Карнаухов В.Н. Люминесцентный анализ клеток
Подождите немного. Документ загружается.


которой совпадает с максимумом спектра импульсного высвечивания (рис.27, а, 1). После
прекращения статической биолюминесценции объект длительное время сохраняет способность
отвечать импульсами высвечивания, на электрическую стимуляцию.
Приведенные данные указывают на то, что биолюминесценция, представляющая собой
излучение в пространство определенных порций энергии, развивается параллельно с усилением
энергетического обмена клеток в ответ на раздражение.
Об изменении энергетического обмена
клеток, точнее - терминальной стадии его, свидетельствует значительное изменение их спектров
люминесценции под действием света.
Веществами, ответственными за полосы люминесценции клеток с максимумами 460-470 и
520-530 нм, относительная интенсивность которых зависит от функционального состояния тканей,
являются обычно восстановленные пиридиннуклеотиды (460-470 нм) и окисленные флавопротеины
(520-530 нм) систем терминального окисления. Совпадение максимумов
спектра биолюминесценции
и, по-видимому, полосы люминесценции окисленного флавопротеина, а также резкое уменьшение
интенсивности люминесценции этого флавопротеина при световой инактивации биолюминесценции
указывает на возможность того, что именно окисленная форма флавопротеина является веществом,
ответственным за генерацию биолюминесцентного излучения, и тогда предположительно
реализуется система 3 (рис.25). Однако несколько необычное для флавопротеинов положение
максимума (505-509 нм) не позволяет исключить существования некоего промежуточного
соединения, функцией которого является трансформация химической энергии в световое излучение,
аналогичного негемовой пероксидазе системы 2 (рис. 25).
Видимый свет подавляет биолюминесценцию клеток, не влияя при этом на их
жизнеспособность (о чем свидетельствует нормальное или даже несколько ускоренное биение
ресничек). Это позволяет предполагать, что биолюминесценция
связана именно с вспомогательной
(рис.25, 2, 3), а не с основной (рис.25, 1) системой терминального окисления.
Микроскопические наблюдения биолюминесценции и люминесценции агрегаций при
увеличениях ×100 - ×1000 показывают, что при электрическом раздражении происходит импульсное
высвечивание отдельных внутриклеточных гранул, обладающих участками с интенсивной желто-
зеленой люминесценцией. Такие овальные гранулы, достигающие 6-8 мкм в диаметре
, наблюдаются
в клетках, расположенных под гребными пластинками в области меридиональных гастроваскулярных
каналов [170-172].
Интересно отметить, что биолюминесцентная система с импульсным высвечиванием
представляет собой, по-видимому, систему, аналогичную лазеру с химической накачкой излучающих
молекул на возбужденный энергетический уровень. По мере увеличения заселенности возбужденного
уровня повышается вероятность перехода одной из молекул в основное
состояние с излучением
кванта света, который индуцирует развитие лавинообразного процесса перехода остальных молекул в
основное состояние, что и приводит к появлению биолюминесцентного импульса. Освещение белым
светом подавляет только способность к импульсному высвечиванию, постоянно стимулируя переход
молекул с возбужденного уровня на основной. При этом излучается то же, что и ранее, общее
количество энергии, но за счет равномерного распределения ее во времени интенсивность
непрерывного излучения резко падает.
2.2. Функциональная активность клетки и регуляция
энергетического аппарата
Среди проблем, решаемых методами микроспектрального анализа, особое место занимают
вопросы регуляции энергетического аппарата клетки [23, 64, 65, 173]. Энергетическое обеспечение -
важнейшая сторона жизнедеятельности живых систем. Любое функциональное проявление от
сокращения мышечной клетки и транспорта ионов через мембрану против градиента концентрации
до синтеза специализированных белков или нуклеиновых кислот должно быть оплачено поставками
энергии из соответствующих энергопроизводящих
систем.
Биологической основой применения спектральных методов к изучению взаимосвязи
энергетических систем клетки с ее функциональными механизмами являются два важных
обстоятельства.
1. Спектры поглощения и люминесценции многих ферментов и коферментов сильно зависят
от того, в какой (окисленной или восстановленной) форме находятся эти компоненты систем
окислительного метаболизма (
Схема метаболизма).
2. Благодаря действию соответствующих систем регулирования, соотношение окисленных и
восстановленных форм компонентов окислительного метаболизма (т.е. скорость выработки энергии)
определяется функциональной активностью клетки.

В качестве иллюстрации этого положения на рис.30 представлено соотношение окисленных и
восстановленных форм компонентов наиболее хорошо изученной м:итохондриальной системы [174]
окислительного фосфорилирования: цитохром с — цитохромоксидаза (
Схема метаболизма) для
разных функциональных состояний митохондрий. Из этого рисунка следует, в частности, что переход
из состояния покоя (состояние 4) в состояние активного обмена (состояние 3) сопровождается
увеличением концентрации окисленных форм пиридиннуклеотида (НАД), флавопротеинов (Фп) и
цитохромов (а + а
3
, с
1
, с, в) и соответствующим уменьшением концентрации их восстановленных
форм. Такая же закономерность характерна и для других систем терминального окисления.
Как уже отмечалось выше, такие компоненты окислительного метаболизма, как
пиридиннуклеотиды и флавопротеины, обладают характерными спектрами люминесценции. Причем,
если пиридиннуклеотиды люминесцируют только в восстановленном состоянии (460-480 им) и
теряют способность люминесцировать при переходе
в окисленное состояние (рис.11), то
флавопротеины, наоборот, люминесцируют только в окисленном состоянии, (520-530 нм) и теряют
способность к люминесценции при переходе в восстановленную форму (рис.12). Поэтому каждое из
приведенных на рис.29 метаболических состояний митохондрий может быть охарактеризовано
Рис. 29. Соотношение
окисленных (светлые
столбики) и
восстановленных (темные
столбики) форм
компонентов дыхательной
цепи митохондрий (рис. 11,
система 2) в разных
функциональных
состояниях.
Действие ингибиторов
приводит к восстановлению
компонентов цепи,
расположенных левее точки
действия ингибиторов, и
окислению компонентов,
расположенных справа от
точки действия ингибитора.
