Меннинг О. Поведение животных. Вводный курс
Подождите немного. Документ загружается.

128
Г лава 4
Мотивация
159
падение. Однако если отдыхающая кошка не видит противника, от-
правится ли она на его поиски? О птицах иногда говорят, что они
«патрулируют» территорию и нападают на встреченного чужака.
Вместе с тем нельзя с уверенностью утверждать, что начало патру-
лирования связано с агрессивной мотивацией. Для этого мы должны
были бы показать, что от момента начала патрулирования и до того,
как птица встретит соперника, пороги агрессивных реакций снижа-
ются, правда, соответствующие измерения провести довольно
трудно.
Однако, рассматривая эту проблему под несколько другим углом
зрения, можно придти к заключению, что при некоторых условиях
агрессивное поведение обладает свойствами подкрепления, т. е.
возбужденное животное осуществляет поведение, способствующее
его агрессивной реакции. Видимо, наиболее изящные примеры, сви-
детельствующие в пользу этого положения, описаны в работе Томп-
сона по бойцовым петухам [453] (гл. 5) и сиамским бойцовым рыбкам
[452]. Важно отметить, что эти животные были отобраны на высокую
агрессивность. Самцы бойцовой рыбки при виде своего зеркального
изображения или модели проявляют полную агрессивную демонстра-
цию с поднятыми жаберными крышками и развернутыми плавниками
и при первом предъявлении им этого раздражителя нападают на него
и кусают. Томпсон держал самцов в резервуарах, где они видели
свое зеркальное изображение в стенке аквариума при простом вык-
лючении внешнего освещения. Подобно тому как в скиннеровских
камерах обучают крыс, он вырабатывал у рыб условный рефлекс.
Крыса для получения пищи должна нажимать на рычаг, чему она
вскоре обучается. Рыбке подвешивали в воде кольцо, и когда она
проплывала через него, лампа автоматически на несколько секунд
выключалась. В стекле при этом появлялось изображение рыбы, на
которую она реагировала агрессивно. В данных условиях самцы обу-
чались проплывать через кольцо и осуществляли это многократно,
по нескольку сот раз в день. Другими словами, когда самцам пре-
доставлялась возможность выбора обстановки, они выбирали та-
кую, в которой могли вести себя агрессивно. Точно так же бойцовые
петухи обучались реакции, которая давала им возможность выпол-
нять агрессивную демонстрацию. Можно возразить, что наличие в
аквариуме кольца само стало стимулом к агрессии, что поддержива-
ло рыбу в состоянии возбуждения. Томпсон показал, что искусствен-
ные предметы, ассоциирующиеся с самцом-соперником, сами по
себе вызывают атаку на них. Даже если это так, нам представляется
неизбежным вывод о том, что для рыбы агрессивное поведение слу-
жит «подкреплением», и, следовательно, в этом отношении сравне-
ние тенденции к драке со стремлением к еде и питью вполне оправ-
данно.
СУЩЕСТВУЕТ ЛИ ПОСЛЕ ВЫПОЛНЕНИЯ АГРЕССИВНОГО АКТА
ФАЗА ПОКОЯ?
Стереотипные комплексы действий при агрессивных демонстра-
циях и атаках можно рассматривать как эквиваленты завершаю-
щих актов, направленных на цель, т. е. на соперника. Однако после
агрессивного поведения трудно обнаружить регулярное возникно-
вение фазы покоя. При других типах поведения, например при еде
и питье, обычно комплекс действий начинается с короткого периода
активации и «разминки» (или «врабатывания», гл. 1).Мыши, напри-
мер, кормятся короткими периодами, прерываемыми паузами, при-
чем длительность таких периодов в первые несколько минут еды,
пока мышь не насытилась, возрастает [499]. Это значит, что, пока
не поступило сигнала обратной связи о «достижении цели» и не нас-
тупила фаза покоя, интенсивность поведения временно увеличивает-
ся. При агрессивном поведении такая активация бывает очень ин-
тенсивной, и если атаки начались, то порог проявления последую-
щих атак часто значительно снижается.
Севенстер [432] и Уилц [508] измеряли агрессивность самца тре-
хиглой колюшки, используя метод, описанный на с. 152. Они обна-
ружили следующий существенный факт: стремление самца кусать
соперника, показываемого в стеклянной пробирке, к концу 1 0-ми-
нутного периода было выше, чем в начале. Если мы дадим животно-
му неограниченное количество пищи па 10 мин, то этого не обнару-
жится. Интересно отметить, что поразительная активация агрессив-
ности при раз начавшихся атаках прямо противоположна картине,
наблюдаемой при биогенных побуждениях. Уилц обнаружил, что в
течение нескольких минут после тестирования агрессивности самец
колюшки остается настолько агрессивно возбужденным, что не спо- Л
собен реагировать на самку. Если самку показать самцу сразу после
исчезновения агрессивных раздражителей, то самец выполняет не-
сколько действий, функция которых, по-видимому, заключается в
снижении агрессивности. После выполнения этих действий его си-
стема половой мотивации способна функционировать. Таким обра-
зом, совсем не очевидно, что вслед за выполнением агрессивного по- II
ведения и устранением соперника наступает фаза покоя. При пос-
тоянном действии вызывающего агрессию раздражителя атаки пре- II
кращаются, однако при изменении раздражителя они возобновля-
ются, подобно тому как смена раздражителей немедленно восстанав-
ливает половое поведение быка, у которого до этого обнаружива-
лось явное половое истощение. Возможно, в таких случаях угасание
реакции правильнее рассматривать как привыкание (с. 259), а не
как покой в результате снижения уровня мотивации.
Таким образом, полученные на животных данные показывают,
что в отличие от побуждения к еде и питью результатом агрессив- ^^^п
ного поведения часто бывает усиление стремления к нападению.
128
Г лава 4 Мотивация 161
Данный вывод достаточно важен как в теоретическом, так и в прак-
тическом отношении, поскольку споры о природе агрессии в связи с
точкой зрения Лоренца о путях контроля агрессии у человека и жи-
вотных продолжаются. Мы обсудим этот вопрос в следующем раз-
деле.
НЕИЗБЕЖНА ЛИ АГРЕССИЯ?
Вопрос формулируется именно гак, поскольку при описании аг-
рессии Лоренц использовал свою психогидравлическую модель,
а в этой модели единственным способом снижения побуждения ока-
зываются выполнение соответствующего поведения, тогда как в от-
сутствие такого «выхода» побуждение накапливается и усиливается.
Если у человека имеется наследственно детерминированное
стремление к агрессии, которое осуществляется по подобному прин-
ципу, то мы должны признать, что избежать всех проявлений агрес-
сии невозможно. При этом, видимо, наилучшим выходом была бы
сублимация или направление агрессивных стремлений по менее
вредоносным путям, нежели путь физического конфликта. Этого
взгляда в сущности и придерживаются Лоренц и Ардри.
