Меннинг О. Поведение животных. Вводный курс
Подождите немного. Документ загружается.


218
Глава 6
Эволюция
238
Рис. 6.6. Три взрослые флоридские хохлатые сойки, ухаживающие за выводком
[507]. Они были окольцованы, поэтому известно, что это двое родителей и птенец
предыдущего выводка, который помогает в выращивании сибсов. Подробнее см.
текст.
ных животных, поэтому, для того чтобы отбор родичей мог действо-
вать, необходим какой-то способ идентификации родственников.
Для общественных насекомых, а также для животных, подолгу жи-
вущих в малых группах, например для приматов, его найти легко
(гл. 8). Значительно труднее идентифицировать родственников у жи-
вотных, которые после размножения расстаются и больше друг с дру-
гом не соприкасаются.
В этом отношении у птиц можно провести интересные сопостав-
ления. Многие тропические и субтропические, птицы ведут относи-
тельно оседлый образ жизни, и потомство их далеко не улетает.
Напротив, молодые птицы зоны умеренного климата разлетаются,
и все они мигрируют на зиму в более теплые районы. В связи с этим
интересно отметить, что явление, иллюстрируемое рис. 6,6, распро-
странено именно среди тропических птиц. На рисунке показаны
три взрослые птицы, участвующие в выращивании птенцов. Однако
не все они являются настоящими родителями. Иногда можно уста-
новить, что дополнительные «помощники» — это молодые птицы
предыдущего выводка, но часто,это бывают просто соседи, у которых
было разрушено гнездо (у многих тропических видов смертность
птенцов в гнездах очень высока). У птиц умеренной зоны таких по-
мощников практически никогда не наблюдается.
Поведение помощников в понятиях отбора родичей объяснить
довольно легко. Молодые птицы помогают родителям выращивать
большее число сибсов, с которыми они связаны таким же близким
родством, как и со своими детьми. Очень часто молодые птицы раз-
множаются плохо, и возможно, что остаться около родительского
гнезда и набрать опыта в сборе корма и т. д. при выращивании второ-
го выводка для них очень благоприятно. Насколько мы знаем, у та-
ких видов дисперсия особей по территории невелика, и вполне веро-
ятно, что рядом гнездящиеся птицы имеют общие гены. Если у одной
из птиц погибло собственное гнездо и уже слишком поздно начать
в этом сезоне строить новое, лучше вместо этого помочь родствен-
никам-соседям вырастить их птенцов. Ни один из этих доводов не
применим к птицам умеренной зоны, которые распределяются по тер-
ритории более широко и часто выращивают лишь один выводок.
Подобные объяснения легко приводить, но их еще требуется дока-
зать, и они далеко не бесспорны. Захави [518] выступил против идеи
об отборе родичей. Он утверждает, что у арабских кустарниц, кото-
рых он изучал, помощники приносят гнездам своих родителей не
столько пользы, сколько вреда, создавая беспорядок и привлекая
хищников. Очевидно, для доказательства гипотезы отбора родичей
важно показать, что гнезда с помощниками оказываются в лучшем
положении в отношении выживания потомства, чем гнезда без них.
Однако в настоящее время достоверных данных такого рода очень
мало. Подробное обсуждение этого вопроса с множеством примеров
дано в книге Брауна [73].
Можно возразить, что, поскольку в случае помощников самопо-
жертвования почти не требуется, это не достаточно убедительный
пример отбора родичей. Есть несколько других примеров, в которых
самопожертвование оказывается более очевидным. Один из наиболее
поразительных — это половое поведение диких индюков, описанное
Уаттсом и Стоксом [489]. Самцы одного выводка индюков по дости-
жении взрослого возраста остаются в группах по два или три и одно-
временно ухаживают за самками на общих токовых площадках.
С самками, которых эти демонстрации привлекают, спаривается
один, наиболее доминантный из всей группы братьев самец (гл. 8).
Другие ему уступают и остаются фактически стерильными. Как это
бывает у многих птиц, для которых характерны демонстрации на
общем токовище (описание его у тетерева см. в гл. 8), привлекает
самок и спаривается с ними только очень небольшое число самцов-
индюков. Отбор родичей благоприятствует менее доминантным сам-
цам, поддерживающим своего брата и помогающим ему добиться
успеха в спаривании путем участия в совместных демонстрациях.
Таким путем они значительно увеличивают совокупную приспособ-
ленность.
Отбор родичей — это полезная концепция, которая расширяет
наши представления об эволюции поведения. Возможно, ее главный
недостаток заключается в относительной легкости построения разно-
го рода недоступных проверке гипотез, которые объясняют малопо-
нятные типы общественных взаимодействий. Далее, как мы уже виде-

218
Глава 6
Эволюция
240
ли в случае помощников, не всегда возможно отделить благоприят-
ные моменты для данной особи от подобных моментов для ее родичей.
Однако если соблюдать определенную осторожность, то на основе
представления об отборе родичей можно строить доступные проверке
гипотезы, и, как мы увидим в гл. 8, это представление играет важную
роль в современном развитии науки об общественном поведении.
МИКРОЭВОЛЮЦИЯ ПОВЕДЕНИЯ
Термин «микроэволюция» используется для описания наиболее
ранних этапов дивергенции между популяциями, приводящей к об-
разованию собственно видов. Изменения в поведении, обусловлен-
ные микроэволюцией группы, относительно легко связать с генети-
ческими изменениями, о которых мы только что говорили.
При сравнении поведения животных близких видов, как правило,
можно обнаружить у них сходные комплексы фиксированных дей-
ствий. Эти комплексы истинно гомологичны, точно так, как гомоло-
гичны элементы скелета разных групп позвоночных — все они вос-
ходят к общему предку. Однако сами эти комплексы под действием
отбора изменяются по-разному. Наиболее часто эти изменения затра-
гивают, во-первых, частоту выполнения комплексов действий и,
во-вторых, их форму, или «акцентирование». Первая категория
изменений не требует комментариев, по своему характеру эти изме-
нения сопоставимы с различиями между линиями, о которых мы
говорили. Из множества примеров приведем три. У разных видов
уток набор всевозможных демонстраций сходен, однако отдельные
комплексы выполняются ими с разной частотой. Некоторые из них
совсем не выполняют определенных действий, хотя имеют соответ-
ствующий генетический материал.
Два вида рода Drosophila: D. melanogaster и D. simulans харак-
теризуются подобными различиями У первого вида наиболее обыч-
ная демонстрация — это вибрация, а у второго — ножницеобразные
синхронизированные движения обоих крыльев [323]. Кларк и др.
