Фёршт Э. Структура и механизм действия ферментов
Подождите немного. Документ загружается.

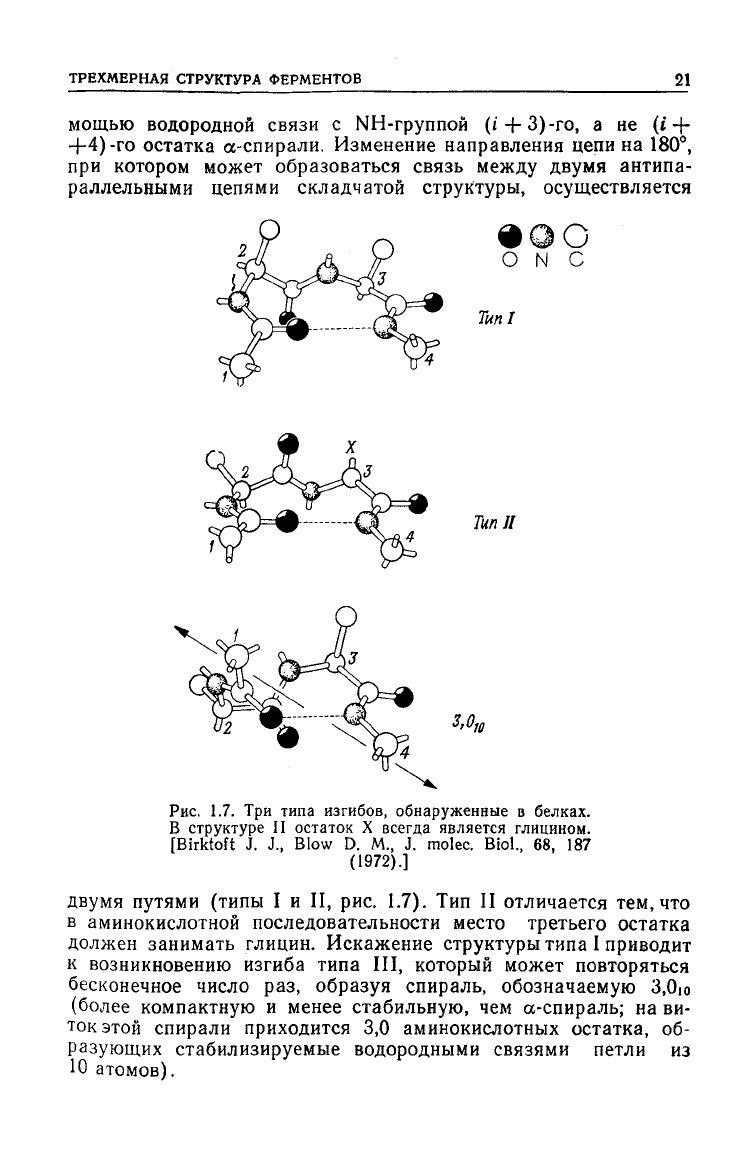
ТРЕХМЕРНАЯ
СТРУКТУРА
ФЕРМЕНТОВ
21
мощью водородной связи с NH-группой (i-J-3)-ro, а не (t-f-
+4)-го
остатка а-спирали. Изменение направления цепи на 180°,
при
котором может образоваться связь
между
двумя антипа-
раллельными цепями складчатой структуры, осуществляется
#0О
О
N С
TunI
Тип
Л
Рис.
1.7. Три типа изгибов, обнаруженные в белках.
В структуре II остаток X всегда является глицином.
[Birktoft J. J.,
Blow
D. M., J. molec. Biol., 68, 187
(1972).]
двумя путями (типы I и II, рис. 1.7). Тип II отличается тем, что
в
аминокислотной последовательности место третьего остатка
должен занимать глицин. Искажение структуры типа I приводит
к
возникновению изгиба типа III, который может повторяться
бесконечное число раз, образуя спираль, обозначаемую 3,0ю
(более компактную и менее стабильную, чем а-спираль; на ви-
ток
этой спирали приходится 3,0 аминокислотных остатка, об-
разующих стабилизируемые водородными связями петли из
Ю атомов).

