Elsevier Encyclopedia of Geology - vol I A-E
Подождите немного. Документ загружается.

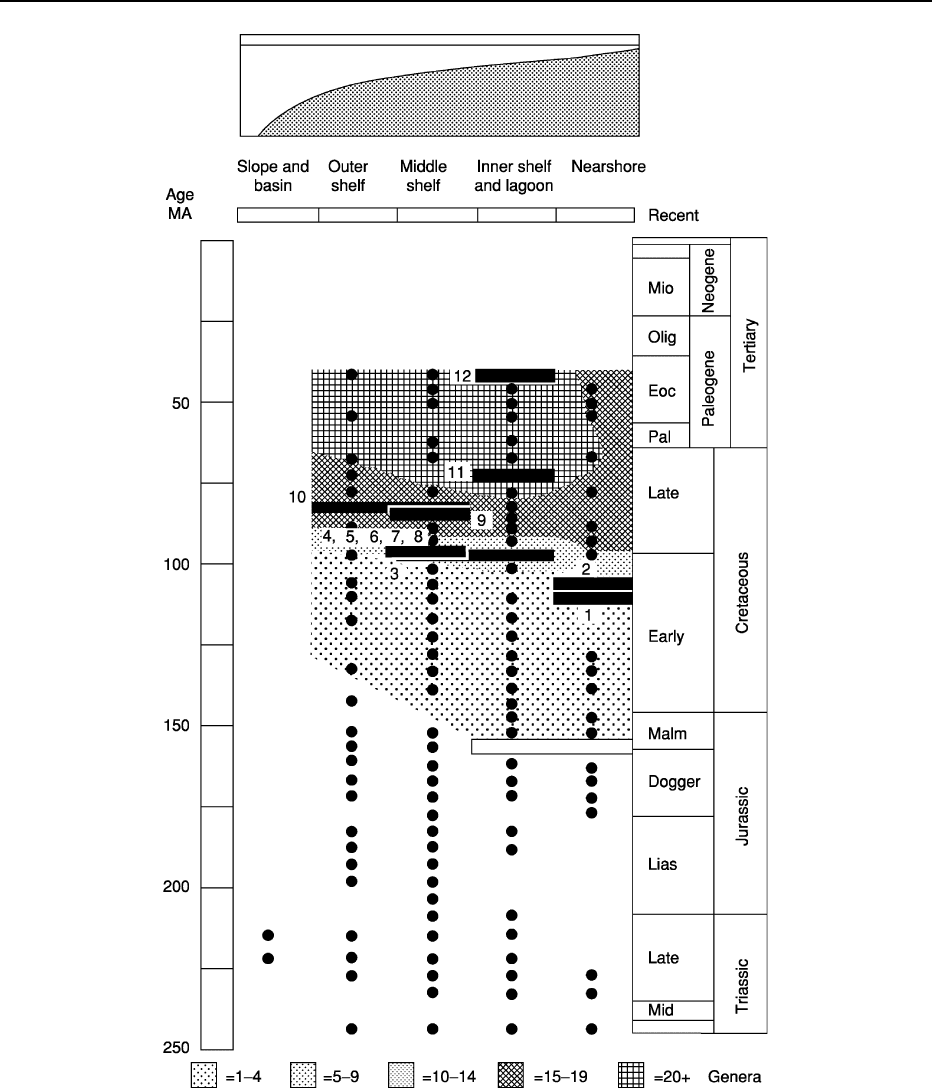
Figure 6 Study exemplifying that the early phases of a history of marine invertebrate clades began in nearshore environments and
subsequently expanded to the outer shelf. In this study the environmental history of the cheilostome bryozoan radiation suggests that
the group originated in nearshore and inner shelf environments (white box). As the group diversified in both numbers of genera
(contours denoted by the different shadings) with the acquisition of evolutionary novelties (numbers associated with black boxes) they
spread to middle and outer shelf habitats. The black dots represent localities yielding bryozoans at different horizons and depositional
environments. Those dots below the shaded areas represent localities at which only non-cheilostomes are found (i.e., cyclostomes
and trepostomes) and these impose a taphonomic control documenting that the cheilostome pattern is not an artifact of preservation.
The dots within the shaded area represent localities at which all kinds of bryozoan are found. Reproduced from Jablonski D, Lidgard
S and Taylor PD (1997) Comparative ecology of bryozoan radiations: origin of novelties in cyclostomes and cheilostomes.
Palaios 12:
505–523. SEPM (Society for Sedimentary Geology).
272 BIOLOGICAL RADIATIONS AND SPECIATION

evolution of cheilostome bryozoans in the Cretaceous
(see Fossil Invertebrates: Bryozoans). It needs to be
emphasized that this is a general pattern; there is no
implication that the ecological shift was the cause
of speciation.
Abiotic Causes – Fragmentation of Areas
Radiations coincident with fragmentation of contin-
ental masses can be documented for a variety of
organisms, but usually only over long timescales.
One example, is the increased diversity of families
of terrestrial amniotes between the Lower Jurassic
and Paleocene following the breakup of Pangaea
(Figure 7). It is very difficult to demonstrate a direct
cause and effect, but there is a considerable shift in the
latitudinal distribution of the continental masses
such that substantial diversification in ecological
conditions occurred leading to a tempting association
with cladogenesis. The breakup of Pangaea also re-
sulted in a considerable increase in available coastline,
Figure 7 Radiations associated with fragmentation of continents. The breakup of Pangaea was associated with increasing provin-
ciality of terrestrial tetrapod families, plotted here from the Middle Jurassic to the Paleocene. This plot shows the percentage of
families of terrestrial tetrapods that occupied one, two and three of the modern continental areas. Data given in table above. The plot
shows that the percentage of families occupying only one of the modern continental areas rose from 8% of the families originating in
the Middle Jurassic to 66% of those originating in the Paleocene. Conversely, 67% of families originating in the Middle Jurassic
occupied three of the modern continental areas and this decreased to just 3% in the Paleocene. Reproduced from Benton MJ (1985)
Patterns of diversification of Mesozoic non-marine tetrapods and problems of historical diversity analysis.
