Меннинг О. Поведение животных. Вводный курс
Подождите немного. Документ загружается.

260 Глава 7
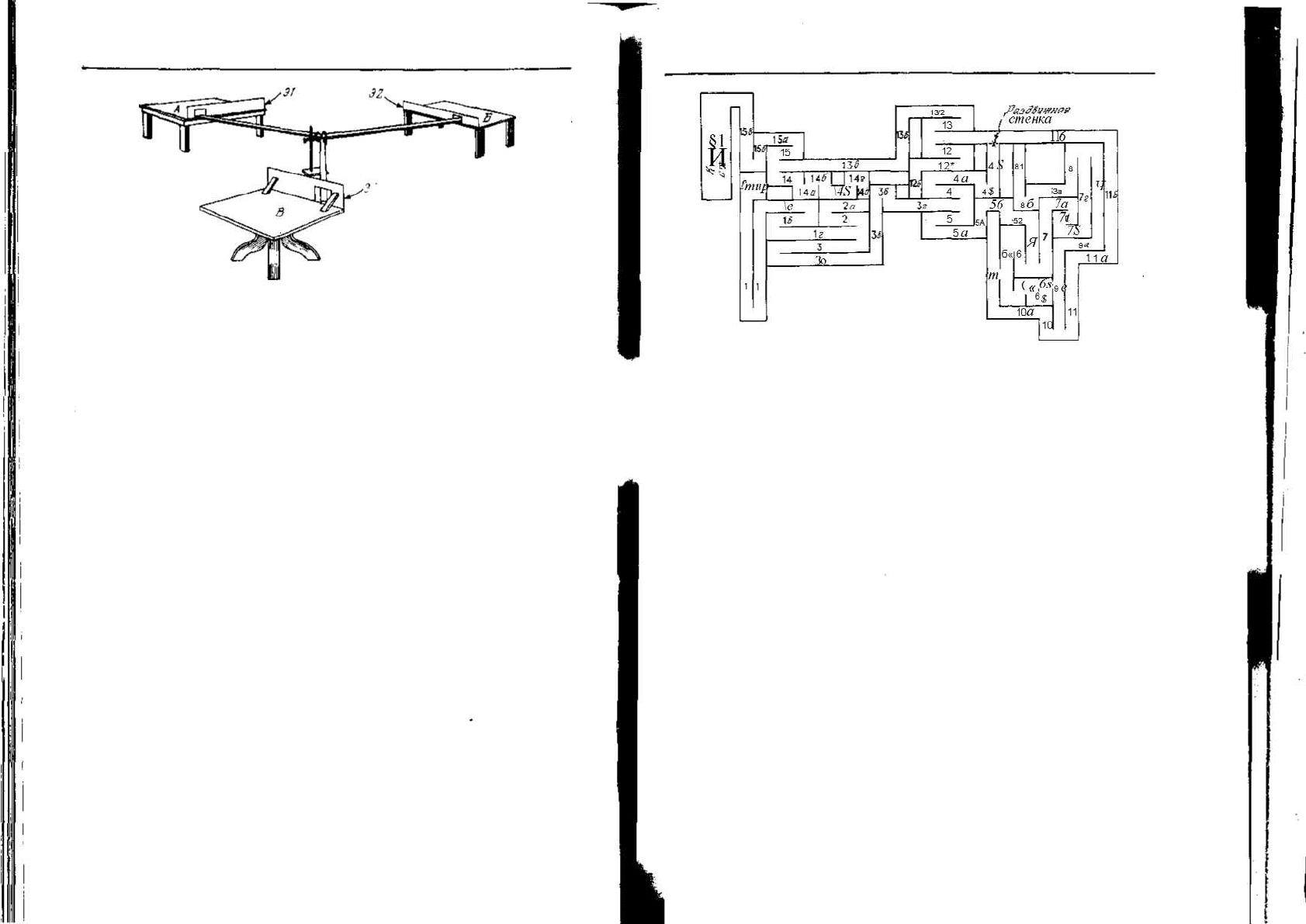
Рис. 7.5. Одна из установок, предложенных Майером для тестирования способ-
ности крыс к рассуждению [321]. Она состоит из трех дорожек длиной 8 футов
(243,8 см), расходящихся из одной центральной точки. Каждая дорожка заканчи-
вается столиком, отличающимся от остальных по размеру, форме и типу. На сто-
ликах устанавливались деревянные экраны (Э1, Э2, ЭЗ) таким образом, чтобы с
одного из них нельзя было видеть, что делается на остальных. После того как кры-
са обследовала все столики и дорожки, ее кормили, например на столике А. Затем
крысу помещали на один из двух других столиков, например S, и отпускали. До-
стигнув центра установки, крыса могла выбрать один из двух путей — на столик
А или столик Б Правильным решением считался выбор столика А. Осмотр уста-
новки предшествовал всем тестам, причем каждый раз крысу кормили, а затем вы-
пускали снова с разных столиков. При случайном выборе доля правильных реше-
ний равна 50%, однако у некоторых крыс она гораздо выше.
дит быстрее, чем если бы животное действительно производило про-
бы, однако возможно, что оно совершает эти пробы «в уме». При
этом предполагается, что животное может формировать понятия и
«рассуждать». Изучение способности животного к «рассуждению»
вряд ли существенно отличается от изучения его способности
к инсайту.
Майер [322] определяет рассудочную деятельность как «... спо-
собность спонтанно комбинировать две или более отдельные реак-
ции при образовании новой, которая эффективна для достижения
желаемого результата». Майер с сотрудниками проверяли способ-
ности крыс к «рассуждению» разнообразными способами, которые
основывались главным образом на том, что крысы находили обход-
ной путь вокруг преграды или кратчайший путь через лабиринт.
Два типичных примера таких задач иллюстрируют рис. 7.5 и 7.6.
Они отражают основные принципы построения соответствующих
экспериментов. Во-первых, во всех случаях существует некоторый
период исследования обстановки, который предшествует основному
опыту. Это свидетельствует о том, что решение задачи в значитель-
ной мере зависит от латентного обучения. Во-вторых, решение за-
дачи оценивается по его скорости: если животное делает ошибки и
нуждается в дополнительном исследовании обстановки, то мы
Обучение
277
Рис. 7.6. Сложный лабиринт, использованный Шепардом для оценки способности
животных к рассуждению [322]. После завершения обучения крыс, секцию, поме-
ченную знаком X, удаляют, тем самым превращая тупик в проход. Обнаружив из-
менение во время прохода по коридору Ив и короткого исследования секции 4в,
некоторые крысы в следующей пробе идут в 4-й отсек (а оттуда в 4а, 46, 4в) вместо
5-го.
заключаем, что оно использует более примитивную форму обучения
методом проб и ошибок.