спектром люминесценции с различным соотношением интенсивностей полос излучения
восстановленных пиридиннуклеотидов (460-480 нм) и окисленных флавопротеинов (520-530 нм).
Для иллюстрации возможностей применения метода люминесцентного спектрального
анализа при изучении взаимосвязи энергетики и функций живой клетки могут быть рассмотрены
результаты исследования рецептора растяжения ракообразных [175]. Особый интерес представляет
при этом наличие в препарате рецептора растяжения нервных и
мышечных клеток двух типов,
значительно различающихся по скорости адаптации их к нагрузке.
В состав препарата (рис.30, а) входят два крупных (70-100 мк) механорецепторных нейрона,
медленно и быстро адаптирующихся к раздражению. Короткие дендриты этих нейронов
оканчиваются на тонких пучках мышц, медленно и быстро адаптирующихся к нагрузке. Аксоны
нейронов направляются в соответствующий ганглий
брюшной цепочки [176-181].
При спектральных люминесцентных исследованиях такого препарата оказалось, что клетки
(нервные и мышечные), различающиеся по скорости адаптации их к нагрузке, характеризуются
различными по форме спектрами люминесценции (рис.30, б-г). Положение полос излучения в
спектрах и изменение их амплитуды при действии на препарат ингибитора гликолиза -
моноиодуксусной кислоты (рис.30, д) -
и разобщителя окисления с фосфорилированием - 2,4-
динитрофенола (рис.30, в, г) - позволяет полагать, что люминесценция нейронов и мышц обязана
своим происхождением восстановленным пиридиннуклеотидам (470—460 нм) и окисленным
флавопротеинам (520 нм) систем окислительного метаболизма.
Соотношение интенсивностей полос люминесценции пиридиннуклеотидов и флавопротеинов
в спектрах свидетельствует о разном метаболическом состоянии энергосистем исследованных клеток.
Медленно адаптирующиеся нервные (рис.30, б) и мышечные (рис.30, в) клетки характеризуются
спектрами с примерно одинаковыми интенсивностями полос восстановленных пиридиннуклеотидов
(460-470 нм) и окисленных флавопротеинов (520 нм). Такое соотношение окисленных и
восстановленных форм компонентов дыхательных систем характерно для тканей, находящихся в
состоянии активного обмена (рис.29, 3). Скорость переноса электронов по дыхательной цепи при
этом максимально высока, и действие разобщителя окисления с фосфорилированием 2,4-
динитрофенола практически не меняет соотношения интенсивностей полос ПН — Н
2
и ФП (рис.30,
в).
Наблюдаемое уменьшение амплитуды спектра связано со значительным поглощением
возбуждающего люминесценцию излучения слоем раствора 2,4-динитрофенола.
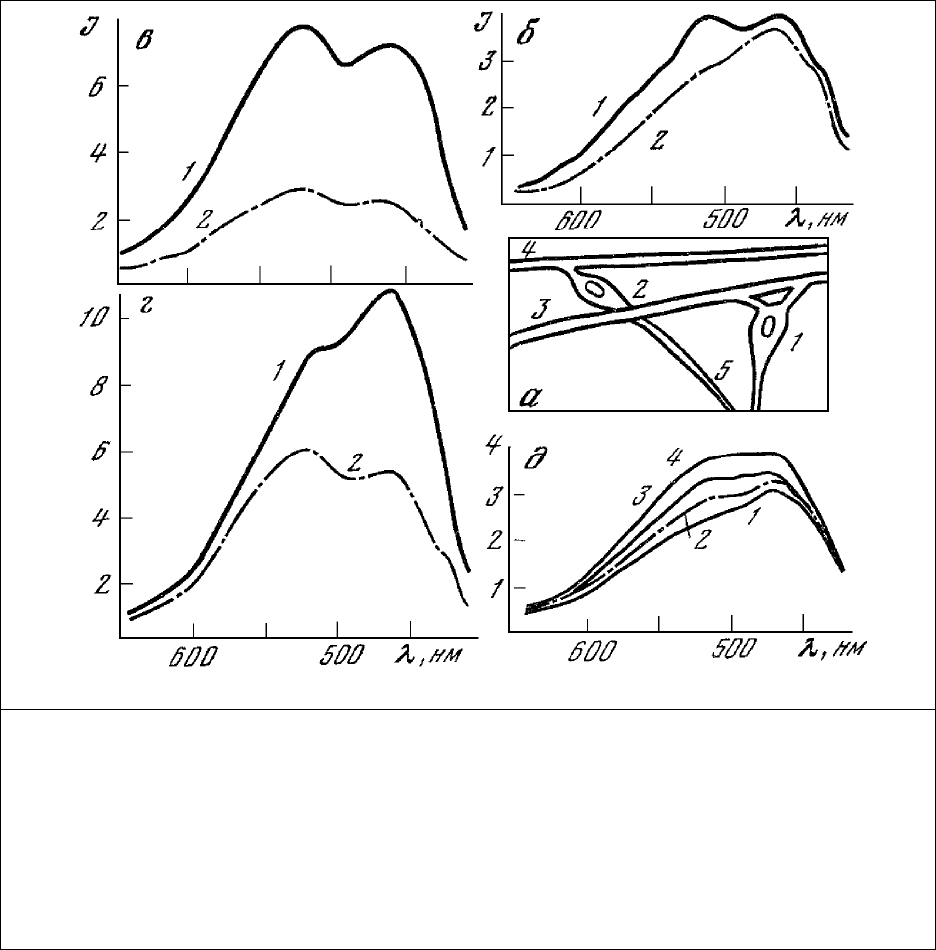
Рис. 30. Схема рецептора и спектры люминесценции нейронов и мышц рецептора растяжения
речного рака.