Special Papers in Palaeon-
tology
33: 185–202. Published by permission of the Palaeontological Association.
BIOLOGICAL RADIATIONS AND SPECIATION 273
along with environmental differentiation, and this
almost certainly influenced the radiation of many
marine organisms.
Origination of an Evolutionary Novelty Leading to
Taxic Diversity
Often the radiation of a particular group has been
linked with the appearance of a particularly significant
character (e.g., development of the cleidoic egg, en-
abling the organisms to become reproductively inde-
pendent of water, or wings to fly). In many of the
usual examples there is very little taphonomic con-
trol, so that it is difficult to separate the appearance of
an evolutionary novelty and increase in species diver-
sity from the preservation bias that may be present in
the fossil record. But there are some examples that
avoid this problem. Cheilostomes are the dominant
bryozoans (see Fossil Invertebrates: Bryozoans) living
today and they are distinguished from other Bryozoa
by the possession of box-shaped zooecia. The fossil
record of cheilostomes begins in the Upper Jurassic,
with a rapid rise in numbers of species and genera in
late Lower Cretaceous. There are two kinds of chei-
lostomes: malacostegans (about 150 Recent species)
and their species descendents, the neocheilostomes
(about 5000 Recent species). Modern malacostegans
release planktotrophic larvae that spend up to about
1 month free living in the water column before set-
tling and beginning colony growth. During this time
there is potential for considerable dispersal as well
as associated potential for continued gene flow.
Neocheilostomes brood their larvae within a special-
ized brood chamber: – the ovicell. The larvae have a
very short, free life span before settling to the sub-
strate (often adjacent to the parent) and beginning
colony formation. Thus, gene flow between popula-
tions may be very restricted and as a consequence
there is the greater potential for genetic divergence
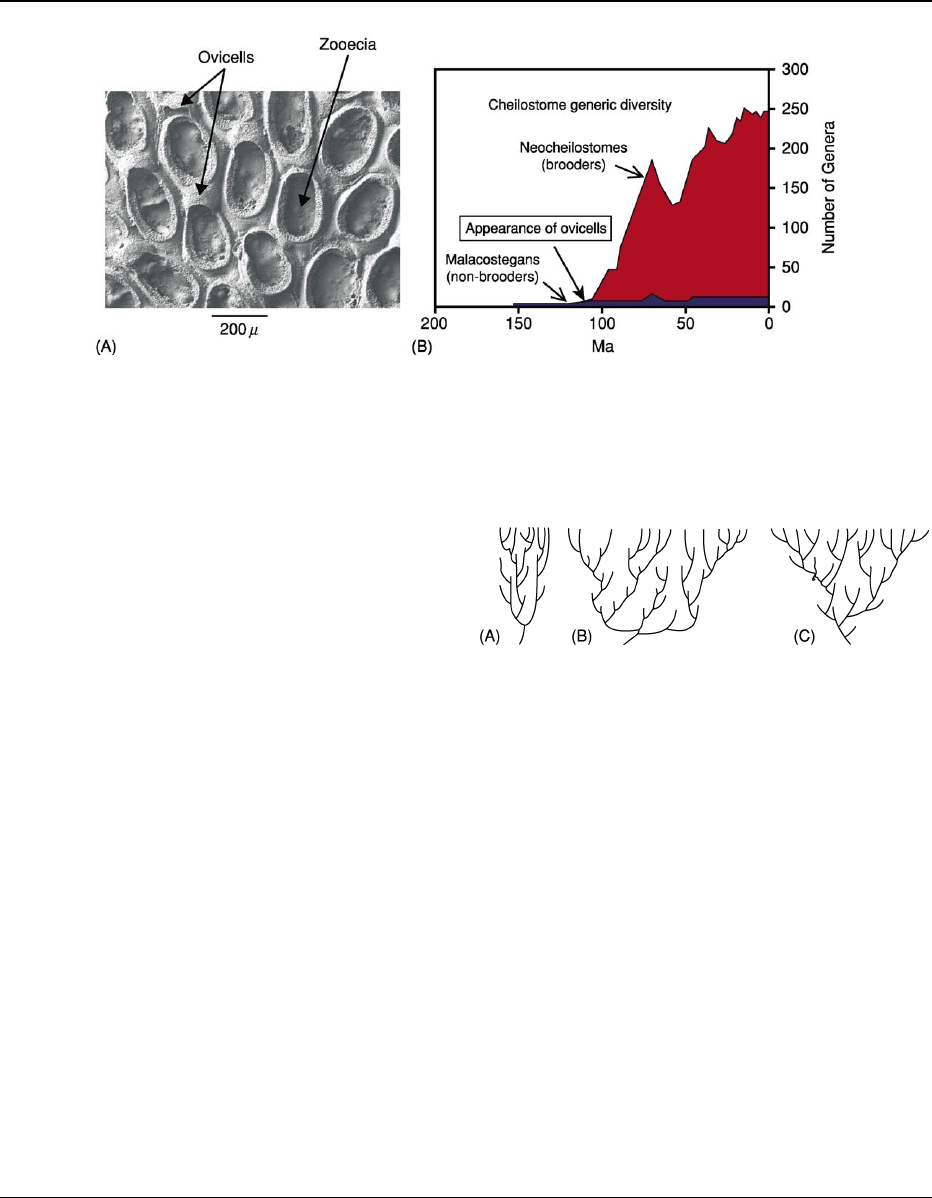
Figure 8 The cause of a clade radiation has often been identified as the evolution of a key innovation. One such example is the
appearance of ovicells (brood chambers) in some cheilostome bryozoans. (A) Welbertopora mutabilis, upper Albian, Fort Worth
Formation, Denton County, Texas. Part of the colony of one of the earliest neocheilostomes with ovicells in which larva develop.