Вероятно, неразумно придавать слишком большое значение обыч-
ным экспериментам с обходом преграды, потому что некоторые на-
секомые обнаруживают необыкновенную способность решать задачи
такого типа. Так, Хебб и Уильяме [207] считают, что хорошим кри-
терием «интеллекта» животных служит преодоление ряда передвиж-
ных препятствий в открытом пространстве с фиксированным местом
старта и цели. Чтобы животные не могли руководствоваться поло-
жением преград относительно цели, их передвигали в каждой пробе
на новое место, так что животное должно было каждый раз решать
новую задачу поиска обходного пути (рис. 7.7). Оценка решения
проводилась по степени прямизны линии пробежки от стартовой
камеры к цели. В модифицированном виде эта методика широко ис-
пользовалась экспериментальными психологами. В настоящее вре-
мя не вызывают сомнений представленные Торпом [455] доказатель-
ства того, что оса Ammophila решает упомянутые выше задачи так
же хорошо, как и собака! В естественных условиях точкой старта
для осы служит место, где она ужалила гусеницу, а целью — гнез-
до, куда она должна доставить добычу. Она никогда прежде не пере-
двигалась по земле, а лишь совершала рекогносцировочный полет
вокруг гнезда. Если во время движения осы на ее пути помещать
преграды, она обычно на некоторое время изменяет направление,
так чтобы обогнуть преграду с минимальным усилием. Удивитель-
ная способность осы к поиску обходных путей основывается, ве-
260
Глава 7
Обучение
278
я
у
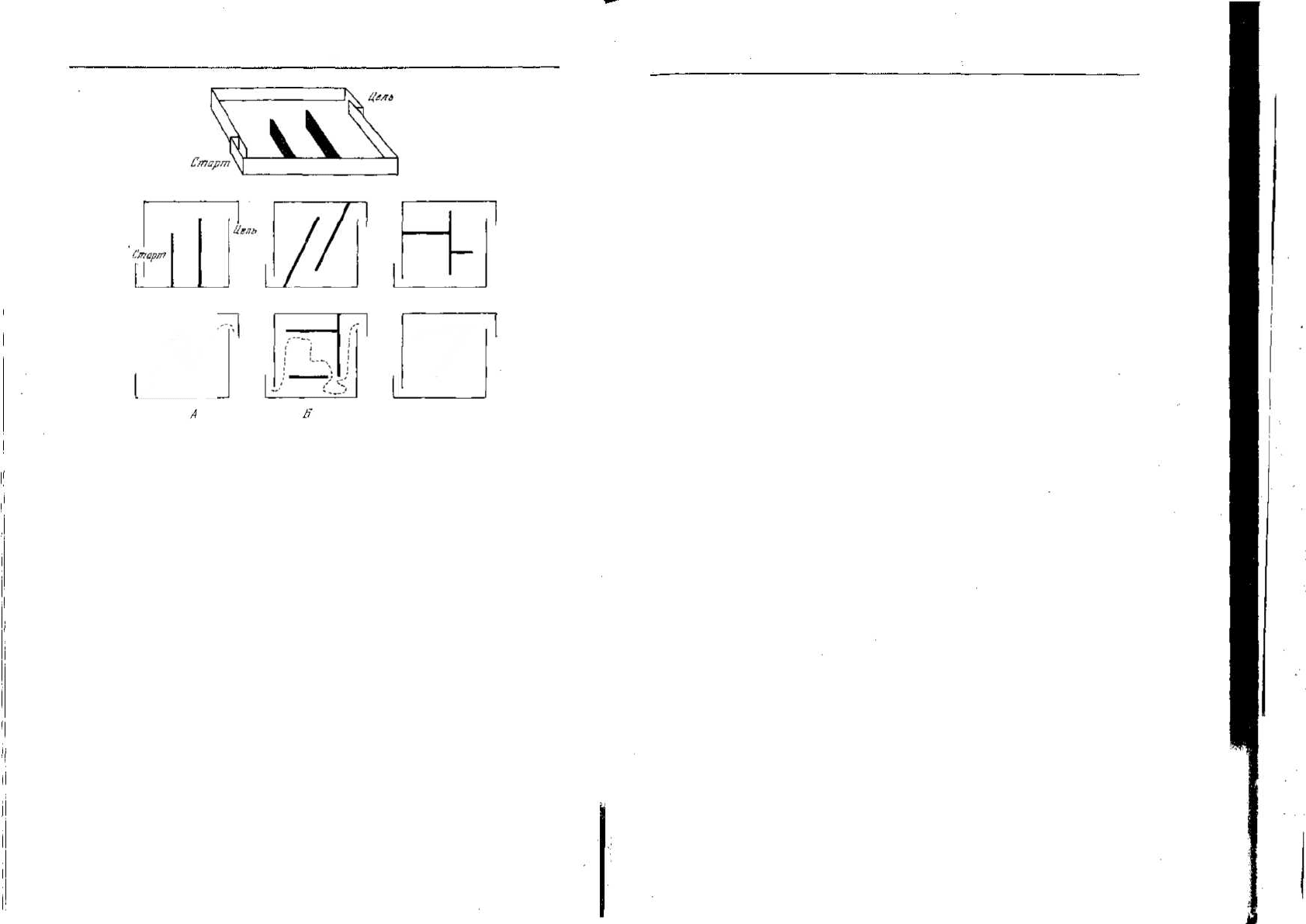
Рис. 7.7. Один из типов ситуации, предложенной для оценки «интеллекта» живот-
ных [207]. Простая огороженная площадка, показанная вверху, имеет передвиж-
ные барьеры. Расположенные ниже схемы показывают серию различных задач на
обход преграды, которые по очереди предъявляют животным. Путь обхода в случае
А означает высокую оценку, а в случае Б — низкую. Оценки за решение многих
задач усредняются для выведения показателя «интеллекта».
роятно, на ориентации относительно Солнца, поэтому вряд ли можно
назвать данное поведение «инсайтом». Это соображение заставляет
нас пересмотреть вопрос о том, могут ли задачи поиска обходных
путей у высших животных служить показателем инсайта.
Классический пример инсайта у животных дают ранние опыты
Кёллера [271] на шимпанзе. Когда гроздь бананов подвешивали
слишком высоко и обезьяны были не в состоянии до нее дотянуться,
они начинали ставить друг на друга ящики или вставляли друг в
друга палки, так чтобы влезть повыше или сбить бананы на землю.
Зачастую они достигали такого решения совершенно внезапно, хотя
и пользовались предшествующим опытом игры с ящиками и пал-
ками (латентное обучение), причем для построения устойчивой
пирамиды из ящиков обезьянам требовался значительный период
проб и ошибок.
Шимпанзе Кёллера использовали знания, полученные в одной
ситуации (некоторые свойства палок и ящиков), и применяли их в
другой ситуации. Не вызывает сомнений тот факт, что у человеко-
образных обезьян и других приматов иногда возможны настоящие
проявления рассудка. Многие владельцы собак приводят примеры,
когда их питомцы совершают разумные действия; не исключено,
что это действительно так, однако нужно постараться исключить
возможность и других объяснений. Торп [457] приводит ряд других
примеров, касающихся поведения птиц и млекопитающих, которое
свидетельствует о наличии мышления у животных (см. гл. 6 и 7 его
книги).
