Меннинг О. Поведение животных. Вводный курс
Подождите немного. Документ загружается.

218
Глава 6
Эволюция 219
I
вида медведок высокоспецифичны; они наверняка играют роль в
половой изоляции между видами (с. 250). Характер песни частично
определяется структурой и размерами передних крыльев, которые
трутся друг о друга, вызывая звук — гребень заднего края левого
крыла трется о зазубренную нижнюю поверхность жилки на правом
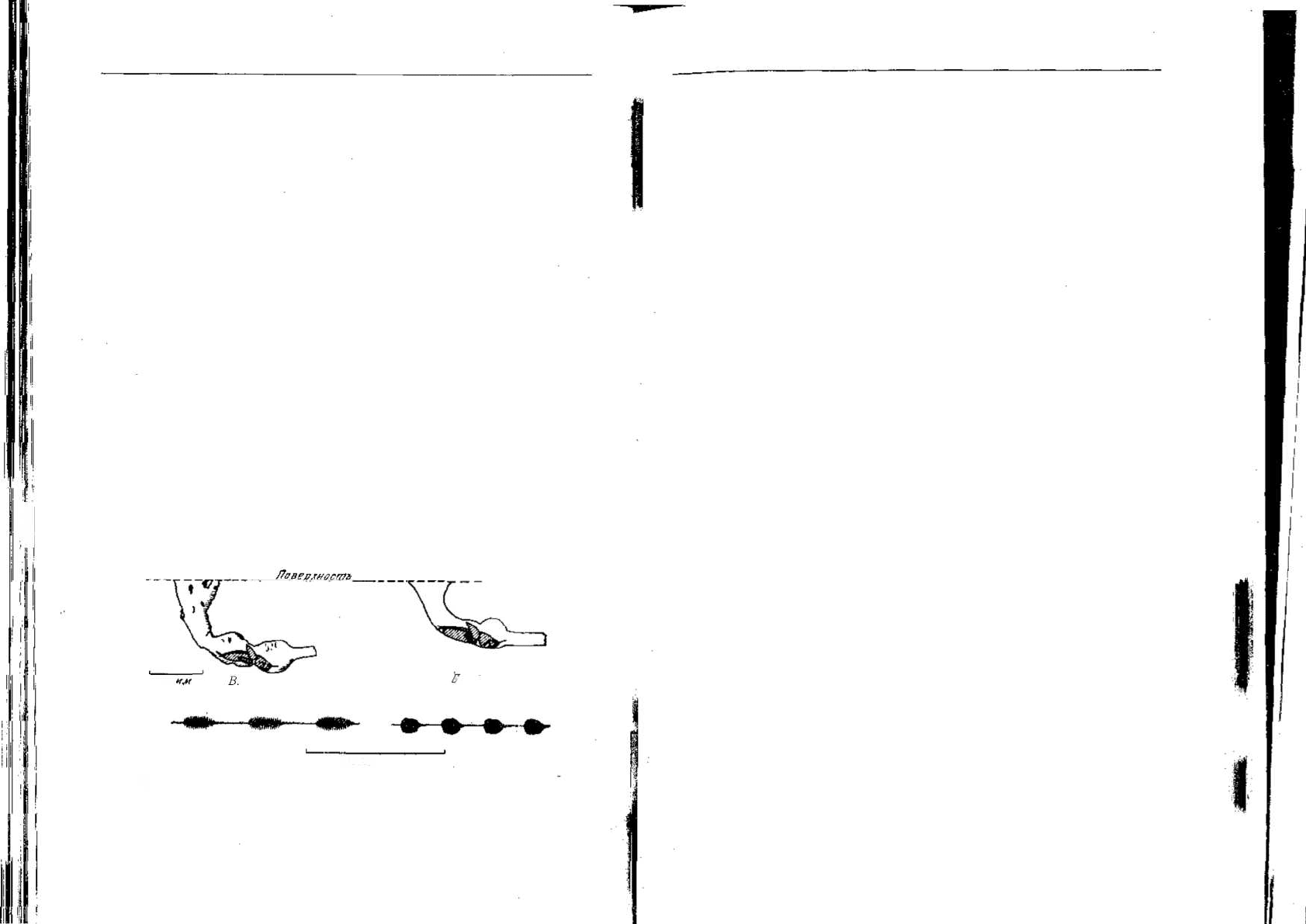
крыле. Из двух видов, изученных Беннет-Кларком [47, 481, у G. gry-
llotalpa, вида с маленькими крыльями, мелкими зубцами гребня и
характеризующегося тихой песней, основная частота песни равна
—1600 Гц (рис. 6.1), тогда как у G. vineae — вида с большими крыль-
ями, глубокими зазубринами и характеризующегося значительно
более громкой песней, она равна 3500 Гц.
И в данном случае мы видим прямую корреляцию между поведе-
нием и морфологией, однако адаптивность проявляется не только в
этом. Самцы каждого из видов роют норки особого типа, показан-
ные на рис. 6.1. Беннет-Кларк обнаружил, что норка имеет форму
рога с расширением на конце, причем физические свойства этого
пространства соответствуют частоте песни, способствуя ее наилуч-
шему распространению.
Во время исполнения песни медведка располагается головой
вниз, так что звук возникает в самом основании рога, как это показа-
но на рисунке. Таким образом, строение крыла, пение и рытье нор-
ки тончайшим образом приспособлены для того, чтобы обеспечить
наиболее эффективную генерацию звука, предназначенного для
привлечения пролетающих над норкой самок.
Еще один пример развития специализированного поведения в
соответствии с требованиями внешней среды животного мы находим
у обыкновенной чайки. Речь идет о ее манере уносить из гнезда скор-
лупу яиц. Раздражители, на которые в этой ситуации реагируют
5q
gryllotalpa
• vtaeae
50 мс
Рис. 6.'). Вид сбоку самцов медведок (Gryllotalpa) двух видов, сидящих в своих
норках [48]. Их голова во время пения обращена вниз. Форма норки напоминает
форму рога. У G. vineae норка с более гладкими стенками, и самец этого вида ис-
полняет более громкую песню. Вместе с тем в обоих случаях форма норки приспо-
соблена к частоте песни (осциллограммы песен приводятся в нижней части рисун-
ка), благодаря чему достигается наибольшая эффективность сигнализации.
птицы, были описаны в гл. 3. Впервые размножающиеся чайки бы-
стро уносят скорлупу, хотя раньше они, естественно, с подобной
ситуацией не сталкивались. Эта деятельность занимает у чаек всего
5 мин в год, н в силу этого ее роль может показаться незначительной.
Тем не менее Тинберген и его сотрудники [4681 четко показали, что
скорлупу важно удалять для маскировки гнезда. В колониях чаек
на птенцов часто нападают вороны, горностаи и лисы, причем эти
хищники отыскивают птенцов с большей легкостью при наличии
поблизости яичной скорлупы как из-за ее запаха, так и из-за белизны
внутренней поверхности (рис. 3.6). Действие естественного отбора
обеспечило надежное выполнение родителями краткого, но жизненно
необходимого комплекса действий, связанного с удалением скорлу-
пы. Очевидно, никакой поведенческий акт, если он с регулярностью
наблюдается в естественных условиях, не может не обладать какой-
то функцией.