28
ГЛАВА
I
3. Построение белковых молекул из отдельных блоков
Расшифровка структуры первых
двух
белков вызвала сенса-
цию.
Оказалось, что единственная полипептидная цепь миогло-
бина образует ряд а-спирализованных палочек, чередующихся с
участками, отличными от р-структуры. Затем было установлено,
что субъединицы гемоглобина идентичны четырем цепям миог-
лобина (рис. 1.8). Однако по мере расшифровки структуры
разнообразных белков картина становилась все более сложной.
Лизоцим и карбоксипептидаза — первые ферменты,
структура
которых была установлена, построены из а-спиралей и р-струк-
тур. Вскоре выяснилось, что молекула а-химотрипсина уложена
почти целиком в виде р-структуры, за исключением
двух
а-спирализованных участков. Стало очевидно, что установить
общие правила укладки полипептидных цепей очень трудно.
Первые шаги в этом направлении сделаны лишь в настоящее
время, когда определена
структура
около ста белков. Выявлены
отдельные регулярно повторяющиеся структурные особенности.
Развиваются методы классификации структур.
Предполагается, что большинство белков можно рассматри-
вать в первом приближении как
структуру
типа сэндвича, каж-
дый слой которой состоит либо из а-спиралей, либо из р-струк-
тур. Недавно Левитт и Чотиа [10] предложили простой и на-
глядный способ изображения таких
структур
(рис. 1.9). Упа-
ковка спиральных и складчатых участков происходит за счет
стэкинг-взаимодействия боковых цепей аминокислот. Внутрен-
няя
область белка имеет плотную упаковку: боковые цепи одного
участка встраиваются
между
боковыми цепями