[95] описали различия в частоте выполнения демонстраций ухажива-
ния у двух видов тропических пресноводных рыб-меченосцев.
Столь же распространены микроэволюционные изменения фор-
мы комплексов действий. При этом в общих элементах демонстра-
ций происходит «акцентирование» разного характера. Проиллю-
стрируем данное положение двумя примерами [325], один из кото-
рых поясняет рис. 6.7. «Долгий крик» чаек эквивалентен территори-
альному пению воробьиных птиц. Птицы двух видов, показанных
на рисунке, осуществляют одинаковую последовательность движе-
ний головы при крике, но «акцентируют» разные части этой после-
довательности, что придает каждой из демонстраций характерный
вид Крэйн 1106] провел тщательное сравнительное исследование
демонстрации ухаживания у манящего краба Пса. Самцы роют на
Обыннавеннау
Серебристая v/«
;
/,,;v
Рис. 6.7. Вариации «акцентирования» при «долгом крике» у двух видов чаек [456].
Последовательность движений слева направо. Сначала голова находится в наклон-
ном положении (Л), при начале крика происходит опускание ее вниз (Б), а за-
тем она отбрасывается назад (В). По мере затухания крика (Г) голова постепенно
опускается. У обыкновенной чайки не акцентируется фаза Б и акцентируется фаза
В, а у серебристой чайки картина обратная.
берегу норки и становятся при входе Одна их клешня значительно
больше другой и ярко окрашена. При виде самки они ритмически
машут этой клешней, и если самка готова к размножению, она при-
ближается. Непосредственный характер подманивающего движения
у каждого вида свой. Род можно подразделить на два подрода как по
поведенческим, так и по морфологическим критериям. К одному из
них, который Крэйн назвал подродом с «вертикальным подманива-
нием», принадлежат виды, самцы которых только немного отставля-
ют клешню и просто поднимают ее вверх и вперед. Представители
подрода с «латеральным подманиванием» отставляют клешню вбок
и опускают вниз, совершая ею более или менее круговой путь.
Для удобства мы выделили два типа изменений — частоту вы-
полнения и форму комплекса действий, однако совершенно очевидно,
что в норме они происходят одновременно и комбинируются с дру-
гими типами изменений. Например, у сверчков при развитии разных
типов песен от общего предкового типа происходит большое число
изменений. При обсуждении этого вопроса Александер [3] приводит
большое число примеров. Песни сверчков варьируют по амплитуде,
Длительности и частоте импульсов. Благодаря этому множество тре-
лей и стрекотаний, присущих сверчкам, сильно отличаются друг от
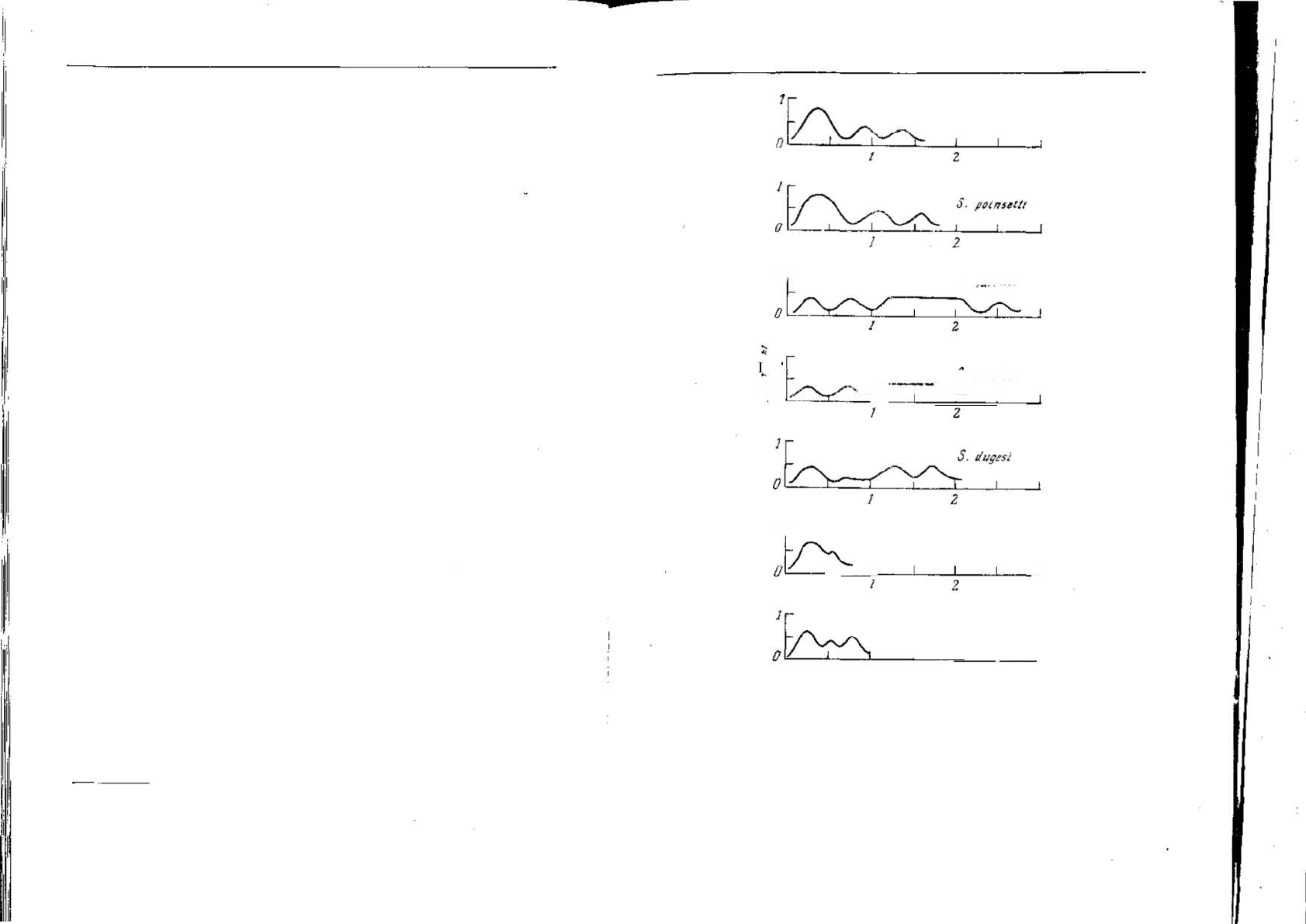
242
Глава 6
друга. Такого же рода факторы должны участвовать в эволюции
пения птиц. Певчий дрозд, Tardus ericetorum, характерным образом
дважды или трижды повторяет каждую фразу песни, прежде чем
переходит к следующей
1
, а его близкий родственник черный дрозд,
Т. merula, этого не делает.
