Голиков А.Н. и др. Физиология сельскохозяйственных животных
Подождите немного. Документ загружается.


21 19
98 Фазы движения конечности лошади
при шаге:
цифры в кружках — для левой конечно-
сти,
для правой — без кружков — показы-
вают последовательность положений дви-
жения конечностей
фазы движения шагом: фазу опира-
ния и фазу висения конечности,
или фазу опоры и фазу переноса ко-
нечности (рис. 98). Шаг считается
правильным, если задние копыта по-
падают в следы передних. Он будет
коротким, если следы задних конеч-
ностей не доходят до передних, и
длинным, когда следы задних конеч-
ностей перекрывают следы передних.
Последний — это хороший шаг. Во
время рыси происходит одновре-
менный подъем передней и задней
конечностей, расположенных по диа-
гонали.
Поступательная сила при шаге
связана с толкательными стимулами
задних конечностей.
Фазу опирания подразделяют на
два отрезка: передний и задний, при-
чем у здоровых лошадей эти отрезки
равны между собой. В передний
отрезок происходит активное со-
кращение мышц разгибателей конеч-
ности, задний — характеризует вре-
мя опирания и отрыв копыта от
почвы. В осуществлении последнего
имеют значение эластические свойст-
ва пальцевого мякиша, мякишных
хрящей, сращенных с ветвями ко-
пытной кости, а также упругость
рогового башмака и стрелки копыта
лошади.
Шаг следует рассматривать как
основной аллюр лошади. При других
аллюрах — рыси, галопе, карьере —
происходят лишь различные соче-
тания движений конечностей, харак-
терных для этих видов движения.
Особое движение лошади — ино-
ходь характеризуется одновремен-
ным движением конечностей тазовой
и грудной соответствующей части
тела, а не перекрестно. Поэтому у
иноходца во время бега слышны не
четыре такта, а только два.
При движении рысью конечности
движутся приблизительно одновре-
менно. Ширина шага до 3,3 м, ско-
рость движения 18 км в час. При
галопе ширина шага возрастает до
3—8 м, а при длинном шаге в ска-
ковом галопе наблюдается свобод-
ный полет лошади. Скорость движе-
ния более 1 км в минуту.
Аллюры. Аллюры — это сложно-
рефлекторные акты. Они приобре-
таются в результате систематиче-
ского тренинга.
Прыжок — один из видов слож-
ных движений, совершенствующихся
и изменяющихся в результате систе-
матических упражнений нарастаю-
щей сложности.
Прыжки могут быть горизонталь-
ными — через ров, вверх — через
барьер, импровизированную стенку
или вниз — с уступа через невидимый
ров (немецкая ловушка), через реку
и др. Прыжок состоит из фазовых
движений: разбег, замедление у пре-
400

пятствия, отталкивание, полет, при-
земление. Вначале лошадь отталки-
вается от почвы передними конечно-
стями, при этом голова закидывается
слегка назад, а туловище становится
в положение «стойки». Затем про-
исходит быстрый, сильный удар тазо-
выми конечностями с одновременным
опусканием головы и броском тела
вперед и вверх. Этот толчок осу-
ществляется всеми мышцами тазо-
вого пояса и разгибательной группой
тазобедренных, коленных и скака-
тельных суставов. В результате ло-
шадь перелетает через препятствие
с согнутыми конечностями. При-
земление происходит на обе грудные
конечности при поднятой голове
(рис.
99). Положение конечностей
при толчке и полете связано с тону-
сом мускулатуры и зависит от сигна-
лов,
подаваемых всадником. Несвое-
временное и неправильное вмеша-
тельство в движение лошади при
прыжке может отразиться на поло-
жении туловища и головы и привести
к рефлекторному изменению положе-
ния конечностей (рис. 100). Поэтому
99 Кинограммы основных фаз прыжка
лошади (по В. И. Дорофееву)
существует правило «не мешай ло-
шади при прыжке». В прыжке исклю-
чительную роль отводят зрительному
анализатору, что указывает на боль-
шую сложность этого аллюра, по-
этому близорукие лошади обычно не
могут «взять препятствие» и не до-
пускаются к тренингу.