другого,
образуя
гидрофобное ядро, плотность которого близка к плотности жид-
кого углеводорода или воска [И]. В этой области мало водо-
родных связей: так, в карбоксипептидазе А их число равно
только 17; они соединяют
друг
с
другом
8 спиралей и одну
складчатую
структуру,
т. е. на каждый спиральный участок
приходится менее
двух
водородных связей. Расположенные во
внутренней области белка доноры и акцепторы, способные обра-
зовывать водородные связи, всегда спарены. Участки, соединя-
ющие
друг
с
другом
спирали и слои, обычно доступны для
растворителя и непротяженны. Таким образом, глобулярные
белки имеют гидрофобное ядро, на поверхности которого распо-
ложены заряженные группы.
Согласно данным Крика [12.], две а-спирали (имеющие оди-
наковое направление) способны плотно упаковываться, если
угол
между
ними составляет 20°; при этом боковые группы одной
спирали встраиваются
между
боковыми группами другой. Дан-
ный
расчет был проделан для случая, когда на один виток
спирали приходится 3,6 аминокислотных остатка. Для более
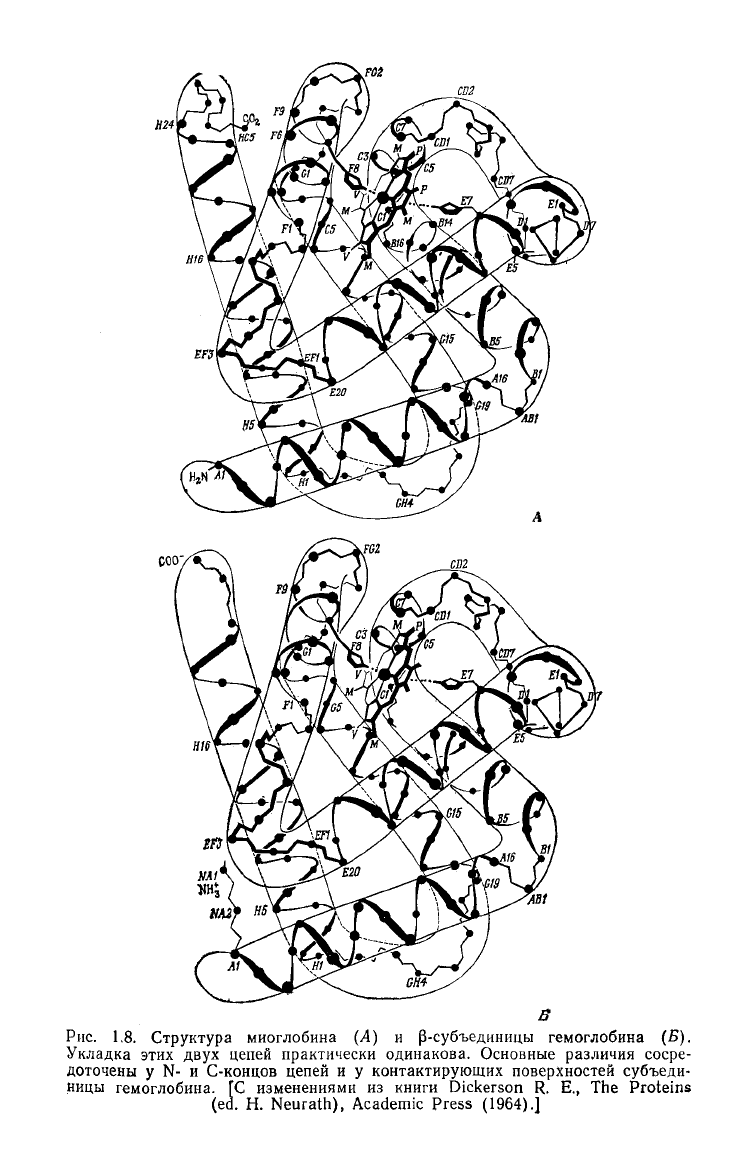
Рис.
1.8. Структура миоглобина (А) и р-субъединицы гемоглобина (Б).
Укладка этих
двух
цепей практически одинакова. Основные различия сосре-
доточены у N- и С-концов цепей и у контактирующих поверхностей субъеди-
ницы
гемоглобина. ГС изменениями из книги Dickerson R. E., The Proteins
(ed. H. Neurath), Academic Press
(1964).]
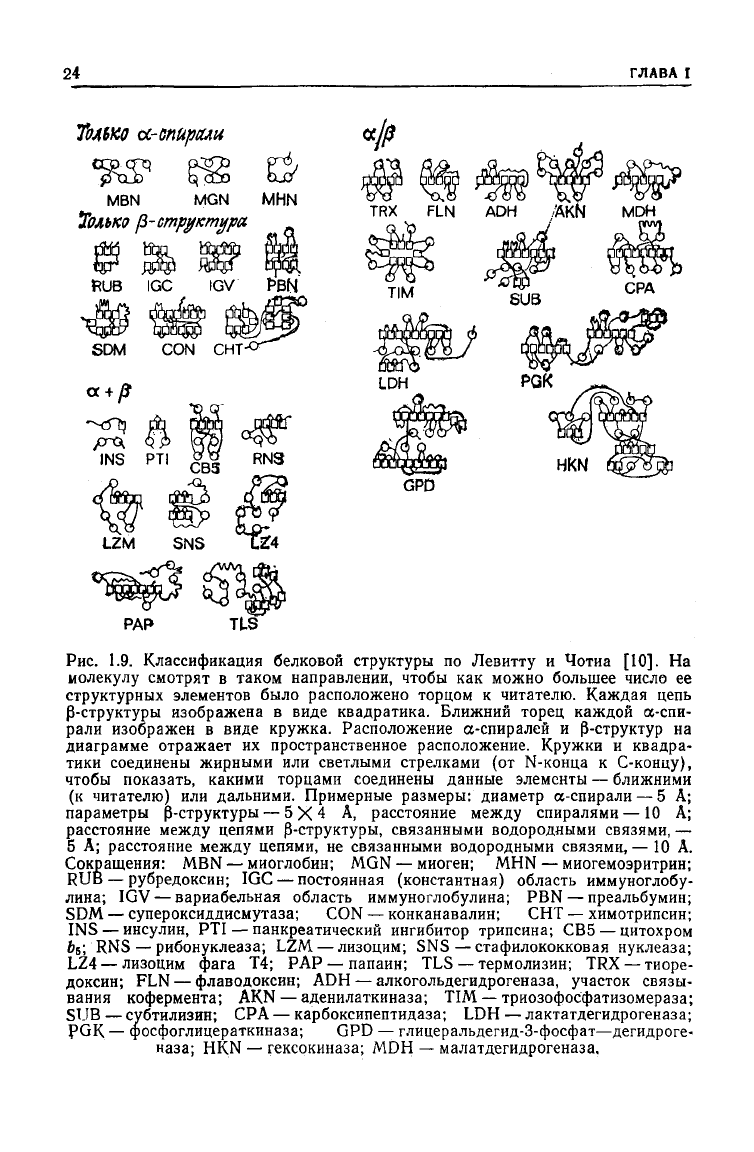
24
ГЛАВА
I
Только
ос-спирали
pXLC
Ц ССО
MBN
MGN MHN
только
/3-структура
TRX FLN ADH /AKN MDH
RUB
IGC IGV
F>BN
GPD
PAP
Рис.
