1147]. This newly-identified
role for Z-DNA — a regulator of the nucleotide template which directs protein
synthesis — may have practical benefits as another element of biological control.
5.4.4 Comparative Features
Table 5.3 summarizes basic features of model A, B, and Z-DNA forms as com-
piled and reconciled from several textbooks. See also Figure 5.8 for sugar analysis
and Figure 5.9 for torsion-angle analysis.
Such model helices can also be constructed from building blocks of nucleo-
tides (for A and B-DNA) and dinucleotides (for Z-DNA), for example using the
geometric variables shown in Table 5.4 [1102].
In practice, departures from model-helix variables occur frequently and the ob-
served ranges in some of these quantities are quite large and differ markedly from
one residue to the next.
For example, analyses of high-resolution DNA structures collected since the
early 1980s indicate a mean twist angle (Ω) for the right-handed B-DNA of around
34
o
(rather than 36
o
), though a broad sequence-dependentrange (roughly 20–50
o
)
is observed. The helical twist of 34
o
corresponds to about n
b
=10.5 bps per turn
(rather than 10), 36
˚
A for the helical pitch, and h =3.4
˚
A for the axial rise (or
rise per residue).
A large field of research is devoted to unraveling the sequence-dependent fea-
tures of DNA. These sensitive patterns are important for numerous biological
functions involving DNA, such as protein binding and transcription regulation.
The next chapter introduces basic elements of DNA sequence effects, DNA hy-
dration and ion interactions, DNA/protein interactions, RNA structure, cellular
aspects of DNA organization, and DNA supercoiling.
162 5. Nucleic Acids Structure Minitutorial
6
Topics in Nucleic Acids Structure: DNA
Interactions and Folding
Chapter 6 Notation
S
YMBOL DEFINITION
h helical rise, or base-pair step separation (e.g., 3.4
˚
A)
k
B
Boltzmann’s constant
n
number of DNA base pairs
n
b
number of base pairs per DNA turn
p
b
DNA bending persistence length
p
tw
DNA twisting persistence length
r
elastic ratio of bending to twisting elastic constants, A/C
s
arclength
A
DNA bending rigidity
C
DNA torsional-rigidity constant
Dy
slide (between successive base pairs)
L
DNA contour length
Lk
linking number (topological invariant)
Lk
0
reference linking number
R
2
mean square displacement
T
temperature
T
c
site juxtaposition time
Tw
total twist (geometric variable)
Wr
writhing number (geometric variable)
θ
b
bend angle (for isotropic polymer models)
θ
R
net roll angle
θ
T
net tilt angle
κ
curvature (also Debye screening parameter)
ρ
roll angle (between successive base pairs)
σ
ΔLk/Lk
0
(superhelical density)
T. Schlick, Molecular Modeling and Simulation: An Interdisciplinary Guide, 163
Interdisciplinary Applied Mathematics 21, DOI 10.1007/978-1-4419-6351-2
6,
c
Springer Science+Business Media, LLC 2010

164 6. Topics in Nucleic Acids Structure: DNA Interactions and Folding
Chapter 6 Notation Table (continued)
S
YMBOL DEFINITION
σ
e
Poisson’s ratio (for an elastic material)
τ
tilt angle (between successive base pairs)
χ
glycosyl (sugar/base) torsion angle
ω
DNA twist rate; also propeller twist (within a base pair)
ω
0
intrinsic twist rate of DNA (e.g., 2π/10.5 radians)
ΔLk
linking number difference with respect to Lk
0
Ω twist angle
It has not escaped our notice that the specific pairing we have pos-
tulated immediately suggests a possible copying mechanism for the
genetic material.
Molecular Structure of Nucleic Acids, J.D. Watson and F.H.C. Crick (1953).