а – медленно (1) и быстро (2) адаптирующиеся нейроны, медленно (3) и быстро (4) адаптирующиеся
мышцы, аксоны (5); б – медленно (1) и быстро (2) адаптирующиеся нейроны; в – медленно адаптирую-
щаяся мышца до (1) и после (2) действия 2,4-динитрофенола в течение 4 мин; г – быстро адаптирующаяся
мышца до (1) и после (2) действия 2,4-динитрофенола в течение 4 мин; д – изменение спектра
люминесценции быстро адаптирующегося нейрона (1) после
действия на препарат моноиодуксусной
кислоты в течение 15 (2), 13 (3) и 16 (4) мин. Длина волны возбуждения 365 нм; размер
фотометрируемого участка 10×40 мкм.
В отличие от этого спектры быстро адаптирующихся нервных (рис.30, б) и мышечных (рис.30,
г) клеток характеризуются ярко выраженным преобладанием полос люминесценции
восстановленных пиридиннуклеотидов. Такое соотношение окисленных и восстановленных форм
компонентов дыхательной цепи характерно для тканей, находящихся в состоянии покоя (рис.29, 4).
Введение в препарат 2,4-динитрофенола приводит, как это и
должно быть при разобщении окисления
с фосфорилированием, к резкому изменению формы спектра люминесценции клеток.
Преобладающими при этом становятся полосы люминесценции окисленных флавопротеинов (рис.30,
г), а сами спектры приобретают форму, аналогичную форме спектров медленно адаптирующихся
тканей (рис.30, б, в), т.е. скорость переноса электронов по дыхательной цепи становится максимально
высокой.
Приведенные на рис.30, д спектры люминесценции показывают динамику изменения быстро
адаптирующегося нейрона (рис.30, а, 2) при введении в препарат ингибитора гликолиза -
моноиодуксусной кислоты. Наблюдающееся при этом увеличение концентрации окисленных
флавопротеинов по отношению к концентрации восстановленных пиридиннуклеотидов
свидетельствует, по-видимому, о снижении скорости восстановления флавопротеинов
дегидрогеназами цикла Кребса за счет блокирования гликолиза моноиодуксусной кислотой при
оставшейся неизменной скорости поступления кислорода в дыхательную цепь.
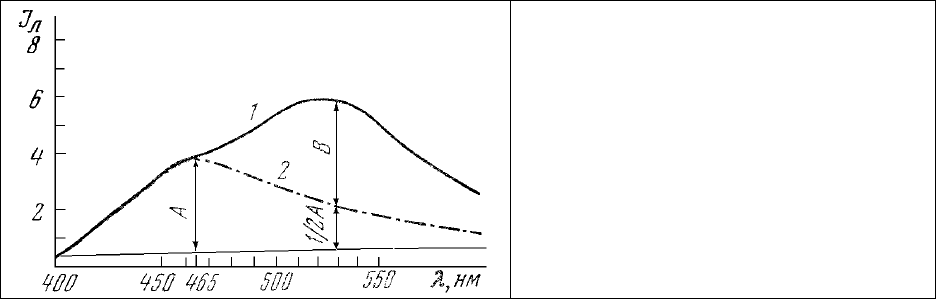
Основываясь на спектрах люминесценции (рис.31), получаемых при возбуждении излучением
Рис. 31. Определение параметра ξ.
–
спектр люминесценции внутриклеточных
митохондрий;
– нормированный спектр люминесценции
раствора НАД Н (1.5⋅10
-4
М) в этаноле для
определения его вклада в максимуме
люминесценции окисленных
флавопротеинов (530 нм).
Длина волны возбуждения 365 нм;
диаметр зонда 10 мкм.
с длиной волны 365 нм, удобно количественно охарактеризовать степень активности
внутриклеточных митохондрий параметром [65].
ξ
=
Ι
фп
/
Ι
пн
== В/А (12)
В этом выражении А представляет собой интенсивность люминесценции восстановленных
пиридиннуклеотидов (НАД⋅Н) в длине волны 465 нм, а В - интенсивность люминесценции
окисленных флавопротеинов, для определения которой необходимо из общей интенсивности
люминесценции в длине волны 530 нм (рис.31, 1) вычесть вклад полосы излучения НАД⋅Н в этой
длине волны. Поскольку интенсивность
люминесценции раствора НАД⋅Н (рис.31, 2) в длине волны
530 нм составляет 0,47 от интенсивности люминесценции этого соединения в максимуме (465 нм)
излучения (см. рис.11, б), для определения величины В с достаточной степенью точности можно из
общей интенсивности в длине волны 530 нм (
Ι
530
) вычитать ½ интенсивности люминесценции
НАД⋅Н в максимуме (
Ι
465
). Тогда выражение (12) для определения параметра ξ приобретает вид:
ξ
= B/A = (
Ι
530
- 0,5
Ι
465
) /
Ι
465
, (12’),
в котором все величины могут быть измерены по спектру люминесценции внутриклеточных
митохондрий (рис.31, 1).
Полученный таким образом безразмерный параметр характеризует степень активности
митохондрий и, как было показано выше (см. уравнение 11, гл.1), не зависит ни от изменения
рассеивающих свойств микрообъекта, ни от изменения таких аппаратурных факторов, как
чувствительность регистрирующей системы,
интенсивность возбуждающего излучения и т. п.
Описываемый препарат рецептора растяжения интересен и в другом отношении. Оказалось
[182], что в его механорецепторных нейронах присутствует два пула митохондрий, различающихся
по своей энергетической активности. Действительно, при микроскопическом излучении нейронов в
свете их собственной люминесценции наиболее высокая плотность люминесцирующих частиц
наблюдается в областях, расположенных вокруг
ядер механорецепторных нейронов (рис.32, 1, 2).
Несколько меньшая плотность таких частиц характерна для областей цитоплазмы, прилегающих к
наружноной мембране нейронов. По данным электронной микроскопии, в околоядерной области и в
примембранных областях цитоплазмы располагаются скопления митохондрий [183].