1.9. Классификация белковой
структуры
по
Левитту
и Чотиа [10]. На
молекулу
смотрят в таком направлении, чтобы как можно большее число ее
структурных
элементов было расположено торцом к читателю. Каждая цепь
Р-структуры
изображена в виде квадратика. Ближний торец каждой а-спи-
рали изображен в виде кружка. Расположение а-спиралей и
Р-структур
на
диаграмме отражает их пространственное расположение. Кружки и квадра-
тики
соединены жирными или светлыми стрелками (от N-конца к С-концу),
чтобы показать, какими торцами соединены данные элементы — ближними
(к
читателю) или дальними. Примерные размеры: диаметр а-спирали — 5 А;
параметры
Р-структуры
— 5X4 А, расстояние
между
спиралями —10 А;
расстояние
между
цепями р-структуры, связанными водородными связями, —
5 А; расстояние
между
цепями, не связанными водородными связями,— 10 А.
Сокращения:
MBN — миоглобин; MGN — миоген; MHN — миогемоэритрин;
RUB
— рубредоксин; IGC — постоянная (константная) область иммуноглобу-
лина;
IGV— вариабельная область иммуноглобулина; PBN — преальбумин;
SDM
— супероксиддисмутаза; CON — конканавалин; СНТ — химотрипсин;
INS
— инсулин, PTI — панкреатический ингибитор трипсина; СВ5 — цитохром
Ьъ\
RNS—
рибонуклеаза; LZM— лизоцим; SNS — стафилококковая нуклеаза;
LZ4 — лизоцим фага Т4; РАР — папаин; TLS — термолизин; TRX — тиоре-
доксин;
FLN — флаводоксин; ADH — алкогольдегидрогеназа, участок связы-
вания
кофермента; AKN — аденилаткиназа; TIM — триозофосфатизомераза;
SUB — субтилизин; СРА — карбоксипептидаза; LDH — лактатдегидрогеназа;
PGK
— фосфоглицераткиназа; GPD — глицеральдегид-3-фосфат—дегидроге-
наза; HKN — гексокиназа; MDH — малатдегидрогеназа.

ТРЕХМЕРНАЯ
СТРУКТУРА
ФЕРМЕНТОВ
25
туго
скрученной спирали (3,55 остатка на виток) этот
угол
уменьшается до 10°. Как отмечал Чотиа [13], р-структуры все-
гда изогнуты; такая изогнутая р-структура энергетически более
выгодна, чем планарная (рис.
1.10).
Белки
— это не рыхлые, случайным образом сформирован-
ные
структуры. Их аминокислотные остатки упакованы так же
плотно,
как и аминокислоты
в
кристаллах [14—16].
Плотность
упаковки (часть
пространства, занимаемого
атомами) для белков со-
ставляет примерно 0,75, а
для кристаллов эта величи-
на
равна
0,7—0,78.
Макси-
мальная плотность упаков-
ки
сферических частиц рав-
на
0,74, а цилиндрических
частиц неограниченной дли-
ны—0,91.
4. Третичная
и
четвертичная
структура
Рис.
1.10. Структура части молекулы
глицеральдегид-3-фосфат — дегидрогена-
зы
из В.
stearothermophilus;
цилиндры —
сс-спирали,
изогнутые стрелки — (5-слои.
(С
любезного разрешения
Biesecker
G. и
Wonacott A.)