Лошади поддаются выездке и по-
казывают сложные фигуры, напри-
мер пируэт — вращательный тип
движения с полным оборотом вокруг
своей оси и лансаду — крутой высо-
кий прыжок, смена ног на галопе;
два элемента — пассаж и пиаффе
могут выполнять только некоторые,
хорошо выдрессированные лошади.
Определенный интерес представ-
ляют особенности движения лакти-
рующих коров и молодняка. При дви-
жении вперед у тяжелой молочной
коровы нижняя часть тазовой конеч-
ности вращается внутрь, а затем на-
ружу. В первой стадии шага, когда
суставы, расположенные дистально
к коленному сочленению, сгибаются,
тазобедренный сустав вращается
так, что и коленное сочленение на-
правлено наружу, а дистальная часть
конечности внутрь. Затем, когда ко-
ленное сочленение разгибается, а
401
15—677
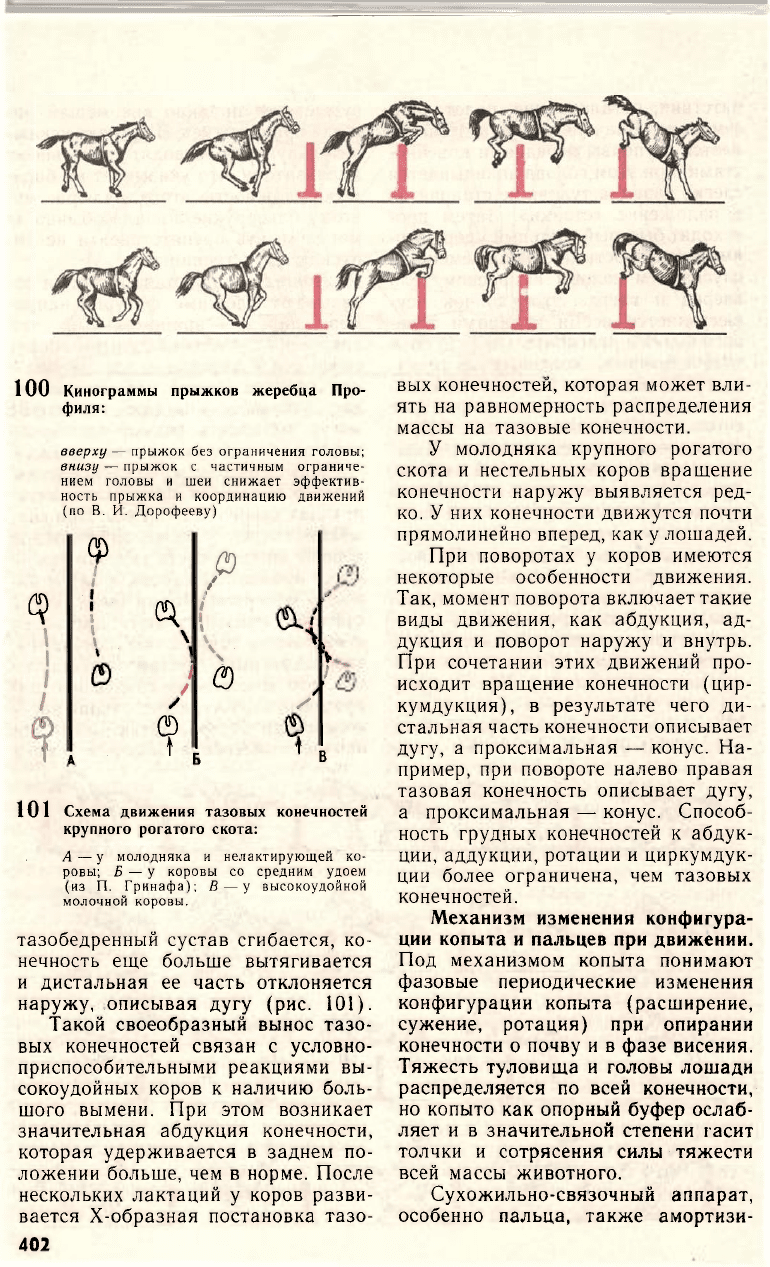
100 Кинограммы прыжков жеребца Про-
филя:
вверху — прыжок без ограничения головы;
внизу — прыжок с частичным ограниче-
нием головы и шеи снижает эффектив-
ность прыжка и координацию движений
(по В. И. Дорофееву)
С?