Для осуществления многих поведенческих актов требуется, что-
бы они очень точно соответствовали аналогичным актам других осо-
бей. Взаимная адаптация обнаруживается в большинстве случаев
коммуникации, о которой мы говорили в гл. 3. В демонстрации уха-
живания, например, реакции одного из партнеров выступают в
качестве фактора, который в процессе отбора формирует поведение
другого партнера, и наоборот.
Чанц и Хирсбруннер-Шарф [472] очень хорошо показали, как
адаптированы друг к другу реакции птенцов и родителей у чистико-
вых. Они исследовали два вида этих птиц — кайру и гагарку,—
которые гнездятся на одних и тех же утесах, но используют для этого
разные места. Гнезда кайр располагаются близко друг от друга на
плоских уступах, тогда как гагарки гнездятся одиночно, используя
трещины и маленькие ниши. Птенцы кайр стремятся сохранять до-
статочно выпрямленную позу, пытаясь удержаться на утесе, кото-
рый обычно бывает мокрым и покрыт слоем помета. Позы птенцов
гагарок более горизонтальны; щели, где располагаются их гнезда,
бывают суше и лучше защищены. Это поведение не зависит от опыта
и не изменяется, если птенцов обоих видов переносят на одну и ту же
поверхность или если птенцы вылупляются в несвойственном для
них месте (при взаимном обмене яиц между этими двумя видами).
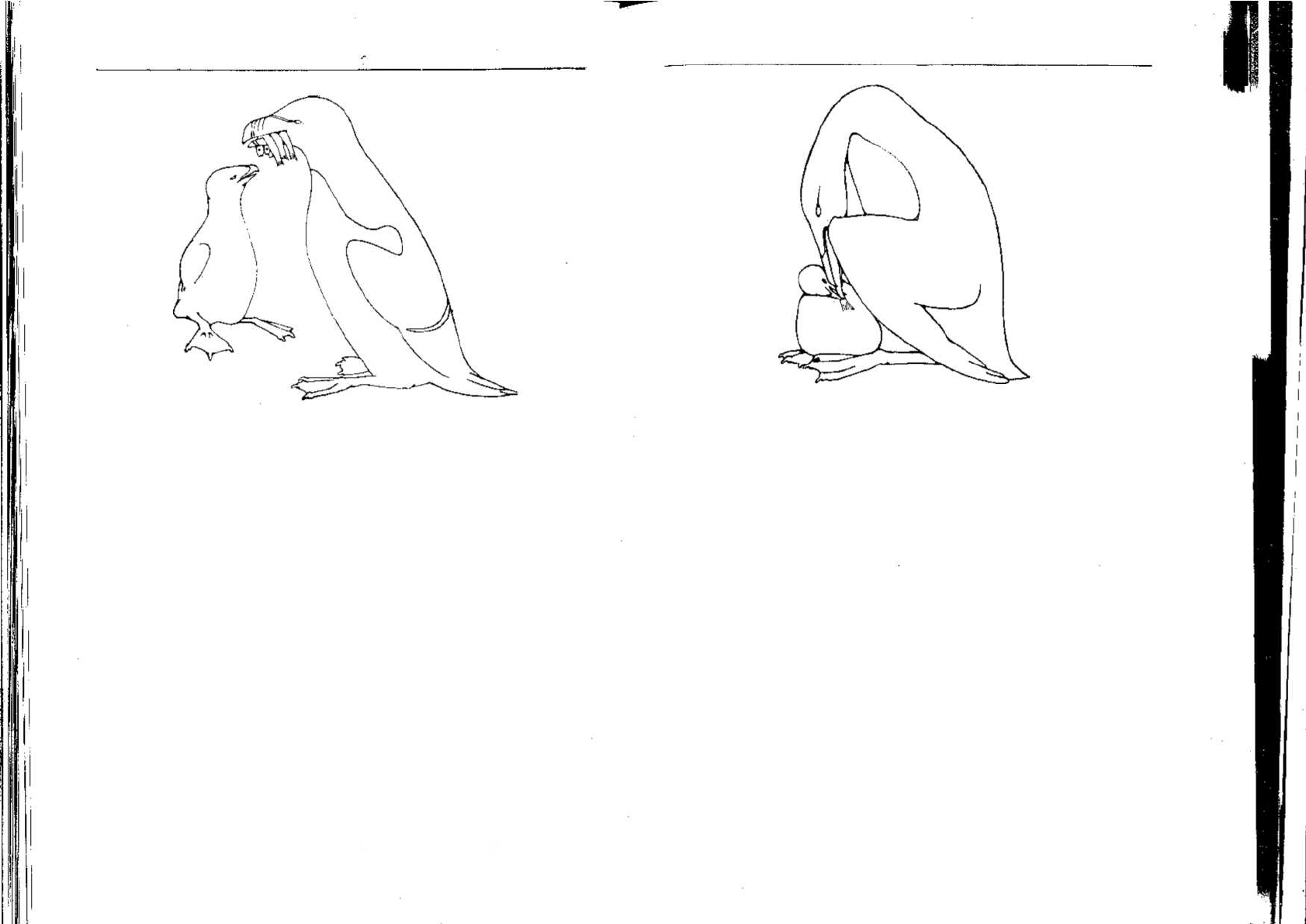
Родители обоих видов приносят птенцам рыбу, однако способы
кормления у них значительно различаются (рис. 6.2 и 6.3). Гагарки
приносят несколько рыб и довольно пассивно стоят в то время, когда
их птенцы клюют пищу. У родителя и птенца достаточно свободного
места, и во время кормления им никто не мешает. Родители кайры
показывают птенцу сначала только хвост рыбы, закрывая кормяще-
гося птенца крыльями и лапами. Птенец пропускает рыбу через
клюв, а затем проглатывает ее, начиная с головы. Такая церемония
кормления обеспечивает некоторую защиту и предотвращает на
густо населенных обрывах воровство.
218
Глава 6 it
Эволюция
220
Рис. 6.2. Гагарка приносит птенцу рыбу, которую он спокойно берет прямо из
родительского клюва 1472].
Рис. 6.3. Способ кормления птенцов у кайр [472]. Птенцу сначала показывается
только хвост рыбы. Птенец его хватает, пропускает через клюв всю рыбу, а затем
проглатывает ее, начиная с головы.