Трехмерную
структуру
белка, состоящего из един-
ственной
полипептидной це-
пи
или из ковалентно свя-
занных цепей, называют его
третичной структурой. Мно-
гие белки состоят из
субъ-
единиц,
не связанных
друг
с
другом
ковалентно. Пространствен-
ная
организация такого комплекса известна под названием чет'
вертичной
структуры. Трехмерная
структура
каждой субъеди-
ницы
по-прежнему называется ее третичной структурой. Изме-
нение
четвертичной структуры означает, что субъединицы сме-
стились
друг
относительно
друга.
Субъединицы
могут
быть образованы как идентичными по-
липептидными цепями, так и различающимися. Как правило,
области контакта субъединиц упакованы так же плотно, как и
внутренняя
часть белков, а расположенные на поверхности
группы и ионы образуют ионные и водородные связи [17].
Изменение
третичной структуры гемоглобина сопровождается
смещением плотно упакованных субъединиц
друг
относительно
Друга.

26
ГЛАВА
1
5. Одинакова ли
структура
фермента в растворе
и
в кристалле?
Этот вопрос является как нельзя более уместным, поскольку
все данные о
структурных
особенностях ферментов и механизме
их действия были получены при исследовании кристаллических
белков. Когда-то ответить на поставленный выше вопрос было
довольно трудно, но имеющиеся в настоящее время данные, как
правило,
дают
положительный
ответ.
Сомнения остаются лишь
относительно активного центра карбоксипептидазы (гл. 12), да
и
то только по поводу подвижной боковой цепи остатка тирози-
на.
Идентичность
структуры
ферментов в растворе и в кристал-
лах подтверждается следующими данными. 1. Несмотря на то
что некоторые ферменты были получены в кристаллическом виде
из
разных растворителей и в различных формах, их
структура
оставалась практически одинаковой (например, субтилизин [18,
19] и лизоцим [20]). 2. Ферменты, принадлежащие к одному
классу, имеют
сходную
структуру
(например, сериновые про-
теазы, см. ниже). 3. При образовании димера а-химотрипсина
контакт
между
мономерами осуществляется одними и теми же
участками как в растворе, так и в составе кристалла [21, 22].
4. Иногда ферменты сохраняют свою активность в кристалличе-
ском
состоянии (например, рибонуклеаза [23, 24] и карбокси-
пептидаза А [25]). Сопоставление активности фермента в рас-
творе и в кристаллическом состоянии является непростой
задачей, поскольку диффузия
субстратов
внутрь кристаллов за-
труднена. Имеется, правда, один
путь
остроумного решения этой
проблемы, состоящий в кристаллизации связанного с ферментом
промежуточного соединения (индолилакрилоилхимотрипсина;
см.
ниже разд. Г. 2) при таком значении рН, когда это проме-
жуточное
соединение устойчиво [26]. При изменении рН, при-
водящем к увеличению реакционной способности промежуточ-
ного соединения, оно гидролизуется, причем реакция характери-
зуется
той же самой константой скорости первого порядка, что
и
в растворе.
Однако если в растворе в равновесии фермент находится в
нескольких разных конформациях, то при кристаллизации
будет
отбираться только одна из этих конформаций. Так, в условиях,
при
которых проводится кристаллизация, значительная часть
а-химотрипсина находится в неактивной конформаций [27],
кристаллизуется же только активная форма фермента.

ТРЕХМЕРНАЯ СТРУКТУРА ФЕРМЕНТОВ 27
В. Классы ферментов
1. Сериновые протеазы
[28—34]
Волнение, вызванное выявлением
того
факта, что белки, свя-
зывающие кислород,— гемоглобин и миоглобин
—•
имеют одина-
ковую третичную
структуру
и выполняют одинаковые функции,
вновь овладело учеными, когда было установлено, что анало-
гичная ситуация имеет место в
случае
сериновых протеаз
млекопитающих. Эти ферменты названы так потому, что они
имеют уникальный по своей активности сериновый остаток, ко-
торый необратимо
реагирует
с фосфорорганическими соедине-
ниями,
например с диизопропилфторфосфатом. Основные пан-
креатические ферменты — трипсин, химотрипсин и эластаза —
кинетически весьма близки и
гидролизуют
пептиды и синтети-
ческие сложные эфиры. Их активность имеет оптимум при
рН
=; 7,8 и определяется состоянием ионизации групп с р/(
а
=
=
6,8. Во
всех
трех
случаях
в процессе реакции образуется
«ацилфермент», в котором карбоксильный фрагмент
субстрата
образует
сложноэфирную связь с гидроксильной группой актив-
ного серина.