Часто для доказательства неизбежности, с которой проявляется
агрессия, приводят множество различных экспериментов, однако
все их можно интерпретировать по-разному. Выращенные в полной
изоляции животные иногда бывают очень агрессивными. Это явле-
ние четко продемонстрировано у мышей и у диких кур, причем у по-
следних Круйт [278] обнаружил, что после нескольких месяцев пре-
бывания в изоляции они начинают нападать просто на перья и, по-
долгу кружась, дерутся с собственными хвостами. Однако хорошо
известно, что у выращенных в изоляции животных сильно изменя-
ются различные типы поведения, поэтому объяснять эти данные
только накоплением стремления к агрессии было бы неоправданно.
Так, например, содержавшиеся в изоляции грызуны обычно очень
возбудимы, причем известно, что у них происходят гормональные
изменения, возможно, как следствие стресса от обстановки, в кото-
рой они находятся. В результате в гонадах самцов может выделять-
ся больше тестостерона, что приводит к усилению стремления напа-
дать.
Лоренц [309] описывает наблюдения за очень агрессивными цих-
лидовыми рыбками Etroplus maculatus. Любители рыб методом проб
и ошибок установили, что для того, чтобы пара Etroplus успешно
размножалась, в аквариуме нужно держать еще одного-двух не учас-
твующих в размножении самцов, которые играют роль «мальчиков
для порки». Участвующий в размножении самец время от времени на
них нападает, однако между членами пары агрессии почти не бывает.
Если же пару Etroplus держать одних, они редко успешно размно-
жаются, поскольку самец постоянно нападает на самку. В хорошо
50
4-0
Г
t>
- 20-
-5
46,57 46,57
13,2/
ML
4,36
Общая
агрессивность
Ш
Агрессия на
другие особи, а
не на партнера
i ул Агрессия',
I'. ;] направленная
на партнера
Группа А
Группа Б Группа В
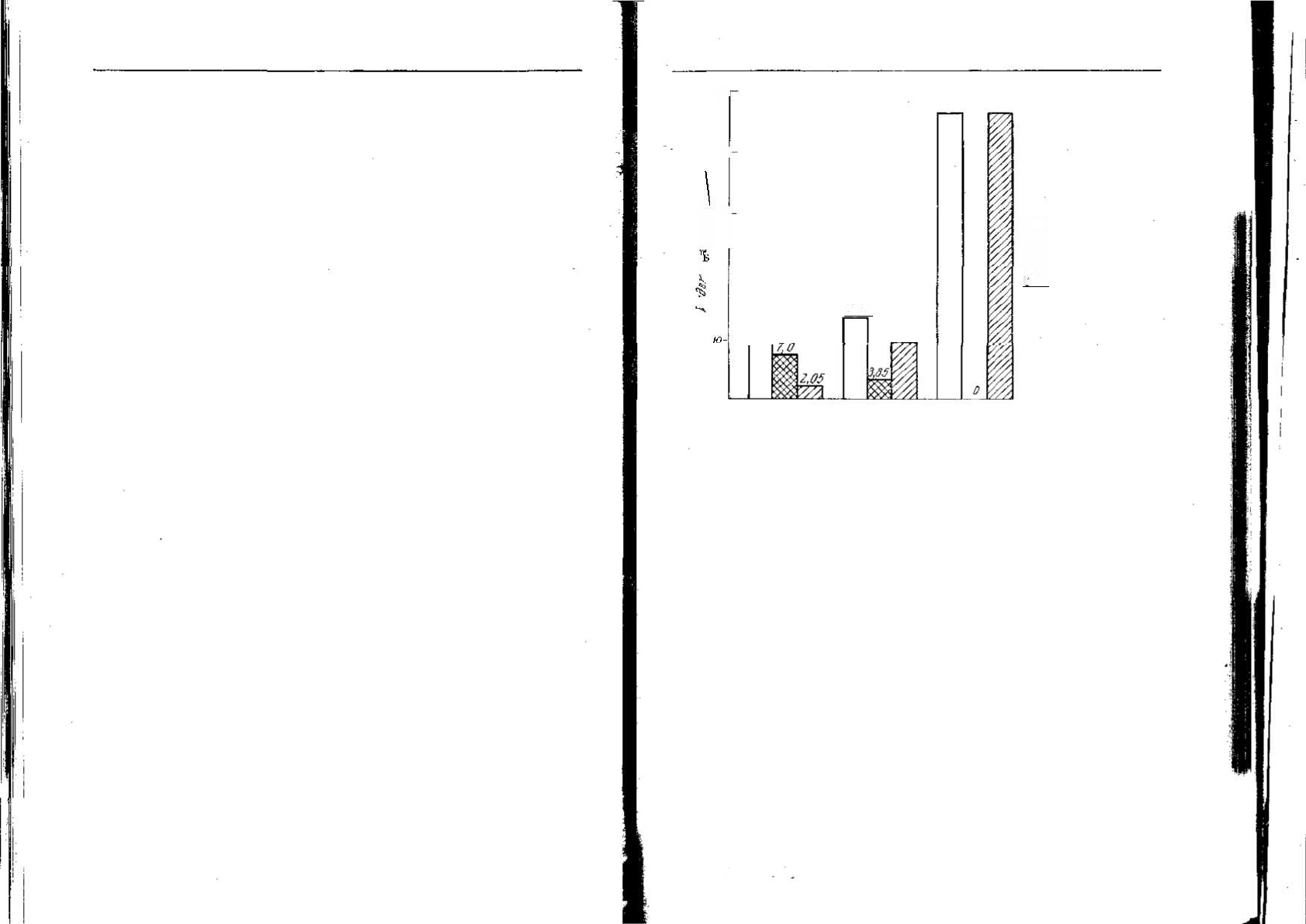
Рис. 4.9. Данные Раза по агрессивному поведению Etroplus maculatus [391]. Ин-
декс агрессивности отражает усредненное в течение 5-минутных периодов число
нападений пары. В группе А пара содержалась в аквариуме с другими рыбами.
В группе Б пара содержалась отдельно, но они могли видеть других рыб через
стекло. В группе В пары были полностью изолированы. Оценивались две категории
агрессивных действий — направленные на других рыб и на партнера (в группе В
только вторая категория).
контролируемых условиях эти данные были подтверждены Раза
[391], результаты которой показаны на рис. 4.9. Если нападать
больше не на кого, то число направленных на самку атак резко уве-
личивается. Лоренц [309] и Эйбл-Эйбесфелдт [143] объясняют эти
результаты накоплением у самца агрессивности, которая должна
найти выход. Однако снижение порога реакции нападения не обя-
зательно означает накопление агрессивности. Как мы будем подроб-
нее говорить в следующей главе, у самцов территориальных живот-
ных присутствие самки обычно в дополнение к половым реакциям
вызывает усиление агрессии. Скорее всего самцы Etroplus во всех
вариантах опытов Раза были активированы. Их агрессивность до
некоторой степени тормозилась тем, что самка вызывает также и по-
ловое стремление, и если присутствовали другие рыбки, то большая
часть атак была переадресована им. Такое переадресование извест-
но в агрессивном поведении человека и животных в случаях, когда
раздражитель, вызывающий агрессию, в силу какой-то причины не
может быть объектом нападения: начальник оскорбляет своего под-
чиненного, а тот в свою очередь дает пощечину посыльному.