6.1 Introduction
This chapter introduces further topics in nucleic acid structure, building upon the
minitutorial of the previous chapter. These topics include DNA sequence effects,
DNA hydration, DNA/protein interactions, and the cellular organization of DNA,
including supercoiling and chromatin structure. The next chapter expands upon
the related topics of alternative hydrogen bonding schemes, non-canonical helical
and hybrid structures, DNA mimics, overstretched and understretched DNA, and
RNA structure and folding.
If I dare write something on any of these topics — given the excellent books and
book sections [100,137,139,163,195,893, 895, 1080,1191,1305] and numerous
articles already written on these subjects — it is only because these important
areas in nucleic acid structure cannot be omitted.
The sequence-induced variations of DNA, for example, profoundly affect the
local structure of DNA and that of DNA complexed with other molecules (water,
ions, ligands, and proteins). Such local variations translate into global structural
effects when considered on the molecular and cellular levels. The associated ener-
getic and dynamic effects, in turn, affect key functional processes, from the action
of regulatory proteins (replication, repair, transcription, etc.) to genome packaging
and processing. This functional influence of DNA sequence on biological activ-
ity has been shown by the works of Olson, Zhurkin, Dickerson, and many others
(e.g., [308, 942]).
Until recently, nucleic acids might have been considered ‘the silent partner’
in regulatory complex formation with proteins. This view is beginning to change
as our appreciation grows for the subtle, sequence-dependent effects of nucleic
acids, including the recently discovered “second genetic code”, the nucleosome
positioning code [1157, 1288,1425].
6.2. DNA Sequence Effects 165
Though presenting these topics in this text contrasts with the very brief and
basic protein chapters, I hope to encourage mathematical and physical scientists
who may not read structural biology texts to pursue research in the area of nucleic
acids. Aspects of DNA and RNA structure that may interest mathematicians, com-
puter scientists, or engineers, include DNA supercoiling; RNA folding prediction;
application of elasticity theory, topology, and knot theory to DNA structure pre-
diction and functional mechanisms; and using DNA as a parallel computer (e.g.,
[40,117,162,825,1155,1156]). The presentations here might introduce readers to
some of the exciting areas of research on DNA.
Besides the excellent texts [139, 195, 1080, 1191], the reader is invited to ex-
plore the wealth of related structural data on the nucleic acid database NDB and
the protein data bank/research collaboratory for structural bioinformatics resource
[128,1365].
As in the previous chapter, we abbreviate base pair and base pairs as bp and
bps, respectively.
6.2 DNA Sequence Effects
6.2.1 Local Deformations
What makes DNA such a rich resource of structural and functional information?
The subtle geometric, electrostatic, and mechanical differences inherent in each
nucleotide (e.g., C), bp (e.g., G–C), or bp step (e.g., AT in the 5
GCGATCGC 3
octamer) combine to affect patterns in local hydration and local helical parame-
ters, such as twist (Ω), tilt (τ ), roll (ρ), and slide (Dy) between two successive
bps (one bp-step), and propeller twist (ω) within each bp, as introduced in the
previous chapter and Figure 5.12.
Recall that twist is the relative rotation between two successive bps about the
helical axis as measured by the change in orientation between the corresponding
C1
–C1
vectors, projected down the helical axis.
Roll and tilt define the deformations between two bp planes along the long and
short bp axes, respectively, perpendicular to the helical axis. The sign conventions
are such that a positive roll angle opens up the minor groove side, while a positive
tilt angle opens up bps in the direction of the phosphate backbone of the first
strand.
Slide describes the translational displacement of neighboring bps along their
long axes, as measured between the midpoint of each long-bp axis. It is considered
positive when the direction is toward the first nucleic acid strand (i.e., positive e2
direction).
The propeller twist defines the angle between the planes of the two bases in
a hydrogen-bonded pair. A positive value corresponds to rotation of each base
in a counterclockwise manner when viewed from its attached sugar/phosphate
backbone [307].