При обмене яиц между гнездами кайр и гагарок (операция, кото-
рую можно успешно провести, если предварительно в течение не-
скольких дней постепенно удалять отметины на яйцах) можно наблю-
дать реакции птенцов и приемных родителей. Во всех случаях птенец
сначала проявлял видоспецифическое поведение, что вызывало боль-
шие трудности при кормлении, в особенности у птенцов гагарок,
реакции которых совершенно не соответствовали церемонии корм-
ления у кайр. Только через довольно большой промежуток времени
приемные родители и птенцы начинали постепенно приспосабливать-
ся друг к другу, а многие из птенцов погибали.Эти результаты указы-
вают на жизненно важное значение развивающихся в процессе эволю-
ции взаимных реакций родителей и птенцов. Мы не всегда осознаем
необходимость подобной взаимной адаптации, когда наблюдаем
гладкопротекающие взаимодействия между родителями и детьми,
хотя совершенно очевидно, что многие животные не могли бы обес-
печить выживание только путем обучения и тренировки.
Тонкость поведенческой экипировки животных, обусловленная
естественным отбором, производит сильное впечатление. Тем' не
менее нам следует помнить, что эволюция редко затрагивает изоли-
рованно какой-либо один признак, особенно признак поведения.
В конечном счете развитие определяется всем организмом в целом,
и имеющуюся наследственную изменчивость отбор использует,
чтобы получить от признака не максимум возможного, а наилучший
компромиссный вариант с учетом остальных признаков.
В гл. 3 мы отметили, что можно изготовить искусственный рели-
зер, который был бы более эффективным, чем естественный. В роли
естественного релизера, например, мог бы выступать участок крыла
бабочки, которое служит также и для полета, а отбор должен найти
компромиссный вариант между этими признаками на уровне ниже
максимального их выражения. Сходную ситуацию мы встречаем
у самцов с их брачным нарядом и песнями. Яркое оперение и гром-
кая песня самца, рассчитанные на то, чтобы на него обратила внима-
ние самка, привлекают также и хищника. Большинство птиц прекра-
щает пение с началом насиживания, а у некоторых брачное оперение
меняется при осенней линьке. Между успехом в размножении и
опасностью нападения хищника должен обязательно существовать
компромисс. Он может устанавливаться на разных уровнях в зави-
симости от обстоятельств. У полигамных видов, таких, как фазаны,
у которых не все самцы достигают успеха в размножении и один
может иметь нескольких самок, развилось исключительно экстра-
вагантное брачное оперение, которое сохраняется у них в течение
всего года. В то же время самцы фазанов не участвуют в насиживании
яиц и воспитании птенцов; этим занимаются только покровитель-
ственно окрашенные самки. Ярко раскрашенный самец, который

г
222
Глава
Эволюция
223
привлекает за сезон шестерых самок, спаривается с ними, а затем
становится жертвой лисы, оставит больше потомков, чем самец с
неярким оперением, который проживет 10 лет, но не привлечет ни
одной самки.
Изучая поведение чаек, Тинберген и его сотрудники обнаружили
прекрасные примеры того, как оно изменяется, чтобы лучше соот-
ветствовать условиям окружения. При исследовании моевок, Rissa
tridactyla, проведенном Каллен [112], выявилось множество их от-
личий от других чаек. Моевки гнездятся на узких уступах скал и
тем самым защищают свои яйца и птенцов от хищников, взимающих
значительную дань с птиц, гнездящихся на земле. Однако подходя-
щие места для гнезд малочисленны, и за них идет ожесточенная кон-
куренция. На обрывах происходят длительные драки, в которых
участвуют и самцы, и самки; иногда сцепившиеся птицы падают в
воду, где они продолжают бить друг друга крыльями. Правда, у
других чаек такие яростные и настойчивые драки неизвестны. Вы-
сокий уровень агрессивности влияет у моевок не только на драки,
но и на демонстрации ухаживания. В них проявляется значительная
доля агрессивных элементов, и в отношениях между самкой и сам-
цом долго после начала сезона размножения сохраняется открытая
агрессивность. В противовес такой агрессивности получили большое
развитие демонстрации умиротворения. Они включают в себя отвора-
чивание головы и опускание шеи — пример умиротворения по типу
«антиугрозы», о котором мы говорили в гл. 5. Наиболее распростра-
ненный тип угрозы — это «кашляние», также уже описанное в гл. 5.
Мы рассмотрели там данные, свидетельствующие о том, что когда
кашляние обнаруживается у других чаек, которым оно не свойст-
венно, это указывает на сильное стремление птиц остаться на месте.
У моевок привязанность к месту гнездования и его защита должны
быть наиболее интенсивными. В соответствии с этим естественный
отбор способствовал формированию компромиссного варианта между
стремлениями к нападению и избеганию, причем с большей склон-
ностью к нападению, чем у других чаек.
Каллен обнаружила у молодых и взрослых птиц значительное
число других модификаций поведения, которые можно связать с
гнездованием на утесах. Позднее Каллен и Ашмол [115], а также
Хейлмен [191] обнаружили сходный ряд адаптаций у гнездящейся
на обрывах тонкоклювой крачки и еще у одного вида чаек — у гала-
пагосской вилохвостой чайки.
«КУЛЬТУРНАЯ» ПРЕЕМСТВЕННОСТЬ В ПЕРЕДАЧЕ НАВЫКОВ
В гл. 2 мы обсуждали, каким образом гены и факторы внешней
среды участвуют в формировании поведения взрослого животного.
Поведение может эволюционировать только, если оно изменяется
и если его изменения передаются от одного поколения к другому.
В таких случаях начинает действовать естественный отбор, способ-
ствующий распространению одних вариантов и уничтожению дру-
гих. Для большинства морфологических и физиологических призна-
ков существует только один способ осуществления таких наслед-
ственных изменений—это генетические изменения, связанные
с мутациями или рекомбинацией уже имеющихся генов.
В процессе эволюции поведение изменяется еще одним спосо-
бом — животные способны обучаться некоторым навыкам от своих
родителей или от других членов группы. Позднее это поведение в
свою очередь послужит моделью, в соответствии с которой будет
модифицировать свое поведение уже потомство данной особи. Пред-
положим, что животное обучилось совершенно новому комплексу
действий, который значительно лучше удовлетворяет некоторой
цели, чем более раннее типичное поведение. Этот комплекс может
передаваться последующим поколениям и постепенно заменять ста-
рый комплекс без каких бы то ни было генетических изменений.
Эволюцию поведения мы начнем рассматривать с примеров этого
относительно редкого, но потенциально очень важного способа пере-
дачи поведения у животных. Мы знакомы с эволюцией культуры
у человека, и многие склонны утверждать, что почти все наиболее
важные человеческие свойства передаются от одного поколения к
другому именно таким способом. Различные языки — очевидный
пример непрерывающейся культурной традиции, в соответствии с
которой в разных популяциях поддерживаются различные типы
поведения.
Разумеется, развитие традиций возможно только у животных,
способных к модификации поведения путем подражания и практики.