Из экспериментов Раза мы видим, что, когда пара содержится в
6 № 147
о

128
Г лава 4
Мотивация
162
изоляции, число нападений очень сильно возрастает. Автор пред-
полагает, что самец Etroplus, так же как и колюшки, наиболее аг-
рессивен вблизи гнезда. Самка же стремится быть около гнезда,
если на нее нападают. Можно, следовательно, ожидать, что самец
около гнезда активирован в максимальной степени. В обычных ус-
ловиях он нападает на других рыб издалека и постепенно удаляется
от гнезда. Затем, поскольку рыбы, на которых он нападает, старают-
ся как можно скорее скрыться с глаз, агрессивность самца Etroplus,
вероятнее всего, снижается до того, как появляется следующий
раздражитель. Чтобы критически проанализировать концепцию ак-
кумуляции энергии и неизбежности агрессивной реакции, требуется
значительно больше информации о поведении рыб, у которых акти-
вация в течение разных периодов времени поддерживается на мини-
мальном уровне.
В нескольких работах на рыбах была сделана попытка получить
такую информацию. Клзйтон и Хайнд [100] наблюдали ход восста-
новления реакций нападения у сиамских бойцовых рыбок. Самцу
предъявляли его зеркальное изображение до тех пор, пока он не
переставал на него реагировать. Для восстановления реакции тре-
бовалось много дней, и, хотя это может быть следствием постепенно-
го накопления агрессивной мотивации, гораздо проще рассматри-
вать данное явление как ослабление процесса привыкания, посколь-
ку мы знаем, в какой степени агрессия зависит от раздражителей.
Вилхелми [501] сходным образом исследовал восстановление реак-
ций нападения у меченосцев, которым перед этим предоставлялась
возможность сражаться с соперником до тех пор, пока один из них
не уступал. И опять период изоляции рыбок на срок от 1 до 8 нед
приводил к постепенному восстановлению реактивности. Вилхелми
предполагает, что причиной таких изменений может быть либо на-
копление побуждения, либо привыкание.
В противоречии с этими данными Хейлигенберг [208] на цихли-
довых рыбках Pelmatochromis обнаружил, что в отсутствие актива-
ции у них происходит ослабление стремления к драке в течение не-
скольких дней. Отметим, что эти рыбки не дрались до полного исто-
щения. Результаты Хейлигенберга не подтверждают точки зрения,
согласно которой в отсутствие возможности драться агрессия всегда
накапливается.
Концепцию Лоренца критиковали исследователи поведения мле-
копитающих, которые отмечали его тенденцию недооценивать влия-
ние на агрессивную мотивацию факторов развития. Скотт [426] дает
четкий обзор многочисленных экспериментов на грызунах, в кото-
рых показано, как воздействием на животных в раннем возрасте мож-
но чрезвычайно сильно изменить уровень их агрессивности. Отно-
сительно просто научить одну мышь всегда нападать на чужака,
тогда как другое животное той же линии можно научить оставаться
совершенно спокойным. Оказывается, изменения в агрессивности,
которые можно получить, создавая различные условия в раннем
возрасте, сходны с изменениями, полученными в селекционном экс-
перименте Лагершпец. Поскольку условия выращивания животных
столь четко влияют на уровень агрессии и поскольку данные, полу-
ченные на животных, неоднозначны во многих отношениях, у нас,
по-видимому, нет оснований соглашаться с тем, что агрессивность
человека неизбежна. Очевидно, мы просто должны признать, что
у человека имеется некая, возможно, наследственная потенциаль-
ная агрессивность, однако при решении вопроса о неизбежности аг-
рессии всегда следует исходить из наличия у человека длительного
периода развития в детстве, а также того сильного влияния, которое
оказывают на человека родители и общество.
ФИЗИОЛОГИЧЕСКАЯ ОСНОВА МОТИВАЦИИ
Продолжение исследований поведения, подобных тем, которые
обсуждались выше, совершенно необходимо, однако важно также
изучить мотивацию на физиологическом уровне и попытаться свя-
зать поведение с определенными событиями, происходящими в нерв-
ной системе. Некоторые из наиболее многообещающих мостов меж-
ду нейрофизиологией и поведением были возведены благодаря рабо-
там, посвященным проблемам мотивации в области физиологической
психологии, которые были выполнены преимущественно американ-
скими учеными. Прекрасное введение в курс физиологической пси-
хологии написано Гроссманом [182]. Ниже мы рассмотрим некото-
рые из исследований в этой области, а начнем с описания того участ-
ка мозга позвоночных, о котором точно известно, что он является
наиболее важным в контроле мотивации, а именно с гипоталамуса.
ГИПОТАЛАМУС
Эта небольшая по объему область (в мозгу человека она по разме-
рам не превышает последней фаланги мизинца) абсолютно необхо-
дима для всего комплекса поведенческих реакций. Блестящую об-
щую оценку гипоталамуса дал Уэлш [482]; ...«этот центр играет до-
минирующую роль в определении путей использования ресурсов ор-
ганизма... Ни одну функцию организма нельзя себе представить вне
прямой или косвенной зависимости от гипоталамуса».
Здесь необходимо привести некоторые сведения по нейроанато-
мии. Более подробное и точное описание нервной системы можно
найти в книге Ромера [397].
Мозг всех позвоночных построен по единому плану. На ранних
стадиях эмбрионального развития он состоит из трех вздутий перед-
него конца спинного мозга. Они называются prosencephalon, mesen-
cephalon и rhombencephalon, или передний, средний и задний мозг.
Эти вздутия возникли в связи с необходимостью принимать возрас-

128
Г лава 4
- Эписриз
•Моомечо/г
Обоцр/пельюя
Мутвица
Зрительный тракт Птот^амуе
(перерезан)' J |
.Щеп I Ме$ея
Telencephalon 'cepfrulon I серяа/ол
\ ,
Prosencephalon
; Meten ,
сер/га Тсп Mye/encepiia/on
fihoMbencephalon
Рис. 4.10. Основные'отделы мозга позвоночных ([397] с изменениями). Мозг позво-
ночных проходит через стадию развития, сходную с изображенной на рисунке,
в особенности мозг птиц и млекопитающих. В мозгу взрослого животного благода-
ря своим большим размерам доминируют большие полушария и мозжечок Они
закрывают все остальное и не позволяют видеть первоначальный план строения.
тающее количество сенсорной информации, приходящей в централь-
ную нервную систему от органов чувств головы. Передний мозг
первоначально был связан с обонянием, средний мозг — со зрени-
ем, а задний — со слухом и чувством равновесия. У большинства
ныне живущих позвоночных исходные функции этих отделов очень
расширились и усложнились, однако соответствующие сенсорные
сигналы идут сначала именно в эти отделы, даже если они после это-
го передаются еще куда-либо.