166 6. Topics in Nucleic Acids Structure: DNA Interactions and Folding
Normal propeller twists are negative in DNA. This is a consequence of
base-pair stacking interactions along polynucleotide strands, as first discussed in
[309, 745]. (Note: some of the earlier sign conventions were opposite than the
current standard).
Propeller twisting can improve bp stacking interactions through enhanced van
der Waals, electrostatic, and hydrophobic contacts. It can also promote stability
through additional hydrogen-bonding contacts. Such contacts involve diagonally-
distant bases in two adjacent residues. Stability is enhanced since the distance that
would result from a perfectly stacked arrangement (perpendicular to the global
helix axis) is decreased due to propeller twisting. Regions of repeating AT bps
exhibit greater propensity than GC for propeller twisting [195,527].
The bending anisotropy in DNA was established over twenty years ago
[1096, 1454]. Namely, DNA prefers to bend more easily into the minor and ma-
jor grooves than other directions, and the preference to major or minor-groove
depends on the nucleotide sequence.
Such bending preferences already emerge for dinucleotide steps, as described
below. However, because sequence context is important, recent work has focused
on the effect of flanking sequences as well (see Subsection 6.2.3).
6.2.2 Orientation Preferences in Dinucleotide Steps
There are 10 unique dimer (dinucleotide) steps in DNA, conventionally specified
on the 5
→ 3
strand. Thus, associated with each dimer step is the complementary
bp step on the opposite strand (e.g., 5
[AC] 3
with 5
[GT] 3
); when the usual
orientation conventions (for tilt, roll, etc.) are applied to the complementary bp
step, the same deformations defined for the main strand result.
Local geometric and energetic trends can be associated with each bp step.
Trends can also be clustered into three broad classes: pyrimidine/purine(Pyr/Pur),
purine/purine (Pur/Pur), and purine/pyrimidine (Pur/Pyr) steps. (Often pyrim-
idines and purines are also denoted by Y and R for short). This is because common
properties such as residue size and electrostatics produce similar geometric and
energetic characteristics with these classes of dinucleotide steps.
The three Pyr/Pur steps are:
TA (TA) CA (TG) CG (CG);
the four Pur/Pur (Pyr/Pyr) steps are:
AA (TT) AG (CT) GA (TC) GG (CC);
and the three Pur/Pyr steps are:
AT (AT) GT (AC) GC (GC).
6.2. DNA Sequence Effects 167
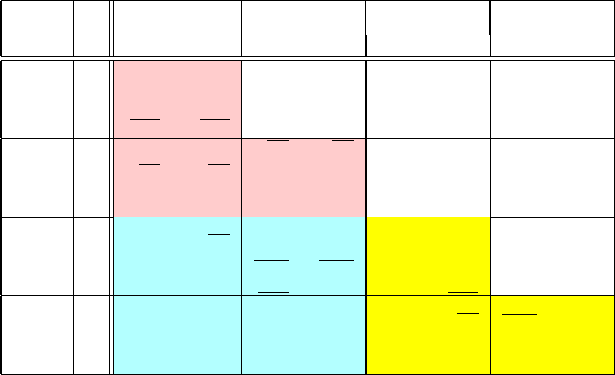
Table 6.1. Base-pair step parameters for roll (ρ), tilt (τ ), and twist (Ω) angles (in degrees)
for free [473] and protein-bound DNA [942] (denoted as DNA and DNA
+
, respectively), as
analyzed from crystal structures of DNA and DNA complexes from the NDB. The overline
and underline symbols indicate the largest and lowest values, respectively, associated with
each angular value in each class (free and complexed DNA). They help distinguish features
among the three dinucleotide classes as well as highlight similarities in patterns between
free and bound DNA of the same bp step. Note a tie for the minimal value of 0.7
o
roll for
AA and GT steps.
