Радченко В.Г. Биология пчел
Подождите немного. Документ загружается.

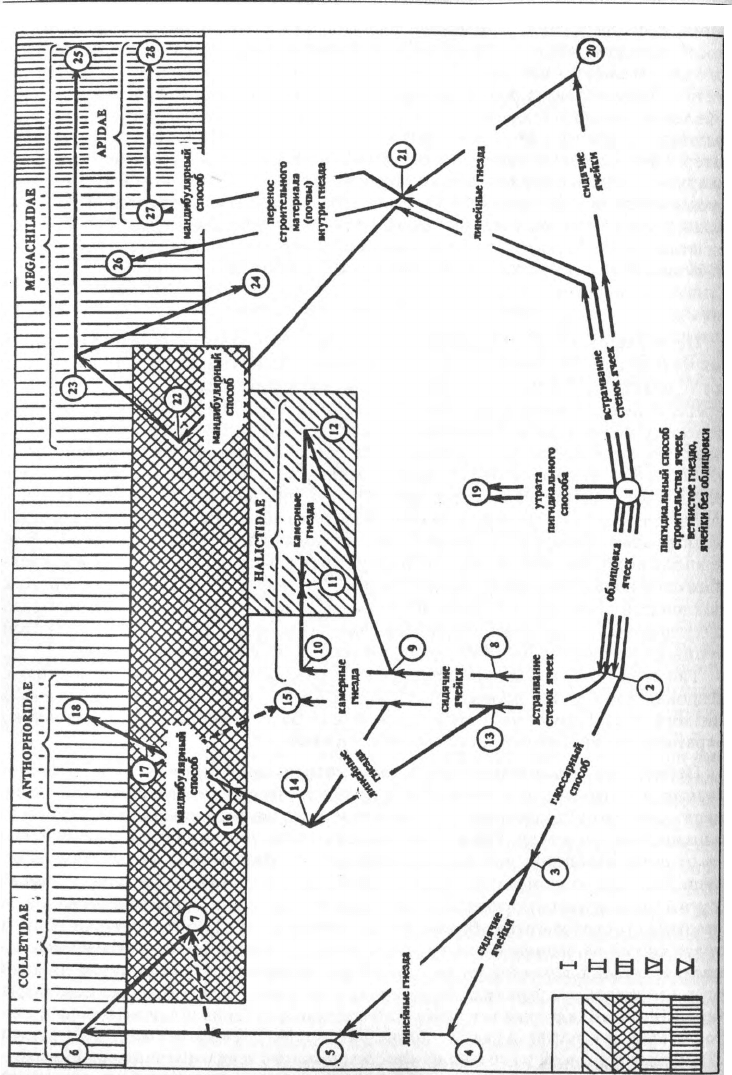
130 Глава 8. Эволюция гнездования пчел
Рис. 142

8.2. Смена субстрата гнездования
131
более сглаженные структуры). Среди самок всех других групп пчел, первично
поселяющихся в почве, такие шипы не встречаются.
Поселение Hylaeinae в растительных материалах. Недостаточно ясным вы-
глядит путь перехода к выгрызанию гнезд в мягкотелых стеблях растений, про-
изошедший у ряда Hylaeinae. С одной стороны, их непосредственные предки
могли поселяться в почве, где строили линейные гнезда, после чего перешли к
выгрызанию гнезд в мягкотелых растениях с дальнейшим переселением в готовые
полости. С другой стороны, они могли перейти к выгрызанию гнезд в раститель-
ных материалах уже после поселения в готовых полостях (например, из-за
недостатка последних), которому непосредственно предшествовало поселение в
почве.
К сожалению, отсутствие адекватных филогенетических схем для Hylaeinae
не позволяет использовать при решении этого вопроса имеющиеся факты о нали-
чии отдельных групп гилеин, приспособленных к поселению лишь в растительных
материалах. В частности, многие представители рода Hyleoides (например,
H.concinna, обитающий в Австралии) строят гнезда в растительном субстрате и
при этом обладают мощными выростами на передних и средних голенях и мощ-
ными мандибулами, что сопряжено только с выгрызанием гнезд в относительно
плотных растительных материалах. Не исключено, что исчезновение скопы у
Hylaeinae также было вызвано поселением в растительных материалах (раздел
7.2). На эту причину, в частности, указывает частичная редукция скопы на
задних голенях у коллетид подсемейства Xeromelissinae. Последнее обстоятель-
ство послужило даже основанием для ошибочного отнесения данной группы к
числу пчел, не имеющих собирательного аппарата (раздел 7.2). В действительно-
сти все изученные представители Xeromelissinae поселяются в растительных ма-
териалах и переносят пыльцу в основном на метасомальной скопе.