ПРИРОДА ГЕНЕТИЧЕСКИХ И МИКРОЭВОЛЮЦИОННЫХ ИЗМЕНЕНИИ
Было бы интересно выяснить, не существует ли общей связую-
щей нити между описанными выше разнообразными генетическими
и микроэволюционными изменениями. Мы подчеркивали, что подоб-
ные изменения имеют количественный характер и многие из них,
выражаясь на языке поведения, непосредственно связаны с измене-
ниями в порогах реакций. Если в одной и той же обстановке два
вида или две линии различаются относительными частотами, с кото-
рыми они осуществляют комплексы действий Л и £ из их общего
набора комплексов, то вполне разумно предположить, что это свя-
зано с различиями в порогах механизмов, управляющих комплек-
сами Л и Б в соответствующих двух видах или линиях. На эти ме-
ханизмы можно воздействовать в различных «участках». Это могут
быть изменения в пороге сенсорной системы, в системах, влияющих
на мотивацию, в двигательной сфере или же любая комбинация
подобных изменений. Изменения порогов происходят как при дей-
ствии генов на нервную систему непосредственно, как у гиперкине-
тических мутантов дрозофилы, описанных на с. 226, так и менее пря-
мым путем — с помощью генов, воздействующих на обмен веществ
или секрецию гормонов, которые в свою очередь влияют на нервную
систему. О достаточно сильном прямом действии генов на нервную
систему имеются лишь отдельные косвенные свидетельства, тогда
как прямых доказательств этому нет. Так, например, различия в
частоте ряда половых реакций у морских свинок разных линий сох-
раняются даже после инъекции им половых гормонов [1851 и тирок-
сина, которые повышают уровень основного обмена [395]. По-види-
мому, различия между линиями обусловлены различиями в порогах
мозговых механизмов полового поведения.
Качественные изменения формы или скорости выполнения комп-
лексов действий также легко объяснить изменениями порогов. Каж-
дый комплекс фиксированных действий должен иметь своего рода «ко-
ординационный центр», структура и свойства которого наследуются.
При активации этого центра выполняются стереотипные действия
на выходе к нижележащим центрам, контролирующим мышцы или
1
Дрозд умный дважды песнь свою поет,
Чтоб знали все: прожить он может снова
Миг вдохновенья вольного шального.
Р- Браунинг (Перевод А. В. Парина)
Эволюция
243
S. cyanegenys
1
г
S. jar.-ovi
1
J
-С
О. or.па t us
"У- .
/г
J— L
5. lorquatus
J
S. mucronatus
n J II
1 2
Время, с
Рис. 6.8. Специфические движения головой (покачивание) у некоторых ящериц
рода Sceloporus [240]. По оси ординат отложена величина отклонения головы, а по
оси абсцисс — время. Четко заметны изменения по амплитуде, скорости и дли-
тельности движений.
группы мышц. Координационный центр возбуждает каждую мы-
шечную группу в определенном порядке в нужное время и на опре-
деленный период. Его влияние зависит от серии тонких взаимодей-
ствий порогов как внутри самого координационного центра, так и
о

218
Глава 6
Эволюция
244
между центрами управления мышцами. Выход координационного
центра может быть полностью «преформирован» или же он может
претерпевать изменения под действием обратной связи от мышц по
мере выполнения комплекса фиксированных действий. У насеко-
мых нам известны центры (видимо, небольшие группы нейронов),
которые контролируют, например, пение у сверчков [238, 239] или
полет у саранчи [504]. Даже после изоляции они генерируют вполне
нормальные выходные сигналы, но теперь эти сигналы уже не могут
изменяться под влиянием обратной связи. Хойл [235] подробно
рассматривает разные типы управления в связи с результатами его
собственной работы на насекомых, которые оказались превосход-
ным объектом для такого рода исследований (см. также гл. 2).
Генетические изменения, влияющие на пороги как внутри коор-
динационного центра, так и в одной из подчиненных мышечных
групп, обусловливают изменения формы комплекса фиксированных
действий. Например, мышцы могут начать сокращаться раньше
или позднее, возбуждаться на большее или меньшее время, иметь
большее или меньшее число активированных мышечных единиц,
что влияет на интенсивность или частоту реакции. На рис. 6.8 дан
прекрасный пример влияния подобных изменений на поведение
ящериц, взятый из работы Хансекера [2401. Самцам рода Sceloporus
свойственны ритмические покачивания головой во время ухажива-
ния, а также при встрече другой особи своего вида, когда они играют
роль сигнала видовой принадлежности. Для каждого вида характер
такого покачивания специфичен. Оно возникает в результате сокра-
щения мышц, вытягивающих передние конечности и поднимающих
и опускающих голову и плечи. В процессе микроэволюции возникло
несколько четко различающихся типов таких движений, связанных
с изменениями, подобными описанным выше. Этот пример особенно
нагляден, поскольку в нем речь идет о работе всего двух групп мышц,
однако нетрудно себе представить сходное возникновение различий
в «акцентрировании», показанных на рис. 6.7.
Итак, гены, влияющие на пороги в нервной системе, играют в
эволюции поведения важную роль. В основном наблюдаемое нами
поразительное многообразие комплексов фиксированных действий
возникло в результате накопления малых количественных измене-
ний, точно так же, как произошло развитие формы тела животных.
РИТУАЛИЗАЦИЯ
Многие из комплексов фиксированных действий, эволюцию кото-
рых мы рассматривали, функционируют как социальные сигналы
и могут быть компонентами демонстраций угрозы, ухаживания или
умиротворения. В гл. 5 мы уже обсуждали происхождение таких
комплексов действий. Обычно они не возникают de novo, а раз-
виваются из «начальных» (или «интенционных») движений — осо-
I
Рис. 6.9. Вытирание клюва в демонстрации ухаживания у трех видов вьюрков
[357].
А. Зебровая амадина, у которой движение неритуализовано. Справа показан са-
мец, который только что сделал такое движение. Б к В. Lonchura striata и L. ри-
nctulata, у которых, по всей видимости, движение стало ритуализованным. У этих
видов самец сохраняет такую позу в течение нескольких секунд, и его ритуали-
зованное вытирание клюва больше похоже на поклон. Во всех ipex случаях слева
самки.