При проведении микроспектрофлуориметрических исследований оказалось, что митохондрии,
расположенные в различных участках приядерной области, обладают почти идентичными спектрами
люминесценции (рис.33, 1-3) и значительно отличаются по форме спектра от митохондрий,
локализованных в примембранных областях цитоплазмы (рис. 33, 4-6). Эти примембранные
митохондрии, в свою очередь, характеризуются почти одинаковыми по форме (соотношение
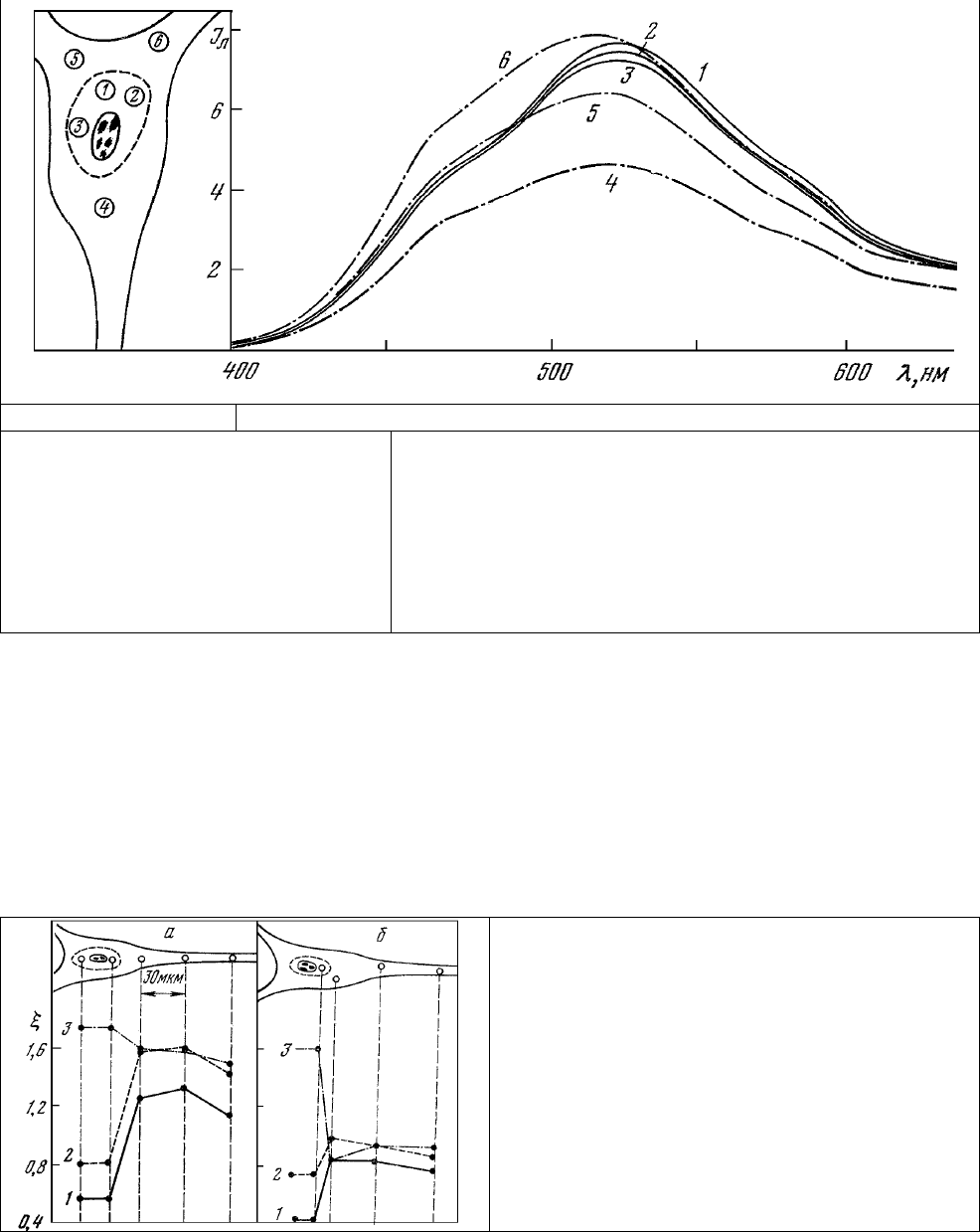
максимумов излучения) спектрами люминесценции. Присутствие в одной и той же клетке скоплений
митохондрий, характеризующихся в один и тот же момент времени разными по форме спектрами
люминесценции, указывает на существование в этих клетках отдельных пулов митохондрий,
находящихся в разных по своей функциональной активности состояниях. В частности, для медленно
адаптирующегося нейрона, приведенного в
качестве примера на рис.33, характерна более высокая
активность приядерного пула митохондрий по сравнению с митохондриями примембранных
областей (рис.33, 3, 5). Не менее часто наблюдается и противоположная ситуация, когда более
активными оказываются митохондрии примембранного пула.
Рис.32 Рис.33
Рис. 32. Рецептор растяжения речного
рака в свете собственной
люминесценции. 1-5 – то же, что на
рис. 31, а; микроскоп МЛ-4, «синяя»
пластинка. Длина волны возбуждения
365 нм светофильтр УФС-6);
запирающий светофильтр – (ЖС-3 +
БС-8).
Рис. 33. Спектры люминесценции приядерной области (1-3)
и периферических участков (4-6) цитоплазмы медленно
адаптирующего нейрона.
Длина волны возбуждения 365 нм; диаметр зонда 7 мкм.