Как показано на рис. 4.10, передний мозг четко подразделяется
на два отдела. Из верхней части (крыши) переднего отдела развива-
ются полушария мозга, которые первоначально были обонятельны-
ми областями, но у млекопитающих стали доминировать над всей
нервной системой. На дорсальной поверхности заднего отдела перед-
него мозга — diencephalon (или промежуточного мозга) расположен
пинеальный орган (шишковидная железа). Когда-то он был связан
с рецептором света, или теменным глазом, который можно увидеть у
некоторых из современных пресмыкающихся. Боковые стенки про-
межуточного мозга утолщены и образуют зрительные бугры, или
таламус — важное «место переключения», где в многочисленных
«ядрах», или скоплениях тел нейронов, соединяются друг с другом
многочисленные проводящие пути. В основании промежуточного
мозга, под таламусом, откуда и пошло его название, расположен
гипоталамус.
Ядра имеются и в гипоталамусе, но они не столь четко очерчены,
как в лежащем выше таламусе. В гипоталамус входят и из него вы-
ходят несколько хорошо заметных проводящих путей, которые сое-
диняют его с полушариями мозга, а также с ниже расположенными
Мотивация
164
отделами. Одной из наиболее существенных структурных особеннос-
тей, имеющих большое значение при изучении поведения животных,
является его тесная связь с гипофизом. Эта железа внутренней сек-
реции, управляющая всей гормональной системой организма (с. 176),
развивается в результате слияния спускающегося вниз выпячива-
ния гипоталамуса и поднимающегося вверх выпячивания ротовой
полости эмбриона. В ножке гипофиза, связывающей его с гипотала-
мусом, проходят нервы и кровеносные сосуды. Сам гипоталамус
обильно снабжается кровью, и капилляры проникают даже в неко-
торые его клетки.
Благодаря связям с другими отделами мозга и гипофизом, а так-
же обильному кровоснабжению гипоталамус хорошо приспособлен
как к выявлению отклонений в метаболизме, так и к включению реак-
ций, которые должны выправлять эти отклонения. Другими слова-
ми, он вполне может быть частью системы гомеостаза, и множество
физиологических данных это подтверждает. Например, поддержа-
ние температуры тела — это одна из наиболее тонких систем гомео-
стаза у млекопитающих и птиц. В гипоталамусе имеются участки,
которые очень чувствительны к изменению температуры крови. Если
искусственно подогревать такой участок с помощью введенных в
мозг проволочек, то у животного тут же начинаются одышка и пото-
отделение. Потоотделение вызывается с помощью периферических
реакций вегетативной нервной системы, а гипоталамус эти реакции
запускает. При охлаждении термочувствительных участков возни-
кает противоположный эффект — животное дрожит, и это еще один
пример вегетативной реакции [482].
Описанные выше факты могут показаться чистой физиологией,
мало касающейся исследований поведения. Однако гомеостаз — это
одно из явлений, которые заставляют ломать барьеры между тради-
ционными областями науки. Проводить границу между физиологией
и поведением при контроле за температурой тела не имеет смысла.
Если крысу кратковременно охлаждать описанным выше спосо-
бом, то она начинает дрожать, что способствует выработке некоторо-
го количества тепла. Мы можем рассматривать это как рефлектор-
ную активность и отнести ее к ведению физиологии, как описано
в гл. 1. Однако при более длительном охлаждении одного дрожания
оказывается недостаточно, и крыса для создания теплоизоляции на-
чинает строить себе новое гнездо или расширять уже имеющееся, если
ей дать для этого материал. Рефлекторная реакция теперь поддер-
ж-ивается сложной поведенческой реакцией. Обе они инициируются
гипоталамусом и обе входят в систему гомеостаза крысы, хотя пост-
ройка гнезда включает в себя более сложные нервные механизмы,
нежели дрожание.

166
Глаза 4
Мотивация
167
ГИПОТАЛАМУС И МОТИВАЦИЯ
Исследования роли гипоталамуса помогли в установлении наи-
более важных связей между мозгом и поведением. Современные фи-
зиологические методики позволяют проводить исследование мозга
с помощью электродов, контролируемого введения химических ве-
ществ или разрушения очень маленьких определенных участков
мозга.
Типичный путь, с помощью которого выявляются физиологичес-
кие механизмы мотивации, мы рассмотрим на примере роли гипота-
ламуса в регуляции жажды. В латеральной области гипоталамуса
имеются клетки, которые реагируют на увеличение концентрации
веществ в крови. Они способны запускать две компенсаторные си-
стемы. Первая из систем, действующая через связь гипоталамуса с
гипофизом, вызывает секрецию антидиуретического гормона (АДГ),
усиливающего реабсорбцию воды почками. Вторая система застав-
ляет животное искать воду и пить ее. Если повреждается связь меж-
ду гипоталамусом и задней долей гипофиза, то секреции АДГ не про-
исходит. В этих условиях почки продолжают выводить значитель-
ное количество мочи, и, чтобы компенсировать это, животное пьет
много воды. Это состояние называется несахарный диабет.
В нормальных условиях количество выпитой воды полностью
соответствует потребности животного, однако если произвести элек-
трическое раздражение детекторов жажды латеральной области ги-
поталамуса или ввести в нее гипертонический солевой раствор, то
потребление воды сильно возрастает. Андерсон [9, 10] давал возмож-
ность козам полностью утолить жажду, а затем через инъекционную
иглу вводил им в гипоталамус концентрированный солевой раствор.
Если кончик иглы не попадал в латеральный гипоталамус, это дава-
ло слабый эффект. При попадании соли в эту область козы неистово
пили воду в течение одной или двух минут после инъекции. Эти жи-
вотные, предварительно полностью утолившие жажду, могли пить
соленые и горькие растворы, которых обычно не касались, даже
если очень хотели пить. Данное состояние коз отличается от неса-
харного диабета, поскольку никакой компенсации потери воды, вы-
веденной почками, у них не было. Они пили воду сверх своих физио-
логических потребностей.
Тот же самый эффект Андерсон [11] вызывал при электрическом
раздражении латерального гипоталамуса. Такое питье часто назы-
вают «связанным с раздражением», поскольку оно длится столько
времени, сколько действует раздражитель (электрический или хи-
мический). Аналогичное явление наблюдали у крыс, причем важ-"
ность латерального гипоталамуса для питья получила дальнейшее
подтверждение при изучении поведения крыс, имеющих поврежде-
ние в этой части мозга [450]. Такие животные перестают пить сов-
сем. Они не пьют воду даже на последних стадиях обезвоживания
ь^ерфагииесхие ггрысы
Рис. 4.11. График потребления пищи по дням контрольными крысами и крысами
с двусторонними разрушениями вентромедиального ядра гипоталамуса [525].
Объяснения см. в тексте.
организма и могут погибнуть при наличии воды, если ее не вводить
им искусственно через трубку в желудок. Удивительно, что крысы
с повреждением латерального гипоталамуса не только не интересу-
ются водой, но и испытывают к ней активное отвращение. При по-
мещении такому животному воды в рот оно не глотает ее, а позволя-
ет ей вытечь, делая при этом такие движения губами и языком, ко-
торые характерны для нормальной крысы, пьющей что-нибудь
очень горькое.