Соответствующие примеры обнаружены у наших ближайших род-
ственников — приматов. Интенсивные и длительные наблюдения
за японскими макаками, Масаса fuscata, показали, что наблюдаемые
между отдельными стаями различия в поведении несомненно проис-
ходят вследствие различий в традициях. Обезьян часто привлекает
искусственно предлагаемая пища — зерно; сладкий картофель и
т. п. К картофелю, как правило, прилипает земля, которую обезьяны
стирают ладонями. Однажды наблюдатели заметили, как одна моло-
дая самка стала опускать картофелины в ручей и их мыть. Она со-
хранила эту привычку, и ей стали подражать сначала один из ее
детенышей, а затем почти все молодые члены ее стаи, и теперь это
явление получило название «субкультуры мытых овощей». Таким
путем между стаями возникли большие различия в способах
питания. Фриш [155] сделал обзор некоторых японских работ на
эту тему. Нет сомнений, что при внимательном наблюдении за при-
матами других групп выявится сходная картина не обязательно в
отношении привычек питания, а, быть может, в отношении исполь-
зования простых орудий, которые несомненно в каждом новом поко-
лении у шимпанзе осваиваются заново. Некоторые элементы комп-

218
Глава 6
Эволюция
219
лексов социального поведения также передаются по традиции [186,
318].
Мы легко признаем существование передачи навыков поведения
по традиции у приматов, однако растет число данных, показываю-
щих, что спорадически она встречается и у других позвоночных.
Галеф [159] сделал хороший обзор этой проблемы. Различные диа-
лекты песен у белоголовой овсянки, описанные в гл. 2, наверняка
поддерживаются благодаря передаче традиций. Молодые самцы
копируют все особенности песни своего отца и ближайших соседей.
Если же самец вырастает в другом месте, то он, как мы знаем, усва-
ивает иной «диалект».
Недавно Нортон-Гриффите [370, 371] обнаружил замечательный
случай передачи по традиции пищевого навыка у кулика-сороки,
Haematopus ostralegus. Эти береговые птицы питаются в большей
степени мидиями и открывают раковины двумя четко различными
способами. При одном птица ударяет клювом по вентральной сторо-
не раковины, которую для этого переносит с камней на подходящий
для этого участок плотного песка. При втором способе птица разре-
зает моллюска клювом через приоткрытые створки, причем моллюск
остается прикрепленным к камням и покрытым морской водой, Нор-
тон-Гриффите показал, что каждая птица пользуется только каким-
нибудь одним способом. Молодые птицы сопровождают родителей
на мидиевые банки и постепенно перенимают их способ, которым
затем исключительно и пользуются. Разные мидиевые банки разли-
чаются по тому, удобно ли на них применять разбивание или разре-
зание, и кулики-сороки выбирают те области, которые больше под-
ходят для освоенного ими метода. Казалось бы при этом создается
основа для настоящей изоляции между разбивающими и разрезаю-
щими мидии птицами, однако, как подчеркивает Нортон-Гриффите,
помимо мидий кулики-сороки употребляют в пищу много других
объектов. К передаче пищевых навыков по традиции относится и
известный случай открывания молочных бутылок синицами [220]
и способ вытаскивания семян из незрелых плодов Daphne rtiezereum
зеленушками — добывать семена из зрелых плодов этого растения
невозможно [381].
Передача навыков по традиции могла влиять и на эволюционное
развитие поведения, поскольку не исключено, что при этом изменя-
лись силы отбора, действующие на животных. Харди [200] проил-
люстрировал это ярким, хотя и гипотетическим примером. Предпо-
ложим, что группа приматов занимает участок, примыкающий к
морю, и начинает использовать для сбора пищи морской берег. Одно
из животных начинает заходить в воду и даже плавать для добывания
пищи. Этот навык распространяется, как распространилось мытье
овощей (в одной из стай японские макаки действительно плавают
в море), и таким путем передачи навыка по традиции группа прима-
тов начинает собирать морскую пищу. Это сразу же отразится на
ходе естественного отбора — будет происходить уменьшение шер-
стяного покрова, развитие жировой подкожной клетчатки и пере-
понок на лапах, т. е. физиологическая адаптация к плаванию. Лю-
бая из этих и другие адаптации к водному образу жизни — которые
ранее не имели никакого значения и, возможно, были бы даже небла-
гоприятными — приобретают высокую селективную ценность про-
сто потому, что изменилось поведение.
Нам не известны примеры эффектов столь сильных, как описан-
ный, но, во всяком случае, среди приматов они должны существо-
вать, хотя могут остаться и не обнаруженными. Однако, несмотря
на то что передача навыков по традиции иногда очень важна, она
встречается достаточно редко, и, как правило, эволюция поведения
происходит при отборе наследственных изменений. В связи с этим
прежде чем обсуждать, какие типы изменений происходят в процес-
се эволюции, необходимо познакомиться с тем, что известно о гене-
тике поведения,
ГЕНЫ И ПОВЕДЕНИЕ
Генетика поведения — очень сложная область исследования.
В генах содержится вся информация, определяющая строение бел-
ков, производимых клеткой. Если исследователя интересует биохи-
мическая генетика, то он довольно просто может установить цепь
причин между геном и нужным биохимическим параметром, так
как эта цепь часто бывает короткой и достаточно ясной. Гораздо
труднее установить причинную связь между генами и морфологиче-
скими вариациями, поскольку в генетике поведения «расстояние»
между генами и конечным продуктом больше, чем где бы то ни было.
На поведение влияет множество факторов: общее состояние обмена
веществ, скорость выделения гормонов, физиология и морфология
мышечной и нервной систем — список этот может быть сколь длин-
ным, столь и сложным. И вот в этом запутанном клубке различных
связей необходимо установить однозначное соответствие между гене-
тическими различиями и различиями в поведении. Разнообразие
явлений поведения еще более усложняет проблему, поскольку нам
известно, что существует генетическое влияние на мотивацию, обу-
чение, общую реактивность, чувствительность к определенным раз-
дражителям, на выполнение фиксированных комплексов действий
и др. Не приходится ожидать, что в развитии такой разнообразной
группы показателей может быть общий генетический путь. Однако
эти трудности не мешали значительному прогрессу генетики поведе-
ния. Хорошие обзоры на эту тему имеются в книгах Фуллера и Томп-
сона [157] и Эрман и Парсонса [142]. Вместе с тем утверждать, что
эта область знаний упорядочена, отнюдь нельзя.
Поведение включает в себя координированный контроль ряда
различных систем организма. На примере плодовой мушки дрозофи-
8
л» 147

11
1!
226
Глава 6
Эволюция
227
И
г
V
'I
лы, изученной генетически лучше, чем любое другое животное,
можно видеть, что гены влияют на поведение разными способами.
У дрозофилы известно много сотен мутантных генов, причем поло-
жение их в хромосомах картировано. Например, ген Ваг определяет
уменьшение числа глазных фасеток в сложных глазах, ген white —
ослабление пигментации глаз, гены forked и hairy детерминируют
число и строение щетинок, гены vestigial и dumpy — изменение фор-
мы крыльев, гены yellow и black — общую пигментацию тела и
т. д.