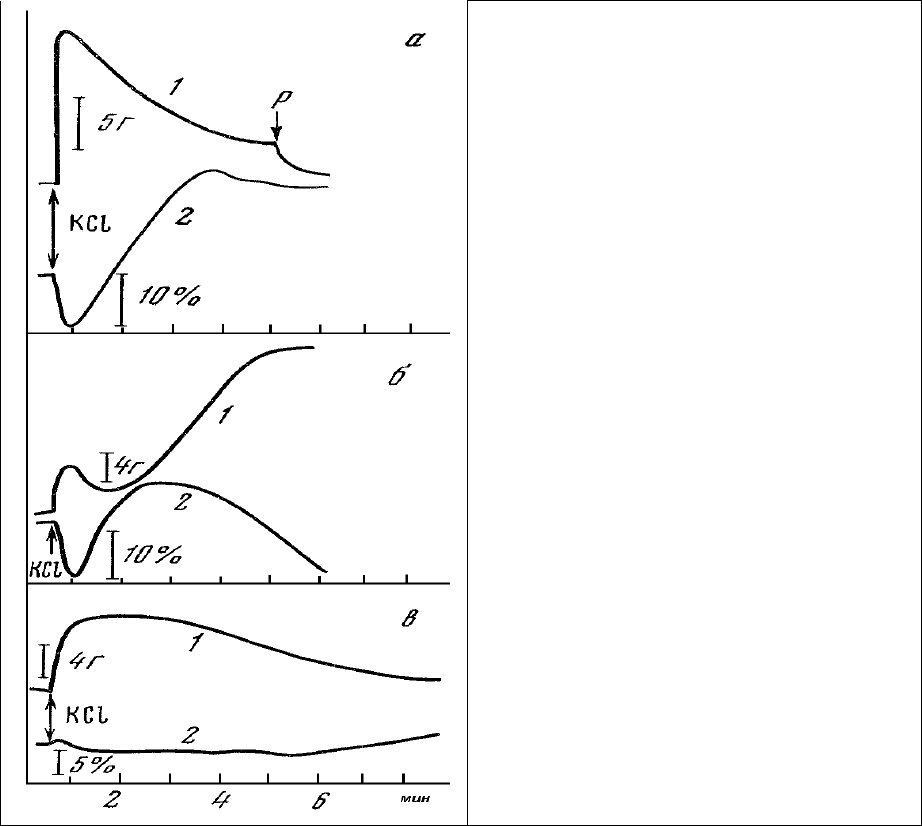
Используя введенный выше безразмерный параметр, характеризующий степень активности
митохондрий, можно проследить как за распределением степени активности митохондрий в
одиночном механорецепторном нейроне, так и за изменением этого распределения при умеренных
физиологических воздействиях на клетку. Приведенные на рис.34, а данные показывают, что в то
время как митохондрии приядерного пула находятся в состоянии
малой активности (ξ=0,6),
митохондри примембранного пула аксона характеризуются высокой активностью (ξ = 0,9÷1,3).
Умеренное растяжение мышц приводит к увеличению активности как приядерных (ξ == 0,8), так и
примембранных (ξ = 1,5÷1,6) митохондрий (рис.35, а, 2). Однако исходное распределение
активностей между двумя упомянутыми пулами митохондрий остается в общем неизменным:
примембранные митохондрии более активны, чем митохондрии приядерной
зоны.
Рис. 34. Распределение состояний митохондрий
(ξ) в двух клетках (а, б) в норме (1), после
растяжения (2) и введения бескальциевой среды
(3) в медленно адаптирующем нейроне рецептора
растяжения речного рака.
Диаметр зонда 7 мкм. По оси ординат –
безразмерный индекс ξ состояния митохондрий.
Резкое изменение соотношений активностей митохондрий разных пулов наблюдается при
замене раствора Харевельда на бескальциевую среду той же тоничности (рис.34, а, 3). При этом
степень активности приядерных митохондрий (ξ == 1,7) становится выше, чем у примембранных (ξ =
1,5÷1,6). Еще более четко изменение соотношения активностей митохондрий двух пулов
прослеживается в случае, приведенном на рис.34, б. Если в исходном состоянии (рис.34, б, 1)
митохондрии приядерного пула (ξ = 0,4) были менее активны, чем митохондрии примембранного
пула (ξ ==0,8÷0,85), то после
замены среды на бескальциевую наблюдается обратное соотношение.
Активность у приядерных митохондрий (ξ = 1,6) становится выше, чем у примембранных (ξ =
0,8÷1,0). По-видимому, именно такое состояние нейронов и зафиксировано на рис. 32.
Приведенные данные указывают на существование в одной клетке двух (приядерного и
примембранного) пулов митохондрий, обеспечивающих энергией, по-видимому, разные
функциональные механизмы,
локализованные в области ядра и в примембранных областях нейрона
соответственно. В настоящее время нет возможности высказать экспериментально
аргументированные суждения о механизме синхронизации активности митохондрий, относящихся к
какому-либо из этих пулов. Неясно, обеспечивается ли эта синхронизация только за счет повышения
скорости потребления энергии соответствующим функциональным механизмом или в основе
наблюдаемого явления
лежит наличие связи между митохондриями одного и того же пула. Такая
связь может быть как прямой, объединяющей наблюдаемые на электронно-микроскопических срезах
митохондрий в единый митохондриальный сросток [184], так и опосредованной через
взаимодействие отдельных митохондрий с какой-либо другой системой.
В этой связи следует отметить, что имеются данные [185] о непосредственной связи
примембранных митохондрий с протяженной системой септального ретикулума, наличие которого
является характерной особенностью исследованных механорецепторных нейронов речного рака
[183]. Возможно также, что все перечисленные механизмы в той или иной мере вносят свой вклад в
наблюдаемую синхронизацию состояния митохондрий, относящихся к одному и тому же пулу.
Как видно из приведенных выше данных, параметр
активности ξ митохондрий достаточно
хорошо описывает поведение системы. Поэтому представляется полезным попытаться более
подробно разобраться в биологическом (биохимическом) смысле этого параметра. Согласно
уравнению (9), интенсивность люминесценции окисленных флавопротеинов в длине волны 520 нм
равна
Ι
Ф
= 2,31 Ι
о
d
о
ε
Ф
k
Ф
c
Ф
, (13)
в то время как интенсивность люминесценции восстановленных пиридиннуклеотидов в длине волны
465—470 нм, измеренная в то же время и в том же участке клетки, составляет
Ι
ПН
= 2,31
Ι
о
d
о
ε
ПН
k
ПН
c
ПН
. (14)
Тогда выражение для определения ξ принимает вид:
ξ
=
Ι
Ф
/
Ι
ПН
=
ε
Ф
k
Ф
/
ε
ПН
k
ПН
⋅
c
Ф
/ c
ПН
, (15)
где ε - молярные коэффициенты экстинкции; k - квантовые выходы люминесценции; c
Ф
-
концентрация окисленных флавопротеинов; c
ПН
- концентрация восстановленных
пиридиннуклеотидов.