\
I
/
/
101 Схема движения тазовых конечностей
крупного рогатого скота:
А — у молодняка и нелактирующей ко-
ровы; Б — у коровы со средним удоем
(из П. Гринафа); В — у высокоудойной
молочной коровы.
тазобедренный сустав сгибается, ко-
нечность еще больше вытягивается
и дистальная ее часть отклоняется
наружу, описывая дугу (рис. 101).
Такой своеобразный вынос тазо-
вых конечностей связан с условно-
приспособительными реакциями вы-
сокоудойных коров к наличию боль-
шого вымени. При этом возникает
значительная абдукция конечности,
которая удерживается в заднем по-
ложении больше, чем в норме. После
нескольких лактации у коров разви-
вается Х-образная постановка тазо-
вых конечностей, которая может вли-
ять на равномерность распределения
массы на тазовые конечности.
У молодняка крупного рогатого
скота и нестельных коров вращение
конечности наружу выявляется ред-
ко.
У них конечности движутся почти
прямолинейно вперед, как у лошадей.
При поворотах у коров имеются
некоторые особенности движения.
Так, момент поворота включает такие
виды движения, как абдукция, ад-
дукция и поворот наружу и внутрь.
При сочетании этих движений про-
исходит вращение конечности (цир-
кумдукция), в результате чего ди-
стальная часть конечности описывает
дугу, а проксимальная — конус. На-
пример, при повороте налево правая
тазовая конечность описывает дугу,
а проксимальная — конус. Способ-
ность грудных конечностей к абдук-
ции, аддукции, ротации и циркумдук-
ции более ограничена, чем тазовых
конечностей.
Механизм изменения конфигура-
ции копыта и пальцев при движении.
Под механизмом копыта понимают
фазовые периодические изменения
конфигурации копыта (расширение,
сужение, ротация) при опирании
конечности о почву и в фазе висения.
Тяжесть туловища и головы лошади
распределяется по всей конечности,
но копыто как опорный буфер ослаб-
ляет и в значительной степени гасит
толчки и сотрясения силы тяжести
всей массы животного.
Сухожильно-связочный аппарат,
особенно пальца, также амортизи-
402
ffi
1
1
P
1
1
I
Щ
•
1
1
i
i
i
i
\
\
Ф
)
у
A
a
1
1
в

рует сотрясение и толчки, причем
сила тяжести распределяется на все
мелкие кости карпального и скака-
тельного суставов. В копыте имеются
специализированные ткани: паль-
цевый мякиш, венчик, роговая кап-
сула, вогнутая подошва со стрелкой
и заворотными углами,— которые
в значительной степени способствуют
ослаблению ударов о почву, особенно
каменистую, и защищают мягкие
ткани от повреждения.
В фазе опирания конечности тя-
жесть тела лошади быстро и сильно
переваливается на задний участок
копыта, на область открытой стороны
стенки, иначе говоря, на область мя-
киша. Происходит сильное перераз-
гибание путового сустава (дорсаль-
ная флексия), путовая и венечная
кости принимают более горизонталь-
ное положение; они как бы продавли-
ваются вниз. Наибольшее напряже-
ние испытывают сухожилие поверх-
ностного пальцевого сгибателя и
межкостный мускул с волярными
связками.
Пальцевый мякиш, испытывая
давление сверху и снизу, становится
более плоским и широким и вместе
с мякишными хрящами раздается
в стороны. Вследствие этого эластич-
ная,
упругая роговая стенка рас-
ширяется в пяточных частях. Челноч-
ная часть также вдавливается в мя-
киш, что имеет существенное значе-
ние в расширении копыта при опоре.
В фазе висения конечности или
последовательного сгибания суста-
вов растянутый мякиш постепенно
освобождается от давления, прихо-
дит к исходному эластичному состоя-
нию'
и подобно упругой резине ме-
ханически отталкивает копыто от
почвы, способствуя сгибанию суста-
вов пальца и помогая сухожилиям
мышц сгибателей.