Названия генов приблизительно соответствуют наиболее замет-
ным из детерминируемых ими эффектов. У генов есть, конечно, и
другие проявления, выявляемые при исследовании обмена веществ,
фертильности, длительности жизни и т. п. Было обнаружено, что
большая часть упомянутых выше генов определяет по меньшей мере
одну из сторон поведения — например, обладающие ими самцы,
менее успешно спариваются с самками, чем нормальные мухи. Одна-
ко, хотя поверхностное описание их влияния на поведение позволяет
всех их объединить в одну категорию, очевидно, один и тот же конеч-
ный результат возникает совсем разными путями. Гены Ваг и white
влияют на зрение: мутантные мухи не видят так хорошо, как нор-
мальные, и им трудно и отыскивать самок и ориентироваться на зри-
тельные сигналы, поступающие от них во время ухаживания. Щетин-
ки — это чувствительные органы осязания, и самцы с генами forked
и hairy имеют соответствующий сенсорный дефект. Важная часть
ритуала ухаживания самца дрозофилы — это вибрация крыла в
горизонтальной плоскости при отведении его вбок. Такая вибрация
возбуждает у самки чувствительные окончания в основании антенн.
Самцы с генами vestigial и dumpy имеют сильно деформированные
крылья и не могут надлежащим образом ими вибрировать. Неудиви-
тельно, что они гораздо реже участвуют в спаривании. Менее оче-
видно, почему самцы с генами yellow и black спариваются реже, чем
нормальные мухи. Показано, что самок отвращает не их аномаль-
ный цвет, поскольку в полной темноте они также обладают очень
низкой конкурентной способностью. В сенсорных органах или в
строении крыльев у них нет никаких явных аномалий, хотя специ-
альными исследованиями это еще не доказано. Не исключено, что
гены yellow и black определяют функционирование нервной системы,
мышц или общего обмена веществ насекомых.
До недавнего времени было невозможно выявить пути влияния
генов на поведение, за исключением случаев очень выраженных
эффектов. У человека гены, ответственные за микроцефалию и фе-
нилкетонурию, обусловливают огромные дефекты в строении и био-
химии мозга, результатом чего является умственная отсталость.
У мышей известно много генов, ответственных за нарушение позы
и равновесия. «Вальсирующие мыши» носятся кругами, когда их
потревожат, другие постоянно качают головой или кружатся. Эти
изменения в поведении коррелируют с дефектами структуры сред-
него уха и мозговых центров, ответственных за чувство равновесия.
Наблюдаемые при этом изменения очень велики и вредны, и по этой
причине они не относятся к важным для эволюции поведения гене-
тическим изменениям, которые, по-видимому, слабо выражены и
благоприятны для организма.
Икеда и Каплан [248, 249] исследовали менее резкие изменения
в поведении и провели их более тонкий анализ. Они изучали группу
мутантов дрозофилы, суммарно именуемых hyperkinetics, поскольку
у них бывают аномально сильные вздрагивания лапок при наркозе,
а при ходьбе обнаруживаются неожиданные подергивания. Исполь-
зуя тонкие нейрофизиологические методы, они выделили в латераль-
ной части грудного нервного сплетения группу мотонейронов, обла-
дающих аномально высокой скоростью спонтанного разряда. Нео-
жиданные залпы активности этих нейронов совершенно независимы
от внешних сигналов. Похоже, что Икеде и Каплану удалось локали-
зовать место действия гена в нервной системе. По предварительным
данным аномальная частота разрядов возникает из-за изменений
в мембране этих нейронов. В этом случае мы действительно можем
объяснить изменения в поведении — хотя еще и достаточно силь-
ные — в понятиях нейрофизиологического действия генов.
В настоящее время идентифицировано и картировано значитель-
ное число генов, имеющих чисто поведенческие эффекты. Когда
дрозофил подвергают действию сильного мутагена, а их потомство
скринируется для выявления отклонений в определенном поведе-
нии, мутации обнаруживаются редко, поэтому необходимо исследо-
вать большое число мух, и, следовательно, поведенческий тест дол-
жен быть относительно простым. Ряд тестов основан на том, что нор-
мальные мухи движутся в направлении источника света, а случайно
возникший мутант так не делает, из-за чего его легко обнаружить.
В других тестах учитывается то, что нормальные самцы осуществля-
ют спаривание в течение нескольких минут, а тех мух, которые не
способны к этому, легко изолировать и использовать в дальнейших
исследованиях.
Бензер [50] и его сотрудники использовали остроумную методику
скринирования генов, детерминирующих фототаксис, хемотаксис,
циркадные ритмы, ухаживание, способность к обучению и т. д.
Мутанты по этим генам потенциально представляют ссбой наиболее
ценный материал для исследования путей влияния генов на поведе-
ние, причем в некоторых случаях удается выявлять и место действия
гена.
Этого добиваются, применяя некоторые удивительные генетиче-
ские приемы, позволяющие получать «мозаичных» особей дрозофил,
часть клеток которых несет изучаемую мутацию, тогда как остальные
клетки нормальны [233, 234]. Исследуя большое число мозаичных
особей, у которых мутантными оказываются разные участки тела,

218
Глава 6
выявляют, в каких именно участках тела мухи должен присутство-
вать мутантный ген, для того чтобы его эффект обнаружился. Напри-
мер, гены, влияющие на циркадные ритмы, проявляются только в
том случае, если мутантной оказывается ткань мозга. Тот же метод
используют для выяснения вопроса, какая часть нервной системы
должна иметь генотип самки или самца, чтобы обнаружилось специ-
фичное для данного пола поведение [234].
Изучение действия отдельных генов — это важное направление
в генетике поведения, однако, если мы хотим понять механизмы
эволюции поведения, необходимо изучить наследование самих комп-
лексов действий. При этом по возможности используют методы клас-
сической генетики. Различающихся по поведению животных скре-
щивают и исследуют их гибриды первого (F) и второго (F
2
) поколе-
ний, а также гибриды, полученные в результате возвратных скре-
щиваний (беккроссы), и т. д.
Здесь сразу же возникает затруднение с выбором для генетиче-
ского анализа «единиц поведения». Это пример одной из проблем,
которые всегда стояли перед генетиками. Гениальное озарение поз-
волило Менделю выбрать для исследования у гороха признаки, кото-
рые очень четко выражены и поддаются подсчету. В случае гороха
каждой единице — высоте растения, цвету семян, текстуре оболочки
семени и т. д.— соответствует один ген, однако маловероятно, чтобы
в случае поведения тоже была бы такая простая корреляция.
Сформулировать правила для выбора «единиц поведения» невоз-
можно. Обычно такой выбор подсказывается самими генетическими
исследованиями. Лучше всего, пожалуй, проводить генетические
исследования комплексов фиксированных действий, поскольку они
четко выражены и достаточно хорошо поддаются подсчету. Более
того, мы знаем, что они устойчивы к воздействиям внешней среды.
Хотя рассматриваемые комплексы представляются филогенетиче-
скими единицами, это не значит, что они обязательно имеют простую
генетическую основу. Разумеется, есть все основания полагать, что
их развитие определяется действием MHOFHX генов, однако частота
выполнения таких комплексов действий или легкость их провока-
ции детерминируются довольно простыми формами наследования,
а это позволяет обнаружить их единую структуру.