(B) Generic plots of non-brooding and brooding cheilostomes showing the dramatic rise in the latter. The presence of Jurassic and
Lower Cretaceous cheilostomes demonstrates that appearance of brooders is not a taphonomic artifact. Images supplied by Dr Paul
Taylor, The Natural History Museum, from unpublished data.
Figure 9 The history of a clade during phylogenetic diversifi-
cation may follow one of several patterns, idealized here. The
horizontal axis represents morphological divergence, the verti-
cal axis time. (A). Morphological evolution constrained, species
origination high. (B) Morphological evolution rapid in early
periods relative to speciation. (C) Morphological evolution and
speciation approximately in step with one another. From Foote M
(1993).
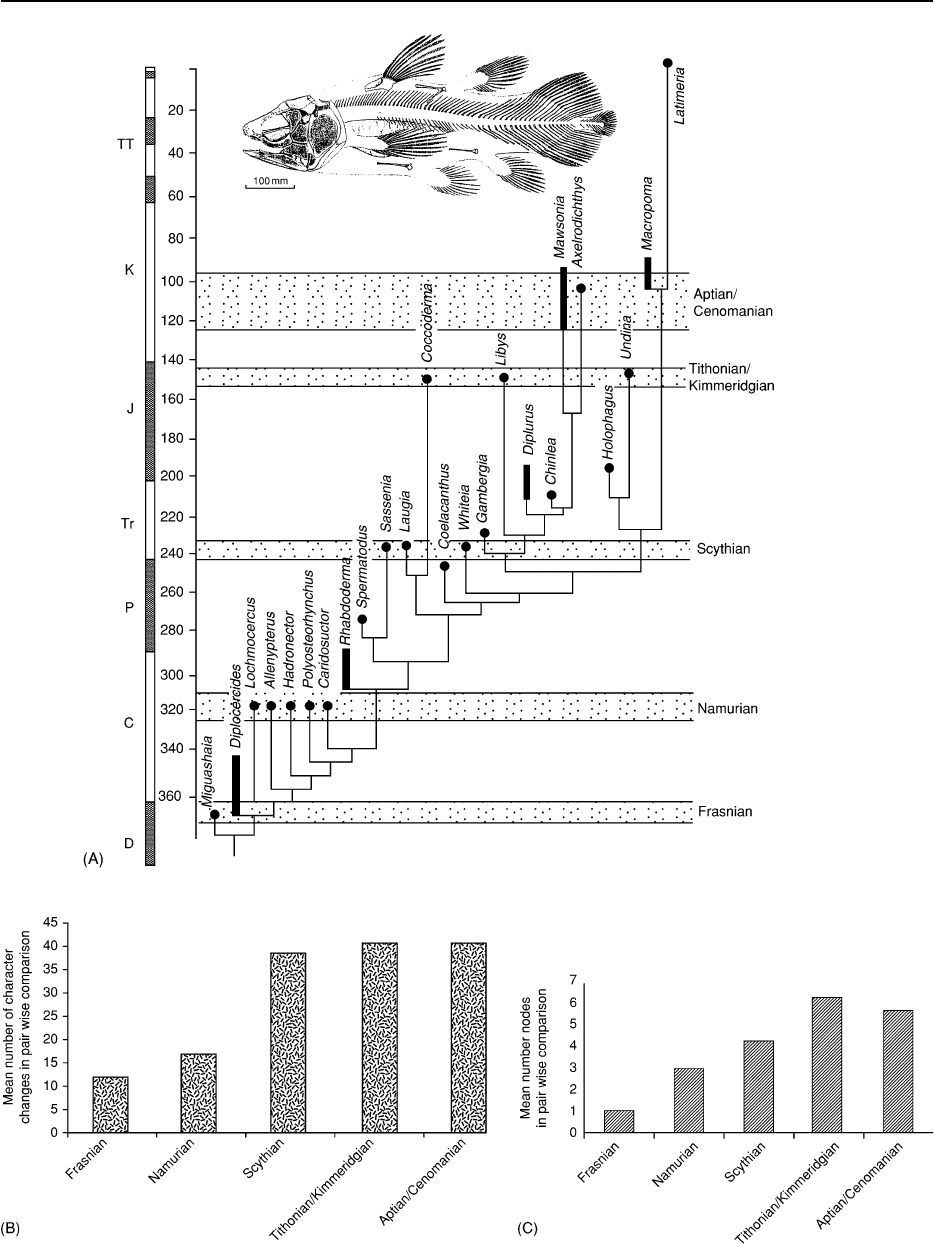
Figure 10 Comparison of morphological and taxic evolution in coelacanth fishes using phylogenetic measures. (A) Phylogenetic
tree showing 5 time bands selected as sample stages. Within each time band two types of comparison were made on a pair wise basis
and then averaged. (B) The mean number of character changes occurring between taxa in each of the five time bands. This is a
measure of morphological disparity. (C) The mean number of nodes (cladogenetic events) occurring between taxa in each of
the five time bands. This is a measure of taxic diversity. In this case morphological diversity tracks taxic diversity except for a slight
insignificant increase in taxic diversity relative to morphological diversity in the early stage (Namurian). Data from Forey PL (1998)
History of the coelacanth fishes. London: Chapman & Hall.