СРАВНИТЕЛЬНОЕ ИССЛЕДОВАНИЕ ОБУЧЕНИЯ
Сравнительная психология имеет большую историю. Ее часто
критиковали, и вполне обоснованно, за то, что она отошла от задач
сравнительного описания и сконцентрировала свои усилия почти
целиком на двух объектах — голубе и белой крысе. Пример такой
критики можно найти в работе Бича [42]. В настоящее время психо-
логи исследуют гораздо более широкий круг животных, и можно
надеяться, что сравнительное исследование способности к обучению
даст информацию как об эволюции обучаемости, так и о степени
общности любых «законов обучения», которые будут сформулиро-
ваны.
Один из подходов, применяемых сравнительными психологами,—
выяснение корреляции между развитием мозга и способностью к
обучению. Детье и Стеллар [133] дали в своей книге замечатель-
ный обзор структуры нервной системы у животных разных система-
тических групп, рассмотрев вопрос о том, насколько сложность по-
ведения может быть связана с развитием мозга. Способность к обу-
чению усиливается в ряду позвоночных, и мы можем грубо связать
это с увеличением конечного мозга. Важен, видимо, не сам по себе
размер мозга; киты и слоны имеют больший мозг, чем человек, но
обладают меньшей, хотя и значительной, способностью к обучению.
Гораздо труднее выявить какую-то закономерность в отношении бес-
позвоночных, которые гораздо более гетерогенны, чем позвоночные.
Известно, что высшие насекомые и головоногие имеют самый боль-
шой мозг среди представителей членистоногих и моллюсков соот-
ветственно и отличаются самой высокой способностью к обучению.
Наиболее важный аспект эволюции мозга позвоночных — рост
мозговых полушарий, особенно их коры, который достигает апогея
у приматов. Однако мы должны избегать простого уравнивания раз-
вития коры и способности к обучению. Например, способности птиц
в прошлом часто недооценивались, потому что в мозгу птиц, отно-
сительно даже большом по размеру, отделы, гомологичные коре мле-
копитающих, развиты слабо. Однако птицы эволюционировали не-
зависимо от млекопитающих в течение 200 млн. лет. В процессе
эволюции у них сформировался мозг другого типа, а по способности
к обучению они в некоторых отношениях уступают лишь приматам.
Противоположный пример уже приводился в гл. 2: хотя среди на-
секомых двукрылые и перепончатокрылые имеют мозг сходного
строения, они чрезвычайно различаются по способности к обучению.

280
Г лава 7
Очевидно, одна только структура мозга не может служить пока-
зателем способности к обучению, и, чтобы изучить эволюцию обу-
чения, нам необходимо сравнить, как различные животные справля-
ются со специфическими поведенческими тестами. При этом мы сразу
сталкиваемся с множеством проблем. Одна из них — выбор пред-
ставителей различных групп животных, которые позволяли бы де-
лать заключения об эволюции способности к обучению. Слишком
просто относить животных к «высшим» или «низшим». Мы, например,
часто видим, что исследователи считают последовательность рыбы,
амфибии, рептилии, птицы, млекопитающие эволюционным рядом
увеличения сложности поведения позвоночных и повышения их
способности к обучению.
При составлении такого ряда из ныне живущих представителей
разных классов не принимается во внимание действительный ход
эволюционного процесса. Мы уже указывали, что млекопитающие и
птицы имеют совершенно различные истории развития. Все ныне
живущие млекопитающие одинаково удалены во времени от общего
предка и все они приспособлены к особому образу жизни. Было бы
наивно ожидать, что способность к обучению у современных костис-
тых рыб (например, у серебряного карася, обычно используемого
при исследовании обучения) точно отражает эту способность у древ-
них рыб, от которых современные костистые рыбы и другие позво-
ночные дивергировали 400 млн. лет назад. Вот почему построение
обоснованной филогении поведения связано с большими трудностя-
ми, ведь поведение не оставляет после себя ископаемых остатков.
Ходос и Кзмпбелл [227] дают убедительный критический разбор
филогенетических построений, сделанных некоторыми специалис-
тами по сравнительной психологии.
Другая проблема, возникающая при сравнительных исследова-
ниях способности к обучению,— создание действительно сравнимых
задач для тестирования различных животных, столь значительно
варьирующих по сенсорному оснащению и способности к манипули-
рованию. Чтобы сравнить способность к различению у осьминога,
пчелы и крысы, нужно использовать совершенно различные приемы
обучения, и в настоящее время нельзя быть уверенным в том, что
предлагаемые этим животным задачи имеют для них одинаковую
трудность и воспринимаются ими одинаково. Уравнивание моти-
вации и подкрепления представляет собой с-ледующую трудность.
Мы уже видели (с. 273), что уровень мотивации влияет на скорость
обучения и иногда определяет даже саму возможность обучения.
Каким же путем можно выровнять уровень мотивации голода у кры-
сы и рыбы? Последняя может жить без пищи неделями, тогда как
первая — лишь несколько дней. Столь же трудно уравнять и под-
крепление для различных животных. Маленький кусочек пищи мо-
жет служить сильным подкреплением для голодного млекопитаю-
щего, но имеет гораздо меньшее значение для рыбы и еще меньшее
Обучение
281
для аннелиды. Легче, пожалуй, уравнять условия наказания, так
как все животные отрицательно относятся к электрическому разд-
ражению. Однако и здесь имеются трудности, потому что электри-
ческое раздражение и вызываемый им страх по-разному влияют на
поведение. Как уже рассматривалось выше (с. 269), животным, ис-
пользуемым в экспериментах по обучению, свойственны различные
внутренние наклонности. Чтобы уравнять влияние наказания, не-
обходимо знать особенности реакций животного на вызывающие
страх раздражители в естественных условиях. Например, аппарат
для выработки условного рефлекса избегания (с. 272) основан на
том, что раздражаемое током животное должно убегать с решетки,
через которую наносится раздражение, постепенно запоминая, что
звонок или свет являются сигналом скорого раздражения. Однако
некоторые грызуны при испуге не бросаются бежать, а наоборот,
застывают и остаются неподвижными, даже если им наносится
электрическое раздражение. У таких животных выработка реакции
избегания будет происходить гораздо медленнее, чем у животных,
которые при появлении опасности спасаются бегством. С точки зре-
ния экспериментатора, активные животные обычно гораздо легче
поддаются обучению любых типов (хороший обзор этих вопросов
имеется в работе Дэни и Ратнера [129]).
Несмотря на все трудности, применив известную изобретатель-
ность и большие предосторожности, можно придумать тест, который
давал бы различным животным достаточный шанс продемонстри-
ровать свои способности. Например, Биттерман [51] описывает
камеру Скиннера для рыб и черепах, обеспечивающую им пищевое
подкрепление. Шнейрла [322] и Воулс [479] показали, что лучшим
подкреплением при обучении муравьев в лабиринте является воз-
вращение в гнездо. Для саламандр [476] и некоторых аннелид [98]
наилучшим подкреплением оказалось на некоторое время оставить
животное в покое!
Несмотря на трудности, возникающие при изучении эволюции
поведения, данная проблема не перестает привлекать внимание ис-
следователей. В связи с этим возникает вопрос о пересмотре нашей
классификации видов обучения, поскольку необходимо выяснить,
имеют ли различные ее категории какую-нибудь филогенетическую
обоснованность, т. е. можно ли их распределить по порядку появ-
ления у представителей разных ступеней эволюционной лестницы,
по крайней мере в той степени, в какой эта лестница включает ныне
живущие организмы.