бенно движений нападения и бегства — или же из смещенной
активности, где наиболее часты движения дыхания, чистки перь-
ев, питья или потребления пищи. Такие демонстрации Тинбер-
ген [461] назвал производной (derived) активностью, отмечая тем
самым, что они происходят от поведения других типов. Эволюцион-
ный процесс, при котором происходит их превращение в социальные
сигналы, называется «ритуализацией». Этот термин предложил Хак-
сли [245] для описания сложной взаимной демонстрации у чомги,

246
Глава 6
о которой мы уже говорили (рис. 5.10). Эта демонстрация развилась
из элементов гнездостроительного поведения этих птиц, причем
в ряде случаев демонстрации ухаживания в процессе эволюции изме-
нились настолько сильно, что без сравнительных данных по видам,
менее изменившимся в этом плане, нам было бы трудно представить
их происхождение. Особенно это относится к некоторым движениям
ухаживания тропических вьюрков-амадин (Erythrurae), показанным
на рис. 6.9. Самец зебровой амадины во время ухаживания часто
производит движения чистки клюва о веточку; возможно, что это
неритуализованная смещенная активность. У родственных зебровым
амадинам Lonchura striata и L. punctulata самец производит поклон и
остается в такой позе около самки с головой, склоненной к ветке.
Сходство этой позы с позой чистки клюва поразительно, и Моррис
[3571 предполагает, что поклон — это сильно ритуализованный
комплекс действий, возникший из движений чистки клюва.
При ритуализации происходят все те изменения порогов и ча-
стот, о которых мы только что говорили, и особенно изменения в
форме движений и-их «акцентировании». Социальные сигналы долж-
ны быть заметными; большинство их искажено по форме и сопровож-
дается специально развившимися релизерами, усиливающими эф-
фект самого движения (см. примеры на рис. 5.3 и 5.11). Ритуализа-
ция начальных движений хорошо описана в обзорах Даньжи [116]
и Эндрью [131. Тинберген [461] и Моррис [355] рассматривают также
другие типы производных активностей.
Нам осталось упомянуть только о двух моментах, связанных с ри-
туализацией. Во-первых, о концепции типической интенсивности —
понятии, введенном Моррисом [356]. Для максимальной эффектив-
ности социальные сигналы должны быть четкими и однозначными.
По этой причине многие из них не изменяются по форме движений
при изменении интенсивности раздражителей. У самца Amadina
fasciata в демонстрации ухаживания есть характерная «распушен-
ная поза». При действии слабых раздражителей он принимает ее на
короткое время, а при действии сильных раздражителей он долго
пребывает в ней и начинает танец ухаживания. В каждом случае
поза при этой демонстрации одинакова (она приведена на рис. 6.10),
т. е. для нее характерна типическая интенсивность. Это можно срав-
нить с позой угрозы у кошки (рис. 5.8), у которой интенсивность
реакции соответствует силе раздражителя. Позы и движения, имею-
щие типическую интенсивность, легко распознаются, однако они
дают мало информации о мотивационном состоянии сигнализирую-
щего животного. Оба способа сигнализации должны иметь свои пре-
имущества и то, какой из них развивается в процессе эволюции, за-
висит от конкретной обстановки, в которой выполняется данная,
демонстрация.
Вторая концепция, о которой нам следует здесь упомянуть,—
это эмансипация (освобождение) ритуализованных комплексов дей-
Эволюиия
247
сдстав-
Рис. 6.10. Распушенная поза ухаживания v самца Z tawinin R
A
fii
«ass п ss g s s
ствий. Например, ухаживание у уток включает движения котооые
явно происходят от смещенной чистки перьев (рис 5.11) и смещенно
го питья. Следуя доводам, которые мы приводили в гл. 5 можно
предположить, что эти комплексы возникли как смещенная а™
0 ЬттСЛеДСТВИе пас^лмажив i
ст о;
м
Л
д
Р °Р ания при конфликте между половьш
стремлением и стремлением к нападению или бегству, когда самцы
ухаживают за самками. Как предполагают, появление чистки перьев
ли питья у предковых форм означало, что конфликтующие ст
л
емле _
У Самцов
ГОДЯТСЯ в равновесии. Самцы! которые проявляли

248
Глава 6
Эволюция
249
смещенную активность более регулярно, были более успешными в
спаривании. Каким-то образом это проявление приковывало внима-
ние самки и, возможно, возбуждало ее половое поведение. У их по-
томков конфликт в ситуации полового поведения уже не был необ-
ходимой предпосылкой выполнения движений чистки перьев и питья.
Они стали практически постоянными частями ритуала ухаживания
и сильно изменились по форме. Тинберген [461] предполагает, что
произошла эмансипация этих движений от исходного управляющего
механизма, и теперь они находятся только под контролем механиз-
мов полового поведения.
Два главных критерия, по которым можно судить об эмансипа-
ции комплекса действий, состоят в следующем. Во-первых, комплекс
выполняется в условиях, отличных от условий, необходимых для
выполнения исходного комплекса, т. е. он более не вызывается ранее
эффективными раздражителями. И, во-вторых, флуктуации в стрем-
лении к выполнению предполагаемого исходного комплекса дей-
ствий не влияют на стремление к выполнению эмансипированного
комплекса. Первый критерий сходен с тем, который используется
для определения комплексов действий как смещенной активности,
тогда как на основании второго критерия можно предсказать, что,
например, состояние жажды не должно влиять на количество де-
монстраций питья, осуществляемых селезнем во время ухаживания.
У нас мало точных доказательств выполнимости этих критериев,
однако реальность существования эмансипации несомненна. Срав-
нительные исследования ясно показывают происхождение элементов
демонстраций ухаживания от движений питья, однако сам комплекс
действий сильно видоизменился. У некоторых видов это не более
чем качание головой, и оно явно не имеет ничего общего с потребле-
нием воды. В настоящее время мы ничего не знаем о том, как проис-
ходил процесс эмансипации (обзор по эмансипации демонстраций уг-
розы написан Блэртон-Джонсом [60]).