По данным Чанса и Вильямса [174], степень окисленности и восстановленности
флавопротеинов и пиридиннуклеотидов меняется в зависимости от функционального состояния
митохондрий (рис.30), и потому концентрация окисленных флавопротеинов в уравнении (15) для
каждого состояния митохондрий может быть определена как
c
Фi
= η
Фi
С
Ф
, (16)
где
η
Фi
— степень окисленности флавопротеинов в данном (i) состоянии; С
Ф
- полная концентрация
флавопротеинов в митохондриях, составляющая 0,72 нмоль/1 мг белка [174].
Аналогичным образом определяется и концентрация восстановленных пиридиннуклеотидов
для определенного функционального состояния митохондрий:
с
ПНi
=
η
ПНi
С
ПН
, (17)
где
η
ПНi
- степень восстановленности пиридиннуклеотидов в данном (i) состоянии; С
ПН
- полная
концентрация пиридиннуклеотидов в митохондриях, составляющая 3,8 нмоль/1 мг белка [174].
Тогда уравнение (15) для определенного (i) состояния митохондрий принимает вид:

ξ
=
Ι
Ф
/
Ι
ПН
=
ε
Ф
k
Ф
/
ε
ПН
k
ПН
⋅
0,72/3,8
⋅
η
Фi
/
η
Пнi
(15')
или
ξ
i
= P
⋅
η
Фi
/
η
Пнi,
(15'')
где
Р = 0,2 ⋅ ε
Ф
k
Ф
/
ε
ПН
k
ПН
- в первом приближении постоянный множитель.
Из сопоставления данных Чанса и Вильямса о соотношении окисленных и восстановленных
форм компонентов дыхательной цепи митохондрий и результатов определения
ξ для митохондрий
нейрона рецептора растяжения (рис.34), приведенных в табл.3, можно видеть, что
ξ оказывается
практически равным отношению степени окисленности флавинов к степени восстановленности
пиридиннуклеотидов и постоянный множитель в уравнении (15'') близок к 1:
Р = 0,2ε
Ф
k
Ф
/
ε
ПН
k
ПН
≈
1, при возбуждении люминесценции излучением с длиной волны 365 нм.
Таблица 3
Количественные отношения окисленных флаеопротеинвв и восстановленных
пиридиннуклеотидов в митохондриях
Функциональное состояние
митохондрий [174]
η
Ф
[174]
η
ПН
[174]
η
Ф
/
η
ПН
ξ = Ι
Ф
/
Ι
ПН
1 0,79 0,9 0,88 -
Истощение (2) 1,0 0
Активное состояние (3) 0,8 0,53 1,51 1,6-1,8
Покой (4) 0,4 0,99 0,6 0,4-0,6
Анаэробиоз (5) 0 1,0 0 -
Таким образом, регистрация спектров люминесценции одиночных клеток и их органоидов
позволяет не только качественно, но и количественно следить за работой энергосистем клетки.
Одним из наиболее «простых» объектов для изучения регуляции энергоаппарата клетки ее
физиологической активностью [186] являются, по-видимому, клетки поперечно-полосатой скелетной
мускулатуры, основная функция которых в организме заключается в
преобразовании энергии
химических связей питательных веществ в механическую работу. Возможность одновременно с
измерением интенсивности люминесценции пиридиннуклеотидов и флавопротеинов регистрировать
и функциональную активность мышцы (силу натяжения) значительно облегчает трактовку
результатов.
В принципе, например, ответственным за изменение интенсивности люминесценции клетки в
длине волны 465—470 нм мог бы являться не только митохондриальный НАД⋅Н, но
и НАД⋅Н из
системы гликолиза, а также и НАДФ⋅Н, как это следует из
Схемы метаболизма. Однако в случае
мышечной ткани, где концентрация НАДФ составляет только около 3% от общего количества
пиридиннуклеотидов [188], учитывая, что квантовый выход НАДФ⋅Н в 3-4 раза ниже такового
НАД⋅Н, вкладом НАДФ⋅Н в изменение люминесценции в области 465-470 нм можно пренебречь
[101, 102]. Вопрос же о необходимости учета влияния гликолитического НАД⋅Н на люминесцентный
сигнал
[103, 104] является более сложным. Хотя определенная синхронность разнонаправленного
изменения интенсивности люминесценции пиридиннуклеотидов и флавопротеинов (рис.30, г)
указывает на митохондриальную природу регистрируемых таким образом пиридиннуклеотидов
мышцы (НАД⋅Н), для выяснения этого вопроса потребовались специальные опыты, результаты
которых приведены на рис. 35 [189]. Сила натяжения, развиваемая мышцей лягушки при калиевой
контрактуре регистрировалась с помощью тензодатчика
из электропроводной бумаги [190].
Развитие натяжения (рис.35, а, 1) при калиевой контрактуре в интактной мышце
сопровождается двухфазным изменением интенсивности люминесценции пиридиннуклеотидов
(рис.35, а, 2). При ингибировании гликолиза моноиодуксусной кислотой изменение люминесценции
пиридиннуклеотидов (рис.35, б, 2) сохраняется, в то время как ингибирование цитохромоксидазы
цианистым калием полностью устраняет их люминесцентную реакцию (рис.35, в, 2) на натяжение
(
рис.35, в, 1) при калиевой контрактуре.
Эти экспериментальные данные позволяют полагать, что изменение люминесценции
мышечных клеток в области 4650-470 нм отражает изменение концентрации именно
митохондриального НАД⋅Н.
Клетки разных тканей, по-видимому, отличаются особенностями механизмов энергетической
регуляции. Поэтому для построения общих представлений необходимы исследования широкого
плана. Однако изучение мышечных клеток вызывает особый интерес в связи с тем, что их основной
функцией является именно преобразование химической энергии в механическую работу, которое
характеризуется при этом и наиболее широким диапазоном развиваемых мощностей.
Рис. 35. Изменение натяжения (10 и
интенсивности люминесценции
пиридиннуклеотидов (2) мышцы лягушки
при калиевой контрактуре [189].
а – нативная мышца, 122 мМ КCl; Р –
отмывка раствором Рингера;
б – мышца после предварительной
инкубации (50 мин) в растворе
моноиодуксусной кислоты (5 мМ); 200 мМ
KCl;
в – мышца после предварительной
инкубации (40 мин) в растворе цианистого
калия (5 мМ); 122 Мм KCl.