Несомненно, наиболее древний вид обучения — привыкание,
и оно, как мы уже указывали, встречается у всех животных. Легко
понять, какие преимущества дает животному способность к привы-
канию; она предохраняет животное от траты внимания на несущест-
венные для него раздражители, что способствует сужению набора
реакций животного.
260
Глава 7
Обучение
283
Какая-то форма ассоциативного обучения должна быть сле-
дующим шагом эволюции обучения, но оказалось, что это доволь-
но большой шаг. Ассоциативное обучение имеет некоторые черты
сходства с привыканием — для того чтобы обучение происходило,
животное должно уметь отличать знакомую ситуацию от незнакомой.
Другие промежуточные этапы в эволюции обучения обнаружить
трудно. Однако не исключено, что явление, обычно называемое «сен-
ситизация», представляет собой стадию в эволюции ассоциативного
обучения, причем в некотором смысле стадию, противоположную
привыканию. Мы говорим о сенситизации, когда после подкрепле-
ния или наказания возрастает реактивность животного на разно-
образные раздражители. Например, Эванс [148] показал, что червь
Nereis (с. 259) иногда высовывается из своей трубки при вспышках
света. В одном опыте такая реакция наблюдалась у 21% червей.
Однако после одного пищевого подкрепления, данного в отсутствие
света, более 60% червей высунулись из трубок в ответ на вспышку
света, произведенную через 30 мин после подкрепления. Подобно
этому, только что накормленный осьминог гораздо охотнее ата-
кует картонные фигуры, опущенные в его аквариум, чем это делает
• голодное животное. Еще более эффективно способствует сенситиза-
ции наказание. После электрического раздражения лап и тараканы,
и крысы спасаются бегством при действии разнообразных новых
раздражителей, которые никак не были связаны с прежним и
которые не вызывали реакции избегания у животных, не подвергав-
шихся сенситизирующему воздействию.
Для того чтобы происходило настоящее ассоциативное обучение,
животные должны начать различать раздражители, связанные с
подкреплением и не связанные с ним. Повышение реактивности во
время подкрепления способствует процессу ассоциативного обуче-
ния. Среди беспозвоночных настоящее ассоциативное обучение обна-
ружено у членистоногих и головоногих моллюсков (осьминога и
родственных ему видов). Данные о наличии ассоциативного обуче-
ния у аннелид и плоских червей почти во всех случаях допускают
и другую трактовку. Часто, например, имеет место не ассоциативное
обучение, а сенситизация (более подробное обсуждение см. в [148]
и [458]).'
Можно было надеяться, что для позвоночных удастся подобрать
хорошую серию тестов на способность к обучению, которая позво-
лила бы выявить разные стадии эволюции
Л
интеллекта». Казалось,
что скорость обучения — качество, восхищающее нас в школьни-
ках,— может служить полезным видом оценки. Однако, как следует
из краткого обзора литературы, одна только скорость обучения мало
о чем говорит. Муравьи и крысы, например, сравнимы по скорости
обучения в довольно сложном лабиринте. Геллерман [167] детально
описывает эксперименты, в которых двух шимпанзе и двух детей в
возрасте двух лет обучали различению белого треугольника на фоне
•
A U В
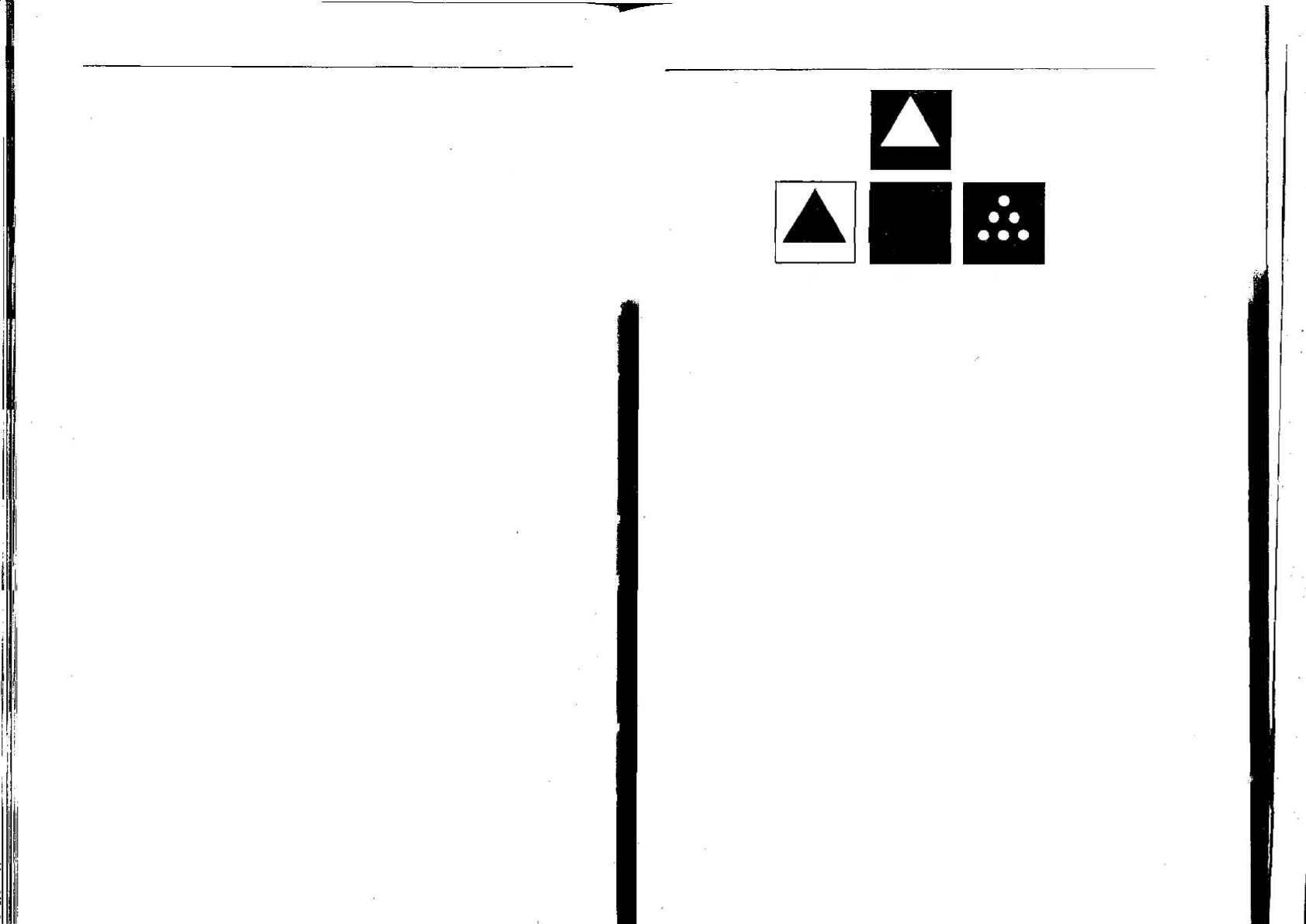
Рис. 7.8. Понятие «треугольности» [206]. Крысы, обученные отвечать на верхнюю
фигуру, реагируют случайным образом на любую из фигур нижнего ряда. Шим-
панзе реагируют на Л и Б, но В выбирают случайно. Двухлетний ребенок узнает
треугольник на всех трех фигурах.