У парнокопытных животных в
фазе опирания пальцы расходятся
в стороны, что ведет к расширению
межкопытной щели. В этот момент
межпальцевые крестовидные связки
растягиваются, а пальцевые мякиши
сдавливаются, амортизируя опира-
ние.
Сила тяжести животного в фазе
опирания конечности о почву пере-
дается на сухожильно-связочный
аппарат, мякиши и роговую капсулу
с многочисленными листочками; все
эти элементы конечности выполняют
роль дистального амортизатора. В
фазе висения конечности потенциаль-
ная энергия, заложенная в растяну-
тых сухожилиях, связках и сжатом
мякише, способствует ее подня-
тию и сближению пальцев.
Центр тяжести и распределение
нагрузки при движении. Центр тя-
жести, обеспечивающий равновесие
животного при движении, деталь-
но выяснен у лошади. Он нахо-
дится в точке пересечения медиаль-
ной плоскости с трансверзальной
у мечевидного отростка грудной ко-
сти и фронтальной плоскости между
нижней и средней третью грудной
клетки. У разных животных центр
тяжести изменяется в зависимости
от массы головы и шеи, а также
положения тела и массы всадника.
Соотношение массы передней части
туловища к задней у лошади со-
ставляет 10:7. Изменение массы той
или иной части туловища перемещает
центр тяжести, вследствие чего утя-
желяется соответствующая часть
тела, что резко нарушает ритм дви-
жения. Например, перегрузка перед-
ней части туловища быстро утомляет
животное.
Работоспособность мышц. Она
зависит от ряда факторов и условий:
1) от правильного чередования
работы и отдыха; оптимальный ритм
движения обеспечивает лучшие ус-
ловия для окислительно-восстано-
вительных процессов в мышцах и
предупреждает утомление;
2) от нормального функциониро-
вания всех систем организма, особен-
но центральной и симпатической
нервной системы, эндокринных влия-
ний, синаптической передачи воз-
буждения с нерва на мышцу, пра-
вильного содержания и кормления
животных;
15*
403

3) продуманный тренинг и пра-
вильное управление животными обес-
печивают наилучшие условия функ-
ционирования всех систем организма
и способствуют выработке полезных
условных рефлексов при выполнении
конкретной задачи;
4) работоспособностьмышцулуч-
шается в процессе тренировки, одна-
ко работающая мышца и организм
утомляются.
Утомление мышцы. В целом ор-
ганизме при работе раньше нервно-
мышечных образований утомляются
нервные центры. При утомлении
мышцы нарушается синаптическая
передача возбуждения с нерва на
мышцу. Так, если мышца в резуль-
тате длительной работы уже не отве-
чает новым сокращением на раздра-
жение двигательного нерва, то ее
можно заставить сократиться, под-
неся электроды от стимулятора не-
посредственно к мышце. Следова-
тельно, утомление в первую очередь
связано с нарушением передачи воз-
буждения с нерва на мышцу, то есть
с недостатком образования ацетил-
холина в синаптических бляшках.
Однако и в самой мышце происхо-
дит ряд биохимических процессов,
характерных для утомления: накап-
ливаются фосфорная кислота, свя-
зывающая ионы Са
+ +
,
молочная
кислота и др.
Перегрузка. Перенапряжение мы-
шечных усилий ведет к быстрому
утомлению. Систематическая чрез-
мерная работа и предъявление жи-
вотному непомерно высоких требова-
ний могут привести к «срыву» —
быстрой утомляемости и нарушению
координации движений.
Непомерная тренировка также
вызывает «срыв», поэтому только
своевременное предоставление жи-
вотному отдыха может восстановить
работоспособность. Животные, ис-
пытавшие перегрузку, долго ощу-
щают ее последствия: у них снижа-
ется сократительная способность
скелетных мышц, расширяются гра-
ницы сердца и др.
При неправильном содержании
животных выделяют понятие «стад-
ное утомление». У свиней при скучен-
ном содержании, недостатке моциона
и свободного передвижения, а также
в связи с гиподинамией или, наобо-
рот, частыми переменами боксов
появляются симптомы повышенной
возбудимости, пугливости, слабости
конечностей, они не могут быстро
и легко ходить и бегать; из-за вы-
деления адаптивных гормонов (но-
радреналина) снижается качество
мяса — «водянистая свинина».