Измерение люминесценции
в длине волны
465 нм, длина волны возбуждения 365 нм.
Исследование люминесцентной реакции НАД⋅Н мышечной клетки в ответ на единичное
сокращение и кратковременную стимуляцию к выполнению работы разной мощности [191]
позволяет полагать, что в скелетных мышцах связь между энергетикой и функцией является
достаточно мягкой благодаря наличию мощных демпфирующих систем-аккумуляторов (запасы АТФ
и креатинфосфата). Запасов макроэргов в этих аккумулирующих системах
достаточно для
энергообеспечения одиночного сократительного цикла и короткой серии (5-10) сокращений. При
этом активация системы окислительного фосфорилирования происходит в течение одной или
нескольких секунд после окончания сократительного цикла и направлена на восполнение запасов
макроэргов в аккумулирующих системах. Таким образом, при нагрузках малой мощности система
окислительного фосфорилирования не принимает непосредственного участия в осуществлении
сократительного цикла.
При более мощных нагрузках (калиевая контрактура), величины которых превышают
мощность аккумулирующих систем, митохондриальная система окислительного фосфорилирования
оказывается непосредственно вовлеченной в осуществление сократительной функции, обеспечивая
развитие фазы расслабления. Сравнение приведенных на рис.36, а, в данных показывает, что
выключение системы окислительного фосфорилирования цианидом (рис.35, в) не препятствует
быстрому развитию контрактуры,
хотя амплитуда натяжения при этом уменьшается, по-видимому, в
связи с уменьшением запасов макроэргов в аккумулирующих системах. Фаза же расслабления
оказывается сильно затянутой в связи с прекращением выработки АТФ в митохондриальной системе
окислительного фосфорилирования. При этом единственным источником АТФ становится, по-
видимому, анаэробный гликолиз. Резкое замедление расслабления (рис.35, в) указывает
на то, что
мощность гликолиза (скорость выработки АТФ) в несколько раз ниже, чем у митохондриальной
системы терминального окисления (рис.35, а).
Учитывая также и малый КПД гликолитической системы по сравнению с окислительной (в 20
раз ниже), можно полагать, что гликолиз представляет собой в мышце не столько альтернативный
механизм производства АТФ, сколько основную систему поставки субстратов в цикл Кребса для
обеспечения работы систем окислительного фосфорилирования. Действительно, выключение
гликолиза
моноиодуксусной кислотой (рис.35, б) приводит к тому, что расслабление, будучи
энергетически не полностью обеспеченным, происходит не до конца, после чего следует новая фаза
сокращения, сопровождающаяся окислением митохондриального НАД⋅Н в связи с прекращением
поставки водорода из цикла Кребса, который, в свою очередь, не получает субстратов из системы
гликолиза, причем в скелетных
мышцах в норме гликолиз является, по-видимому, основным
поставщиком субстратов в цикл Кребса.
Как упоминалось выше, способность мышечных клеток осуществлять функционирование в
широком диапазоне мощностей представляет особый интерес, и потому служила объектом
специальных исследований [169, 192]. Было установлено, что при длительной работе мышцы
лягушки с большой мощностью (высокая частота стимуляции) происходит изменение
путей
выработки энергии, включение, по-видимому, менее экономичных систем с более высокой
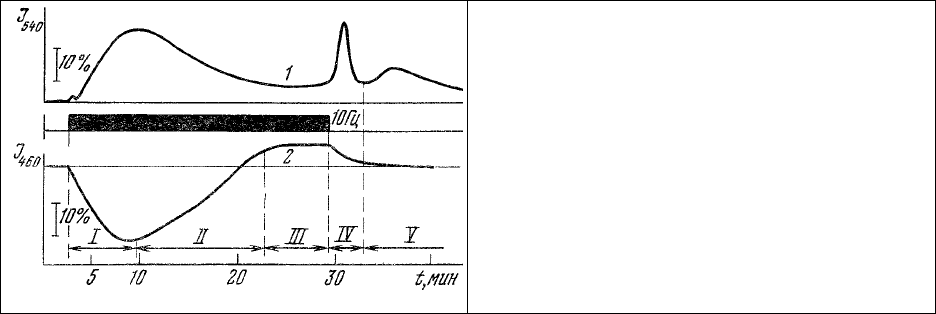
мощностью. Приведенные на рис.36 данные показывают, что в первой фазе процесса наблюдается
обычная для дыхательной системы митохондрий [174] реакция увеличения окисленных (рис.36, 1) и
уменьшения восстановленных (рис.36, 2) форм компонентов цепи терминального окисления НАД
⋅
H-
цитохромоксидаза. Однако максимальная мощность этой системы оказывается, по-видимому, не
достаточной для обеспечения сократительного аппарата в этом режиме и потому в фазе II
наблюдается переход на иной путь терминального окисления. По окончании переходного процесса
(фаза II) энергообеспечение сократительного аппарата уже полностью осуществляется
энергосистемой высокой мощности (фаза III), т.е. происходит своего рода
адаптация энергетики
клетки к наложенной на нее нагрузке. Одним из указаний на наличие такой адаптации является
колебательная реакция окисленных флавопротеинов (рис.35, 1) на снятие нагрузки (фаза IV).
Описанная реакция энергетической системы мышечных клеток проявляется только при
относительно высоких мощностях (частотах стимуляции) нагрузки. При малой мощности (до 1-1,5
Гц) переключения путей терминального окисления не
происходит и наблюдается только описанный
Чансом и Вильямсом [174] переход митохондриальной системы из состояния покоя в состояние
активного обмена. Ввиду того, что данные эксперименты проводились на мышечных клетках in situ,
наблюдаемые эффекты не могут быть отнесены за счет каких-либо нарушений жизнеобеспечения
тканей.
Рис. 36. Изменение концентрации окисленных
флавопротеинов (1) и восстановленных
пиридиннуклеотидов (2) мышечной клетки
при длительной стимуляции с частотой 10 Гц.
Длина волны возбуждения 365 нм. I – V –
фазы адаптации энергоаппарата клетки.