черного квадрата, который был связан с пищей, от простого черного
квадрата. Один ребенок усвоил эту дифференцировку с одной пробы,
тогда как другому потребовалось 200 сочетаний, а обоим шимпанзе
свыше 800 сочетаний для достижения критерия 19 правильных вы-
боров из 20. В сходном опыте, поставленном на крысах, большинству
из них требовалось от 20 до 60 проб, хотя их обычно слегка наказы-
вали за неправильные ответы и вознаграждали за правильные. Рас-
хождения такого рода многочисленны как в пределах одного вида,
так и между разными видами, и у нас нет веских доказательств, что
скорость начального обучения в простых ассоциативных задачах
отличается не только у разных позвоночных, но и у позвоночных
и высших беспозвоночных.
Ясно, однако, что скорость — лишь один из аспектов обучения;
немаловажным также является вопрос, что выучивается. Напри-
мер, как поясняет рис. 7.8, хотя шимпанзе тратят больше времени
на выработку упомянутого выше дифференцирования треугольни-
ков, они при этом больше узнают о признаке «треугольное™», чем
крысы. Один из показателей «ума» — способность установить в тес-
тах подобного типа правильный баланс между генерализацией и
дифференцированием. Если мы рассмотрим более сложные формы
обучения, то сможем по крайней мере выявить некоторые градации
в способности к обучению среди позвоночных.
В этом отношении полезно исследовать формирование установки
обучения. Согласно одной из точек зрения, если животное способно
сформировать установку обучения, то это означает, что оно может
не только обучиться решать какую-то задачу, но усвоить принцип,
лежащий в ее основе, и сможет устойчиво увеличивать скорость
обучения при предъявлении серии сходных задач. Харлоу [201]
описал основную методику, применяемую для приматов.

260
Глава 7
Обучение
284
Обезьяне показывают пару разных предметов, например спичеч-
ную коробку и подставку для яиц. Под спичечной коробкой незави-
симо от места, где она находится, обезьяна всегда находит подкреп-
ление, а подставка для яиц с подкреплением никак не связана.
После ряда проб обезьяна сразу начинает выбирать спичечную ко-
робку. Тогда ей предлагают новую пару предметов, причем кубик из
детского строительного набора подкрепляют, а половинку теннис-
ного мяча — нет, Чтобы усвоить, какой предмет нужно выбирать,
обезьяне требуется почти столько же времени, как и в первый раз.
Затем снова предъявляют новые предметы и т. д. После нескольких
десятков таких задач обезьяны решают каждую новую задачу
гораздо быстрее, хотя, взятая отдельно, она не менее трудна,
чем предъявлявшиеся вначале. Приблизительно после 100 таких
задач при предъявлении пары новых предметов обезьяна выбирает
один, и, если он подкрепляется, в каждой последующей пробе она
выбирает только его. Если же выбранный обезьяной при первом
предъявлении предмет не получил подкрепления, то во всех после-
дующих пробах она выбирает только второй, подкрепляемый пред-
мет. Таким образом, обезьяна улавливает принцип задачи или, по
терминологии Харлоу, формирует «установку обучения».
Формирование установки обучения — это лишь один вариант
методики, основанной на применении последовательных попыток
дифференцирования. Вероятно, более простой ее вариант — «по-
следовательные переделки», В этом случае животных сначала учат
выбирать предмет А. Когда животное научится это делать, выбор
предмета А перестают сопровождать подкреплением и начинают
подкреплять выбор предмета Б, который раньше не подкрепляли.
Когда эта первая переделка дифференцирования будет выработана,
снова начинают подкреплять выбор предмета Л и т. д. Если при каж-
дой последующей переделке животное будет обучаться быстрее,
чем при предыдущей, то это свидетельствует о том, что оно усвоило
принцип задачи.
Способность к формированию установки обучения раньше рас-
сматривалась как свойство только высших млекопитающих, но те-
перь мы знаем, что на самом деле это не так. Уоррен [484] обобщил
сравнительные данные по рыбам, черепахам, птицам, крысам, кош-
кам и приматам. Хотя дальнейшие исследования могут показать
другое, по этому критерию рыбы, у которых установка обучения не
формируется, вероятно, отличаются от всех остальных позвоночных.
Недавно Мэкинтош [319] представил убедительные данные, что ось-
миноги способны к успешному обучению последовательным пере-
делкам, так что в этом отношении высшие беспозвоночные превос-
ходят низших позвоночных. На насекомых подобные исследования
еще не проводились.
Хотя скорость образования простой дифференцировки мало от-
личается у животных разных систематических групп, скорость фор-
4
т
40
I
г
I во
1
1
4
70
50
40
,» Обезьяны - резусы
Беличьи абезьюш
\Мгрутоаые обезьяны
> Квшки Крысы
Слрчайныи уровень
I I J
I . I I I I J
200
BOO
1000
Задачи
Moo
_I L
1800
Рис. 7.9. Скорость формирования установки обучения у разных животных [484].
В каждой новой задаче первый выбор животного должен быть случайным, но если
оно усвоило принцип задачи, то уже второй выбор будет верным. Обратите внима-
ние, как много времени требуется, чтобы у крыс и белок правильные выборы при
втором предъявлении задачи превышали случайный уровень. Многие обезьяны до-
стигают почти 100%-ного уровня в пределах 400 задач.
мирования установки обучения у них меняется очень существенно.
Рис. 7.9 показывает, насколько быстрее формируется установка
обучения у обезьян, чем у крыс, у которых постепенное улучшение
начинается лишь после 800 последовательно предъявленных задач.
При формировании установки обучения и в более сложных зада-
чах превосходство приматов несомненно, однако это, по-видимому,
количественное превосходство. У нас нет доказательств, что они
обладают какими-то способностями, которые не имели бы предшест-
венников у более низкоорганизованных млекопитающих.
Трудно сдержать чувство смутной неудовлетворенности от такого
заключения. Видимо, слишком ограниченная искусственность мно-
гих экспериментов по обучению не позволяет отдать должное не-
правдоподобной приспособляемости и изобретательности высших
приматов, таких, как шимпанзе, В данном случае мы несомненно
находимся под влиянием их сходства с нами, особенно в том отно-
шении, что они, как и мы, способны манипулировать предметами.
Мы можем недооценивать ум других животных, потому что у них
нет хороших рук и хорошего зрения. Мы осознали, насколько за-
мечателен ум дельфинов лишь недавно, потому что их строение и
среда обитания столь отличны от нашей.
Как упоминалось в предыдущей главе, исследования поведения
шимпанзе в естественных условиях показали, что они могут выби-
рать и обрабатывать простые орудия. Гудолл [169] описывает, что
шимпанзе жевали листья и использовали их как губку, чтобы доста-
I во
J
I

i!