Описанные эффекты, возникающие в результате раздражения
или повреждения определенной области мозга, могут привести к
выводу, что латеральный гипоталамус ответствен за состояние, ко-
торое на уровне поведения мы называем стремлением к питью. До
какой степени оправдан такой вывод? Прежде чем обсуждать этот
вопрос, мы остановимся на сходных данных, связанных с участием
гипоталамуса в контроле приема пищи.
Млекопитающие и птицы в норме сохраняют свой вес достаточно
постоянным, регулируя соответствующим образом количество по-
требляемой пищи. Если крысам давать высококалорийную пищу, то
они едят мало, если же корм смешать с непитательной клетчаткой,
то они поедают его в значительно большем количестве. Мы уже упо-
минали о том, что крысы с повреждением центральной области
гипоталамуса, а именно его вентромедиального ядра, теряют способ-
ность к такому чувствительному контролю. На рис. 4.11 сравнивает-
ся потребление пищи крысой с разрушенными вентромедиальными
ядрами и ложнооперированной контрольной крысой. Через нес-
колько дней, по окончании связанного с операцией периода подав-
ленного приема пищи, крыса с разрушением в мозгу начинает пое-
дать гигантские количества пищи, по меньшей мере в четыре раза
большие, чем нормальные животные. Эта так называемая динами-
ческая фаза гиперфагии длится примерно 3 нед. По окончании ее
потребление пищи постепенно снижается и устанавливается на уров-
не, при котором животное съедает примерно вдвое больше нормы.
I •

Мотивация
169
168
Глава 4
!
Само собой разумеется, что гиперфагические животные делаются
невероятно толстыми и очень малоподвижными.
Электрическое раздражение вентромедиального ядра вызывает
у нормальных крыс подавление приема пищи, поэтому данную об-
ласть гипоталамуса часто называют «центром насыщения». Центр
определяет момент, когда животное съело достаточно, и тормозит
дальнейший прием пищи. (Под термином «центр» в данном случае
имеется в виду группа нервных клеток, организованных для выпол-
нения общей функции: мы будем обсуждать центры ниже.) Следует
отметить, что у гиперфагических животных контроль над потреб-
лением пищи потерян не полностью. На рис. 4.11 видно, что иногда
они съедают значительно меньше, чем в ранней динамической фазе.
Разрушение вентромедиального ядра не предупреждает наступле-
ние насыщения, а сильно поднимает его порог.
Центр, комплементарный по функции вентромедиальному ядру,
т. е. усиливающий потребление пищи, расположен в латеральном
гипоталамусе и тесно связан с областью питья. Проведенные на кры-
сах эксперименты по изучению жажды, аналогичные описанным
выше, показали, что «пищевой центр» инициирует поисковое пище-
вое поведение. С помощью вживленных электродов можно вызвать
зависимое от раздражения кормление, а повреждение этой области
приводит к истощению от голода в присутствии пищи. Пищевой и
питьевой центры тесно связаны анатомически и взаимодействуют
друг с другом сложным образом. Для подробного знакомства с по-
ведением при таких взаимодействиях мы отсылаем читателя к очень
хорошему обзору Тейтельбаума и Эпштейна [450].
До какой степени «нормально» действие пищевой и питьевой си-
стем при поведении, зависимом от раздражения, если, как следует
помнить, этот термин относится не к «нормальному», а только к ис-
• кусственному химическому или электрическому раздражению са-
мого мозга. Поскольку не исключено, что еда и питье такого типа
есть просто следствие рефлекторной реакции на раздражение нерв-
ных путей, контролирующих соответствующие комплексы движений,
было бы существенно выяснить, будет ли у животного при раздра-
жении обнаруживаться нормальное поисковое поведение. Раздра-
жением других участков мозга, например областей двигательной
коры больших полушарий, можно вызвать хорошо координирован-
ные движения губ и языка.
Некоторые из наблюдений Андерсона, по-видимому, позволяют
исключить возможность простой рефлекторной реакции, так как
козы при раздражении проявляли все признаки обычной жажды.
Они шли в угол своего загона и искали поилку, т. е. демонстрирова-
ли нормальное поисковое поведение, а не просто насильственно
пили. В случае пищевого поведения Куне и др. [1031 показали, что
крысы, кормящиеся во время раздражения, также способны нау-
читься нажимать на рычаг для добывания пищи, причем эта реакция
I
может переноситься и проявляться у них в состоянии нормального
голода.,
Хотя описанные результаты очень убедительны, по-прежнему
остаются некоторые неясности, на которые недавно обратили внима-
ние Валенстейн и др. [475]. Во многих экспериментах показано, что
тех животных, у которых еда и питье определяются раздражением,
значительно легче отвлечь от цели, нежели животных с нормальной
мотивацией. Это справедливо по крайней мере для крыс: обычно не-
большого добавления в их пищу хинина бывает достаточно, чтобы
они перестали на нее реагировать. (Мы уже упоминали о том, как
легко отвращаются от пищи при добавлении в нее хинина крысы с
гиперфагией.)
Было отмечено, что скорость, с которой крысы пьют воду, лакая
ее из трубки, обычно постоянна. Как правило, лишение животных
воды ведет просто к удлинению периодов лакания, но не влияет на
его скорость. Вместе с тем Уайт и др. [496] сообщили, что при зави-
симом от раздражения питье скорость лакания у животных изме-
няется в зависимости от силы электрического раздражения. Эти дан-
ные заставляют предполагать в данном случае участие центров,
связанных с организацией движений при питье.
Для того чтобы мы могли разрешить этот вопрос, необходимы
дополнительные факты, но даже на основании имеющихся данных
не приходится ожидать, что одни и те же детали организации будут
одинаковыми у разных позвоночных или даже у разных млекопи-
тающих. Тем не менее можно не сомневаться, что гипоталамус
стабильно выполняет свою основную функцию детектора физиологи-
ческого дисбаланса. Его клетки реагируют на температуру, коли-
чество питательных веществ, воду и концентрацию гормонов в кро-
ви. При возникновении любого локального дисбаланса его детектор-
ные области инициируют физиологические и поведенческие реакции,
как мы уже об этом говорили выше.
Хотя несомненно гипоталамус важен для инициирования состо-
яния мотивации, утверждать, что ее контроль зависит только от
гипоталамуса, мы не можем: в управлении мотивацией участвуют
также и многие другие области мозга. Гроссман [183] приводит мно-
го данных, свидетельствующих о влиянии на еду, питье, половое и
агрессивное поведение других участков переднего мозга. Во многих
экспериментах производят повреждения различных областей мозга
и выявляют влияния этих повреждений на поведение. Так, было об-
наружено, что повреждение большей части областей больших полу-
шарий не влияет специфически на мотивацию животного, однако
имеется ряд существенных исключений. Многие из них связаны с
разрушением наиболее фронтальных участков полушарий и группы
проводящих путей, называемых вместе обонятельным мозгом или
лимбической системой. Эта система располагается в основании боль-
ших полушарий и имеет связи с гипоталамусом. Часть ее структур,

170
Глава 4
упрощенно говоря, связана с обонянием, однако у высших животных
она явно участвует в организации более общих аспектов поведения
и, по-видимому, влияет на активность гипоталамических центров.