Систематическая и интенсивная
работа мускулов способствует уве-
личению массы мышечной ткани,
такое состояние мышцы называют
рабочей гипертрофией. В ее основе
лежит увеличение массы цитоплазмы
мышечных волокон и числа содержа-
щихся в них миофибрилл, сопровож-
дающееся увеличением диаметра
каждого волокна. Происходит акти-
визация синтеза нуклеиновых кислот
и белков, повышается содержание
веществ, доставляющих энергию со-
кращения (гликогена, АТФ). Проти-
воположное состояние рабочей ги-
пертрофии — атрофия мышц от без-
деятельности. Она возникает в тех
случаях, когда скелетные мышцы
в силу ряда причин бездействуют
или слишком мало участвуют в дви-
гательных актах всего тела, напри-
мер при обездвиживании конечности
после длительного наложения гипсо-
вой повязки, повреждения сухожи-
лий или нервов, отсутствия и недоста-
точности моциона, при клеточном
содержании. Особый вид нейроген-
ной атрофии возникает в случаях
повреждения периферических нер-
вов,
когда мышца лишается нервной
импульсации и обречена на посте-
пенное отмирание вследствие нару-
шения трофики. Ведущее значение
в этих процессах имеет выключение
афферентных импульсов (А. Н. Голи-
ков,
1961).
Тренинг. С физиологической точ-
ки зрения тренинг — выработка по-
лезных условнорефлекторных актов
404

у животных в спортивных или хозяй-
ственных целях. Задача тренинга —
выработать у животного высокую
работоспособность при выполнении
движений определенного спортив-
ного или хозяйственного характера
(развитие быстроты, силы, выносли-
вости, точности движений); совер-
шенствовать координацию между
важнейшими функциями организма.
Образование двигательных навы-
ков происходит по принципу выра-
ботки сложных условных рефлексов,
в формировании которых важней-
шая роль принадлежит коре полуша-
рий мозга, анализаторам экстеро- и
интерорецепторов.
В процессе тренинга кора полу-
шарий мозга получает сигналы из
внешней и внутренней среды и на
базе врожденных двигательных реф-
лексов происходит стойкое закрепле-
ние новых двигательных актов, до-
стигающих весьма высокого совер-
шенства как у лошадей, обезьян,
зебр,
так и у птиц и водоплавающих
животных. В результате многократ-
ных повторений образуется динами-
ческий стереотип, и его соблюдение
и подкрепление обеспечивают наи-
больший эффект. Наоборот, его на-
рушение может вредно сказаться
на организме животного.
Тренинг рекомендуется проводить
систематически и последовательно,
увеличивая сложность упражнений.
Но при этом нужно учитывать
индивидуальные качества животно-
го,
тип его высшей нервной деятель-
ности, следить за состоянием сердеч-
но-сосудистой системы и внутренних
органов. Показатели этих систем
могут служить объективным крите-
рием эффективности тренинга.
ФИЗИОЛОГИЧЕСКИЕ
ПОКАЗАТЕЛИ
ТРЕНИРОВАННОСТИ МЫШЦ
Систематическая интенсивная ра-
бота мышц приводит к увеличению
массы мышечной ткани. Мышца
увеличивается в поперечнике, пре-
имущественно в результате разраста-
ния саркоплазмы. Изменяется хими-
ческий состав мышцы, возрастает
количество белков, особенно глико-
гена, увеличивается диаметр каждо-
го мышечного волокна. В мышце
происходит усиленный синтез белков
и нуклеиновых кислот, увеличивается
содержание веществ, доставляющих
энергию для мышечного сокращения:
аденозинтрифосфата, креатинфос-
фата. В результате сила и скорость
сокращения мышцы возрастают. Уве-
личение массы мышечной ткани при
систематической работе приводит к
рабочей гипертрофии мышц.
Показатель тренированности
мышц — функциональное состояние
организма, зависящее от способа тре-
нинга, содержания животных (выгуль-
ное,
стойловое), обеспечения мо-
ционом.