Рис. 142. Основные направления эволюции гнездования пчел
1 – первопчела (ближайший общий предок надсемейства Apoidea), Rophitinae, Dasypodinae,
некоторые Anthophorinae; 2 – некоторые Colletidae, Stenotritidae, Oxaeidae, некоторые Halictidae,
? Meganomiinae, группа Melitta, некоторые Anthophoridae; 3 – некоторые Colletes; 4 – отдельные
Colletes; 5 – многие Colletes; 6 – Xeromelissinae, многие Hylaeinae, некоторые Colletes; 7 – некоторые
Hylaeinae; 8 – некоторые Halictinae; 9 – многие Halictinae; 10 – некоторые Halictinae; 11 – отдельные
Halictini, многие Augochlorini; 12 – некоторые Halictini, отдельные Augochlorini; 13 – некоторые
Anthophorinae; 14 – отдельные Proxylocopa; 15 – некоторые Anthophorinae; 16 – Clisodon; 17 – многие
Xylocopinae; 18 – некоторые Xylocopinae; 19 – Fideliidae, Pararhophitini, Hesperapis trochanterata;
20 – отдельные Anthophorinae; 21 – некоторые Anthophorinae; 22 – Lithurginae; 23 – многие
Megachilinae; 24 – некоторые Megachilinae; 25 – отдельные Megachilinae; 26 – Ctenoplectra, Tetrapedia;
27 – некоторые Euglossinae; 28 – большинство Apidae.
I - гнезда в почве, II - гнезда в гнилой древесине, III - гнезда в стеблях и древесине, IV - гнезда в
готовых полостях, V - гнезда на открытых местах
Когда была закончена верстка настоящей книги вышли 2 статьи (R.-A. Alsina, C.D. Michener. Univ.
Kansas Sci. Bull., 1993. Vol. 55, N 4. P. 124-162; F.A. Silveira. Ibid, N 5. P. 163-173), в которых
пересмотрен статус и филогенетическое положение ряда групп высших («длиннохоботковых») пчел.
Дополнения и изменения, связанные с этими работами, мы уже не могли внести в корректуру книги.
Отметим только, что близость семейства Ctenoplectridae с трибой Tetrapediini (Anthophoridae) и с
семейством Apidae, показанная ранее по биологическим параметрам 1-м автором настоящей книги
(Радченко, 1992б, с. 22; также см. раздел 8.1 и 8.3), получает в работе Силвейры (Silveira, 1993, р.
169, 172-173) морфологическое подтверждение. Эти таксоны (в работе Силвейры принятые соответ-
ственно как Ctenoplectrini, Tetrapediini и «apine line») являются сестринскими и все основные биологи-
ческие преобразования, связанные с переходом к гнездованию в готовых полостях, происходили,
по-видимому, еще у их общего предка, т.е. одной, а не двумя параллельными линиями (ведущими на
рис. 142 к кружкам с номерами 26 и 27).
132
Глава 8. Эволюция гнездования пчел
8.3. Переход к поселению в готовых полостях и на открытых местах
Поселение в готовых полостях независимо возникало не менее 10 раз среди
представителей 5 семейств пчел (Colletidae, Ctenoplectridae, Megachilidae, Antho-
phoridae и Apidae). Общим для всех групп пчел, перешедших к строительству
гнезд в готовых полостях, было уменьшение зависимости ячеек от окружающего
субстрата, которое достигается разными способами в зависимости от того, какое
из четырех реализованных направлений было выбрано.
Первое направление касается представителей семейства Colletidae, которые
сооружают ячейки из относительно плотной целлофаноподобной пленки. Данная
пленка является не столько облицовочным, сколько строительным материалом
(раздел 6.2), позволяющим возводить ячейки «глоссарным» способом в готовых
полостях.
Вторым направлением, которым шли представители Ctenoplectra (Ctenoplec-
tridae) и Tetrapedia (Anthophorinae), очевидно, было встраивание стенок ячеек из
приносимой почвы, взятой в другом месте гнезда, подобно тому, как поступают
многие Halictinae и Anthophoridae (раздел 6.2). Появившаяся способность транс-
портировки строительного материала (почвы) позволила в дальнейшем исполь-
зовать и готовые полости, куда пчелы доставляют этот материал извне. При этом
сохраняется пигидиальный способ сооружения ячеек из почвы, которую самки
Tetrapedia и, очевидно, Ctenoplectra скрепляют с помощью растительного масла.
По третьему направлению эволюционировал предок Megachilinae и некото-
рые Xylocopinae. Оно осуществлялось через промежуточный этап выгрызания
гнезд в растительных материалах. Основу данного направления составил переход
от пигидиального к мандибулярному способу возведения ячеек, сначала из мате-
риала субстрата (из опилок), а затем и из иных материалов. Подробное обоснова-
ние рисуемого пути перехода представителей этих групп пчел к поселению в
готовых полостях излагается в следующем разделе.
