Elsevier Encyclopedia of Geology - vol I A-E
Подождите немного. Документ загружается.


of fossils that actually lived together and (ideally)
interacted with one another to the extent that the
grouping itself achieved a degree of stability – and
therefore recognizability – as a result of these
interactions.
Because properly defined assemblage zones reflect
ecological relations, this type of zone tends to be
restricted to particular facies. The ability of such
zones to track spatial shifts in environments through
time provides them with a distinctive utility in terms
of palaeoenvironmental analyses. However, this util-
ity comes at a price, and that price is a relatively
reduced ability to achieve long-distance chronostrati-
graphical correlations. For this reason, assemblage
zones of benthic organisms tend to be most useful
on local and regional scales. Assemblage zones of
planktonic organisms do perform well in terms of
chronological correlations, but the degree to which
such organismal groupings are maintained by close
inter-specific interactions is debatable.
Like Oppel zones, the specific criteria used to rec-
ognize assemblage zone boundaries are flexible. Not
all of the groups present in the zone’s ‘type area’ need
be present to recognise the zone in remote locations.
Unlike Oppel zones though, there is an objective and
independent rationale underlying this definitional
latitude. In the case of assemblage zones, one seeks
to recognise a set of dependent ecological relations
among species and between organisms and their
environment that transcend mere faunal and/or floral
lists. The objective reality of such patterns in nature is
well established by numerous studies of modern
faunas and floras and is reasonably well understood
from a theoretical point of view. Oppel zones, on the
other hand, are unified only in the vague sense that
the species used to recognize the zone are thought to
be useful in chronostratigraphical analysis. Although
there is certainly ample justification for suspecting
that, in many cases, the biostratigraphic ranges
of the species in different regions and habitats will
coincide, there is much less justification for regard-
ing these organisms as part of transcendent causal
association than is the case with assemblage zones.
As a result of the flexible manner in which assem-
blage zones are defined, the same groups can be used
to define different assemblage zones (e.g., a coral–
bryozoan assemblage zone and a coral–foraminiferal
assemblage zone can have zone-defining taxa in
common) and different members of the same eco-
logical association can be used to define different
assemblage zones. Assemblage zones have been used
frequently in areas where suitably short-ranging taxa
are not present or have not been studied.
By convention, the name of an assemblage zone
should be based on two or more taxa that figure
prominently in the zone’s definition (e.g., Eponides–
Planorbulinella Assemblage Zone, Eodicynodon
Assemblage Zone).
Interval Zone
An interval zone (also referred to as an interbiohor-
izon zone, gap zone, or a partial-range zone) is de-
fined as a body of strata delineated by the region
between two distinctive biostratigraphic horizons,
but that has no distinctive biostratigraphic identity
of its own (Figure 8). The boundaries of interval
zones can be marked by a wide variety of criteria.
These zones typically represent the undefined regions
between other types of zones; especially taxon range
zones.
An interval zone’s existence assumes a complemen-
tary relation with the underlying and overlying bios-
tratigraphically defined horizons that serve as their
inferior and superior boundaries. As with all other
types of biozones, the traditional one-dimensional
concept of biozone geometry (Figure 8A) can mask
the more complex geometries evident in two and
three-dimensional conceptualizations (Figure 8B). In
particular, interval zones are confined geographically
to only those regions in which the defining biozones
overlap. So long as one’s region of interest is confined
to the geographical area encompassed jointly by the
zone’s defining taxa, recognition of the zone can be
made with confidence. Outside this geographic envel-
ope, though, recognition of an interval zone becomes
problematic, if not impossible.
By convention, interval zones are either named for
the taxa used to define their boundaries (e.g., Globi-
gerinoides sicanus–Orbulina suturalis Interval Zone)
or for a taxon that occurs in the interval, but is not
itself used in the zone definition (e.g., Globigerina
ciperoensis Zone).