Один из немногих примеров в этой области — работа Ротенбухле-
ра [400, 401] на медоносных пчелах. Некоторым линиям пчел свой-
ственно «гигиеническое поведение», состоящее в том, что, когда
личинка гибнет в ячейке, рабочие особи вскрывают ячейку и выбра-
сывают личинку. При отсутствии гигиенического поведения пчелы
так не делают и ячейки остаются закрытыми. Все гибриды между
линиями не проявляли гигиенического поведения, что, видимо, сви-
детельствует о доминантном характере признака негигиенического
поведения. При возвратном скрещивании гибридов с особями гигие-
нической рецессивной родительской линии Ротенбухлер получил
Эволюция
219
замечательные результаты. В 29 колониях, полученных в результате
возвратных скрещиваний, он обнаружил следующее:
1. В девяти колониях пчелы вскрывали ячейки с мертвыми ли-
чинками, но оставляли их в улье.
2. В шести колониях они не вскрывали ячеек, однако выбрасы-
вали личинок, если ячейки вскрывал экспериментатор.
3. В восьми колониях пчелы не вскрывали ячеек и не выбрасыва-
ли личинок, т. е. не обладали гигиеническим поведением.
4. В шести колониях пчелы вскрывали ячейки и удаляли личи-
нок, т. е. обнаруживали гигиеническое поведение.
Соотношение этих четырех классов колоний примерно одинаково,
поэтому результаты можно объяснить исходя из действия двух пар
аллелей, одна из которых ведает комплексом действий по вскрыва-
нию ячеек, а другая — удалением мертвых личинок. Аллели, опре-
деляющие отсутствие гигиенического поведения, доминантны. Обо-
значим их «£/» для вскрывания и «R» для удаления, тогда их «гигие-
нические» аллели будут обозначаться как «ш> и «г». Рабочая пчела
гигиенической линии имеет генетическую конституцию иигг, а неги-
гиенической — UURR. Все рабочие пчелы и матки гибридов должны
иметь генотип UuRr и не обладают гигиеническим поведением. По-
скольку самцы пчел (трутни) гаплоидны, т. е. имеют только один
набор хромосом, для получения колоний беккроссов Ротенбухлер
скрестил 29 гибридных трутней с матками двойной рецессивной
гигиенической линии. Трутни имели все четыре комбинации аллелей:
UR, иг, Ur и uR, а результат скрещивания был следующим:
Трутни
UR
Ur
uR
иг
Матки
1
, uuRr
1
, Г
l
U ' (
1 2 3 4
(группа в вышеприведенном списке)
Простота полученного результата не означает, что развитие
нервных сетей, регулирующих комплексы действий вскрывания
ячеек и удаления личинок, контролируется одним локусом. По дан-
ным Ротенбухлера, рабочие пчелы, не обладающие гигиеническим
поведением, в действительности осуществляют такие комплексы дей-
ствий, однако с очень низкой частотой и при действии более сильных
раздражителей. Аллели U и и действуют как переключатели, опре-
деляющие порог комплекса действий вскрывания ячейки фактически
по типу «все или ничего».
Мы знаем и другие примеры комплексов фиксированных дей-
ствий, которые наследуются полностью или не наследуются совсем.
X
иигг

218
Глава 6
Эволюция
230
Демонстрация ухаживания у уток состоит из серии комплексов
действий, большую часть которых можно наблюдать почти у всего
семейства. Одну такую демонстрацию Лоренц [304] назвал «вниз-
вверх». Она заключается в том, что селезень опускает клюв в воду,
а затем резко поднимает голову, всплескивая воду. Эта демонстра-
ция отсутствует у нырка, Nettion flavirostre, и шилохвости, Defila
acuta, хотя имеется у многих их родственников. В то же время она
проявляется во всей полноте у гибридов F
x
между этими двумя вида-
ми [308, 481]. Наиболее вероятное объяснение данных результатов
состоит в том, что группа генов, необходимых для демонстрации
«вниз-вверх», имеется у обоих родительских видов, однако естествен-
ный отбор выключил проявление этой демонстрации, повысив ее
порог. Это повышение порога было достигнуто у обоих видов с по-
мощью разных групп генов. При комбинации этих групп генов в
гибридах Fj их влияние уменьшилось, порог снизился и данная
демонстрация стала проявляться в нормальной обстановке. Для
подтверждения этой гипотезы необходимо исследовать гибриды F2 и
беккроссы, но, к сожалению, как это нередко бывает, эти гибриды
совсем не дают потомства.
Мы привели два примера влияния генов на пороги, когда их
действие по типу переключателя определяет, проявится или нет
данный комплекс действий. Известны и другие гены-переключатели,
контролирующие действие групп генов, которые наследуются как
единый блок [434]. У некоторых бабочек в процессе эволюции на
крыльях появились рисунки, имитирующие узор на крыльях насе-
комых неприятного вкуса, благодаря чему бабочки приобрели неко-
торую защиту от хищников. Развитие таких узоров находится под
контролем многих генов, однако имеется единственный ген-переклю-
чатель, который определяет, будут они работать или нет. Подобно
этому, в поведении можно ожидать, что естественный отбор благопри-
ятствует группировке в блок тех генов, которые вносят свой вклад
в проявление поведенческого признака во всей его полноте, если он
оказывается полезным для особи. Чем теснее связаны такие гены, тем
меньше опасности, что у особей будет отсутствовать какая-то их
часть и развитие окажется несовершенным.
Работа Юинга [150] подтверждает это положение. В ней показа-
но, что специфический характер брачной «песни» (исполняемой сам-
цом дрозофилы при демонстрации ухаживания с помощью вибрации
крылом) у двух видов дрозофил — Drosophila pseudoobscura и D.
persimilis — определяется Х-хромосомой. F,— гибридные самцы
исполняют «песню» материнского вида, поскольку свою Х-хромосо-
му они получают от матери. В детерминировании характера песни
должно участвовать много генов, и их совместное расположение в
Х-хромссоме вряд ли случайно. Мысль о том, что генные блоки вооб-
ще характерны для системы, ответственной за наследование комплек-
сов фиксированных действий, крайне привлекательна. Чтобы дока-
ill
зать это, необходимы дополнительные исследования, однако найти
для них подходящий генетический объект очень трудно.
Наиболее интересно то, что когда у животного комплексы фикси-
рованных действий проявляются в определенной последователь-
ности, у гибридов часто нарушается именно последовательность
комплексов действий, а сами комплексы остаются без изменений.