274 BIOLOGICAL RADIATIONS AND SPECIATION
BIOLOGICAL RADIATIONS AND SPECIATION 275
and, eventually, speciation. The ovicells and hence
brooding of larvae can be identified in the skeleton
of the neocheilostomes (Figure 8A). Plots at the gen-
eric-level of malacostegans and neocheilostomes
show little change in the former, but a dramatic rise
in the latter (Figure 8B), suggesting strongly that the
acquisition of brooding is responsible for the radi-
ation. It is pertinent to note that very similar patterns
involving similar contrasting reproductive strategies
have been observed in turritellid gastropod evolution.
Morphological and Taxic Evolution During
Radiations
Radiations involve particular patterns of morpho-
logical as well as taxic evolution. Figure 9 shows
three types of relationship between morphological
and taxic evolution during clade history. Studies
examining the relationship between these two aspects
may be phylogeny dependent or independent, and
several different methods have been used to analyze
both. Much literature suggests that the pattern in
Figure 9B is common.
One study of the radiation of coelacanth fishes
(see Fossil Vertebrates: Fish) used a strictly phylogen-
etic approach for both measures (Figure 10). The
phylogeny of 24 genera was sampled at five time
intervals, which collectively spanned the Upper Dev-
onian to Upper Cretaceous. Therefore, this study
dealt with a radiation that was long term and in-
volved few taxa and, while demonstrating a method,
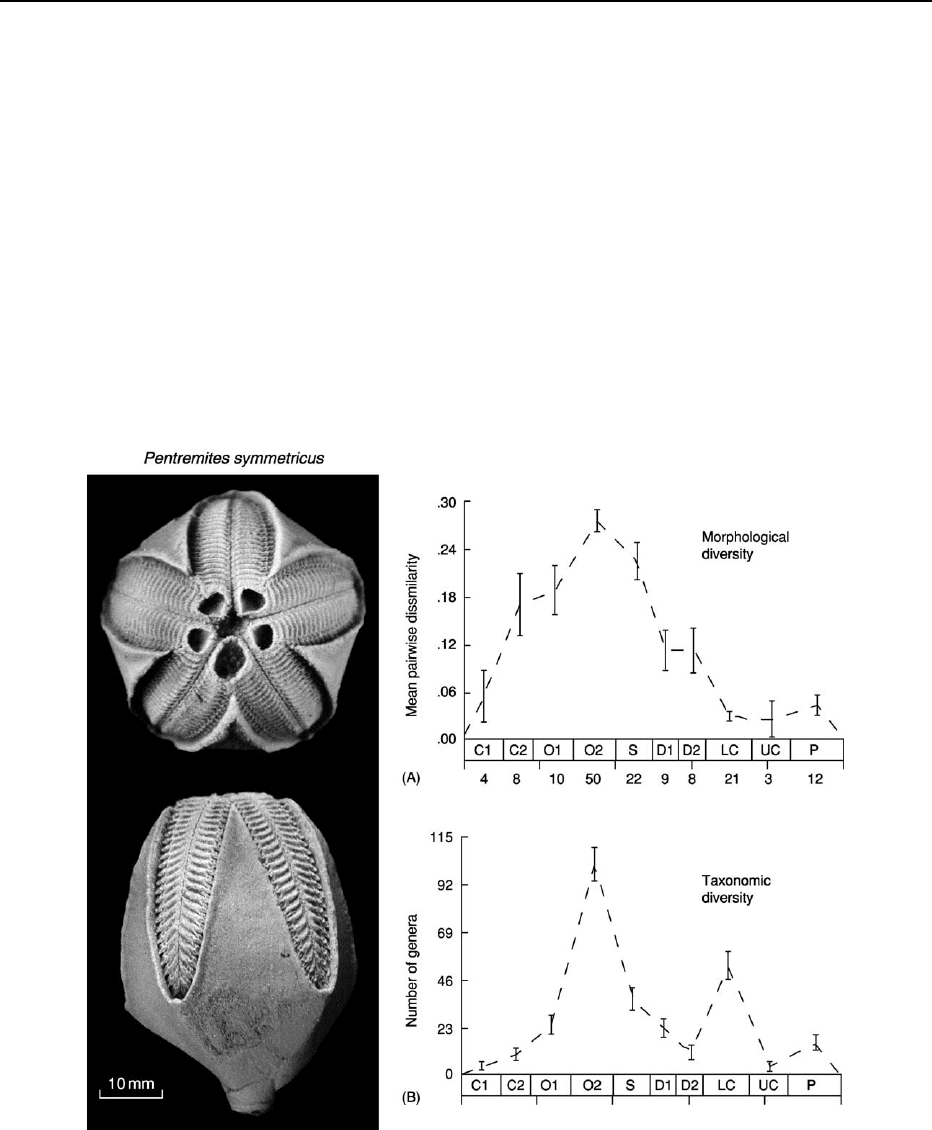
Figure 11 Comparison of morphological and taxic evolution in Blastozoa echinoderms using non-phylogenetic measures taken at
approximately equal time slices from Lower Cambrian to Permian. (A) Mean pair wise dissimilarity, calculated by comparing the
number of dissimilar codes for 65 characters between each pair and averaging over the number of comparisons made. The number of
genera (one species of each was examined) is given against each time slice examined. This gave an average measure of
morphological disparity. (B) Number of genera at each time slice as a measure of taxic diversity. This study showed a rapid burst
of morphological evolution that was not accompanied by taxic evolution in the early stages of clade history (C2 and O1). Note that
quite the reverse trend was evident later in clade history (LC). Time slice abbreviations: C1 – Lower Cambrian, C2 – Middle and
Upper Cambrian, O1 – Lower Ordovician, O2 – Middle and Upper Ordovician, S – Silurian, D1 – Lower Devonian, D2 – Middle and Upper
Devonian, LC – Lower Carboniferous, UC – Upper Carboniferous, P – Permian. (After Foote 1992, with permission of author).