Acme Zone
An acme zone (also referred to as a peak zone, flood
zone, or epibole) is defined as a body of strata delin-
eated by the region of ‘maximal development’ of a
taxon (e.g., species, genus, family), but not its total
range (Figure 9). In this context, the term ‘maximal
development’ is meant to be used flexibly. In some
cases it might refer to an initial increase and subse-
quent decrease in the relative abundance of a taxon
that takes place within the confines of its biostrati-
graphic-geographic range. In others, it might refer to
an increase/decrease in body size, an increase or de-
crease in diversity, etc. Since these aspects of a taxon’s
evolutionary/ecological history tend to be strongly
associated with local and regional conditions, it is
on these spatial scales that acme zones have their
302 BIOZONES
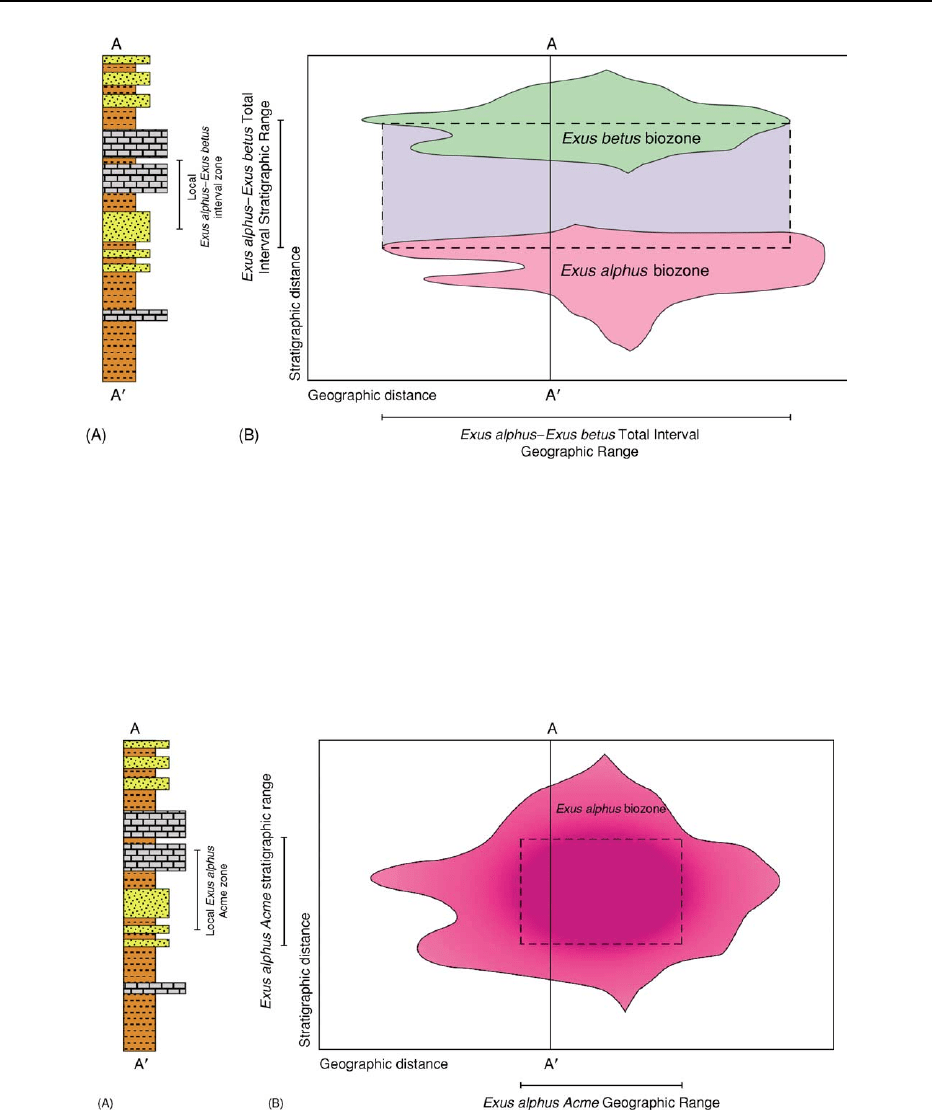
Figure 8 One-dimensional (A) and two-dimensional (B) representations of an interval biozone. In this example the zone begins at
the geographic acme of the hypothetical species Exus alphus and encompasses all strata between this extinction datum and the
overlying Exus betus geographic expansion datum. Globally, this interval between taxon range biozones defines a spatio-temporal
region within which the interval biozone’s inferred Total Stratigraphic Range (vertical) and Total Geographic Range (horizontal) are
defined (dashed box). In local sections however, the biozone will be represented by a one-dimensional interval whose boundaries are
included within this global biozone. Note the strong diachrony of the biozone boundaries. Intervals of strata that lie outside the
biozones’ geographical boundaries cannot be referred to the biozone
per se, but can be placed within the corresponding biochrono-
zone (¼ time interval represented by the Total Stratigraphic Range). Many other definitional geometries are also possible (e.g.,
interval between two species’ last appearances, interval between two species’ first appearances).