Необычное гнездостроительное поведение гибридных попугаев-
неразлучников, описанное в гл. 2, служит этому примером. Лоренц
[307] и Рамсей [389] описали поразительные примеры такого рода
в поведении ухаживания у уток. У селезня кряквы, Anas platyrhyn-
chos, так же, как и у других кормящихся на поверхности воды уток,
репертуар ухаживания состоит из десяти высокостереотипных комп-
лексов действий, часть которых иллюстрирует рис. 6.4. Приведен-
ную последовательность: «встряхивание клюва», «свист с ворчанием»,
«потряхивание хвостом» — Лоренц назвал обязательной последо-
вательностью, хотя Рамсей отмечает, что иногда у кряквы и у ее
близкого родича — американской черной утки, Anas rubipres —
он наблюдал порядок: «потряхивание хвостом, встряхивание клю-
ва, свист с ворчанием». У селезней-гибридов этих двух видов Рамсей
наблюдал последовательность: «свист с ворчанием, потряхивание
хвостом, встряхивание клюва» и другие комбинации, никогда не
наблюдавшиеся ни у одного из родительских видов. Ясно, что гене-
тический контроль последовательности выполнения комплексов
независим от контроля самих фиксированных действий, однако мы
слишком мало знаем о механизмах поведения и поэтому более деталь-
но рассмотреть это положение пока не в состоянии. В своей работе
на гибридах меченосцев Xiphophorus Франк [153] обсуждает не-
сколько других примеров аномальных комбинаций комплексов
фиксированных действий и дает также хороший обзор наблюдений за
гибридами вообще.
В то время как гены-переключатели, о которых говорилось выше,
очень сильно влияют на уровень порогов поведенческих актов, зна-
чительно чаще обнаруживаются гены, вызывающие относительно
небольшие количественные изменения. Мы уже упоминали о сниже-
нии успеха в спаривании самцов дрозофилы, Drosophila melanogaster,
имеющих ген yellow. Бесток [21] показала, что ухаживание этих
самцов слабее стимулирует самок, чем ухаживание нормальных,
потому что они реже осуществляют вибрацию, хотя в остальном их
демонстрация ничем не отличается от нормы. У дрозофилы известно
еще несколько примеров подобного типа, поскольку у нее легко по-
лучать линии, различающиеся только по одному гену [325].
В большинстве исследований, выполненных на других животных,
используются или инбредные линии, которые различаются большим
числом генов, или линии, которые были отобраны на различия по
произвольно выбранному признаку поведения. В этих случаях реги-
стрируют количественные различия в частоте выполнения одних и

„Встряхивание
„ Свист с ворчанием"
„ Потряхишание .
Рис. 6.4. Одна из наиболее частых демонстраций ухаживания у утки-кряквы, состоящая из последовательно использу-
е
мых тр
ех ком
пл
ексов
д
е
й
ст
в
и
й [307]. У селезней-гибридов эти комплексы выполняются в иной последовательности.
а & я ь -о
o s
и
я я
Jg%
9
^ о
а = * ss еь
'Р
™
т.
*
ггг
А £ = о п
- н Ol V
p-ril
Е s о Е -I
К 0» X'
.
к
О~ ' "
Ш
р
<•0
§
а
л

218
Глава 6
вали, как в результате селекции изменился уровень агрессивности
мышей (гл. 4). Сходным путем и удивительно быстро — всего за
три поколения — произошло изменение частоты, с которой домашние
петухи осуществляют комплексы полового поведения [511]. Мен-
нинг [324] проводил в течение ряда поколений селекцию дрозофилы
по признаку малой и большой скорости спаривания (рис. 6.5). Ско-
рость спаривания — это комплексный признак, который включает
в себя взаимодейтвие самца и самки, причем анализ показывает,
что при селекции происходит количественное изменение поведения
у особей обоего пола. Самцы из «быстрых» линий чаще выполняли
более быстрые движения ухаживания, чем самцы из «медленных»
линий. И наоборот, самки из «быстрых» линий легче стимулирова-
лись самцами как своей, так и других линий, чем самки из «медлен-
ной» линии.
Вместе с изменением частоты выполнения комплексов фикси-
рованных действий при селекции изменяются также более «общие»
стороны поведения, такие, например, как уровень локомоторной
активности и эмоциональности. Рундквист [408] отбирал две линии
крыс, проявлявших соответственно высокую и низкую активность
в колесе, присоединенном к их клетке, куда они могли по желанию
входить. Он получил очень небольшие изменения в сторону усиления
активности, но добился очень больших изменений в сторону ее ос-
лабления — через 12 поколений селекции крысы неактивной линии
совершали за 15 дней по 6000 оборотов колеса, тогда как контроль-
ные крысы совершали за это время по 100 000 оборотов. Хотя у нас
нет конкретных данных, скорее всего изменения подобного типа
влияют на многие стороны поведения крыс. Это относится, в ча-
стности, к эмоциональности, которую тщательно изучал Бродхэрст
[68, 69]. Он отбирал крыс на эмоциональность и ее отсутствие (или
на реактивность и нереактивность крыс, как он это называл) в об-
становке, вызывающей у животных умеренный страх, т. е. при поме-
щении их на ярко освещенную площадку. Раньше мы уже упоминали
об этом тесте «открытого поля», когда говорили о влиянии раннего
опыта на эмоциональность (гл. 2). Реактивными называли тех крыс,
у которых в «открытом поле» был высокий уровень дефекации и мо-
чеиспускания и которые оставались неподвижными, часто распласты-
ваясь на одном месте. К ним, по-видимому, относятся животные,
характеризующиеся высоким уровнем физиологической активации
(гл. 5). У нереактивных крыс не было дефекации, и они свободно раз-
гуливали по площадке. В экспериментах Бродхэрста отбор оказался
весьма эффективным. Полученные им линии различались по ряду
физиологических и поведенческих показателей. Например, реактив-
ные крысы медленно обучались простой реакции избегания удара
электрического тока — их обучению мешала развившаяся у них
интенсивная активация (гл. 7).
Успех селекционного эксперимента означает, что естественные
Эволюция
235
популяции содержат большое число генов, влияющих на поведение
в количественном отношении. В процессе естественного отбора реак-
ции поведения поддерживаются на оптимальном уровне, и вместе
с тем благодаря характерной для животных громадной изменчивости
может осуществляться отбор. Изменчивость — это основа эволюции
и, как мы увидим далее, изменения, возникающие при так называе-
мой микроэволюции поведения (например, на ранних стадиях видо-
образования), достаточно просто связать с влияниями мутаций и
искусственного отбора, которые мы только что рассмотрели. Однако,
прежде чем обсуждать микроэволюцию, необходимо познакомиться
с еще одним аспектом генетики и эволюционного учения, который
имеет важное значение для поведения.