Photograph supplied by Dr AB Smith.
276 BIOLOGICAL RADIATIONS AND SPECIATION
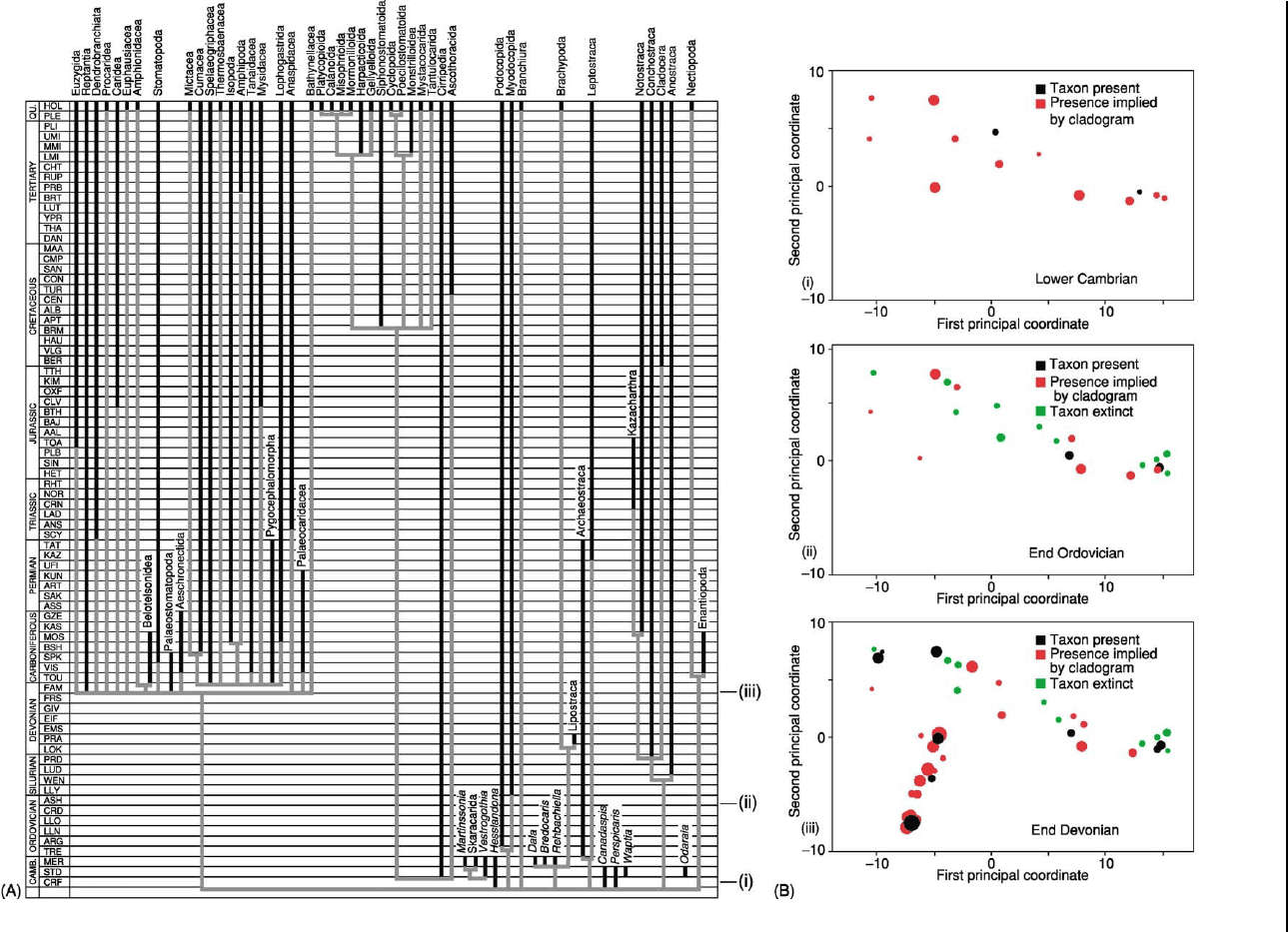
Figure 12 (Continued)
BIOLOGICAL RADIATIONS AND SPECIATION 277

may not be typical. There was very little taphonomic
control in this study. At each interval the distance, as
measured by number of character changes along the
inferred lineage between terminal taxa, was averaged
over all pair wise comparisons. This gave an average
morphological distance for successive time periods in
the clade’s history. The values for successive time
periods could then be compared to each other and,
in turn, compared to the number of cladogenetic
events within each time period (which provided a
measurement of taxic diversity). In this case the
taxic diversity tracked morphological diversity except
for the early stages where taxic diversity slightly
exceeded morphological diversity.
Some other studies have not considered the phylo-
genetic relationships within clades (although mono-
phyly of the entire clade has been accepted). Figure 11
shows the results of such a study of the radiation of
members of the echinoderm subphylum Blastozoa
(see Fossil Invertebrates: Echinoderms (Other Than
Echinoids)). In this study, one species each from
147 genera, ranging in time from the Lower Cam-
brian to Permian, were examined for 65 morpho-
logical characters. Ten stratigraphic intervals were
chosen and within each interval pair wise compari-
sons between characters coded for each of the genera
were made and averaged. The phylogeny was not
considered here. One common belief is that morpho-
logical diversity is disproportionately high compared
to taxic diversity at the beginning of a radiation
and that such a relationship is reversed later on. The
blastozoan study appears to confirm this since the
morphological diversity was far greater than
the taxic diversity in the mid-Cambrian to Lower
Ordovician; that is, in the early stages of clade
history. The reverse relationship was true in the
Lower Carboniferous.