Как мы упоминали уже в гл. 5, при обсуждении вопросов, касаю-
щихся смещенной активности, для того чтобы смещенная активность
«прорвалась», необходимо достаточно стабильное состояние равнове-
сия между конфликтующими стремлениями. Такое равновесие не мо-
жет сохраняться идеальным достаточно долго, и, видимо, существу-
ет ряд состояний конфликта, при которых оба стремления достаточно
близки по величине, чтобы на короткое время могло проявиться сов-
сем иное поведение. Роуэлл [402] называет такой ряд рядом «эффек-
тивного равенства». Не исключено, что один из начальных этапов
ритуализации заключался в расширении ряда эффективного равен-
ства между находящимися в конфликте стремлениями и, следова-
тельно, в увеличении частоты проявления смещенной активности.
Блеет [57] подробно обсуждает некоторые вопросы эмансипации и
концепцию ритуализации в целом.
До сих пор при обсуждении ритуализации мы касались исключи-
тельно вопросов эволюции демонстраций ухаживания и угрозы,
в отношении которых этот термин первоначально применялся. Сле-
дует отметить, что и в других случаях можно обнаружить такую
тенденцию в действии отбора, когда поведенческий элемент, прояв-
ляющийся в одних ситуациях, служит исходным материалом для ри-
туализации при других обстоятельствах.
Многие тропические пресноводные виды рыб, такие, как гурами
и бойцовые рыбки, строят гнезда из воздушных пузырьков. Самец
набирает на поверхности воды воздух и выдувает пузырьки с обо-
лочкой из слюны, которые он располагает среди водных растений в
виде гроздьев, прилегающих к поверхностной пленке. После ухажи-
вания и спаривания в это гнездо из пузырьков он помещает оплодо-
творенную икру. Брэдок и Брэдок [64] предположили, что сооруже-
ние пузырьковых гнезд последовало за эволюцией воздушного дыха-
ния. Последний навык эволюционировал в связи с совершенно дру-
гими потребностями — как реакция на низкое содержание кислоро-
да в воде тропических рек и озер, когда рыбы постоянно набирали
воздух у поверхности воды. При таком поведении рыба обычно вы-
пускает несколько пузырьков, которые, не лопаясь, задерживаются
у поверхностной пленки. Имея это в виду, нетрудно представить,
каким образом пузырьковые гнезда постепенно превращались в
хорошо снабженные кислородом укрытия для развивающихся икри-
нок.
Другим замечательным примером эволюции с использованием
случайных удобных обстоятельств представляется язык танца медо-
носной пчелы, который мы подробно обсуждали в гл. 3. В основе
этой системы коммуникации пчелы лежит ее способность переводить
угол своего пути относительно Солнца в угол отклонения от направ-
ления силы тяжести при ее перемещении на вертикальной стенке
сота. Эта способность кажется на первый взгляд весьма необычной,
и ее хочется объяснить каким-то уникальным эволюционным событи-
ем. Однако мы знаем, что свет и сила тяжести используются для
. ориентации большинством насекомых и, следовательно, в их взаимо-
связи нет ничего необычного. Например, муравьи, которые бегут
по горизонтальной поверхности под определенным углом к свету,
при выключении света и превращении поверхности в вертикальную
резко меняют курс, принимая такой же угол относительно направ-
ления силы тяжести [478]. Некоторые жуки ведут себя сходным обра-
зом. Возможно, что у пчел произошла модификация системы ориен-
тации, свойственной всем насекомым. Насколько нам известно, ника-
кие другие насекомые не используют эту способность для коммуни-
кации, однако потенциально это вполне возможно. Естественному
отбору, столкнувшемуся с требованием максимальной эффективно-
сти в обеспечении кормом колоний медоносных пчел, удалось создать
такое замечательное средство коммуникации на основе столь обыден-
ного и малозаметного свойства.

250
Глава 6
Эволюция
251
!
I
. I
, i.
ПОЛОВАЯ ИЗОЛЯЦИЯ
Родственные виды в естественных условиях обычно не скрещива-
ются между собой, поскольку это для них неблагоприятно. Хотя
у гибридов могут обнаруживаться некоторые признаки «гибридной
силы», они обычно бывают полу- или полностью стерильными. Даже
если гибриды способны к размножению, они редко размножаются
так успешно, как любой из родительских видов. У последних имеет-
ся набор генов, который в течение многих поколений отбирался как
наилучший для данных условий среды. Гибриды имеют промежуточ-
ный набор генов, не обеспечивающий им столь хорошего приспособ-
ления к условиям жизни, как в случае одного или другого родителя.
У растений известны случаи, когда межвидовые гибриды с успехом
расселились в новых условиях, однако у животных, если это и слу-
чается, то крайне редко.
Следовательно, выбор полового партнера из членов своего вида
представляется благоприятным для вида. Это особенно важно для
самок; самцы обычно спариваются в течение жизни несколько раз,
тогда как, например у насекомых, самки спариваются только од-
нажды. Если они спарятся с самцами не своего вида, то могут остать-
ся бесплодными. В соответствии с этим, как правило, способность
к различению других особей более сильна у самок, чем у самцов.
Самки не могут позволить себе рисковать, тогда как самцы могут,
и очень сильная способность к различению может быть для них даже
неблагоприятной. Для самца бывает лучше рискнуть тем, что какое-
то спаривание не даст результата, чем упустить шанс осуществить
такое спаривание, которое обеспечит появление потомства. У дрозо-
фил, например, самки более разборчивы, чем самцы, и они довольно
часто отвергают посторонних поклонников, что препятствует меж-
видовой гибридизации.
Половую изоляцию можно определить как «поведенческую
преграду гибридизации между видами или популяциями». Она ока-
зывается одним из проявлений более общего явления — репродук-
тивной изоляции, подробно обсуждаемой Майром [3391, которая
представляет собой один из главных путей воздействия поведения
на эволюцию популяций животных.
Степень контакта двух родственных видов в естественных услови-
ях сильно варьирует. Даже если два вида совместно существуют
в одной и той же области, они редко занимают один и тот же участок
обитания, так как конкуренция способствует их специализации.
В Англии пеночка-теньковка Phytloscopus collybita и пеночка-вес-
ничка P. trochilusжиsут в одних и тех же лесах и строят гнезда на
земле. Однако на кормежку теньковки направляются на высокие
деревья, тогда как веснички кормятся на нижних ветвях деревьев
и в кустарнике. Ясно, что у них произошла специализация, тогда
как на Канарских островах, где веснички неводятся, теньковки за-
нимают также и их нишу.