Nuc. Pur A Pur G Pyr T Pyr C
DNA DNA
+
DNA DNA
+
DNA DNA
+
DNA DNA
+
ρ 2.6 3.3
Pyr τ 0.0 0.0
T Ω 40.0 37.8
ρ 1.1 4.7 6.6 5.4
Pyr τ 0.6 0.5 0.0 0.0
C Ω 36.9 37.3 31.1 36.1
ρ 0.5 0.7 2.9 4.5 −0.6 1.1
Pur τ −0.4 −1.4 −2.0 −1.7 0.0 0.0
A Ω 35.8 35.1 30.5 31.9 33.4 29.3
ρ −0.1 1.9 6.5 3.6 0.4 0.7 −7.0 0.3
Pur τ −0.4 −1.5 −1.1 −0.1 −0.9 −0.1 0.0 0.0
G Ω 39.3 36.3 33.4 32.9 35.8 31.5 38.3 33.6
Let us first examine in Table 6.1 the published average roll (ρ), tilt (τ),
and twist (Ω) values for these 10 dinucleotide steps as collected over a set of
crystallographically-determined for free (unbound) DNA systems [473](firstcol-
umn in each dinucleotide entry) and protein-bound DNA systems [942] (second
column). For updated values, consult newer works by the authors.
This 4 × 4 table is symmetric as presented, so the upper triangle is not filled.
As ordered, the three top matrix entries (upper triangle cluster: TA, CA, CG) cor-
respond to Pyr/Pur bp steps; the four lower square entries (for AA, AG, GA, GG)
correspond to the Pur/Pur steps; and the three entries in the lower triangle cluster
(AT, GT, GC) correspond to Pur/Pyr steps. Note that the six maxima (overline
symbols) associated with roll, tilt, and twist values for both free and bound DNA
occur in the Pyr/Pur cluster; four extremes (lower bounds) are noted in the Pur/Pur
cluster, and three in the Pur/Pyr group. (The minimal value of 0.7
o
average roll
displacement occurs for both AA and GT steps).
This observation is related to the decreasing overall flexibility associated with
these dinucleotide steps, with Pyr/Pur steps most easily deformed. Note also the
similarity in trends for the free and protein-bound DNA entries.
The observed trends in each dinucleotide step can be explained in part by
the tendency of DNA to adjust local geometry so as to improve bp stacking
(overlap) interactions. Since the bp-plane areas are water insoluble, stacking

168 6. Topics in Nucleic Acids Structure: DNA Interactions and Folding
energies can be strengthened by reducing the area between successive bps via
local variations, and/or enhancing longer-range interactions through formation of
bifurcated hydrogen bonds (between successive bps on opposite strands).
See Box 6.1 for a discussion of the inherent flexibility of Pyr/Pur and Pur/Pur
steps, including the contribution of some examples of biological systems where
flexibility in Pyr/Pur steps has been noted.
Box 6.1: Inherent Flexibility of Pyr/Pur and Pur/Pur Steps
Pyr/Pur Favorable Configurations. Pyr/Pur steps (i.e., TA, CA, and CG) can produce
favorable stacking in one of two general configurations: a combination of large positive
roll and small negative slide,ornear-zero roll and small positive slide,bothinassociation
with propeller twisting in the two bps [195] (see Figure 5.12 of Chapter 5).
In the former arrangement (large positive roll and small negative slide), the purine bases
slide toward each other to improve the cross-chain stacking between them. The concomi-
tant large positive roll inclines the smaller pyrimidine partners to maintain the favored
propeller twist in both bps. In the latter (near-zero roll and small positive slide), the large
purine bases slide away from each other to avoid a steric clash between them since
the propeller twisting of the two bps brings them closer together. The roll is zero since,
in the same strand, stacked pyrimidines and purines remain parallel to one another in this
propeller-twisted arrangement. (See illustrations in [195]).
Given at least these two options for favorable stacking, the Pyr/Pur class of dinucleotide
steps generally displays a wide range of roll values when many structures are analyzed.
CG Example in E2. The high positive roll associated with a CG dinucleotide step has
been used to explain the importance of the central CG step in the DNA-binding sequence
of the bovine papillomavirus E2 protein [1074], whose sequence is ACCGACGTCGGT.