И наконец, четвертым направлением, приведшим к поселению в готовых
полостях, двигался предок Apidae. В настоящее время сложно судить о возможных
промежуточных этапах на этом пути, поскольку остается неясным вопрос о пер-
вичности появления восковых желез у предка данного семейства. Как известно,
воск продуцируется всеми непаразитическими представителями 3 подсемейств
Apidae (Bombinae, Meliponinae, Apinae), тогда как самки 4-го подсемейства –
Euglossinae – не выделяют воск, а сооружают ячейки из смолы, смешанной с
другими материалами (раздел 6.1). Вместе с тем у самок Euglossa cordata
(Euglossinae) на предпоследнем метасомальном тергуме обнаружены структуры,
сходные по строению с восковыми железами других представителей Apidae (Cruz-
Landim, 1963, 1967). Эти структуры имеют очень маленькие размеры и не функ-
ционируют как железы, и поэтому в принципе их можно считать рудиментами
восковых желез, что должно интерпретироваться как первичность использования
воска всеми апидами и дальнейшая утрата его производства у эуглоссин. Если
принять такую версию, то вопрос о переселении предка апид в готовые полости
решается примерно таким же образом, как и для указанных выше групп из
Colletidae, которые возводят ячейки из плотного целлофаноподобного секретор-
ного материала.
Если же придерживаться другой, на наш взгляд, более реалистичной версии о
возникновении восковых желез лишь у указанных выше 3 подсемейств (при этом
вполне вероятно, что восковые железы даже среди этих подсемейств возникали
независимо, по крайней мере, 2 раза, о чем свидетельствует разная локализация

8.4. Эволюция гнездования Megachilidae
133
данных желез на теле у самок; раздел 6.1 ), а структуры, обнаруженные у Е. corda-
ta, рассматривать как зачатки желез (что, кстати, не отрицается; см.: Cruz-
Landim, 1967), то появление у предка Apidae в качестве строительного материала
именно смолы, переносимой в обножках, имеет логически более последователь-
ное обоснование.
Все те Apidae, которые используют при строительстве гнезд минеральные или
растительные материалы, приносят их в виде обножек, а не с помощью мандибул
(Zucchi et al. 1976; Sakagami, 1976). Поэтому начальная эволюция гнездования
этой группы, скорее всего, шла по тому же пути, каким двигались Ctenoplectra и
Tetrapedia (см. выше). При этом предок Apidae при строительстве ячеек мог
начать использовать для скрепления частиц минеральных веществ растительную
смолу взамен, например, растительного масла. В подтверждение выдвигаемой
гипотезы, помимо чисто логического, свидетельствует и морфологический аргу-
мент: преобразование скопы на ногах у Apidae в специальные «корзиночки»
(раздел 2.5), скорее всего, стало следствием перехода к транспортировке смолы,
которую было трудно счесывать с опушенной скопы. Добавим, что многие апиды,
перешедшие к изготовлению ячеек из воска, также собирают в «корзиночки»
смолу и используют ее при строительстве (разделы 9.5-9.6).
Переход к сбору смолы привел и к изменению самого способа строительства
ячеек от пигидиального к мандибулярному, поскольку обработка смолы с по-
мощью пигидиальной пластинки крайне затруднена. В результате у этой группы
пчел появилась возможность лепить ячейки не только внутри готовых полостей,
но и на открытых местах. Дальнейшая эволюция гнездования Apidae сопровож-
далась переходом к преимущественному использованию секретируемого воска,
что, благодаря его высокой пластичности, позволило видам Apis достичь вершины
совершенства в строении гнезда среди пчел.
8.4. Эволюция гнездования Megachilidae
Это семейство характеризуется наибольшим разнообразием в местах строи-
тельства гнезд и используемых материалов, поэтому рассмотрение эволюции его
гнездования заслуживает особого внимания.
Прежние версии. Высказаны два разных мнения о происхождении у мегахи-
лид основных типов гнезд.
Как можно понять из работ Малышева (1911, 1913, 1936), он предполагал
существование двух параллельных путей в развитии их гнезд. Так, одни пчелы,
применяющие для изготовления ячеек минеральные вещества (в основном по-
чву), первично поселялись в почве, где самки начали встраивать стенки ячеек из
этого субстрата. Переселившись в готовые полости в растительном субстрате, они,
по мнению Малышева, сохранили способность лепить ячейки из почвы, доставляя
ее в виде грязи в гнездо извне. При этом поселение в растительном материале
позволило пчелам перестать строить из приносимой почвы боковые стенки ячеек,
роль которых стали выполнять естественные стенки полости. Другие пчелы, ис-
пользующие для строительства ячеек растительные материалы, на его взгляд,
первоначально поселялись в различных полостях в растениях, где сооружали из
пережеванных растительных материалов лишь перегородки между ячейками. В
ходе дальнейшей эволюции такие пчелы стали сооружать из растительных мате-
риалов все стенки ячеек, что позволило им разнообразить места устройства гнезд,
в том числе перейти к гнездованию в почве. Представления Малышева в значи-
тельной степени противоречат монофилетичности не только подсемейства Mega-
chilinae в целом, но и всех входящих в него триб и родов, поскольку среди их
134
Глава 8. Эволюция гнездования пчел
представителей широко встречается использование разных материалов (мине-
ральных или растительных), в том числе и смешанных.