The Cambrian explosion must be the most spec-
tacular of all radiations that the planet has witnessed.
Yet it remains one of the most enigmatic. Within
15 million years of the beginning of the Cambrian
period, where tiny shelly faunas are first detected, a
wide variety of skeletonized animals appeared, al-
ready well diversified and exquisitely adapted. Today
they are classified amongst the major invertebrate
phyla. Molecular evidence, alongside phylogenetic
studies on the first appearing fossils all point to there
being a long Precambrian history to many lineages.
But exactly what form this history took is very uncer-
tain. Steven Jay Gould, in his book Wonderful Life,
– suggested that when skeletonized animals first
appeared in abundance in the Cambrian there was a
remarkably diverse array of body forms and that
many of these were weeded out by extinctions leading
to differential survival of a few body forms that
eventually gave rise to our modern fauna. In other
words, morphological disparity was greatest in the
early phase of the animal lineage history and gradually
became canalized. On a broad scale this theory echoed
the radiation pattern shown by the blastozoan echino-
derms mentioned above. Gould’s thesis was grounded
in a large part on the arthropods, and especially the
crustacean-like animals, conspicuous in the famous
Burgess Shale Lagersta
¨
tten, but was largely anecdotal,
with little phylogenetic evidence or any precise meas-
ures of morphological disparity applied. More
detailed studies comparing levels of disparity in Cam-
brian and Recent arthropods (see Fossil Invertebrates:
Arthropods) showed that there was very little differ-
ence between then and now. More specifically detailed
studies of the early evolution of the Crustacea have
shown that morphological disparity grew in concert
with taxic diversity (Figure 12).
The study of the pattern of radiations in the fossil
record has implications for our understanding of post-
extinction recovery and potentially for our efforts to
put conservation efforts in place. If there are general
patterns that emerge that can be associated with
Figure 12 Morphological disparity of crustaceans tracked through time against a phylogeny. In this study one exemplar taxon from
each of the crustacean orders or suborders was coded for 135 characters. (A) A cladistic analysis plotted against stratigraphy resulted
in the consensus tree shown here. The actual stratigraphic ranges are shown in black and those which must be assumed because
of the shape of the tree in grey (ghost ranges). Notice that most of the species of modern crustacea did not appear until late Palaeozoic.
(B) Plots showing disparity of the clades present through three time slices. The disparity is measured by making pair wise
comparisons between the coded characters for each of the taxa present in any one time interval (the ghost range taxa were coded
according to reconstructed ancestral states). Some differential weights were applied according to the particular type of character and
in this it differed from the Blastozoan study (Figure 11). A principal coordinate analysis was then carried out to illustrate the difference
in morphospace between the taxa. In the diagrams the first three principal coordinates are shown, the third coordinate being at right
angles to the paper (filled symbols positive and open symbols negative). The black symbols represent taxa with a range extending
through the stage illustrated, red symbols represent taxa implied by ghost ranges and the green symbols are taxa extinct at stage
illustrated. This diagram shows that both morphological disparity and taxic diversity increased from the Cambrian to Upper Devonian
at which time a new area of morphospace (bottom left) was occupied. Reproduced from Wills MA (1998) Crustacean disparity through
the Phanerozoic: comparing morphological and stratigraphic data.
Biological Journal of the Linnean Society of London 65: 455–500.
278 BIOLOGICAL RADIATIONS AND SPECIATION

specific ecological conditions, then some predictions
may be made as to where and what to protect.
See Also
Evolution. Fossil Invertebrates: Arthropods; Bryo-
zoans; Echinoderms (Other Than Echinoids); Gastropods.
Fossil Vertebrates: Fish.
Further Reading
Benton MJ (1985) Patterns of diversification of Mesozoic
non-marine tetrapods and problems of historical diversity
analysis. Special Papers in Palaeontology 33: 185–202.
Cheetham AH (1986) Tempo of evolution in a Neogene
bryozoan: rates of morphological change within and
across species boundaries. Paleobiology 12: 190–202.
Erwin DH and Anstey RL (1995) Introduction. In: Erwin
DH and Anstey RL (eds.) New approaches to speciation
in the fossil record, pp. 1–10. New York: Columbia Uni-
versity Press.
Foote M (1992) Paleozoic record of morphological diver-
sity in blastozoan echinoderms. Proceedings of the Acad-
emy of Natural Sciences 89: 7325–7329.
Foote M (1993) Discordance and correspondence between
morphological and taxonomic diversity. Paleobiology
19: 185–204.
Forey PL (1998) History of the coelacanth fishes. London:
Chapman & Hall.
Jablonski D, Lidgard S, and Taylor PD (1997) Comparative
ecology of bryozoan radiations: origin of novelties in
cyclostomes and cheilostomes. Palaios 12: 505–523.
Lazarus DB (2001) Speciation and morphological evolu-
tion. In: Briggs DE and Crowther PR (eds.) Palaeobiology
II, pp. 133–137. Oxford: Blackwell Scientific.
Levington JS (2001) Genetics, Palaeontology and Macro-
evolution, 2nd edn. p. 617. Cambridge: Cambridge
University Press.
Nehm RH and Geary DH (1994) A gradual morphologic
transition during a rapid speciation event in marginellid
gastropods (Neogene; Dominican Republic). Journal of
Paleontology 68: 787–795.
Schluter D (2000) The ecology of adaptive radiation.
Oxford: Oxford University Press.
Stanley SM (1979) Macroevolution: pattern and process.
San Francisco: WH. Freeman.