This step contributes to the needed deformation of the DNA-binding region — a large
overall bending toward the protein.
TG Example in CAP. The characteristic flexibility of another Pyr/Pur step, TG (equiv-
alent to the CA step discussed above), has also been used to explain the importance of the
central TG dinucleotide step in the DNA-binding sequence of the catabolite gene-activating
protein (CAP) in E. coli, a highly conserved dinucleotide segment in both monomers of
this dimer protein in different CAP-binding sites. The binding of this complex requires
substantial distortion of the DNA, largely modulated by a 40
o
kink at this central TG step
[163, 1143].
TA-Rich Regions in Functional Sites. Furthermore, the combination of an observed
small energetic barrier to unwinding in Pyr/Pur steps (though not fully understood) and the
intrinsic curvature associated with consecutive AT bps (equivalently, AA or TT dinucle-
otide steps) has been used to explain the prevalence of TA-rich regions in key functional
sites. Examples of such sites are TATATATA, also known as the adenovirus E4 promoter,
and TATAAAAG, in the adenovirus major late promoter, both of which binds to transcrip-
tion proteins. These proteins (like the TATA-binding protein TBP in eukaryotes) must

6.2. DNA Sequence Effects 169
unwind DNA, and hence the relatively low barrier to twisting is well exploited. Note also
that the standard, Watson-Crick (WC) AT bp has one fewer hydrogen bond than a WC GC
bp, which has three, and thus the AT bp should be easier to deform.
Pur/Pur Tendencies. It has also long been noted that the Pur/Pur AA dinucleotide step
prefers an orientation where the two successive AT bps are propeller-twisted (i.e., they
deviate from planar bp orientations) with near zero roll values (as bases remain parallel to
their neighbors on the same strand) [195]. This out-of-plane bending shortens cross-residue
distances between the adenines on one strand and the thymines on the opposite strand and
allows offset, cross-strand hydrogen bonds to form between an oxygen of thymine and a
nitrogen of adenine on the major groove side. This interaction also implies close contact
between an oxygen of thymine and carbon of adenine on the minor groove side.
The large positive roll characteristic of CG dinucleotide steps and low roll associated
with AA (or TT) steps are particularly consistent trends noted through various structural
analyses [274].
6.2.3 Orientation Preferences in Dinucleotide Steps
With Flanking Sequence Context: Tetranucleotide Studies
The importance of bp flexibility in the structural and functional properties of DNA
as well as of DNA/protein, DNA/RNA and other complexes combined with the
significance of flanking sequence effects has led to extensions in recent sequence-
effect studies from dinucleotide to tetranucleotide steps. The latter represent the
minimal structural unit that can reveal near-neighbor bp structural patterns.
There are 136 unique tetranucleotide steps, and unraveling these patterns has
become an international initiative called the Ascona B-DNA Consortium (ABC).
See [133, 321,705] for emerging information from MD simulations of many sol-
vated DNA systems containing all unique tetramer sequences. Such studies reveal
fine patterns for sequence context, and they are being used to compile a database
of geometric parameters for all dinucleotide steps to address all possible flanking
bp combinations.
For example, these modeling and simulation studies have revealed that while
Pyr/Pur steps are intrinsically flexible, they are least affected by the neighboring
units. Their flexibility is especially pronounced when these Pyr/Pur steps occur
near the intrinsically rigid Pur/Pur or Pur/Pyr steps due to their significant impact.
ABC-driven dynamics simulations are also uncovering a large degree of backbone
fluctuations and persistent canonical and non-canonical B-DNA sub-states [321].
6.2.4 Intrinsic DNA Bending in A-Tracts
Now consider all these sequence effects taken together. For heterogeneous, or
mixed sequence DNA, all these differing trends will very roughly average out on
a global scale for free DNA. In other words, these trends will accumulate slowly