Несколько иного мнения на происхождение гнезд у мегахилин придерживался
Гутбир (1916), считавший, что у этой группы сооружение ячеек не только из
минеральных, но и из растительных материалов, принесенных извне, берет нача-
ло в почве, и только после этого произошла их миграция в растительный субстрат.
Эйкуорт с соавторами (Eickwort et al., 1981), рассматривая эволюцию гнездо-
вания всех Megachilidae, также пришли к выводу, что эта группа возникла как
поселяющаяся в почве, где у них развилась способность собирать для облицовки
ячеек растительные материалы (например, вырезанные кусочки листьев или
лепестков) как альтернатива к использованию секреторной облицовки для влаго-
непроницаемой защиты ячеек.
В качестве основного примера, подтверждающего их взгляды, Эйкуорт с соав-
торами приводят особенности гнездования представителей подсемейства
Fideliinae, относимого ими к Megachilidae [но в действительности являющегося
самостоятельным семейством Fideliidae, не имеющим ни одной надежной синапо-
морфии с Megachilidae (Michener, Greenberg, 1980; Michener, 1983а)]. Неубеди-
тельно также и объяснение Эйкуорта, что ограниченность распространения
Fideliidae аридными зонами связана лишь с отсутствием у этих пчел облицовки
ячеек, из-за чего увеличивается опасность гибели потомства от влажности и/или
грибковых заражений запасенной пищи. Известны несколько групп пчел (напри-
мер все Rophitinae, Dasypodinae, некоторые Panurginae и Anthophorinae), кото-
рые, поселяясь в почве, не облицовывают ячейки секреторными материалами
(раздел 6.2) и тем не менее благополучно существуют в умеренных зонах. При
этом ни в одной из этих групп не появилось изготовление стенок ячеек из расти-
тельных материалов.
В подтверждение первичности выкапывания гнезд предком Megachilidae в
почве указанные авторы приводят неудачный пример с Lithurginae, выгрызающи-
ми гнезда в растительном субстрате. Кроме того, Гутбир (1916) и Эйкуорт с
соавторами (Eickwort et al., 1981) расценивают факт строительства многими ме-
гахилинами всех стенок ячеек из растительных материалов в таком, на первый
взгляд, малогигроскопичном материале, каким является древесина, как косвен-
ное доказательство первичности возникновения облицовки ячеек растительными
материалами именно в почве. В то же время Гутбир, рассматривая причины
наличия секреторной облицовки всех стенок ячеек у Clisodon furcatus (Antho-
phoridae), сам связывал это с большой гигроскопичностью гнилой древесины, в
которой данный вид выгрызает гнезда, а Малышев ( 1913), в свою очередь, отмечал
чрезвычайную гигроскопичность сухой сердцевины стеблей растений, часто ис-
пользуемых пчелами для поселения. Добавим также, что сохранение строитель-
ства всех стенок ячеек даже в действительности малогигроскопичных материалах
можно легко объяснить большой пластичностью гнездостроительных инстинктов
мегахилин, часто использующих самые разнообразные места для устройства
гнезд (раздел 5.1 ), в том числе, например, различные трещины, где пчелы должны
уметь возводить все стенки ячеек.
В качестве дополнений, которые действительно можно было бы использовать для подтверждения
приведенной выше гипотезы Гутбира и Эйкуорта с соавторами, укажем следующие примеры. В
частности, по данным Мариковской (1968), Osmia flavicornis изготавливает в почве (на засоленном
такыре) ветвистые гнезда, располагая в них по одной ячейке на конце каждого бокового хода. При этом
ячейки имеют утрамбованные и отполированные земляные стенки. На дне ячейки самка помещает
шаровидный хлебец, откладывает на него яйцо и запечатывает ячейку земляной крышкой.
Еще более интересные сведения приведены Жанвье (Janvier, 1955) о строении гнезда Osmia
arequipensis, обнаруженного в Боливии. Данное гнездо, находившееся в почве, имеет ветвистую форму
с одной ячейкой на конце каждого бокового хода. Ячейки овалоидной формы с гладкими, лощенными
8.4. Эволюция гнездования Megachilidae
135
стенками, сверху покрытыми лепестками мальвы. При этом, по мнению Жанвье, самка сглаживает и
полирует стенки ячеек с помощью метасомы.