Было показано, что у собак, кошек и других млекопитающих по-
вреждения лобных долей полушарий и лимбической системы влия-
ют на половое и пищевое стремление, причем иногда исключительно
сильно. Например, собаки с повреждениями части лобных долей на-
чинают непрерывно есть, поедают несъедобные материалы и дела-
ются очень толстыми. Кошки с повреждением области миндалины
(часть лимбической системы) делаются гиперсексуальными и пыта-
ются спариваться с самыми разными животными и неодушевленны-
ми предметами (подробнее см. [182]).
Ряд данных свидетельствует о том, что в регуляции потребления
пищи и воды участвуют проводящие пути переднего мозга, образу-
ющие петлю. Проводящие пути, раздражая которые можно получить
соответствующее поведение, идут из гипоталамуса, проходят через
разные лимбические структуры и прослеживаются снова в направ-
лении гипоталамуса. Конечно, такой круг не изолирован, навеем
его протяжении имеются связи с другими структурами, однако воз-
можно, что поддержание активности внутри подобной петли являет-
ся одной из основ мотивационного состояния.
ОБЗОР ДАННЫХ ПО УПРАВЛЯЮЩИМ ЦЕНТРАМ
В заключение обсудим данные в пользу нейрофизиологической
основы мотивации и рассмотрим, в частности, может ли активность
определенных центров гипоталамуса вызвать специфическую моти-
вацию.
«Центр» — довольно расплывчатое понятие, однако обычно под
этим термином понимают относительно небольшую группу нервных
клеток, имеющих общую функцию. Для исследования организации
мозга применяются различные методики. Одной из наиболее распро-
страненных в физиологической психологии методик является пов-
реждение мозга и исследование влияния этого повреждения на по-
ведение. При выявлении центров методом повреждений опасность
получения ошибочных результатов очень велика, потому что мозг —
это поразительно сложная структура, составленная из мельчайших
единиц. Прижигание его участков или отрезание кусков — это сли-
шком грубые способы, вряд ли приемлемые для работы с ним. При
удалении тел клеток почти всегда повреждаются проводящие пути,
идущие от разрушаемой структуры или проходящие в этом районе.
Это может повлиять на функционирование участков мозга, к кото-
рым идут эти пути, и которые не обязательно находятся вблизи мес-
та разрушения; в результате интерпретация результатов очень зат-
руднена. Например, Рейнолдс [392] предполагает, что вентромеди-
альное ядро гипоталамуса не является «центром насыщения». Он
Мотивация
171
считает, что вследствие разрушения происходит постоянное «раз-
дражение» путей, соединяющих это ядро с «пищевым центром» гипо-
таламуса, и гиперфагия вызывается стимуляцией именно «пищевого
центра».
Чтобы подчеркнуть проблемы, возникающие при интерпретации
результатов, получаемых с помощью методик повреждений, Грего-
ри [178] использует следующую яркую аналогию. Размышляя о
том, как далеко могут заходить и в какой степени полезны для пони-
мания работы мозга сравнения мозга с различными механизмами,
он говорит: «Удаление из электронной схемы нескольких далеко
друг от друга расположенных сопротивлений вызывает в приемнике
вой, однако из этого не следует, что вой непосредственно обуслов-
лен удалением этих сопротивлений или что между ними существует
прямая связь. В частности, мы не должны считать, что в нормальной
схеме функция этих сопротивлений заключается в торможении воя.
В то же время, когда с аналогичной проблемой встречаются нейрофи-
зиологи, они постулируют существование «тормозных областей».
Если мы хотим надежно идентифицировать управляющие цент-
ры, то мы должны данные, полученные с помощью методик повреж-
дений, подтвердить в экспериментах другого типа. Предположим,
нам требуется показать, что определенный отдел мозга отвечает за
включение специфического поведения, связанного с определенной
целью. В идеальном случае мы надеемся использовать методики, пе-
речисленные в табл. 4.1, и получить соответствующие результаты.
В ней дана гипотетическая серия опытов и предполагаемые резуль-
таты чересчур упрощены, однако для «пищевого центра» латераль-
ного гипоталамуса и вентромедиального «центра насыщения» экс-
периментальные данные имеются
Если мы сможем получить такие результаты, то один аспект фун-
кции данного отдела мозга нам удастся идентифицировать доста-
точно надежно, однако никогда не следует полагать, что это все, что
он выполняет. Например, при повреждении вентромедиального ги-
поталамуса изменяются и другие стороны поведения крысы. Живот-
ные делаются менее пугливыми и более подвижными в новой обста-
новке, а также сильнее реагируют на болевые раздражители [184].
Некоторые из этих эффектов характерны для «синдрома гиперфа-
гии», на котором мы останавливались и который нелегко понять,
если ограничиваться представлениями об усилении пищевой моти-
вации. Иными словами, нам нельзя обойтись без тщательного иссле-
дования всех поведенческих эффектов повреждения и раздражения
мозга. Физиология и этология в этом отношении помогают друг
другу.
1
Интересно отметить, что у этих центров обнаруживаются свойства, анало-
гичные свойствам «центральной структуры» и «анализатора» поведенческой моде-
ли Дейча (рис, 4.5),
I ^
I J
lis

128
Глава 4
Таблица 4,1
Методика Ожидаемые результаты
1. Удаление участка мозга
2. Раздражаемая область ( электрически
А
\ химически
области 3. Химическое
мозга
подавление
4. Регистрация нормальной электриче-
ской активности из данной области
у бодрствующего, свободно пере-
мещающегося животного
Определенное поведение не обнаружи-
вается даже при оптимальных для
него условиях
Определенное поведение обнаружива-
ется даже при совсем неподходящих
условиях
Определенное поведение не обнаружи-
вается даже при оптимальных для
него условиях
Активность высока при осуществлении
данного определенного поведения и
низка в его отсутствие
Данных о существовании центров, управляющих другими систе-
мами, помимо пищевого поведения, пока недостаточно. В области
переднего гипоталамуса выделены участки, захватывающие из кро-
ви половые гормоны и контролирующие половое поведение; более
проблематично существование центров агрессии и страха. Напри-
мер, у кошек реакция атаки и «ярости» возникает при раздражении
многих участков гипоталамуса и лимбической системы. В таких слу-
чаях следует быть особенно осторожными в толковании результатов,
поскольку реакция ярости может возникать в ответ на болевые раз-
дражения любого типа, а электроды могут раздражать пути прове-
дения боли. Тем не менее, хотя отдельные элементы реакции
«ярости» кошки — шипение, шерсть дыбом, выгибание спины — мо-
жно вызвать раздражением ряда отделов мозга, для проявления
всего комплекса этой реакции необходима целостность гипоталамуса.