Полезно при этом напомнить одну из работ Хилла [193], в которой, сравнивая скорость
потребления кислорода и выполняемую мышцей работу, он показал, что коэффициент полезного
действия преобразования мышцей химической энергии в механическую оказывается наиболее
высоким (≈25%) при частотах сокращения 1—2 Гц. При частотах выше 1 Гц КПД резко падает.
Несомненно, что исследования регуляции энергоаппарата
мышечных клеток находятся в настоящее
время еще в стадии феноменологического накопления сведений. Однако значение этого этапа нельзя
недооценивать, так как при изучении живой клетки в целом выявляются те особенности систем,
которые обычно элиминируются при деструкции клеток в соответствии с требованиями методов
классической биохимии. Большой интерес, в частности, представляют сведения о
регулирующей
роли кислорода, в работе энергосистем клеток разного типа [187, 194]. При параллельной
регистрации концентрации НАД⋅Н и дыхания [195] мышц был обнаружен любопытный феномен
задержки дыхания (рис.37, 2) и быстрого восстановления митохондриального НАД⋅Н (рис. 37, 1) в
момент включения стимуляции мышц с относительно высокой (более 1,5 Гц) частотой. Величина
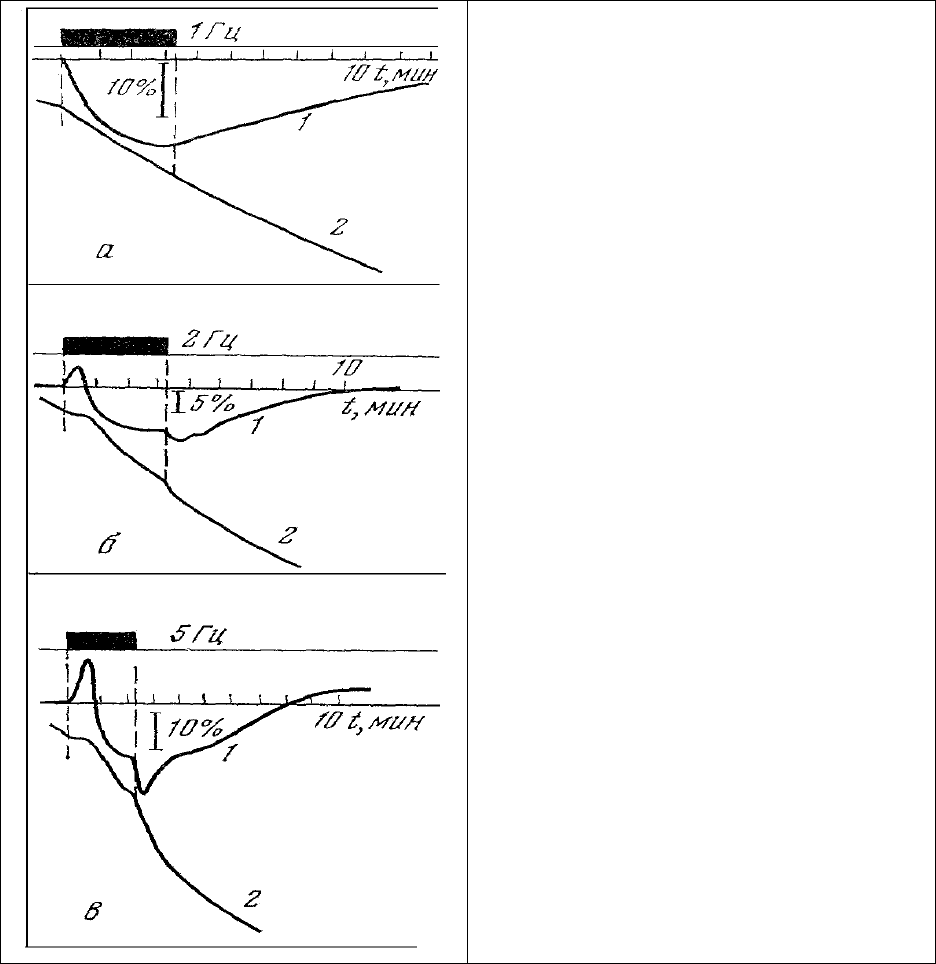
эффекта растет с увеличением частоты. По-видимому, в этом случае мы сталкиваемся с проявлением
действия неизвестной нам обратной связи между активностью системы окислительного
фосфорилирования и скоростью потребления ею кислорода. Увеличение скорости потребления
кислорода в момент выключения нагрузки делает предположения о наличии обратной связи в
достаточной мере обоснованными для дальнейших экспериментальных исследований
природы этой
связи.
Рис. 37. Изменение люминесценции НАД Н
(460 нм) (1) и скорости потребления кислорода
(2) мышцей лягушки при стимуляции с
частотой 1 (а), 2 (б) и 5 (в) Гц [195].
Заканчивая главу, следует подчеркнуть особые преимущества люминесцентного
спектрального анализа в изучении физико-химических процессов, протекающих непосредственно в
живой, функционирующей клетке. Возможность возбуждать люминесценцию излучением,
фокусируемым на объект исследования тем же микрообъективом, который используется и для сбора
света люминесценции, позволяет изучать клетки, лежащие на поверхности или на глубине 200—300
мкм от нее,
не выделяя их из организма животного. Таким образом, в отличие от метода
абсорбционной спектрофотометрии, где толщина объекта не может быть более 100—150 мкм, метод
люминесцентного спектрального анализа позволяет производить исследования клеток in situ. При
этом отпадает необходимость в наиболее сложной части экспериментов по физиологии клетки, а
именно в поддержании адекватных условий внешней среды
для выделенной из организма клетки. Эта
задача возлагается на само подопытное животное [169, 196-198].
Именно такой подход был использован, например, при изучении адаптации энергоаппарата
мышечных клеток, обсуждавшейся выше (рис.36). Живую лягушку помещали в препаровальную
камеру и закрепляли в ней в таком положении, чтобы ее конечности, закрепленные лигатурами к
бортикам камеры в слегка
растянутом состоянии, оказались в удобном для дальнейшей работы