Такие необычные для мегахилин гнезда могли бы служить более убедительными примерами для
утверждений не только о первичности гнездования этой группы в почве, но и о возникновении здесь
облицовки ячеек растительными материалами. Вместе с тем описанные выше гнезда O.flavicornis
принадлежали, однако, скорее всего, другому виду пчел или, как и в случае с O.arequipensis, были
результатом поселения в старом гнезде другого вида. В частности, одинаковая с O.arequipensis архи-
тектура гнезда приведена в той же работе Жанвье для «Halictus» pacis, относимого в настоящее время
к роду Caenohalictus. Это доказывается прежде всего тем, что самки всех Megachilidae не способны
полировать стенки ячеек, поскольку у них отсутствует пигидиальная пластинка, с помощью которой,
собственно, и становится возможной такая обработка почвенных стенок.
Столь подробный критический анализ приведенных примеров вызван тем, что
нами отрицается не только использование растительных материалов для строи-
тельства ячеек предками Megachilidae, поселявшимися в почве, но и возможность
самого становления семейства при гнездовании в почве.
Новая гипотеза. Становление семейства Megachilidae, на наш взгляд, про-
изошло после переселения его предка в растительный субстрат. В пользу данного
утверждения свидетельствует, во-первых, как уже отмечалось выше, редукция
скопы на задних голенях у Megachilidae, которая, вероятнее всего, вызвана вы-
далбливанием гнезд в растительных материалах. Во-вторых, выявленной нами
синапоморфией этого семейства являются шипы на вершинах передних и средних
голеней самок (такие шипы иногда преобразованы в более или менее закруглен-
ные пластинки у видов, вторично вернувшихся к поселению в почве; например,
у Trachusa). Наличие этих шипов у пчел связано с гнездованием в растительных
материалах и не встречается среди самок других групп пчел, первично поселяю-
щихся в почве. В-третьих, строение мандибул самок большинства Megachilidae
(за исключением нескольких специальных случаев, как, например, Chelostoma
maxillosum), скорее, приспособлено к выдалбливанию гнезд в растениях, чем в
почве. Кроме того, ни в одной другой группе пчел, выкапывающих гнезда в почве,
не возник мандибулярный способ строительства ячеек. И наоборот, все случаи
перехода к мандибулярному способу сооружения ячеек (раздел 6.2) связаны либо
с выгрызанием гнезд в растительных материалах, либо с поселением в готовых
полостях, также обычно находящихся в растениях. В-четвертых, Lithurginae,
считающиеся наиболее генерализованным подсемейством мегахилид, гнездятся
исключительно в растительном субстрате. И наконец, в-пятых, представляется
более реалистичным и логически последовательным путь перехода к использова-
нию растительных материалов пчелами, поселявшимися в растительном субстра-
те, а не в почве.
Рассмотрим последний аргумент подробнее. Перейдя от выкапывания гнезд в
почве к их выгрызанию в растительном субстрате, предок мегахилид стал исполь-
зовать этот субстрат (опилки) для устройства ячеек и одновременно отказался от
пигидиального способа строительства, начав использовать с этой целью мандибу-
лы, которыми они выгрызают полости и укладывают (часто в пережеванном виде)
строительный материал. Именно на этом этапе остановились Lithurginae. Прочие
Megachilidae (подсемейство Megachilinae) полностью перешли к поселению в
готовых полостях. Этому способствовали относительная ограниченность числа
растительных материалов, в которых пчелы способны устраивать гнезда, а также
возможность использования пустотелых стеблей растений, облегчающих задачу
выдалбливания гнезда. Процесс протекал примерно следующим образом.
Сначала пчелы использовали готовые полости в растениях, в которых они
продолжали строить ячейки из опилок, снимаемых со стенок полости. Затем они
начали транспортировать опилки также из мест, расположенных снаружи гнезда,
после чего полностью перешли на доставку строительных материалов извне.
136
Глава 8. Эволюция гнездования пчел
В свою очередь, независимость строительных материалов от субстрата гнезда
позволила резко расширить диапазон используемых материалов и привела (в том
числе) к сбору не только растительных материалов (таких как частицы стеблей,
листьев, смол и т.п.), но и минеральных (почва, камешки). В результате отдель
ные виды фактически во всех родах мегахилин независимо перешли к использо
ванию самых различных строительных материалов.
В пользу рисуемого пути эволюции гнездования мегахилид свидетельствует и то, что аналогичный
процесс протекает параллельно у Xylocopinae (Anthophoridae). Так, многие ксилокопины, например
Ceratina okinawana и С. flavipes (Maeta, Gôukon, 1980), Xylocopa artifex, X. auripennis и X. ciliata (Iwata,
1964; Sakagami, Laroca, 1971), наряду с выдалбливанием гнезд в плотных растительных материалах,
нередко поселяются в пустотелых стеблях растений (тростнике и бамбуке). При этом в качестве
строительного материала для перегородок между ячейками они используют опилки, отгрызенные с
внешних сторон стеблей. Опилки доставляются в гнездо с помощью мандибул, что, очевидно, также
стало одной из основных причин перехода этих пчел к мандибулярному способу строительства ячеек.