Основное различие
между
трипсином, химотрипсином и эла-
стазой состоит в их специфичности. Трипсин специфически гид-
ролизует пептиды, состоящие из лизина и аргинина, и эфиры
этих аминокислот; химотрипсин расщепляет полипептидную
цепь по фенилаланину, тирозину и триптофану, имеющим боль-
шие гидрофобные боковые цепи; специфичность эластазы про-
является в ее действии на такие небольшие гидрофобные моле-
кулы, как аланин. Установление
структуры
кристаллических
ферментов показало, что полипептидные остовы
всех
трех
фер-
ментов при наложении их
друг
на
друга
практически совмеща-
ются, за исключением участков, где добавлено или пропущено
несколько аминокислот (рис.
1.11).
Различие же в специфично-
сти этих ферментов обусловлено небольшими изменениями в
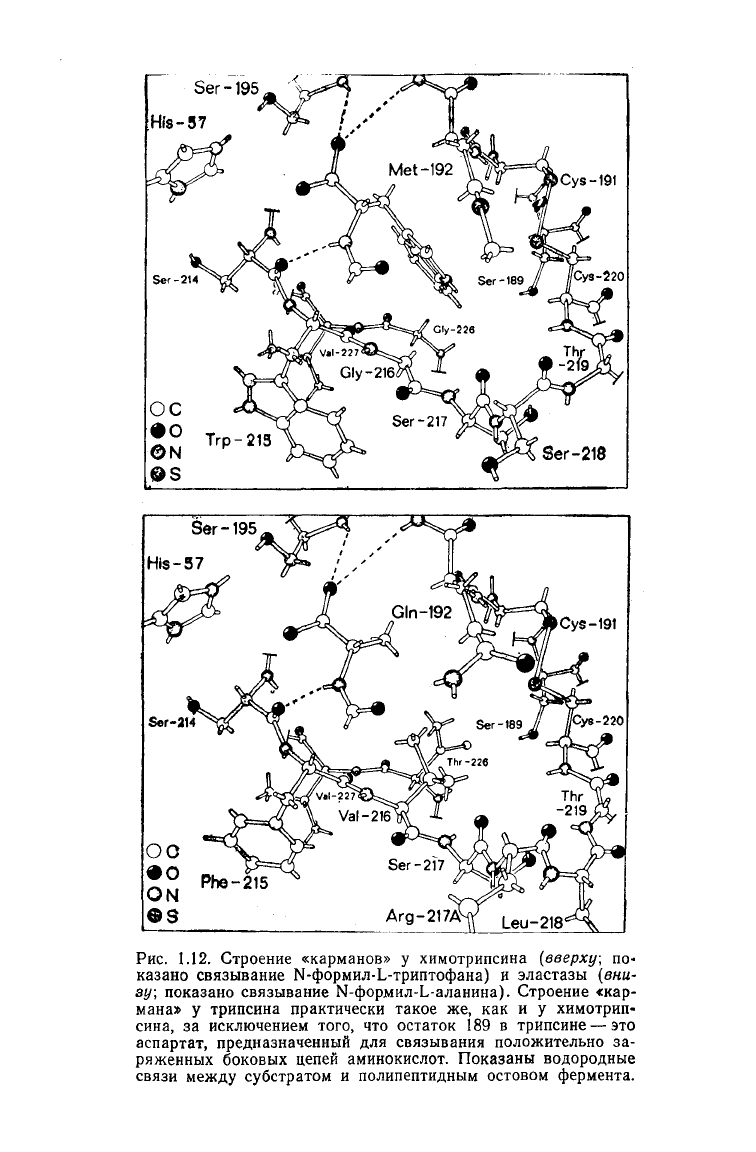
строении «кармана», связывающего боковую цепь аминокисло-
ты. В молекуле химотрипсина имеется четко выраженный кар-
ман,
связывающий большие гидрофобные боковые цепи [35].