Недавно Валенстейн и сотр. [475] подвергли сомнению специфич-
ность этих предполагаемых центров. Их подход заставляет нас опять
обратиться к обсуждению проблемы общего и специфического по-
буждений, с которой начиналась эта глава. Как обнаружили Вален-
стейн и сотрудники, раздражением одной и той же точки латераль-
ного гипоталамуса можно вызвать разные типы поведения — корм-
ление или питье — в зависимости от того, что происходило с живот-
ным ранее в данной обстановке. Они предположили, что раздраже-
ние вызывает у животного общую активацию и облегчает осуществ-
ление любой реакции, которая, употребляя их термин, была «препо-
тентна» (prepotent). На такую препотентность могут влиять внутрен-
нее состояние организма и внешние раздражители. Эксперименталь-
ные методы, использованные ими, и полученные результаты слиш-
ком сложны, чтобы их излагать здесь. Можно составить хорошее
Мотивация 173
представление о них, если прочесть напечатанную в Science дискус-
сию между Уайзом [509, 510] и Валенстейном [473, 474]. Следует
еще раз подчеркнуть потребность в хороших исследованиях поведе-
ния и необходимость знания того, как именно ведет себя животное в
различных экспериментальных условиях.
Уайз отстаивает существование специфических побуждений, хо-
тя и не отрицает, что центры голода и жажды расположены очень
близко один от другого и могут возбуждаться через один и тот же
электрод. Диффузное распространение тока — это обычный потен-
циальный источник артефактов в исследованиях по раздражению
мозга, и по этой причине Миллер [3491, Гроссман [181] и другие
предпочитают использовать химическую стимуляцию. Центральная
нервная система в биохимическом отношении крайне разнообразна,
и это делает химическую стимуляцию значительно более избиратель-
ной, чем нанесение повреждений или электрическое раздражение.
И электрический заряд, и химическое вещество распространяются
от точки их нанесения и воздействуют на некоторые соседние «цепи»,
однако если на электрическое раздражение реагируют все нейроны,
то каждая цепь реагирует только на некоторые специфические ве-
щества. Это означает, что при последовательном введении несколь-
ких разных веществ каждый раз можно воздействовать на отдельную
«цепь» нейронов, и поэтому результаты таких экспериментов гораз-
до проще интерпретировать. Пользуясь подобной методикой, Грос-
сман смог избирательно усиливать либо еду, либо питье, вводя в ла-
теральный гипоталамус через одну и ту же иглу разные химические
вещества. Поскольку эффекты были очень четкими, маловероятно,
чтобы латеральный гипоталамус был бы столь недифференцирован-
ным, как это полагает Валенстейн.
Какой бы точки зрения в этом споре ни придерживаться, боль-
шинство исследователей согласятся, что нельзя ограничивать цент-
ры побуждения единственной маленькой группой нейронов, по-
скольку в качестве единиц мозговых функций фигурируют большие
по размеру и более диффузные группы нейронов. Центры голода
и жажды наиболее близки к подобному идеалу, однако даже и в
этом случае локализовать их точно невозможно.
Изящные эксперименты Тейтельбаума и Эпштейна [450] показа-
ли, что клетки латерального гипоталамуса обладают значительной
пластичностью. Давая крысам, переставшим совсем принимать пищу
после разрушения этой области, вкусную жидкую пищу, можно до-
биться того, что животные начинают нормально есть. Сначала у них
восстанавливается аппетит, а затем способность регулировать по-
требление пищи. Это означает, что функцию нейронов пищевого
центра взяли на себя другие клетки. Способность к компенсации эф-
фектов повреждения в мозгу обнаруживается достаточно часто, что
также свидетельствует в пользу не слишком жесткой локализации
функций. Этот вывод подтверждается и тем непонятным фактом, о
II

128
Г лава 4
Мотивация
174
i II
И!
!i'
'!
1
;:i i
м i;
t
котором мы упоминали в связи с работой Валенстейна, а именно что
раздражение одного и того же участка в разных случаях не всегда
дает одинаковый результат.
Фон Холст и фон Сент-Пол [230] в своей работе по раздражению
различных участков мозга кур описывают многочисленные подоб-
ные случаи. Нередко они связывали изменения эффектов со спон-
танными изменениями реактивности птицы к определенным раздра-
жителям. Активность одной управляющей системы может влиять на
активность других и изменять порог участка, который раздражает-
ся электрически. Это и определяет те пути, по которым распростра-
няется раздражение.
Нельзя ожидать, что при искусственном раздражении, так же
как и при естественной внешней стимуляции, будет возбуждаться
изолированно только одна мотивационная система. В следующей
главе мы рассмотрим более детально, как в поведении животных
выражается одновременная активация двух разных систем.
ГОРМОНЫ И МОТИВАЦИЯ
Мы только что обсуждали ряд путей, с помощью которых пове-
дение животного можно связать с обменом веществ организма. Ли-
шение животного воды и пищи вызывает поведение, направленное
на восстановление дефицита, и мы отметили, что одновременно с по-
веденческой реакцией происходят гормональные сдвиги. Жажда ве-
дет к выделению антидиуретического гормона задней долей гипофи-
за. Здесь мы подробнее рассмотрим взаимодействие между гормона-
ми и поведением. Эта тема заслуживает пристального внимания, по-
скольку продуцирующие гормон железы внутренней секреции и нер-
вная система имеют общие функции коммуникации и координации
внутри организма, а также между организмом и внешним миром.
Гормоны — это химическая система связи, которая, видимо, так же
стара, как и нервная система. Разумеется, частично одна из них раз-
вилась из другой. У всех представителей животного царства мы об-
наруживаем в нервной системе нейросекреторные клетки. Это мо-
дифицированные нейроны, которые способны как переносить специ-
фические химические вещества по своим аксонам, так и выделять их
в кровоток. Иногда эти клетки собраны вместе наподобие кардиаль-
ных тел у насекомых, которые имеют тесные связи с нервной систе-
мой и с кровеносным руслом. Гипофиз позвоночных развивается в
результате слияния нервной и эпителиальной тканей и сохраняет
прочную связь с гипоталамусом. Выделяя разные гормоны, гипофиз
регулирует активность всех остальных желез внутренней секреции
и в свою очередь регулируется центральной нервной системой. По-
добная система управления позволяет обеспечивать соответствую-
щую гормональную реакцию на изменение среды, которое воспри-
нимается нервной системой. Наиболее известный пример этого —
регуляция размножения у млекопитающих или птиц в определенное
время года. Изменение длительности дня, воспринимаемое зрением,
вызывает изменения в активности гипоталамуса, который стимули-
рует гипофиз. Последний в свою очередь секретирует гормоны, ко-
торые вызывают в организме изменения, связанные с началом раз-
множения.
При всех взаимосвязях функции двух коммуникативных систем —
эндокринной и нервной — остаются в значительной степени комп-
лементарными. Нервная система передает информацию только с по-
мощью залпов импульсов. Ее состояние может очень быстро менять-
ся, и она мало подходит для передачи постоянного и неизменного
сигнала в течение длительного периода времени. Временной масш-
таб функционирования нервной системы характеризуется малыми
величинами — от миллисекунд до нескольких минут. Эндокринная
система не способна реагировать столь быстро, однако ее клетки мо-
гут длительно и стабильно поддерживать секрецию гормона, если
это необходимо,— в течение месяцев. Кроме того, разносясь с кро-
вью, гормоны могут попадать в любую клетку тела, тогда как нерв-
ная система управляет главным образом мышцами.