Вполне возможно, что у ксилокопин может также развиться способность транспортировать для соору
жения ячеек и другие материалы.
Основные направления эволюции. Дальнейшая эволюция гнездования
Megachilidae осуществлялась в двух направлениях. Первое направление связано
с упрощением строительства ячеек. Оно ведет, прежде всего, к прекращению
возведения боковых стенок ячеек, роль которых стали выполнять естественные
стенки полости. При этом у пчел обычно сохраняется способность к сооружению,
по крайней мере, части боковых стенок, например при поселении в больших или
щелеподобных полостях. Такое поведение, в частности, отмечено у Osmia coeru
lescens (Verhoeff, 1891). Наибольшее упрощение в строительстве ячеек привело
к исчезновению и перегородок, т.е. к появлению гнезд вообще без ячеек, обнару
женных у Metallinella brevicornis (Радченко, 1978). Основную роль в возникнове
нии таких гнезд, вероятнее всего, сыграло то, что самка M. brevicornis (в отличие
от большинства других видов одиночных пчел) откладывает каждое яйцо еще до
окончания формирования провизии. При этом яйцо помещается в специальную
воздушную камеру внутри корма. В результате в поведении M. brevicornis от
сутствует обязательное для подавляющего большинства других одиночных пчел
запечатывание ячейки сразу после откладки яйца, а продолжающееся наполне
ние ее пищей привело к выпадению акта строительства и крышки ячейки.
Тип гнезда, обнаруженный у M. brevicornis, конвергентно развивается и у
некоторых других мегахилид, также откладывающих яйца в период запасания
корма, в частности у Lithurge fuscipenne (Гутбир, 1916), L. atratiforme (T.Houston,
1971), L. chrysurum Roberts, 1978) и y Megachile policaris (Krombein, 1967). Вполне
вероятно, что гнезда без ячеек могут быть найдены и среди прочих видов, поме
щающих яйца в корм до окончания его фуражировки, например у видов подрода
Cephalosmia из рода Osmia (рис 132). Попутно отметим, что взгляд Робертса
(Roberts, 1978) на отсутствие перегородок у видов Lithurge, как на шаг в направ
лении к «прогрессивному» кормлению личинок (раздел 6.4), представляется не
убедительным, поскольку эти пчелы не имеют доступа к своим личинкам,
располагающимся внутри корма.
В отличие от описанных выше тенденций, направленных на упрощение работ
по сооружению ячеек, второе направление в эволюции гнездования Megachilinae,
наоборот, связано с усложнением таких работ. В частности, использование в
строительстве ячеек прочных материалов, таких как комочки почвы, камешки,
песчинки и т.п., скрепленных смолой и (или) секретом слюнных желез, позволил
ряду видов отказаться от гнездования в готовых полостях и перейти к лепке ячеек
на открытых местах. Предположение Гутбира (1916) о возникновении гнезд от
крытого типа непосредственно у пчел, поселявшихся в почве, на наш взгляд,
малореалистично. Именно поселение вне почвы, особенно в щелеподобных поло
8.5. Основные тенденции в эволюции 137
стях, где часть стенок ячеек непосредственно подвержена влиянию внешних
условий, способствовало использованию пчелами строительных материалов, ус
тойчивых к неблагоприятным условиям.
Например, отмечены случаи строительства не только части стенок, но и отдельных ячеек вне
полостей самками Hoplitis adunca и Anthocopa bidentata (Радченко, 1981б). Обычно эти виды
поселяются в различных полостях в древесине, в том числе щелеподобных, где строят ячейки из
комочков почвы и камешков, скрепленных смолой (Maneval, 1932; Grandi, 1935). Правда, в тропиках
отмечены гнезда Anthidium truncatum с ячейками из ватоподобного материала, расположенными
открыто на листьях растений (Friese, 1909). По нашему мнению, такие гнезда, скорее всего, были в
действительности устроены пчелами внутри молодых завернутых листьев, поскольку трудно даже
представить, как могли самки возвести ячейки из «ваты» на открытом месте, не говоря уже о высокой
гигроскопичности этого материала, требующего дополнительной защиты в виде стенок полости.
И наконец, в качестве отдельного, третьего, направления в эволюции гнездо
вания Megachilinae можно рассматривать вторичный переход отдельных видов к
выкапыванию гнезд в почве или к выгрызанию их в растительных материалах
(чаще всего в мягкотелых стеблях растений). Такой возврат к самостоятельному
изготовлению полостей для устройства гнезд, очевидно, связан с постоянным
недостатком подходящих готовых полостей для устройства гнезд в естественных
условиях. Вероятно, эта же причина сыграла основную роль и в переходе к
сооружению гнезд открытого типа.