Taylor PD and Larwood GP (eds.) (1990) Major evolution-
ary radiations. Oxford: Clarendon Press.
Wheeler QD and Platnick NI (eds.) (2000) Species concepts
and phylogenetic theory. New York: Columbia Univer-
sity Press.
Wills MA (1998) Crustacean disparity through the Phaner-
ozoic: comparing morphological and stratigraphic data.
Biological Journal of the Linnean Society of London 65:
455–500.
BIOSEDIMENTS AND BIOFILMS
M R Walter and A C Allwood, Macquarie University,
Sydney, NSW, Australia
ß 2005, Elsevier Ltd. All Rights Reserved.
Introduction
Throughout most of the geological record we find
traces of the past inhabitants of our planet. Bones,
woody stems, and hard outer carapaces provide evi-
dence of the passing of species, entombed by the
sediments that were deposited around them. How-
ever, the organisms that yield these relatively familiar
kinds of fossils represent only the evolutionary
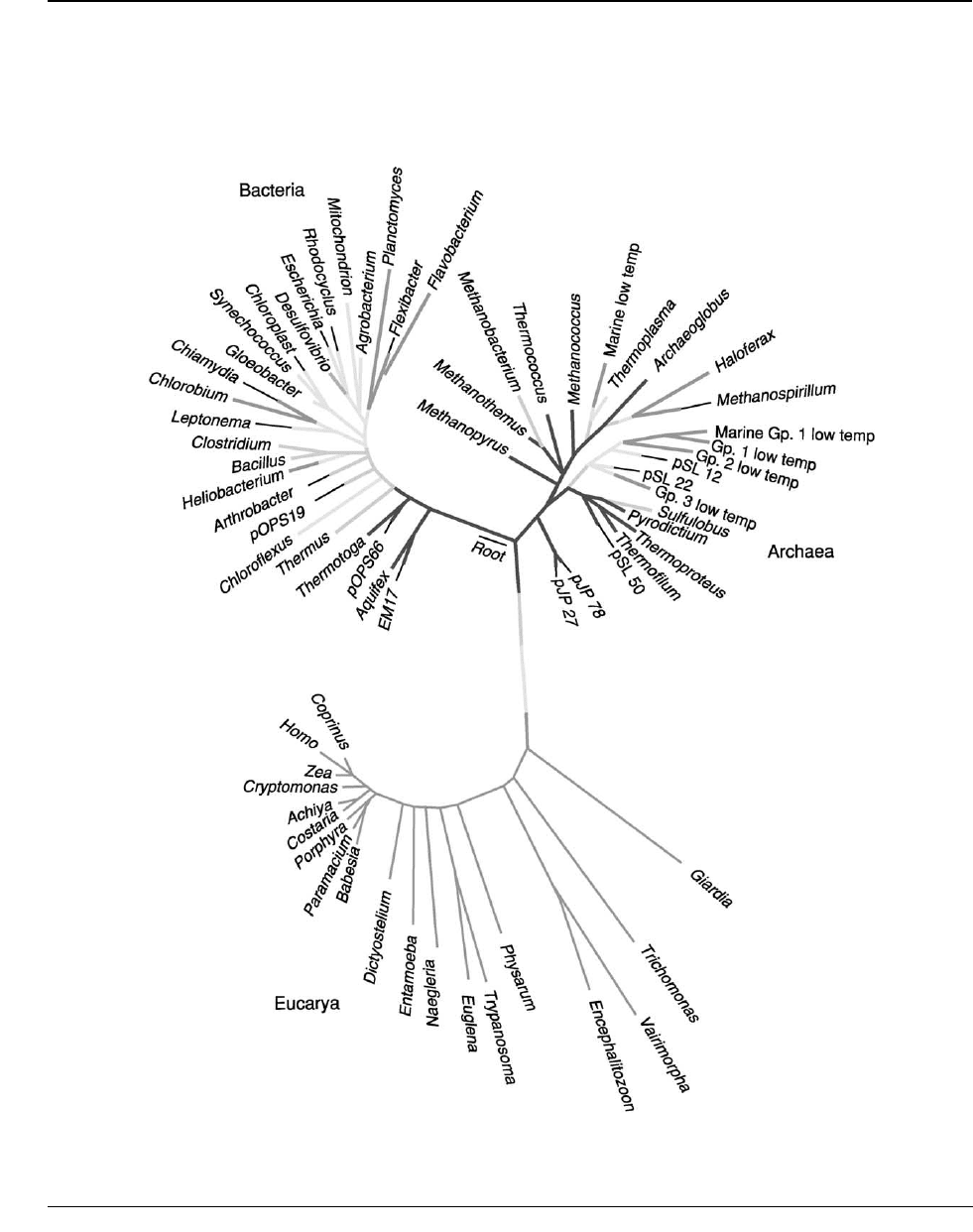
branch tips of the ‘Tree of Life’ (Figure 1), and have
risen only recently in the history of life on Earth. The
bulk of the ‘Tree of Life’, and most of the history of
life on Earth (Figure 2), is represented by microscopic
organisms. Unfortunately, these tiny, soft-bodied
creatures are not readily fossilized. Moreover, where
fossils are preserved, their tiny size, simple morphology,
and imperfect preservation make them extremely diffi-
cult to recognize. This challenge increases in the search
for the earliest fossils on Earth within rocks that have
been subjected to over 3 billion years of geological
processes: not only are the fossils likely to be heavily
degraded, but the very nature of the host sedimentary
deposits can be quite different to younger deposits, so
that the misinterpretation of unfamiliar structures can
be a risk.
The challenging search for the oldest traces of life
is further limited by the availability of suitable rocks.