В молекуле трипсина на дне аналогичного кармана вместо
Ser-189
находится аспартат [36]. Отрицательно заряженная
карбоксильная группа
Asp-189
образует
ионную связь с поло-
жительно заряженной аммонийной или гуанидиниевой группами
на
конце цепи лизина или аргинина. Два глицина, расположен-
ные у
входа
в карман химотрипсина, в
случае
эластазы замеще-
ны
валином
(Val-216)
и треонином (Thr-226) [37]. Это предот-
вращает проникновение в карман больших боковых цепей и обе-
спечивает связывание небольшой по размерам боковой цепи
аланина (рис.
1.12).
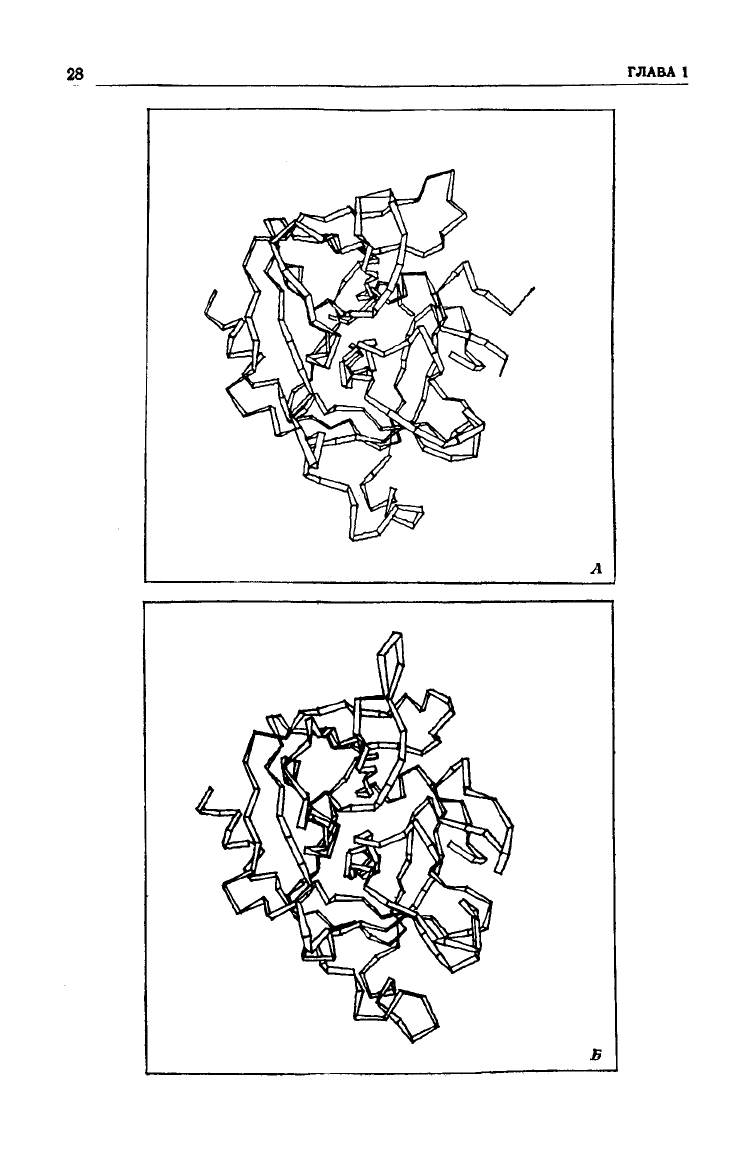
28
ГЛАВА
1
ТРЕХМЕРНАЯ
СТРУКТУРА
ФЕРМЕНТОВ 29
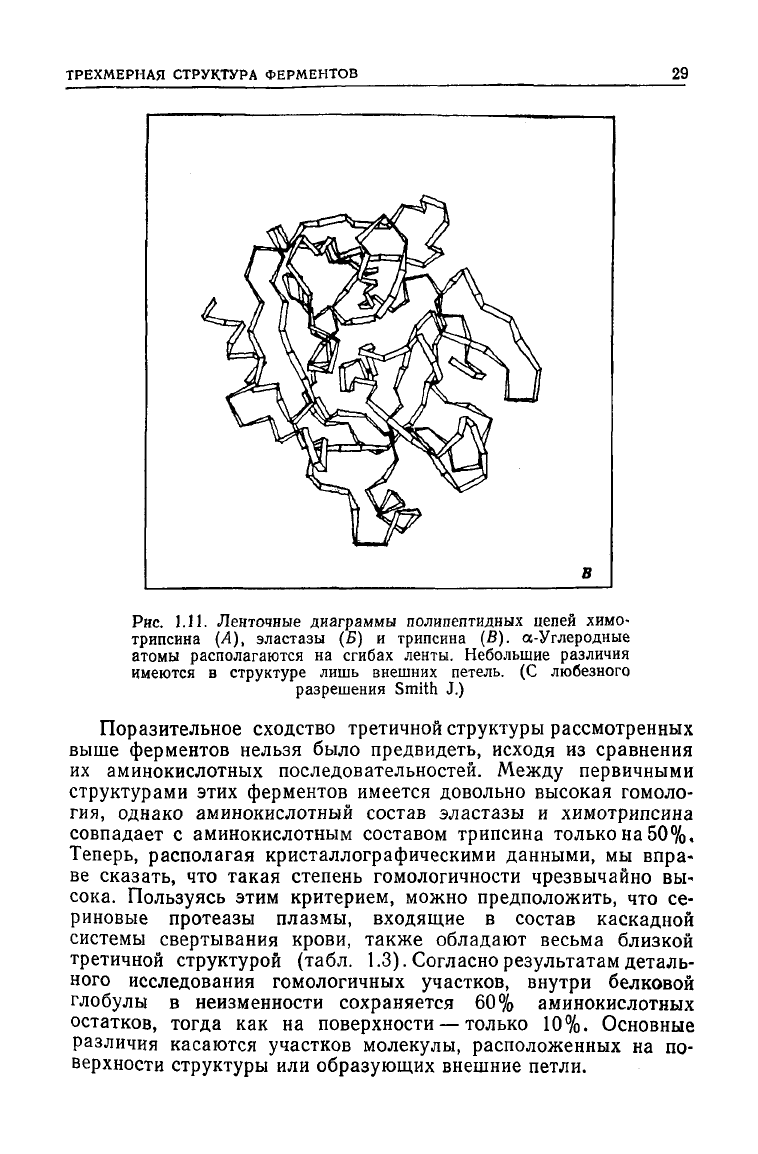
Рис.
1.11. Ленточные диаграммы полипептидных цепей химо-
трипсина
(А), эластазы (Б) и трипсина (В). ос-Углеродные
атомы располагаются на сгибах ленты. Небольшие различия
имеются в
структуре
лишь внешних петель. (С любезного
разрешения Smith J.)
Поразительное
сходство
третичной структуры рассмотренных
выше ферментов нельзя было предвидеть, исходя из сравнения
их аминокислотных последовательностей. Между первичными
структурами этих ферментов имеется довольно высокая гомоло-
гия,
однако аминокислотный состав эластазы и химотрипсина
совпадает с аминокислотным составом трипсина только на 50%,
Теперь, располагая кристаллографическими данными, мы впра-
ве сказать, что такая степень гомологичности чрезвычайно вы-
сока.
Пользуясь этим критерием, можно предположить, что се-
риновые протеазы плазмы, входящие в состав каскадной
системы свертывания крови, также обладают весьма близкой
третичной структурой (табл. 1.3). Согласно результатам деталь-
ного исследования гомологичных участков, внутри белковой
глобулы в неизменности сохраняется 60% аминокислотных
остатков,
тогда
как на поверхности — только 10%. Основные
различия
касаются участков молекулы, расположенных на по-
верхности структуры или образующих внешние петли.
Ser-195
His-97
Ser-218
Cys-191
Рис.
1.12. Строение «карманов» у химотрипсина
(вверху;
по-
казано
связывание Ы-формил-Ь-триптофана) и эластазы (вни-
зу; показано связывание Ы-формил-Ь-аланина). Строение «кар-
мана» у трипсина практически такое же, как и у химотрип-
сина,
за исключением того, что остаток 189 в трипсине — это
аспартат, предназначенный для связывания положительно за-
ряженных боковых цепей аминокислот. Показаны водородные
связи
между
субстратом и полипептидным остовом фермента.