8.5. Основные тенденции в эволюции
Анализ эволюционных изменений в биологии пчел, даже ограниченный толь
ко 8 рассмотренными признаками, характеризовавшими первопчелу (табл. 3),
показывает, что эти изменения могут быть объяснены двумя общими адаптивны
ми тенденциями, свойственными для эволюции и большинства других процвета
ющих групп животных: 1) обеспечение условий для экспансии, 2) усиление
заботы о потомстве.
Обеспечение экспансии. Первая тенденция отражает различные адаптации,
обеспечивающие расширение возможностей для обитания групп как в географи
ческом, так и в биотопическом аспектах, а также способствующие повышению
численности и плотности организмов. Эти адаптации в итоге приводят к разнооб
разию жизненных стратегий и ресурсов, которые способны осваивать и усваивать
организмы данной группы. Географическая и экологическая экспансия у пчел
обеспечивалась прежде всего расширением возможностей для гнездования, что
достигалось как за счет овладения новыми субстратами для устройства гнезд
(песчаная почва, древесина, мягкотелые или полые стебли, раковины моллюсков
и другие «готовые» полости), так и благодаря ослаблению зависимости от конк
ретных субстратов у тех видов и групп пчел, которые перешли к устройству ячеек
из различных приносимых (кусочки листьев или лепестков, растительный «пух»,
смола, глина и т.п.) или секретируемых материалов в готовых полостях или на
поверхности субстрата. Меньшее значение имело расширение диапазона кормо
вых ресурсов – сбор пыльцы с цветков растений, которые первоначально «не
предназначались» для пчел (один из немногих примеров – переход к сбору очень
крупных пыльцевых зерен в зобик, что, в частности, привело к утере собиратель
ного аппарата у некоторых Colletidae; раздел 7.2), и использование в качестве
корма для личинок, помимо пыльцы и нектара, также растительных масел (раз
дел 2.1 ) или такой специальной пищи, как «маточное молочко» у медоносных пчел
и ткани погибших животных у Trígona hypogea (Roubik, 1982).
Конечно, трофика пчел определяет специфику соответствующей им жизнен
ной формы – насекомогоантофила. Более того, именно сопряженная эволюция с
цветковыми растениями позволила пчелам достигнуть процветания. Но к такой
138
Глава 8. Эволюция гнездования пчел
жизненной форме принадлежала и первопчела, поэтому в строгом смысле изме-
нения по данному биологическому признаку в пределах пчел не произошло (не
считая указанного выше единичного случая). Исключением из этого утверждения
не является и переход некоторых групп пчел из разных таксонов к клептопарази-
тизму на других пчелах, так как имаго и личинки клептопаразитических пчел
питаются той же пищей (пыльцой и нектаром), что и остальные пчелы. Следова-
тельно, клептопаразитизм у пчел, в какой бы форме он не был выражен («сфекод-
ный» тип, «номадный» тип, «социальный паразитизм» шмелей-кукушек или
«облигатное грабительство» некоторых Meliponinae; раздел 3.4), по своей сути
является лишь специфическим или частным вариантом внутри жизненной формы
«насекомое-антофил». Вместе с тем клептопаразитизм, конечно, можно рассмат-
ривать как одно из средств, способствовавших если не экспансии пчел, то, по
крайней мере, их таксономическому, морфологическому и биологическому раз-
нообразию, числу и разнообразию занимаемых ими экологических ниш.