Figure 9 One-dimensional (A) and two-dimensional (B) representations of an acme biozone. In this example the zone begins at the
first substantial increase in the relative abundance of the species (symbolized by dark shading) of the hypothetical species
Exus alphus
and encompasses all strata in which the species is regarded as abundant. Globally, these abundance-based data define a spatio-
temporal region within which the interval biozone’s inferred Total Stratigraphic Range (vertical) and Total Geographic Range
(horizontal) are defined (dashed box). In local sections, however, the biozone will be represented by a one-dimensional interval
whose boundaries are included within this global biozone. Note the strong diachrony of the biozone boundaries. Intervals of strata that
lie outside the biozones’ geographical boundaries cannot be referred to the biozone
per se, but can be placed within the corresponding
biochronozone (= time interval represented by the Total Stratigraphic Range). Many other definitions of acme are also possible (e.g.,
increased size, increased ornamentation).
BIOZONES 303
greatest utility. Care must be taken when defining
such zones, that the recognition criteria employed
are as explicit and objective as possible. Nevertheless,
the popularity of such zones in regional basin ana-
lyses (particularly in commercial biozonations)
stands as testimony to their practical utility. By con-
vention, interval zones are named for the taxon used
in its definition (e.g., Emiliana huxleyi Acme Zone,
Tylosaurus Acme Zone).
Other Types of Biozones
Although the foregoing descriptions represent the
most frequently used biozone types, other types do
exist. These include such exotica as barren zones,
coiling-direction zones, negative association zones,
species pre-lap zone, species post-lap zone, etc. In
addition, biozone types exist that are not defined on
fossils per se, but rather on the trace fossils left in the
sediment as a result of animal and plant activity (e.g.,
track zones). All such zones are valid only to the
extent that they are useful to the stratigrapher and
geologist, and only to the extent that their definition
is based on unambiguous observational evidence. As
was noted by Hedberg in 1971, ‘Time and usage will
then be the surest means of determining whether it is
desirable for such [zones] to persist’.
Biozones and Biochronozones
Ever since the days of Smith, Lyell, and Oppel, the
purpose of establishing biozones has been to achieve
chronostratigraphical correlations. Other types of
stratigraphic intervals (e.g., key beds, magneto-
chrons, isotope zones) may be based on boundaries
whose emplacement is effectively synchronous over
geological time-scale. None is uniquely identifiable in
the same manner as biozones, however. Indeed, the
use of biostratigraphic methods to place, in an ap-
proximate temporal framework, the observations
upon which these other types of chronostratigraphic
inference depend, is so common it is rarely even men-
tioned. This failure to give biostratigraphical data due
credit for the basic contribution it makes would not
be so unfortunate, were it not for the fact that bios-
tratigraphy has come to be viewed by many as a
lacklustre and unimaginative field of study. Nothing
could be further from the truth. Biostratigraphic data
underpin not only most of what we know about
geological time over the last 600 million years, but
are critical to studies of palaeogeography, palaeoecol-
ogy, extinctions, diversifications, and patterns of
morphological evolution. In order to remind oneself
of the importance of biostratigraphical data, it is
worth recalling that the lack of a detailed time-scale
in which to place Proterozoic and Archaean events is
not due to a lack of palaeomagnetic reversals, cata-
strophic depositional events, isotopic excursions,
etc., but rather to the lack of fossil biotas on which to
base a coherent, finely resolved, and widely accessible
stratigraphy.