Энергетические процессы в мыш-
цах связаны с распадом органиче-
ских веществ и образованием тепла,
но характер сокращения зависит от
вида миофибрилл. Тетанические мио-
фибриллы отличаются способностью
длительно сокращаться вследствие
обратимой деформации молекул со-
кратительных белков. Способность
к сокращению исчезает при темпера-
туре 50 °С. Фазные миофибриллы со-
держат белковые нити, которые при
сокращении укорачиваются, теряя
это свойство уже при 42 °С.
В процессе тренировки животных
важное значение имеет ресинтез
АТФ,
непрерывно расщепляющейся
при сокращении мышц, и накопле-
ние в миофибриллах свободных
ионов Са
+ +
.
Нарушение ресинтеза
АТФ,
например при отравлении, мо-
жет привести к полному исчезнове-
нию АТФ и креатинфосфата, вслед-
ствие чего «кальциевый насос» пере-
стает работать и мышца может
прийти в состояние длительного све-
дения (контрактуры).
ДВИЖЕНИЕ ПТИЦ
Особенности движения птиц свя-
заны с полетом и с передвижениями
405

на местности. Стояние кур требует
большего напряжения мышц конеч-
ностей, чем у четвероногих. Центр
тяжести проходит у них вне площади
опоры, несколько спереди от тазобед-
ренного сустава. Большую роль в
поддержании равновесия играют
хвост, крылья и длинная шея; они
помогают увеличивать массу той или
иной части тела, и птицы пользуются
этим, изменяя положение шеи или
крыла. Особенности расположения
сухожильно-связочного аппарата
птиц при сидении на ветке или на-
сесте обеспечивают прочное сгибание
пальцев, надежный охват ветки без
напряжения мышечной силы. У водо-
плавающих птиц плавательные пере-
понки выполняют роль гребных ве-
сел,
а перемещение головы опре-
деляет направление движения. Пла-
ванию способствует устройство су-
ставов пальцев, при их разгибании
происходит одновременное растяже-
ние плавательных перепонок. Пла-
вание и полет птиц — это сложно-
координированные физиологические
акты, регулируемые центрами голов-
ного и спинного мозга, мозжечка
и высокоразвитой периферической
нервной системы.
МЕХАНИЗМ РЕГУЛЯЦИИ
ДВИЖЕНИЯ
Во всяком движении, каким бы
простым оно ни казалось, всегда уча-
ствует целый ряд мышц, одновре-
менно или последовательно сокра-
щающихся. Сокращение одной груп-
пы мышц через центральную нервную
систему вызывает изменение в со-
стоянии другой, антагонистической
группы мышц. Такая согласован-
ность в работе отдельных мышц и
мышечных групп, участвующих в
движении животного, достигается
координирующими влияниями со сто-
роны центральной нервной системы.
Механизмы взаимосвязанной иннер-
вации в форме одновременной и по-
следовательной индукции обеспечи-
вают согласованность работы всех
мышц, участвующих в поддержании
равновесия тела и при движении
животного. Центры спинного мозга
регулируют последовательность со-
кращения мышц конечностей, харак-
терную для акта ходьбы, в этом мож-
но убедиться на спинальных живот-
ных. Однако в целом организме
регуляция двигательных актов осу-
ществляется в основном стволовой
частью мозга, мозжечком и под-
корковыми центрами.
При любой форме движения поток
биоэлектрических импульсов от од-
них мышц усиливается, от других
ослабевает, меняя свою частоту и
амплитуду. Импульсы, возникающие
в рецепторах двигательного анали-
затора, поступают в центральную
нервную систему и по проводящим
путям спинного мозга достигают раз-
личных отделов коры больших полу-
шарий.
Основной путь к коре полушарий
состоит из трех групп нейронов.
Первые нейроны лежат в межпозво-
ночных чувствительных ганглиях
(или в ганглиях черепномозговых
нервов); вторые — в ядрах про-
долговатого мозга; третьи — в цен-
трах промежуточного мозга.