Забота о потомстве. Вторая из сформулированных выше главных адаптивных
тенденций в эволюции пчел – усиление заботы о потомстве – включает различные
изменения в их биологии:
1) адаптации, направленные на защиту личиночного корма и самих преима-
гинальных фаз от высыхания и переувлажнения – облицовка ячеек гигрофобными
секретируемыми материалами или растительными маслами (разделы 6.2-6.3),
встраивание стенок ячеек из растительных и/или минеральных материалов
(Ctenoplectridae, Megachilinae, Tetrapediini), лепка ячеек из смолы (Euglossinae)
и/или воска (прочие Apidae), отчасти расположение ячеек группами в камерах,
облицовка хлебцев секреторным веществом (раздел 6.4) ;
2) адаптации, направленные на предохранение корма от загрязнения почвой
– встраивание и/или облицовка стенок ячеек (раздел 6.2), специальные средства
для защиты жидкого корма (разделы 6.4, 7.7) ;
3) адаптации, направленные на защиту потомства и запасаемого корма от
микробных инвазий – использование секреторных выделений или веществ расти-
тельного происхождения, обладающих антимикробной активностью, для обра-
ботки стенок ячеек и запасаемого корма (раздел 6.3) ;
4) адаптации, направленные на защиту преимагинальных фаз от паразитов и
хищников, а также от механического уничтожения – устройство гнезд глубоко в
земле (например, у ряда видов Andrena и Stenotritidae; раздел 5.2), рассредото-
ченная (неагрегированная) закладка гнезд, что связано с преодолением инстин-
кта строить их вблизи материнского гнезда и с активным расселением (например,
у многих Andrena и у Melitta leporina; раздел 5.1 ), строительство гнезд с ячейками,
расположенными далеко друг от друга в горизонтальной и/или вертикальной
плоскостях или сооружение одноячейковых гнезд в очень рыхлом субстрате (раз-
дел 5.3), устройство ячеек в готовых полостях в твердом субстрате (раздел 5.1),
встраивание добавочных стенок ячеек из материала субстрата (раздел 6.2), изго-
товление временных крышек, которыми пчела закрывает ячейки после приноса
очередной порции корма (раздел 6.4; эта адаптация также связана и с обеспече-
нием чистоты запасаемого корма; см. выше пункт 2), маскировка входного отвер-
стия гнезда (раздел 5.1) ;
5) адаптации, обеспечивающие комплексную защиту потомства и заботу о
нем – инспектирование самкой ячеек, ее работа «сторожем» в гнезде с развиваю-
щимся потомством и, наконец, различные формы социальной жизни (гл. 9-13).

139
ЧАСТЬ III. СОЦИАЛЬНАЯ ЖИЗНЬ:
ФОРМЫ ПРОЯВЛЕНИЯ, ПРОИСХОЖДЕНИЕ
И ЭВОЛЮЦИЯ
Надсемейство Apoidea является одной из 5 групп насекомых (также термиты
– отряд Isoptera, муравьи – надсемейство Formicoidea, большинство складчато-
крылых ос подсемейства Vespinae и роющая оса Microstigmus comes из семейства
Sphecidae), у которых известна настоящая социальная жизнь («эусоциальность»,
раздел 9.1). В каждой из этих групп социальная жизнь возникала независимо.
Более того, среди пчел эусоциальность появлялась многократно. В настоящее
время она известна в следующих семействах пчел: Halictidae (в трибах Augochlo-
rini и Halictini; гл. 10), Anthophoridae (в трибах Allodapini, Ceratinini и Xylocopini;
раздел 9.4) и Apidae (все 4 подсемейства: Euglossinae – раздел 9.4, Bombinae – гл.
11, Meliponinae – раздел 9.5 и Apinae – раздел 9.6). В отличие от пчел все виды
муравьев и термитов социальные (или реже паразитируют на других видах).
Другим отличием этих двух групп от пчел является обладание ими лишь одной
формой социальности, а именно – эусоциальностью. Кроме того, многим терми-
там и муравьям свойственно наличие морфологически четко различимых каст
среди нерепродуктивных (рабочих) особей, чего нет у пчел, даже достигших
высших уровней эусоциальности. Зато среди пчел представлены различные фор-
мы социальности, в том числе такие, которые занимают промежуточное положе-
ние между одиночным и эусоциальным образом жизни. Примерно такое же
разнообразие форм социальности характерно и для складчатокрылых ос, однако
именно изучение биологии пчел позволило реконструировать основные пути и
факторы становления социальности у жалоносных перепончатокрылых (гл. 12-
13), к которым относится подавляющее большинство социальных видов насеко-
мых.
ГЛАВА 9. ФОРМЫ ПРОЯВЛЕНИЯ СОЦИАЛЬНОЙ ЖИЗНИ
У ПЧЕЛ
9.1. Терминология и классификация форм социальности
Эусоциальность. Под настоящей социальностью, или эусоциальностью
(eusociality), большинство современных авторов (E.Wilson, 1971,1975; Hermann,
1979; Starr, 1979, 1985; Andersson, 1984; Кипятков, 1985, 1986; Росс и др., 1985;
Fletcher, Ross, 1985; Gadagkar, 1985b; Schuster, Schuster, 1985; Захаров, 1991, и
др.) понимают такие случаи совместного обитания в одном гнезде взрослых осо-
бей, когда, во-первых, они представлены двумя генерациями, во-вторых, имеется
кооперация в выполнении различных работ и, в-третьих, репродуктивные функ-
ции распределены между ними (наличие каст). Сразу отметим, что в дальнейшем
тексте под термином «генерация» (generation, поколение) понимается совокуп-
ность особей, относящихся к одной и той же ступени в генеалогической последо-
вательности «родители – дети – внуки и т.д.» Выводком (brood) называется порция
потомства матери, по времени закладки и отрождения отдаленная от других
порций. Термин «эусоциальность» впервые был введен Батрой (Batra, 1966с, р.
375). Некоторые авторы (например: Michener, 1969а, р.305, 1974, р.46; Кипятков,