This having been said, the distinction between bio-
zones and chronozones must always be borne in
mind. All biozone boundaries are diachronous by
definition and, without recourse to radioisotopic
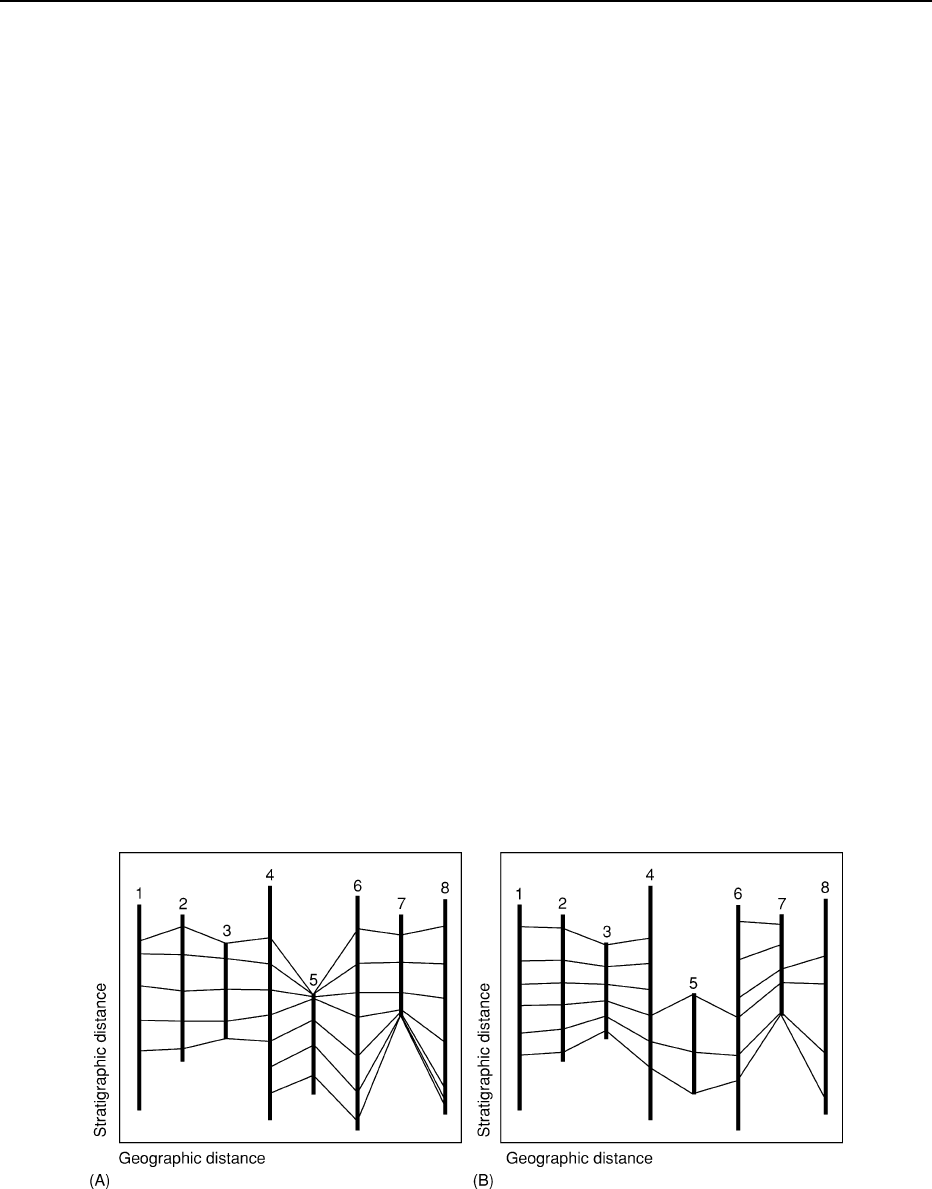
Figure 10 Results of multivariate biostratigraphic analyses on simulated biostratigraphic data from various geological sections
numbered 1 to 8 using the graphic correlation (A), as implemented by constrained optimization) and ranking and scaling (B) approaches.
These methods focus on the estimation of global, composite, taxon range biozones from datasets containing a series of biostratigraphic
first and last appearance datums for two or more stratigraphic successions or cores. Such approaches offer more consistency,
objectivity, and higher reproducibility of results than either qualitative or pairwise quantitative approaches to biostratigraphic
analysis. In these diagrams, connecting lines between the eight sections represent best-estimate lines of biochronostratigraphic
correlation.