Наличие двусторонней афферент-
ной и эфферентной связи между
мышцами и центрами создает замк-
нутую цепь, в которой нервные им-
пульсы, распространяющиеся по дви-
гательным волокнам, вызывают со-
кращение мышц. В свою очередь,
мышца сигнализирует в центры о ка-
честве выполняемой работы (сильно,
слабо, быстро, медленно и т. д.).
Эта обратная связь поддерживает
необходимый уровень функциони-
рования всей мышечной системы
в данной ситуации (движение по пря-
мой, по кругу, галоп, прыжки и др.).
Проприорецепторы мышц при со-
кращении последних воспринимают
раздражение, которое передается в
соответствующие нервные центры,
вызывая ответную реакцию. Благо-
даря сигналам, поступающим от
проприорецепторов мышц, осущест-
406

вляются шейные тонические реф-
лексы, имеющие большое значение
для движения животных. Для осу-
ществления движения важное значе-
ние имеют импульсы, идущие от кож-
ных чувствительных рецепторов. Ве-
лика роль мозжечка и лабиринтов,
центры которых связаны с ядром
Дейтерса. Сигналы, идущие от лаби-
ринтов, регулируют положение голо-
вы,
а это влияет на тонус мышц
конечностей. Например, если лошадь
должна сдвинуть с места значитель-
ный груз, то она вначале опускает
голову, а это перемещает центр тя-
жести и увеличивает тонус задних
конечностей, обеспечивающих толчок
при движении вперед. Если же голо-
ва поднимется кверху, то тонус тазо-
вых конечностей будет ослаблен и
лошадь не сможет «взять с места».
Управление движениями обуслов-
лено совместной деятельностью всех
уровней нервной системы, рассматри-
ваемых многими учеными с позиций
биологической кибернетики. Предмет
биологической кибернетики состоит
в изучении специфических для живых
существ общих принципов и конкрет-
ных механизмов целесообразного
саморегулирования и активного вза-
имодействия с окружающей средой.
Физиология и биокибернетика взаим-
но дополняют друг друга. Биоки-
бернетика помогает понять сложные
процессы саморегуляции движения.
В частности, она рассматривает
мышцу как структурный элемент
локомоторной системы с множествен-
ными обратными связями. Регуляция
работы мышц конечностей осуществ-
ляется тремя типами сенсорных ре-
цепторов: рецепторы мышцы, реаги-
рующие на изменения ее длины;
рецепторы сухожилий, чувствитель-
ные к изменению их напряжений;
рецепторы суставов, реагирующие на
изменение положения конечностей.
Все виды рецепторов играют важную
роль в координации движения,
но наибольшее значение имеют ре-
цепторы первой группы, расположен-
ные параллельно мышечным пучкам.
При растяжении мышц они воз-
буждаются и передают информацию
о степени данного растяжения. В ре-
зультате этого в центрах возникают
командные импульсы, возвращаю-
щиеся по эфферентному пути к мыш-
це и вызывающие ее сокращение.
Считают, что в нервной регуляции
деятельности мышц участвуют мини-
мум три подсистемы. Первая под-
система определяет сократительную
функцию мышц. Она состоит из мото-
нейронов и мышц с расположенными
в них проприорецепторами. Импульс-
ная информация в ней распростра-
няется от клеток передних рогов
спинного мозга к мышцам и от мы-
шечных рецепторов в обратном на-
правлении через заднекорешковую
систему к спинномозговым центрам
и вновь к мотонейронам. Вторая
подсистема обеспечивает оптималь-
ный уровень возбудимости проприо-
рецепторов; третья, состоящая из
аксонных коллатералей мотонейро-
нов и вставочных нейронов Реншоу,
предназначена для саморегуляции
мотонейронов.
Гамма-регуляция мышечного то-
нуса осуществляется двумя группами
мотонейронов спинного мозга: круп-
ными нейронами, вызывающими со-
кращение волокон скелетных мышц,
и мелкими у-мотонейронами, изме-
няющими натяжение внутриверетен-
ных мышечных волокон и повышаю-
щими чувствительность проприо-
рецепторов мышц. Изменение на-
пряжения мышц под влиянием
импульсов у-мотонейронов вызы-
вается гамма-регуляцией. Актив-
ность последних находится под кон-
тролем ретикулярной формации ство-
ла мозга. В поддержании тониче-
ского напряжения мышц участвует
сложная многоуровневая система
нервных центров, находящихся под
контролем коры больших полушарий.