Очевидно, что выбор характерного для вида местообитания со-
ставляет часть нормальной репродуктивной изоляции. У Drosophila
pseudoobscura и D. persimilis, двух близкородственных видов дрозо-
фил в юго-западных областях США, ареалы значительно перекры-
ваются. В лаборатории они гибридизуются особенно хорошо при
низких температурах, но, хотя в природе были пойманы и обследова-
ны десятки тысяч мух, среди них не было найдено ни одного есте-
ственного гибрида. В пределах единого местообитания оба вида ищут
различные микроусловия, D. pseudoobscura — более сухие и светлые,
a D. persimilis — более прохладные и влажные. Такая разница в
предпочтении местообитаний приводит к достаточно эффективной
изоляции этих видов. Даже в условиях лаборатории самки D. pseudo-
obscura не сразу принимают самцов D. persimilis и наоборот, по-
скольку у них различаются брачные «песни» (с. 230). У большинства
животных изоляция на основе местообитания усиливается развитием
в процессе эволюции высокоспецифичных сигналов, которые дают
животному возможность отыскивать особь своего вида иногда на
значительном расстоянии.
Среди насекомых у сверчков, кузнечиков и цикад, а среди позво-
ночных у лягушек и жаб самцы в сезон размножения издают «хоро-
вые крики», которые привлекают самок (о насекомых см. [2], а о
бесхвостых амфибиях см. [53]). В одной и той же области могут
жить несколько разных видов: даже из одного пруда или лужи мо-
жет раздаваться пение самцов нескольких видов лягушек. Удиви-
тельно, как сильно различаются крики разных видов и как самки
реагируют на крики только своих самцов. Можно не сомневаться,
что специфичный характер их «песен» есть прямой результат отбора.
Например, Блэр [52] описывает два вида лягушек Mycrohyla olivacea
и М. carolinensis, имеющих на юге США общий обширный ареал,
причем первый вид занимает более западные, а второй — более
восточные области. На крайних удаленных участках их ареалов, где
практически обитает один вид, хоровые крики самцов очень сходны.
Однако ближе к центру, там, где их ареалы перекрываются и оба
вида используют для размножения одни и те же водоемы, крики де-
лаются непохожими и легко различимыми. В природных условиях
гибриды попадаются, и Блэр показал, что кроме криков других
преград к гибридизации нет. Самцы рассматриваемых видов, как и
у всех Апига, пытаются спариться почти с любым предметом соот-
ветствующего размера, а самки, помещенные с самцами чужого
вида, с готовностью их принимают.
Пердек [380] приводит сходный пример у двух близкородствен-
ных видов кузнечиков, Chorthippus brunneus и С. biguttulus. Они
очень сходны морфологически и встречаются в одних и тех же обла-
стях, однако их песни сильно различаются, и опять же самки привле-
каются пением самцов только своего вида. Если подманить самку к
самцу чужого вида с помощью магнитофонной записи песни самца

252
Глава 6
Эволюция
253
ее вида, то ухаживание и спаривание протекают нормально и появ-
ляется гибридное потомство.
По-видимому в случаях, подобных вышеприведенным, происхо-
дит перекрывание местообитаний двух популяций, имевших в прош-
лом сходные крики. Однако данные популяции были в течение мно-
гих поколений в географической изоляции и за это время разошлись
по множеству других признаков. В таких случаях отбор благоприят-
ствовал возникновению различий в криках, поскольку появление
гибридов было бы неблагоприятно. Александер и Бигелоу [5], опи-
савшие призывные крики большого числа видов сверчков и кузнечи-
ков, совместно живущих в областях на востоке США, обнаружили,
что у всех этих видов, за одним исключением, крики сильно различа-
ются. Иногда виды бывают столь сходны между собой морфологиче-
ски, что сначала их различали только по песне. Исключение со-
ставляют сверчки Acheta pennsylvanicus и A. veletis, живущие в од-
ной области и имеющие похожие песни. Однако это исключение под-
тверждает правило, поскольку A. veletis и A. pennsylvanicus дости-
гают половой зрелости соответственно весной и осенью и, следова-
тельно, самцы разных видов призывают самок к спариванию в раз-
ные сезоны года. Эти два вида разошлись от общего предка, специа-
лизируясь на раннем и позднем времени размножения. Тот факт, что
в течение их развития песни остались идентичными, свидетельствует
о следующем: если не происходит активного отбора на различия в
криках, то крики остаются очень устойчивыми, по-видимому, пото-
му, что любые отклонения могут быть «не поняты». Наиболее убеди-
тельно этот вывод подтверждается на примере трех видов сверчков
из рода Gryllus — европейского G. campestris, G. bermudensis с Бер-
мудских островов и G. firmus с востока США. Они, по-видимому,
изолированы друг от друга в течение многих тысяч поколений, и, в то
время как по строению они сильно различаются, их призывные
песни остались почти идентичными [4].
Между призывом самца и ответом самки существует взаимная
адаптация. Если отбор благоприятствовал дивергенции песен, то
изменения в поведении особей одного пола должны влиять на выбор
среди партнеров противоположного пола тех, которые дают правиль-
ную реакцию. Не исключено, что неожиданные сильные изменения
окажутся неблагоприятными, поскольку противоположному полу
может быть трудно «следовать» за ними.
Хоровые крики —это только один из типов половой изоляции.
В других группах животных особи находят себе подобных по запа-
ху, характеру окраски или комбинации этих черт. Характер окраски
явно очень важен у некоторых птиц. На одном пруду могут собирать-
ся для демонстраций ухаживания утки двух-трех видов, однако все
селезни раскрашены по-разному.
Различия в запахах очень важны для половой изоляции насеко-
мых. Некоторые девственные самки ночных бабочек привлекают сам-
цов своим высокоспецифичным запахом [423]. У дрозофилы химиче-
ские сигналы видовой идентичности производятся, когда самец
приближается к самке и касается ее кончиком лапки [325].
Если исключить особенности пения, само поведение ухаживания
редко используется для идентификации вида. Различия в поведении
между близкими родственниками, как мы видели, главным образом
количественные. По-видимому, отбор на дивергенцию наиболее эф-
фективен в том случае, когда окраска или запах изменяются очень
резко. Особенности ухаживания способствуют половой изоляции
тем, что характерные для этой демонстрации позы и движения уси-
ливают заметносгь специфической окраски или запаха.
Половая изоляция дает нам пример эволюции различий в поведе-
нии как фактора, способствующего различию между видами. Следу-
ет, однако, отметить, что иногда отбор благоприятствует противопо-
ложной тенденции и способствует эволюции сходства с животными
другого вида — опять-таки для их блага. Что касается окраски,
то здесь наиболее известен пример мюллеровской мимикрии. Мно-
#
ДрозЗ \
(Turc/us merula) I
Г
3 5
Большая синица
(.Parus major)
!