304 BIOZONES

data, it is never correct to regard local biozones as
being equivalent to chronozones. Even when the
physical data necessary for absolute dating are avail-
able, it is no simple matter to accurately interpolate
the ages of biozone boundaries from radioisotopically
dated horizons.
Fortunately, several methods exist whereby strati-
graphers can compare sets of biostratigraphical ob-
servations in different successions or cores and,
through use of a few simple rules, create a summary
of the chronostratigraphically relevant data con-
tained in all such sections/cores. Graphic correlation
(Figure 10A) and ranking and scaling (Figure 10B) are
two of the more common quantitative methods
used for bridging the gap between observational
biostratigraphies and the ideal of a ‘biochronostrati-
graphy’. The ongoing incorporation of raw bio-
stratigraphical data into internally consistent and
synthetic composite successions based on the ap-
plication of these methods, along with improve-
ments in consistent identifying fossil morphologies
(e.g., via automated object recognition) promise to
take the science of biostratigraphy, and its foundation
concept – the biozone – into the twenty-first century.
There it will, once again, serve as the single most
useful contrivance in the practical stratigrapher’s
toolkit.
Glossary
Constrained optimization A mathematical technique
involving the search for the optimal condition or
structure of a system of observations given one or
more consistently applied external rules.
Depositional hiatus A horizon within a body of sedi-
mentary rock that represents a gap in time due to
the nondeposition of sediment, active erosion, or
structural complications.
Diachrony The condition of taking place at different
times.
Facies A stratigraphic body distinguished from other
such bodies by a difference in appearance or com-
position.
Homotaxis The condition of occupying the same
position in a sequence.
Isochrony The condition of being created at the same
time.
Microfossil Fossil shells or other hard parts of tiny
organisms or parts of organisms studied with the
aid of a microscope.
Phylogenetic systematics A method of determining
evolutionary relations between taxa based on the
nested patterns of shared characteristics derived
from the same pre-existing condition.
Stratum (strata) A tabular section of a rock body that
consists of the same type of rock material.
Taxon (taxa) Generalized term for any level within
the Linnean hierarchy of biological classification
(e.g., order, family, genus).
See Also
Analytical Methods: Geochronological Techniques.
Mesozoic: Triassic; Jurassic; Cretaceous; End Cret-
aceous Extinctions. Palaeozoic: Cambrian; Ordovician;
Silurian; Devonian; Carboniferous; Permian; End Per-
mian Extinctions. Precambrian: Vendian and Ediacaran.
Sequence Stratigraphy. Stratigraphical Principles.
Tertiary To Present: Paleocene; Eocene; Oligocene;
Miocene; Pliocene; Pleistocene and The Ice Age. Time
Scale.
Further Reading
Cubitt JM and Reyment RA (1982) Quantitative strati-
graphic correlation. Chichester: Wiley.
Gradstein FM, Agterberg FP, Brower JC, et al. (1985)
Quantitative stratigraphy. Dordrecht: D. Reidel.
Hedberg HD (1971) Preliminary report on biostratigraphic
units. Montreal, Canada: International Subcommission
on Stratigraphic Classification.
Hedberg HD (1976) International stratigraphic guide: a
guide to stratigraphic classification, terminology, and
procedure. New York: John Wiley & Sons.
Kauffman EG and Hazel JE (1977) Concepts and methods
in biostratigraphy. Stroudsburg, Pennsylvania: Dowden,
Hutchinson & Ross, Inc.
Mann K, Lane HR, and Stein J (1995) Graphic correlation.
Tulsa: Society of Economic Paleontologists and Mineral-
ogists, Special Publication 53.
Miall AD (1984) Principles of sedimentary basin analysis.
New York: Springer-Verlag.
Rawson PF, Allen PM, Brenchley PJ, et al. (2002) Strati-
graphical procedure. London: The Geological Society.
Reyment RA (1980) Morphometric methods in biostrati-
graphy. London: Academic Press.
Shaw A (1964) Time in stratigraphy. New York: McGraw-
Hill.