ОТБОР РОДИЧЕИ И СОВОКУПНАЯ ПРИСПОСОБЛЕННОСТЬ
Как высказался Сэмюэль Батлер, курица — это способ, которым
яйцо производит другое яйцо. Это утверждение может показаться
странным, но оно содержит важную биологическую истину. Сущест-
вует точка зрения, согласно которой размножаются и оставляют по-
томков только гены, а тело — просто сложная упаковка, создающая
им защиту и повышающая шансы на успех. Эту точку зрения умно и
решительно отстаивает в своей книге «Ген-эгоист» (The selfish gene)
Доукинс [122]. Для выяснения полезности этой концепции нет необ-
ходимости рассматривать все аргументы автора. Если за единицу
отбора принимать гены, а не организм, то размножение и родитель-
ское поведение можно рассматривать как средство, увековечивающее
гены. У каждого потомка половина генов происходит от одного
из родителей, и оставление двух потомков в среднем обеспечивает
постоянство частоты любого из генов от поколения к поколению. До
этого момента наши рассуждения могли представляться достаточно
академичными, поскольку с таким же успехом мы можем рассуждать
в терминах более знакомого нам представления о размножении ор-
ганизмов. Однако идея отбора генов наводит на мысль о еще одной
возможности, поскольку генами данной особи кроме нее самой обла-
дают не только ее непосредственное потомство, но и другие род-
ственники. Сюда же относится одно послеобеденное замечание Хол-
дейна, который, подсчитав что-то быстро на обороте меню, восклик-
нул: «Я готов пожертвовать жизнью во имя четверых внуков или
восьмерых кузенов». Если кто-то имеет гены, ответственные за пове-
дение самопожертвования, то у таких родственников, о которых го-
ворит Холдейн, в среднем будет столько же соответствующих генов,
сколько у него самого.
Этот подход служит основой важной концепции, согласно кото-
рой отбор благоприятствует альтруизму, т. е. жертвованию соб-
ственными непосредственными интересами, даже жизнью, если вслед-
ствие этого достаточное число животных, родственных сданным, уве-

218
Глава 6
Эволюция
236
личит свои шансы на выживание. Нам знакома эта точка зрения для
случая, когда родители приносят свои интересы в жертву для сохра-
нения потомков, однако теперь мы можем распространить ее и на
родственников. Приспособленность особи приблизительно выража-
ют как число оставленных потомков. Приспособленность гена (или
генотипа) можно расширить, включив в нее всех выживших особей,
которые имеют его копии, и это мы назовем «совокупной приспособ-
ленностью». Смит [439] в своей книге, содержащей блестящий обзор
современной эволюционной теории, для описания отбора, в котором
учитываются не только непосредственные потомки, но и другие
родственники, предлагает термин «отбор родичей».
Отбор родичей часто затрагивает гены, контролирующие пове-
дение, особенно поведение, направленное на других животных. В по-
следнее время возрос интерес к роли этого отбора в эволюции всех
типов общественного поведения. Этот интерес в большой степени
был вызван двумя статьями Гамильтона [199]. Гамильтон обратился
к проблеме, долгое время озадачивавшей зоологов. У насекомых
истинная общественная жизнь, т. е. обитание в сообществах с пере-
крывающимися поколениями, стерильными рабочими особями и
совместным выкармливанием потомства (гл. 8), развилась только
в двух отрядах — у Isoptera (термиты) и Hymenoptera (муравьи,
пчелы, осы). Самое замечательное состоит в том, что по существую-
щим данным у перепончатокрылых общественный образ жизни воз-
никал независимо одиннадцать раз, а может быть, и больше. Ясно,
что появление сообщества в эволюции — процесс не простой, иначе
сообщества обнаруживались бы и в других группах насекомых;
по-видимому, данный отряд — перепончатокрылые — имел к этому
какое-то предрасположение.
Гамильтон обратил внимание на уникальный характер детерми-
нирования пола у перепончатокрылых. У них имеется гаплодипло-
идия, т. е. самцы гаплоидны и развиваются из неоплодотворенных
яиц, а самки диплоидны и развиваются нормальным путем из оплодо-
творенных яиц. Диплоидный организм получает половину хромосом
от матери, половину от отца. При образовании гамет, которые впо-
следствии развиваются в спермин и яйцеклетки, происходит редук-
ционное деление (мейоз), в процессе которого число хромосом умень-
шается вдвое. Гомологичные материнские и отцовские хромосомы
составляют пары и обмениваются участками генетического материа-
ла. Затем хромосомы, составляющие пары, в случайном порядке
расходятся в дочерние клетки. Это означает, что, хотя в каждой
гамете имеется полный гаплоидный набор хромосом, доли материн-
ских и отцовских генов в нем варьируют, но в среднем они состав-
ляют по 50% . У гаплоидного самца перепончатокрылых не происхо-
дит редукционного деления с уменьшением числа хромосом вдвое;
все его гаметы, следовательно, идентичны и содержат просто копии
его набора хромосом.
Одна из наиболее поразительных черт колонии перепончато-
крылых заключается в том, что она обычно состоит из одной семьи —
из матки с ее многочисленным потомством, большую часть которого
составляют самки. Гамильтон проанализировал влияние гапло-
диплоидии на генетические взаимоотношения внутри такой семьи.
Поскольку нормальное потомство получает по гаплоидному набору
хромосом от каждого из родителей, можно сказать, что его срод-
ство и с матерью, и с отцом составляет '/
2
. Как мы говорили выше,
гаметы, из которых образуется потомство, варьируют по своему ге-
нетическому составу и в среднем содержат 50% одинаковых генов.
Разные потомки одних и тех же родителей имеют в среднем общими
половину отцовских и половину материнских генов. Таким образом,
среднее генетическое сродство одного сибса с другим составляет
7
2
XV
2
(от отца) + V2XV2 (от матери) = 7
2
.
Поэтому, насколько это касается генов, особь столь же близко род-
ственна своему сибсу, как и родителям. Дочь медоносной пчелы
развивается из оплодотворенного яйца своей матери — пчелиной
матки,— и ее родство с матерью, как и обычно, составляет '/
2
, одна-
ко родство между сестрами не такое. Поскольку все гаметы их от-
ца — гаплоидного трутня — идентичны между собой, все пчелы-
дочери имеют 50% общих генов, полученных ими от отца. Из полу-
чаемых от матери генов у них также 50% общих, так что их родство
друг с другом равно
х
/
2
(
от
отца) + '/
2
X
V
2
(от матери) =
3
/
4
.
Следовательно, самки перепончатокрылых имеют с сестрами больше
общих генов, чем с матерями.
В сообществе перепончатокрылых размножается только очень
небольшое число дочерей, большую же его часть составляют сте-
рильные рабочие особи, которые помогают матери выращивать не-
скольких молодых привилегированных маток. Однако, как подчер-
' кивает Гамильтон, в понятиях репликации генов это — благопри-
ятное обстоятельство. Если перед молодой самкой в эволюционном
плане имеется «выбор» между тем, чтобы покинуть колонию и вос-
питывать собственных дочерей или остаться в ней и помогать матери
в выращивании все большего числа сестер, она выберет последний
вариант.
Хотя наверняка у насекомых существовали и другие предпо-
сылки для эволюции общественного поведения [267], можно не сом-
неваться, что предположение Гамильтона о важной роли гаплодип-
лоидии в удивительном развитии перепончатокрылых правильно.
Отбор родичей проявляется с особой силой в связи с особенностями
механизма определения пола. Подробнее с гипотезой Гамильтона
можно ознакомиться в работе Вильсона [506]. Имея представление
об отборе родичей, мы можем в несколько ином свете рассмотреть
эволюцию общественного поведения, и в особенности актов явного
альтруизма или самопожертвования. Отбор не может благоприят-
ствовать жертвованию приспособленности особи во имя неродствен