0.5
Время, с
Рис. 6.11. Спектрограммы звуков при криках тревоги пяти видов ворьбьиных птиц
трех разных семейств [333]. Они поразительно сходны по форме и высоте. Во всех
случаях звук трудно лоцировать.
'i t lr
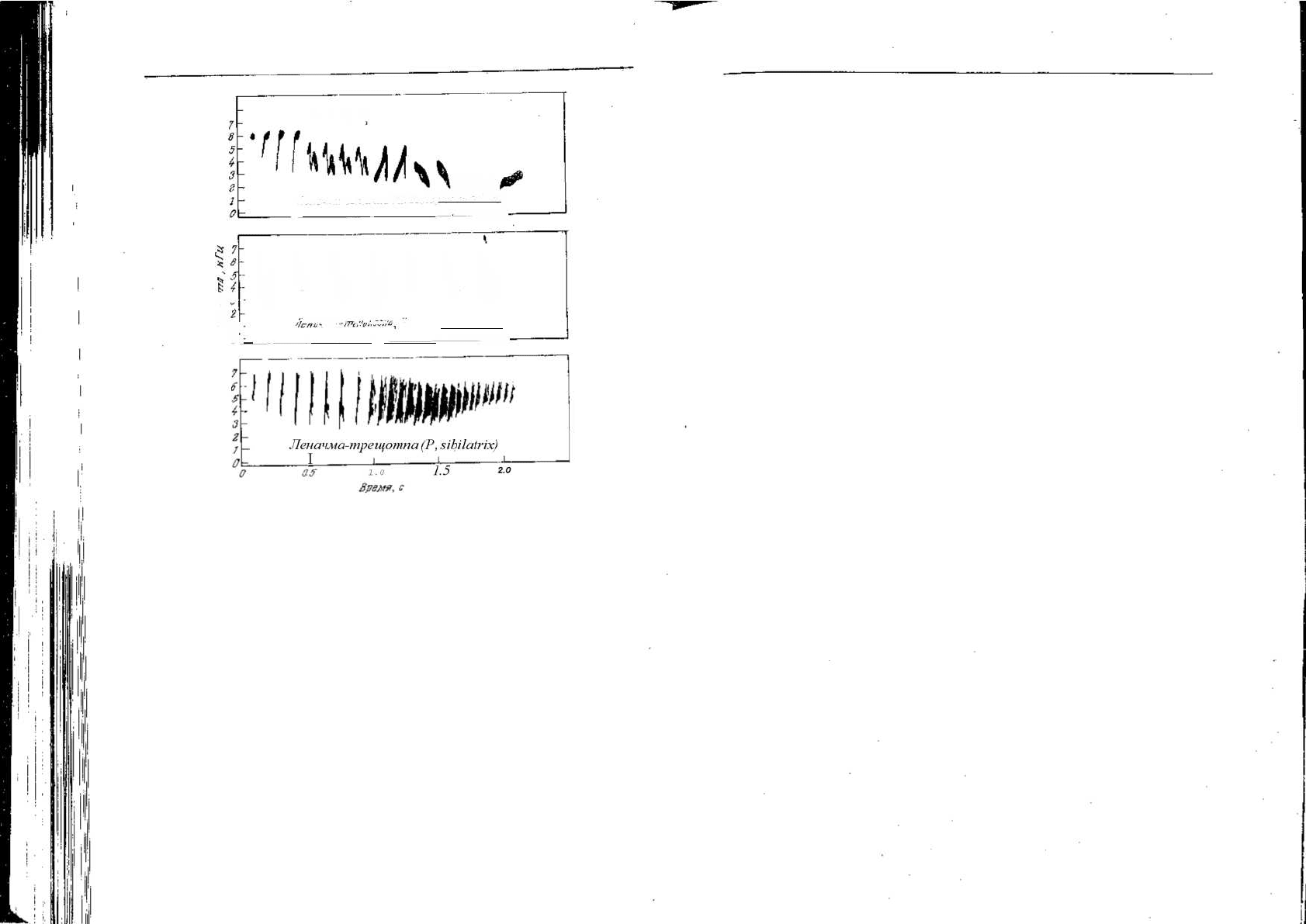
218
Глава 6
Эволюция
254
I
\ M \
Г /'
\ N
mi
Пеноню-мсничяарНзИозсориз tnnMlus)
_J I I L
I 3-
I
* /
-
Ob
A
И Ц i U
.•• •• .•••.--•- •• collybita)
L l_ £ L
Рис. 6.12. Спектрограммы звуков песни трех близкородственных видов европей-
ских пеночек [333]. Эти виды очень сходны по цвету оперения и живут на одной
территории. Их песни, вероятно, должны, по крайней мере частично, способ-
ствовать половой изоляции Звуки их песен резко различаются по форме и харак-
теру. Общее свойство песен заключается в том, что их источник обнаруживается
очень легко.
гие насекомые, имеющие неприятный вкус (например, некоторые
гусеницы, божьи коровки), или жалящие насекомые (осы, шмели)
имеют яркие полосы или пятна черного и желтого или черного и
оранжевого цвета. Такой же тип окраски встречается у некоторых
рыб, амфибий, ящериц и змей — в каждом случае это бывает связано
с каким-нибудь мощным приспособлением к защите от врагов. Отбор
должен благоприятствовать тому, чтобы ядовитое или неприятное
на вкус животное имело заметную раскраску и заметное поведение
или по крайней мере не старалось прятаться. В результате одного
неприятного опыта хищники обучаются избегать контакта с подоб-
ными особями. Мюллер предположил, что сходный тип предупреж-
дающей окраски используется многими животными. Напрашивается
вывод, что у этих животных в эволюции вырабатывалось сходство
друг с другом, так как каждый вид получает выгоды от предупреж-
дающей окраски других внешне сходных видов. Хищник, избегаю-
щий неприятной на вкус добычи определенного вида, будет избегать
и другую, похожую на нее, не обучаясь этому отдельно. Ясно, что
эта ситуация благоприятна для особей любого вида жертвы.
Конвергенция особенностей окраски обнаруживается также у
птиц тех видов, которые образуют смешанные стаи для кормежки
(более подробно значение стай мы обсудим в гл. 8). Мойнан [359]
описывает ряд американских тропических видов птиц — западно-
панамских танагр, славок и вьюрков, которые образуют смешанные
стаи и имеют в основном черную и (или) желтоватую окраску, иногда
с белыми пятнами. Птицы этих видов стремятся собираться и дер-
жаться вместе, а сходство окраски, возможно, эволюционировало
для поддержания спаянности группы. Мойнан называет это явление
«социальной мимикрией».