Циркулирующие в крови гормоны обычно рассматриваются в по-
ведении животных как первичные мотивирующие факторы; этологи
часто предполагают, что они непосредственно воздействуют на соот-
ветствующие центры мозга, усиливая побуждение. И действитель-
но, имеются убедительные примеры того, как гормоны явно вынуж-
дают животное совершать определенные акты поведения даже в са-
мых неблагоприятных обстоятельствах. Например, Блум и Фидлер
[58] показали, что после инъекции гипофизарного гормона пролак-
тина (см. ниже) содержавшиеся в изоляции самцы рыбы Cretiilab-
rus ocellatus производят вентилирующие движения плавников, ха-
рактерные для родительского поведения. По форме и по функции
они сходны с вентилирующими движениями у колюшек, которые
описаны на с. 152. После инъекции самцы Crenilabrus вентилируют
воду в пустом аквариуме, в котором отсутствуют нормальные внеш-
ние раздражители, вызывающие вентилирование: С02 в воде, опло-
дотворенная икра, гнездо, место, где оно находится, и т. д., а коли-
чество времени, проведенное за вентилированием, зависит от дозы
пролактина. Примеры такого типа убедительно свидетельствуют о
центральной мотивирующей роли гормонов, но гормоны влияют на
поведение и другими способами.
Прежде чем обсуждать примеры действия гормонов, необходимо
дать краткое описание тех аспектов эндокринной системы позво-
ночных, которые наиболее важны для поведения. Подробные обзоры
на эту тему написаны Горбмэном и Берном [170], а также Остином
и Шортом [18].

128
Г лава 4
Мотивация
90
ГИПОФИЗ
Гипофиз выделяет несколько гормонов, которые воздействуют
на функционирование других эндокринных органов. Таким путем
гипофиз эффективно управляет всей эндокринной системой-. Боль-
ше всего нас интересуют гонадотропины, которые действуют на го-
нады и вызывают как рост зародышевых клеток, так и развитие тка-
ней половых желез, в свою очередь тоже секретирующих гормоны.
Существуют два главных гонадотропина: фолликулостимулирую-
щий гормон (ФСГ) и лютеинизирующий гормон (ЛГ). Оба они полу-
чили свои названия по их действию на женские гонады, или яич-
ники, однако они секретируются и у самцов и имеют у них сходные
функции. У самок ФСГ и ЛГ необходимы для роста яйцеклеток и
их выхода в яйцевод.
Третий, важный для поведения гормон гипофиза — пролактин,
называемый также лактогенным гормоном или лютеотропным
гормоном СЛТГ) дает множество физиологических эффектов. Из-
вестно, что он имеется у позвоночных разных классов, однако у него
часто бывают совершенно различные функции и органы-мишени
(те части организма, рост и функция которых изменяются под влия-
нием гормона). Интересно отметить, что большая часть его эффектов
относится к родительскому поведению в самом широком смысле.
Как уже упоминалось, у самцов колюшек и некоторых других
рыб пролактин стимулирует вентилирование икры. Он способству-
ет насиживанию у кур (но не у всех птиц), секреции молочка у го-
лубей, а также росту молочных желез и секреции молока у млеко-
питающих.
Название лютеотропный гормон происходит от еще одной его
важной функции у млекопитающих — он необходим для поддержа-
ния желтого тела яичника и стимуляции выделения им прогестерона
(см. ниже).
ГОНАДЫ: ЯИЧНИКИ И СЕМЕННИКИ
Уже много столетий люди знают, что кастрация глубоко влияет
на поведение и на особенности строения тела позвоночных. В от-
личие от этого у беспозвоночных такого влияния почти или совсем
нет. Гонады выполняют роль важных эндокринных желез только у
позвоночных, у которых при стимулирующем влиянии гипофизар-
ных ФСГ и ЛГ в специальных секреторных клетках продуци-
руются половые гормоны. Женские гормоны называются эстро-
генами, а мужские — андрогенами. Все эти гормоны относятся к
стероидам и имеют сходную химическую структуру. Хотя у разных-
групп позвоночных секретируются несколько различные стероиды,
гормоны одной группы животных обычно эффективны для других.
Наиболее распространенным андрогеном млекопитающих явля-
ется тестостерон.
Половые гормоны ответственны за развитие вторичных половых
признаков, а также за рост репродуктивной системы и образование
в ней яйцеклеток и спермы. Обычно в строении тела самцов и самок
имеются постоянные различия, которые поддерживаются в тече-
ние всей жизни взрослой особи. Усиление этих различий при дей-
ствии половых гормонов обычно происходит в сезон размножения.
У оленей самцов можно отличать от самок на протяжении всего го-
да, однако сезонный рост рогов происходит только в период размно-
жения.
Наконец, нужно упомянуть еще один стероидный гормон, кото-
рый продуцируется яичником. После выхода яйцеклетки ее пустой
фолликул увеличивается и образует заметную желтоватую структу-
ру на поверхности яичника — желтое тело. Оно начинает секрети-
ровать прогестерон, под влиянием которого слизистая матки подго-
тавливается для принятия яйцеклетки и развития ее в бластоцисту.
Прогестерон тормозит также сокращение мышц матки, что необхо-
димо для продолжения беременности. Его справедливо называют
гормоном беременности, однако встречается он не только у млеко-
питающих. Образования, сходные с желтым телом, обнаружива-
ются при созревании икринок у рыб, амфибий и яиц у пресмыкаю-
щихся; правда, о наличии и действии прогестерона у этих живот-
ных мы знаем очень мало. У птиц имеется прогестерон, который поч-
ти наверняка выделяется яичниками, хотя желтые тела у них не за-
метны. Известно, что прогестерон продуцируется у самцов птиц не-
которых видов (видимо, в семенниках).
ВЛИЯНИЕ ГОРМОНОВ НА ПОВЕДЕНИЕ
Как упоминалось выше, исследователи поведения обычно обра-
щали внимание на центральное действие гормонов. Однако для
этих очень активных химических соединений характерно то, что
они часто вызывают бурный рост в разных органах-мишенях. Про-
изводимые ими изменения могут косвенно влиять на поведение
таким образом, что порой это трудно заметить.
Например, у самок млекопитающих эстроген вызывает рост мат-
ки, изменения в эпителии влагалища и рост протоков молочных же-
лез. У птиц эстроген, иногда действуя вместе с прогестероном, вы-
зывает увеличение яйцеводов и всей репродуктивной системы.
У некоторых видов он вызывает также выпадение перьев на вент-
ральной поверхности тела и усиленную васкуляризацию этого
участка-(т. е. образование наседного пятна). Под влиянием пролак-
тина у млекопитающих при беременности увеличиваются молочные
железы, а у голубей в последние дни инкубации набухает зоб.
Все органы-мишени снабжены нервами, и информация о проис-
ходящих в них изменениях может участвовать в создании обратной
связи с нервной системой и влиять на поведение. Лерман [287]
J