286
Глава 7
вать воду из углублений. Кроме того, они подбирали веточки и
очищали их от боковых побегов, пока не доводили до размера,
подходящего для доставания термитов. Такие наблюдения нас не
удивляют, потому что собственные предки человека — австралопи-
теки — использовали кости как оружие против других животных
и обрабатывали кремниевые орудия. Размер их мозга составлял от
650 до 680 см
3
, немногим больше мозга современной гориллы [99].
От австралопитека до человека с объемом мозга около 1500 см
3
произошел огромный скачок, и некоторые авторы утверждают, что
человек располагает способностями, которые даже в слабой степени
не обнаруживаются в поведении человекообразных обезьян. Однако
предок человека находился на стадии человекообразной обезьяны
сравнительно недавно, и существовали хорошо приспособленные
промежуточные формы, подобно Homo erccius, которые имели мозг
объемом 1000—1200 см
3
. Харлоу [202] указывает, что использова-
ние человеком символического языка дает ему огромное преиму-
щество перед обезьянами, которые языка не имеют, однако этот
пробел представляет собой лишь один шаг на пути к человеку.
Возможно, что современным человекообразным обезьянам не хва-
тает лишь немногого до уровня, необходимого для появления языка.
Келлог [264] описывает несколько безуспешных попыток научить
шимпанзе говорить. Никому не удавалось научить их произносить
более одного-двух членораздельных слов. Однако это может быть
обусловлено просто их неспособностью к артикуляции (у человека
гортань и буккальная полость гораздо более специализированы для
речи 12901) и тем, что общение с помощью жестов гораздо более ес-
тественно для них. Действительно, недавно было проведено два за-
мечательных эксперимента по обучению шимпанзе, которые пока-
зали, что шимпанзе способны использовать символический язык с
удив11тельной легкостью. Гарднеры [164, 165] использовали язык
жестов, принятый-у глухих в Америке. Примак [3 8# использовал
для обозначения отдельных предметов и слов цветные кусочки плас-
тика разнообразной формы. В обоих случаях в опытах использо-
вали по одной самке шимпанзе, причем их учили не просто узнавать
предметы и связывать их с определенным жестом или символом, но
заставляли использовать эти жесты и символы, чтобы просить
обозначаемую ими вещь и общаться с экспериментатором. Обе обезь-
яны овладели словарем в несколько дюжин слов. Не возникает сом-
нений в замечательной способности шимпанзе усваивать названия
предметов и некоторых действий, таких, как «входи, Джим», «играть
в прятки», и изменять фразы при помощи, например, слова «больше».
Спор об истинной природе этих возможностей обезьян продолжается.
Обладают ли их предложения какой-то простой грамматической
конструкцией? Гарднеры утверждают, что они могут сопоставить
их способности и способности детей первых лет обучения речи [163,
165].
Обучение
287
Эти важные эксперименты открывают увлекательные перспек-
тивы на будущее, правда, они требуют невероятного терпения и
времени. В лаборатории Гарднеров шимпанзе общалась с людьми
весь период бодрствования и не использовала других средств ком-
муникации, кроме знакового языка. Сейчас этому языку обучают
небольшие группы молодых шимпанзе. Что они будут «говорить»
друг другу, когда овладеют таким средством коммуникации? Как
бы то ни было, уже полученные результаты заставили нас полностью
пересмотреть наши представления о том, чего могут достигнуть
600 см
3
мозга.

289
ОРГАНИЗАЦИЯ СООБЩЕСТВ
До сих пор мы обсуждали вопросы, освещающие организацию
поведения отдельного животного. Однако часто мы сталкивались
с необходимостью рассмотреть аспекты, связанные с взаимодейст-
вием особей, например агрессивное поведение или ухаживание. Фак-
тически все животные хотя бы часть времени живут парами или бо-
лее крупными группами. У некоторых видов социальность — это
доминирующая сторона их жизни, и теперь мЫ остановимся на жиз-
ни животных в сообществах как таковой.
Сплоченность и организованность сообществ животных часто
бывает поразительной. В таких группах отдельную особь легко по-
терять из виду и можно не заметить, что ее поведение формируют те
же силы, которые формируют поведение особей, принадлежащих к
видам, живущим поодиночке. Естественный отбор действует на
отдельные особи, и реакции общественного животного на других
членов группы эволюционируют для его наибольшего блага.
Понятие «организация сообщества» относится к популяциям, а не
к особям и описывает характер взаимодействий между членами
вида. В некоторых случаях, например у различных общественных
насекомых, структура сообщества бывает достаточно жесткой и ви-
дбспецифичной. Как мы увидим дальше, у позвоночных организа-
ция сообщества значительно более динамична и может меняться в
зависимости от обстоятельств. Разумеется, это понятие относится не
только к высокосоциальным видам. И тигры, которые обычно живут
и охотятся в одиночку на большой территории и (за исключением пе-
риода размножения) избегают контакта друг с другом, и медоносные
пчелы, которые проводят всю свою жизнь в густонаселенной коло-
нии, дают нам примеры организации сообществ, хотя и совсем раз-
ных типов.
Любое взаимодействие между двумя особями одного вида можно
совершенно обоснованно называть «общественным поведением».
Этого критерия придерживаются в своих книгах Тинберген [463]
и Димонд [137], однако здесь, рассматривая общественное поведе-
ние разных типов, мы сосредоточим внимание на так называемых
сообществах животных. Истинное сообщество — это нечто большее,
чем супружеская пара или мать с детенышами. Оно представляет
собой стабильную группу, члены которой поддерживают интенсив-
Организация сообществ
291
н.ую коммуникацию и находятся в некоторых относительно постоян-
ных отношениях друг с другом.
В животном царстве существуют самые разнообразные объедине-
ния и группы, однако далеко не все из них отвечают критериям,
приведенным выше. К двум в некотором смысле противоположным
примерам относятся муравьи и слоны. Образующие колонию мура-
вьи живут в общем гнезде, которое они совместно строят. Царица
(матка) и ее стерильные дочери, которых часто подразделяют на
две группы — рабочих и солдат,— имеют характерное, свойствен-
ное им строение.
Сообщество африканских слонов организовано значительно ме-
нее жестко. Стадо из 40—50 голов обычно возглавляет старая сам-
ка — матриарх. Ее потомство двух или даже трех поколений нахо-
дится вблизи от нее — эта маленькая группа составляет ее семью.
С семьей матриарха объединяются другие семьи, образуя то, что
Дуглас-Гамильтон [138] называет «семейной единицей». Молодые
самцы сначала остаются со своими семьями, однако после полового
созревания живут поодиночке, но обычно близко от других самцов,
иногда объединяясь с ними. Они вступают в краткий контакт с
«семейной единицей», а затем спариваются с самками в эструсе.
В таких семейных группах слоны живут по 40—50 лет.
В сравнении с этими истинными сообществами организация
стаи птиц или рыб представляется значительно более простой, хотя
особи могут оставаться вместе месяцами. Совершенно очевидно, что
применять понятие «сообщество» к стае дафний, собравшихся в бо-
гатом пищей участке, или группе дрозофил на гниющем фрукте не-
верно. Дрозофилы и дафнии образуют скопления, поскольку их
всех привлекает источник пищи. Они реагируют на присутствие
друг друга, например распределяясь в пространстве таким обра-
зом, чтобы не касаться друг друга, но не образуют сообщества.