В гл. 3 мы упоминали о том, что многие мелкие птицы реагируют
на крики тревоги птиц других видов точно так же, как и на крики
своего вида, и что в процессе эволюции произошла конвергенция
характера таких сигналов, в результате которой возник общий сиг-
нальный раздражитель. Мерлер [329] рассматривает свойства, кото-
рыми должен обладать «идеальный» крик тревоги, чтобы в наимень-
шей степени служить хищнику указанием на местонахождение кри-
чащей птицы. Помимо всего прочего этот крик должен иметь по-
стоянную частоту и начинаться и кончаться плавно. Резкое начало
звуков способствует их локализации, поскольку позвоночные могут
лоцировать источник звука только по сопоставлению времени, за
которое он достигает обоих ушей. На рис. 6.11 показаны спектро-
граммы криков тревоги ряда мелких птиц, относящихся к разным
семействам,— крики эти очень сходны между собой и удовлетворяют
требованиям, изложенным выше. Это совсем не похоже на террито-
риальное пение трех европейских видов пеночек (рис. 6.12). В данном
случае между родственными видами, живущими в одной области,
произошла дивергенция вследствие влияния половой изоляции и
необходимости демонстраций.
Л

ГЛАВА 7
ОБУЧЕНИЕ
Обучение
257
На протяжении всей книги нам приходилось упоминать об обуче-
нии и о том, как оно влияет на различные аспекты поведения живот-
ных, о котором здесь идет речь; теперь мы сосредоточим внимание на
самом обучении. Обучение вызывает изменения в поведении, часто
довольно продолжительные, и поэтому было бы целесообразно рас-
смотреть его прежде всего как форму развития поведения. Однако
до сих пор подход к обучению был иным, и, как мы видели, занима-
лись этим исследователи самых разных направлений. В этой главе
мы рассмотрим явление обучения в целом и в различных его прояв-
лениях. Кроме того, мы кратко коснемся различий в способности к
обучению у животных разных групп.
Торп 1457] определил обучение «как процесс, состоящий в появ-
лении адаптивных изменений индивидуального поведения в резуль-
тате приобретения опыта». Это определение подчеркивает два суще-
ственных признака обучения. Первый заключается в том, что в норме
обучение проявляется в появлении адаптивных изменений и, как
уже мы говорили в гл. 2, обучение и инстинкт — это два способа
обеспечения животного набором адаптивных ответов на окружающие
его условия. В норме оба этих способа взаимодействуют и, естествен-
но, имеют между собой много общего. В случае инстинкта в процессе
эволюции популяции происходит отбор индивидуумов, несущих
оптимальный набор генов. В случае же обучения происходит отбор
оптимальных реакций отдельного животного. Рассел [410—413]
проводит аналогию между обучением и эволюцией в наиболее полной
и убедительной форме.
Второе важное положение, вытекающее из определения Торпа,
состоит в том, что, строго говоря, обучение представляет собой про-
цесс, который мы не можем обычно измерять непосредственно. Мы
оцениваем то, что животные запомнили как результат обучения.
Поскольку нам легче общаться с испытуемыми-людьми, то во мно-
гих отношениях они служат лучшим объектом для исследования
обучения, чем животные. Например, нашу память можно проверить
двумя способами; первый способ — «воспроизведение» — состоит
в том, что испытуемый должен повторить устно или записать ряд
бессмысленных слогов, который он заучивал до этого. Второй спо-
соб — «узнавание» — состоит в том, что среди набора бессмыслен-
ных слогов надо выбрать то, что заучивалось ранее. Задача на узна-
вание обычно легче, чем задача на воспроизведение, потому что в
первом случае испытуемый обеспечен раздражителями, которые,
так сказать, «подстегивают» его память и облегчают процесс узна-
вания. Если мы научим крысу проходить лабиринт, то мы, разуме-
ется, не можем попросить ее нарисовать карту этого лабиринта на
бумаге. Единственный способ понять, что она заучивает и запоми-
нает,— посадить ее обратно в лабиринт и наблюдать за ее поведе-
нием. Если крыса делает ошибки, у нас нет возможности узнать, в
чем причина. В том ли, что она не может правильно научиться, или
в том, что не может воспроизвести усвоенное.
Это сталкивает нас с проблемой механизмов обучения. Что про-
исходит в нервной системе в процессе обучения животного? Когда
молодая выводковая птица впервые затаивается при сигнале тре-
воги, издаваемом родителями, она должна использовать преформи-
рованные наследственные пути в мозгу, благодаря которым слухо-
вой сигнал легко «добирается» до двигательной системы, контроли-
рующей затаивание. Когда крыса учится нажимать на рычаг в скин-
неровской камере, должны устанавливаться новые связи, потому
что до обучения рычаг не вызывает у крысы специального ответа.
Мы знаем также, что просто предъявления рычага недостаточно;
для обучения необходимо наличие закрепления. В какой-то части
механизма обучения учитываются результаты действий крысы,
и если они «положительны», то повышается вероятность того, что
реакция повторится в следующий раз при возникновении той же
ситуации. Далее, мы знаем, что в какой-то части нервной системы
хранится более или менее непрерывная «запись» процесса обучения,
которая может служить для «консультации» или воспроизведения в
будущем.
Изучение процесса регистрации, или следов памяти, и физиоло-
гических механизмов, лежащих в основе их формирования, в те-
чение десятилетий было областью активных исследований. Это одна
из спорных проблем биологии, и добиться прогресса в ее изучении
нелегко. Мы ограничимся обсуждением лишь поведенческих аспек-
тов обучения. Хорошим введением в проблему механизмов обучения
служит обширная серия обзоров, охватывающих многие стороны
ее исследований, изданная под редакцией Розенцвейга и Беннета
[398].
До недавнего времени вопросами обучения животных занима-
лись главным образом экспериментальные психологи. Вслед за
Уотсоном, заложившим основы «необихевиоризма», они почти на
полвека сосредоточили свое внимание на способности к обучению у
доместицированных белых крыс. На базе этих работ возникло не-
сколько школ, в частности школы Халла, Толмена и Скиннера.
Каждая из них пыталась построить систему законов поведения, ко-
торые могли бы более или менее точно предсказать, при каких ус-
9
№ 147
ii II