Трофическое влияние импульсов
вегетативной нервной системы тоже
имеет определенное значение в про-
цессе регуляции функции скелетных
мышц. Импульсы симпатической
407

нервной системы воздействуют на
мионевральные синапсы, служащие
местом перехода возбуждения с дви-
гательного нерва на мышцу. Сила
мышечных сокращений утомленной
мышцы увеличивается, если раз-
дражаются симпатические волокна.
Координация движения зависит
от образования тепла в организме.
Чем интенсивнее работа, тем больше
образуется тепла, особенно в жаркое
время года и при насыщенном влагой
воздухе. В таких условиях тепло-
отдача затрудняется, что может при-
вести к нежелательным последствиям
(тепловой удар). Если количество
молочной кислоты, образующееся в
результате значительной физической
нагрузки, может возрасти до 500 %
и более, не принося вреда, то перегрев
тела (гипертермия) на 5 %, т. е. на
2 °С, уже опасен, причем в
первую очередь нарушается коорди-
нация движений в силу высокой
чувствительности центров гипота-
ламуса и ретикулярной формации
ствола мозга. В координации дви-
жения и регуляции мышечного тону-
са важная роль принадлежит моз-
жечку. Последний оказывает также
трофическое влияние на мышцы,
выражающееся в восстановлении
обмена веществ и работоспособ-
ности.

Глава 16
АДАПТАЦИЯ
СЕЛЬСКОХОЗЯЙСТВЕННЫХ
ЖИВОТНЫХ
Под физиологической адаптацией
(от лат. adaptatio — приспособле-
ние) понимают совокупность морфо-
физиологических процессов в орга-
низме, лежащих в основе его при-
способления к конкретным условиям
существования во внешней среде.
В результате адаптации повышается
устойчивость организма к низкой и
высокой температуре воздуха, не-
достатку кислорода, воды, освеще-
ния, виду корма и многим другим
факторам, связанным с экологи-
ческими условиями обитания и пове-
дения разных видов животных. Из-
менение условий среды, естественно,
вызывает функциональные сдвиги
в организме, которые при определен-
ных условиях могут переходить в со-
стояние болезни.
По эколого-генетической класси-
фикации физиологическую адапта-
цию подразделяют на видовую —
унаследованную, индивидуальную,
характеризующую отдельную особь,
и популяционную. Последняя харак-
теризует группу организмов данно-
го вида (породы), развивающуюся
в определенных условиях среды
(А. Д. Слоним, 1962).
Одни виды животных приспосо-.
бились к холодному климату, дру-
гие — к умеренному, третьи — к вы-
сокогорным условиям (яки, ламы),
а некоторые — к тропическому кли-
мату. Имеются различия и в при-
способляемости животных к обита-
нию в умеренной зоне. Одни живот-
ные при относительно оптимальных
условиях хорошо используют скуд-
ные кормовые средства, но обладают
низкой продуктивностью, другие ну-
ждаются в высококачественных кор-
мах и наиболее эффективно превра-
щают их в продукты, потребляемые
человеком. В полупустынных райо-
нах разводят обычно высоконогих,
сухих по телосложению животных,
а в районах влажных низменностей,
наоборот, животных рыхлой консти-
туции.
Влияние среды на организм мо-
жет иметь решающее значение. Жи-
вой организм — это в высшей степе-
ни саморегулирующаяся система,
сама себя поддерживающая, восста-
навливающая, направляющая и да-
же совершенствующая. Однако орга-
низм подчинен периодически повто-
ряющимся изменениям внешней сре-
ды:
смене дня и ночи, сезона года,
приливов и отливов в прибрежных
зонах морей и океанов, температур-
ным перепадам и многим другим эко-
логическим и климатогеографиче-
ским явлениям, которые необходимо
учитывать при определении особен-
ностей адаптации сельскохозяйст-
венных животных.
Предложена классификация адап-
тации, учитывающая природные и
технологические факторы, которые
409