ПРЕИМУЩЕСТВА ЖИЗНИ В ГРУППЕ
Мы высказали выше предположение, что естественный отбор
формирует реакции особи внутри группы точно так же, как он влия-
ет на любые другие приспособления животного к среде. Следова-
тельно, можно ожидать, что жизнь в любой группе, будь это скоп-
ление или истинное сообщество, оказывается благоприятной для
составляющих ее членов.
Работы Элли и его школы [6] сыграли важную роль, показав,
каким образом скопления дают преимущества особям, их составля-
ющим. Иногда благоприятное влияние почти целиком определяется
физическими размерами группы. Дафнии не способны жить в ще-
лочной воде, а продукты дыхания множества особей создают иног-
да достаточно кислую среду, так что содержание щелочи в ней сни-
жается до приемлемого уровня. Таким образом, там, где отдельная
10 № 147
Г пава

290
Г лава
Организация сообществ
291
особь гибнет, группа выживает. Культуры дрозофилы плохо раз-
виваются, если в них слишком много яиц, так как вылупляющимся
личинкам при этом не хватает корма, в то же время слишком малое
число яиц почти так же плохо, поскольку малое число личинок не в
состоянии хорошо взрыхлить среду, чтобы сделать ее годной в пищу.
Для самки оказывается благоприятным откладывать свои яйца рядом
с чужими, поскольку потомство при этом получает преимущество.
Стаи птиц и рыб представляют собой примеры групп, которые
обладают лишь некоторыми признаками сообщества, но часто ока-
зываются более сложными, чем простые скопления. В объединении
их в группы определенную роль играют физические факторы, как,
например, в случае с императорскими пингвинами, которые при
инкубации яиц во время антарктической зимы стоят, прижимаясь
друг к другу. Это способствует удержанию тепла, так как птицы,
находящиеся снаружи, двигаются больше, чем те, что стоят в центре
группы: в результате происходит постепенное перемещение пинг-
винов, и поэтому все они имеют относительна одинаковое укрытие.
Однако между особями иногда происходят взаимодействия, кото-
рые никак нельзя отнести к просто физическим.
Одно из наиболее очевидных преимуществ спаянной группы,
члены которой реагируют на поведение друг друга, заключается в
защите от хищников. При наличии нескольких особей, стоящих на
страже, приближение хищника вряд ли останется незамеченным,
а как только он обнаружен, сигнал тревоги слышат все сразу.
В табл. 8.1 приводятся реальные цифры, свидетельствующие о свя-
зи уровня бдительности скворцов с размерами группы. Пауэлл
[382] наблюдал за скворцами, кормящимися на площадке, где он
мог показывать им чучело сокола. Он определял частоту, с кото-
рой птицы, кормящиеся в одиночку или группами, прерывали кор-
Л
жку, чтобы посмотреть вверх, а также латентный период их взле-
та после появления сокола. Стаи снимались с места одновременно,
а птица, которая первой замечала хищника, взлетала первой и
Таблица 8.1
Кормление, наблюдение и время реакции у скворцов, сидящих поодиночке
и в группе (подробности в тексте (382])
Одиночные Группа из Группа из
ПТИЦЫ
5 птиц 10 птиц
Процент времени, проведенного за
53
70 88
кормежкой
Процент времени, проведенного за на-
47
30 12
блюдением
Время реакции на модель летящего 4,1 с
Не измеря- 3,2 с
сокола
лось
А*
4
Рис. 8.1. Реакция стаи скворцов на приближение хищной птицы [460].
обычно при этом издавала крик тревоги. Отметим, что птицы в стае
заметно реже взглядывали вверх и взлетали быстрее, чем одиноч-
ные. Хорошо известно, что даже полсекунды промедления при стре-
мительном броске хищника может означать смерть.
С уменьшением необходимости проявлять бдительность у птиц
должно освобождаться больше времени для кормежки и другой ак-
тивности. Даже если хищник обратил группу птиц в бегство, все
равно объединение в группу представляет дополнительные преиму-
щества, поскольку они могут предпринять согласованные защитные
действия. На рис. 8.1 показан такой маневр стаи скворцов. Точно
так же ведут себя некоторые рыбы, которые при малейшей тревоге
собираются вместе. Хищники редко нападают на отдельных осо-

№
293 Г лава
Организация сообществ 311
310
l
!
I
11
I !
II i i
I
Рис. 8.2. «Круговая оборона» группы мускусных быков в канадской тундре. При
приближении хищника они собираются вместе, причем снаружи группы стоят
старые животные, принимающие на себя атаку. (Фото Д. Уилкинсона, Information
4
Canada Phototheque.)
бей в плотной стае; их наиболее распространенная стратегия — это
наброситься на группу и вызвать ее рассеивание, в результате чего
они имеют возможность нападать на отделившееся от группы изо-
лированное животное.
Колониально гнездящиеся птицы — чайки и крачки — оказы-
вают поразительное сопротивление вторгающемуся в их колонию
хищнику, например лисе: Они совершают на нее совместные атаки,
хотя каждая птица реагирует более или менее индивидуально, за-
щищая свою собственную территорию. В более высокоорганизован-
ных сообществах способы защиты от хищников бывают еще более
эффективными. Мы уже говорили, что жизнь в группах обеспечи-
вает павианам защиту от леопардов — когда объединяются нес-
колько взрослых самцов, ни один леопард не может им противо-
стоять/На рис. 8. 2 показана круговая оборона стада мускусных
быков, которую они осуществляют при приближении волков; ха-
рактерно, что при этом старые животные прикрывают молодых.
Защита от хищников — это только одно из преимуществ, ко-
торыми обладает группа. Колония луговых собачек, постоянно об-
грызая высокие травинки, поддерживает открытой большую об-
ласть, давая возможность расти мелким травянистым растениям»
которыми они питаются. Кормежка стаями облегчает птицам поиск
пищи, потому что группа сильнее, чем отдельная особь, нарушает
целостность местообитания и вспугивает добычу, а также потому»
что вид кормящейся птицы привлекает к этому месту других. Кребс
и др. [277] показали, что, когда в стае больших синиц одна птица
обнаруживает пищу, остальные быстро меняют стратегию поиска й
концентрируют внимание на том участке и на тех местах деревьев,
где была найдена пища. Гнездование колониями иногда также обес-
печивает преимущества в добывании пищи, особенно если в какой-
то области источники пищи распределены неравномерно. Кребс [276]
изучал колонии большой голубой цапли и обнаружил, что птицы,
безуспешно летавшие за пищей, нередко парят над колонией и ле-
тят вслед за теми особями, которые, прилетев с кормежки, быстро
улетают снова. К последним относятся птицы, которым посчастли-
вилось найти косяк рыбы. Колониальное гнездование краснокрылого
трупиала, а также огромные скопления, которые образуют вороны,
скворцы и другие птицы в течение зимних месяцев, видимо, имеют
преимущества, подобные описанным выше. Группы, узнающие о
хороших местах кормежки, раньше улетают из стаи и, возвращаясь,
улетают снова, сопровождаемые другими особями. В течение всей
зимы каждая особь, по-видимому, выигрывает от этого, так как бла-
годаря многочисленности птицы имеют возможность использовать
большее число пригодных для кормежки мест [517].