There are just three occurrences of Early Archaean
sedimentary rocks, from remnant Archaean cratons in
Greenland (ca. 3.8 Ga), South Africa, and Western
Australia (3.4–3.5Ga; Figure 2). The rocks of Green-
land, however, have been subjected to pressures and
temperatures that have obliterated original depos-
itional fabrics. Thus, there are just two potentially
fossiliferous sedimentary sequences remaining from
the era during which life arose on Earth. Over recent
BIOSEDIMENTS AND BIOFILMS 279
decades, these areas have become the focus of studies of
the Earth’s earliest biosphere and, despite the imagined
impossibility of finding microbial remains in rocks
of such antiquity, there have indeed been several
‘fossil’ discoveries. Evidence for life at ca. 3.5 Ga com-
prises three different types of ‘fossil’: (1) microscopic
bacteria-like structures, interpreted as microfossils;
(2) chemical signatures, interpreted as resulting from
Figure 1 The ‘Tree of Life’ showing the genetic relationships between all living organisms. All branches are microbial, except for
Coprinus, Homo, and Zea, at the extreme tip of the lower branch.
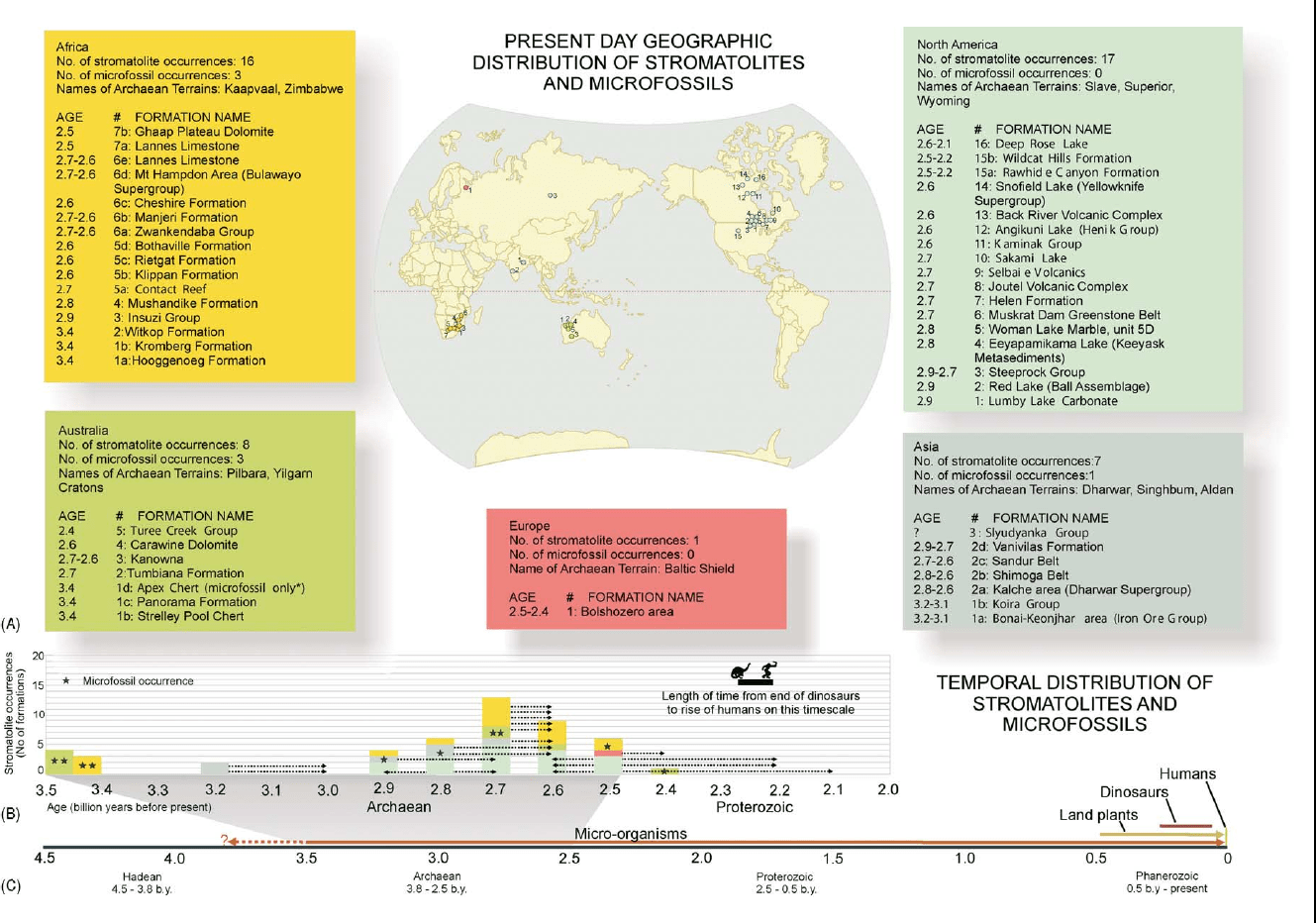
f0010 Figure 2 (A) Geographical distribution of Archaean stromatolite occurrences on the surface of the Earth today. All occurrences are
clustered on remnants of Archaean crust in Africa, Australia, North America, Europe, and Asia. The oldest known stromatolites and
microfossils occur in the Pilbara craton of Australia and the Kaapvaal craton in South Africa. (B) Temporal distribution of Archaean
stromatolite and microfossil occurrences. Data are coloured by continent. The age of some occurrences is approximate, and some
may extend into the Proterozoic, as indicated by dotted lines. (C) Geological time-scale highlighting the dominance of microbes
throughout most of the Earth’s history. Higher organisms represent only the most recent fraction of the history of life on Earth.
280 BIOSEDIMENTS AND BIOFILMS
BIOSEDIMENTS AND BIOFILMS 281