В некоторых случаях хищники также получают преимущества
от жизни в группе. У львов, гиен и гиеновых собак во время охоты
члены группы координируют свои действия. Их стратегия зачастую
заключается в том, что одни особи гонят добычу к месту, где в за-
саде находятся другие. Известно также, что собаки по очереди пре-
следуют антилопу до ее полного изнеможения. Превосходный обзор
различных способов охоты у хищных опубликован Крууком [280].
Еще один фактор, который благоприятствует особям, существу-
ющим в группе,— это стимуляция синхронизации размножения.
В гл. 4 мы говорили об опытах, показавших, что гормональные цик-
лы самок некоторых птиц и млекопитающих изменяются в присут-
ствии самцов. Большое число морских птиц гнездится в плотных
колониях, хотя в пределах колонии отдельные особи яростно защи-
щают свою территорию. Дарлинг [119] одним из первых отметил,
что в таких колониях птицы получают много зрительных и слуховых
раздражителей от демонстраций, которые постоянно происходят в
начале сезона размножения. Под влиянием такой стимуляции, ис-
10* № 147

294
Глава 8
i ,
"i!
ходящей как от соседей, так и от партнера по паре, ускоряются и
синхронизируются циклы размножения птиц в пределах колонии.
Дарлинг предполагает, что это воздействие благоприятно, посколь-
ку оно позволяет сократить до минимума длительность периода раз-
множения, когда птицы столь уязвимы. Если откладка яиц и вылуп-
ление происходят синхронно, то хищники, которые способны съе-
дать ежедневно только ограниченное количество яиц и птенцов, на
короткое время оказываются как бы «завалены» ими, так что для
каждого отдельного гнезда возможность нападения уменьшается.
Экспериментальное подтверждение так называемого эффекта
Дарлинга получить нелегко, однако кое-что в этом отношении дают
замечательные длительные наблюдения за моевками, проведенные
Коулсоном [105]. Он показал, что более успешно размножаются
птицы, гнездящиеся ближе к центру колонии, а не к ее краям. Они
откладывают больше яиц и в более ранние сроки; не приходится
сомневаться в том, что одним из факторов, обеспечивающих этот
эффект, оказывается социальная стимуляция, имеющая место в
центральной части колонии. Центр обладает большей привлека-
тельностью, ив начале сезона моевки яростно сражаются за места в
центральной части колонии.
Наконец, при обсуждении преимуществ жизни в группе мы долж-
ны кратко рассмотреть взгляды Уинн-Эдвардса, изложенные им в
его знаменитой книге «Дисперсия животных в связи с их обществен-
ным поведением» [514]. Уинн-Эдвардс придерживается точки зре-
ния, прямо противоположной той, которую мы до сих пор развива-
ли в этой главе. Мы рассматривали, какие преимущества приносит
жизнь в обществе особям, его составляющим. Уинн-Эдвардс пред-
полагает, что этим вопрос далеко не исчерпывается и что в эволюции
общественной жизни животных 'часто особь жертвует собственными
«эгоистическими» интересами для благополучия группы в целом.
Он предполагает, что только таким способом животные могут су-
ществовать в пределах ресурсов, обеспечиваемых внешней средой.
Если бы животные всегда размножались с максимальной скоростью,
то они очень скоро съели бы всю пищу, а, как утверждает Уинн-
Эдвардс, в природных условиях этого почти никогда не случается.
Эволюционировали те группы, у которых скорость размножения
имеет связь с наличными ресурсами. Животные поддерживают ско-
рость размножения на более низком уровне по сравнению с макси-
мальным, и в устройстве их сообществ имеются важные регулятор-
ные механизмы. Поскольку группы стабильны и в большой степени
репродуктивно изолированы от других, отбор действовал на сами
группы, устраняя те, которые не смогли адаптироваться, т. е. име-
ли размер, не соответствующий объему ресурсов.
С представлением о групповом отборе связаны большие трудно-
сти. Предположим, как это предлагает Уинн-Эдвардс, что колония
птиц существует в условиях обеспечения ее пищевыми ресурсами,
Организация сообществ
295
откладывая меньшее, чем максимальное, количество яиц. В дейст-
вительности многие морские птицы откладывают по одному яйцу
и не откладывают нового, если оно гибнет. Размер кладки должен
иметь наследственную основу, поэтому если в течение короткого
времени пищи оказывается достаточно, то как отбор может предот-
вратить распространение генов, под действием которых птицы от-
кладывают по 2 яйца? Даже если мы предположим, что в процессе
эволюции установился признак «только 1 яйцо» — хотя трудно по-
нять, как это может произойти, если не допустить, что это благопри-
ятно не только для всей колонии, но и для особей в отдельности,—
то популяция с этим признаком будет внутренне неустойчивой сис-
темой, поскольку ей будет угрожать распространение новых мута-
ций, дающих кратковременные преимущества отдельным 'особям.
Хороший обзор этого положения и представлений Уинн-Эдвардса
в целом написан Уинсом [498].
В своих последующих работах Уинн-Эдвардс [515] сам признал
малую вероятность существования механизмов группового отбора.
Эволюция общественного поведения и явного альтруизма гораздо
правдоподобнее объясняется непосредственными преимуществами,
получаемыми отдельной особью, которые усиливаются отбором ро-
дичей и совместно с последним влияют на совокупную приспособлен-
ность [210]. Тем не менее Уинн-Эдвардс был первым, привлекшим
внимание к взаимоотношениям между поведением и структурой по-
пуляций животных, и его идеи, хотя и отвергнутые позже, оказали
воздействие на многие более поздние исследования общественного
поведения.
Один из четких примеров эффекта, существование которого он
предполагал, дает исследование шотландских куропаток на верес-
ковых пустошах Шотландии. Уотсош486] и его коллеги показали, что
размер популяции этих куропаток определяется территориальным
поведением. Осенью петухи дерутся на токовых площадках (а мы
теперь знаем, что чем лучше местообитание, тем сильнее они де-
рутся), и какая-то часть их захватывает территории. Потерпевшие
неудачу птицы изгоняются из долины. Следующей весной размно-
жаются только владельцы территорий; птицы, оказавшиеся лишни-
ми, остаются по сути стерильными; во всяком случае, у них зареги-
стрирована более высокая смертность в тех неблагоприятных ус-
ловиях, куда их заставили выселиться. Территориальное поведение
ограничивает эффективный размер популяции и ставит ее в зави-
симость от наличия пищевых ресурсов вересковой пустоши в дан-
ный период.
Надежное подтверждение взаимосвязи между поведением и регу-
ляцией размеров популяции получить нелегко. Здесь недостаточно
нескольких сезонных исследований, проведенных на одном виде.
Для того чтобы получить однозначный ответ, необходимо проводить
наблюдения в течение примерно 25 лет. Ценность гипотезы Уинн-
10*