BIOZONES 305
BRAZIL
F F Alkmim and M A Martins-Neto, Universidade
Federal de Ouro Preto, Ouro Preto, Brazil
ß 2005, Elsevier Ltd. All Rights Reserved.
Brazil in the Geological Scenario
of South America
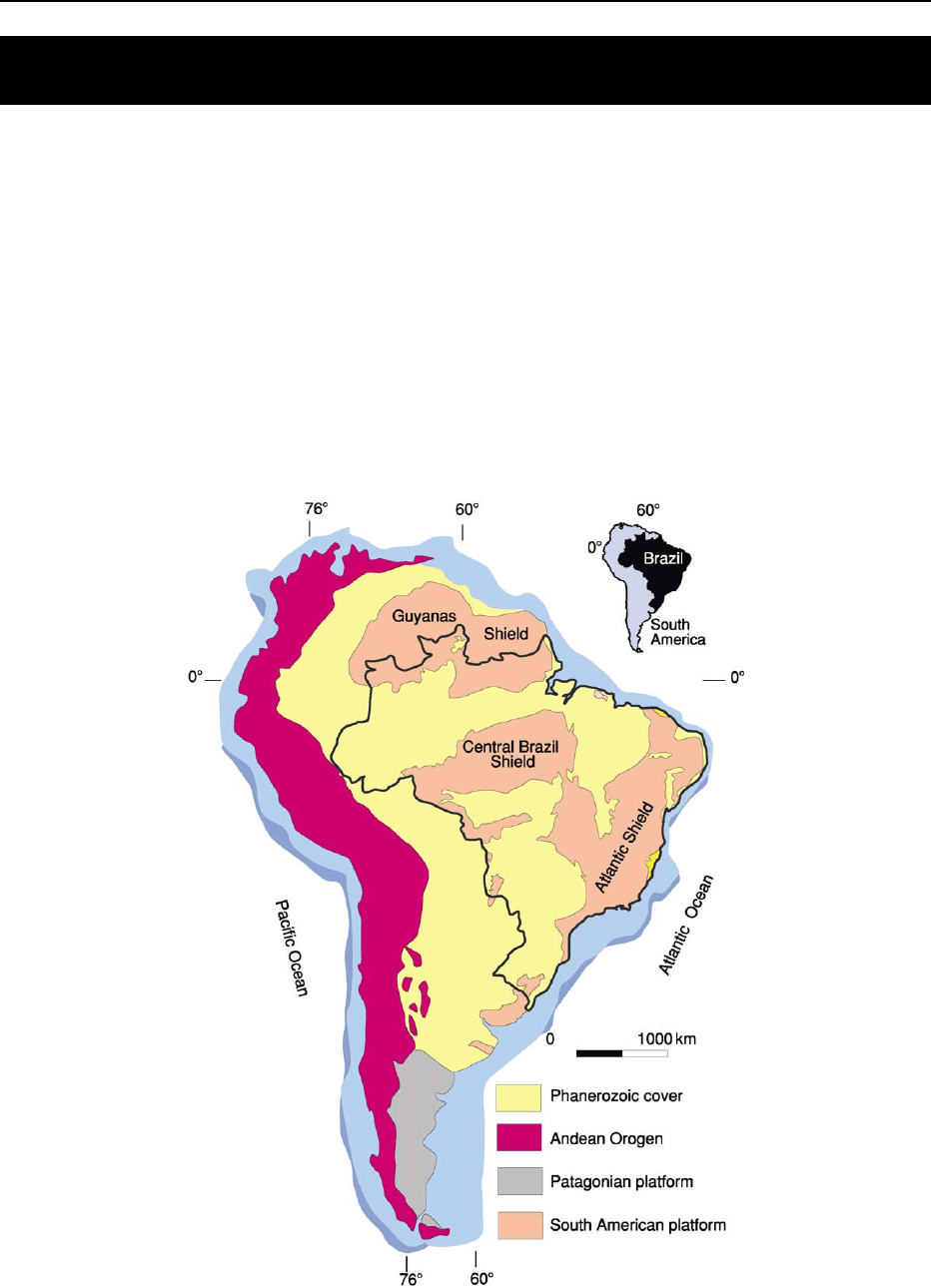
The South American continent comprises five major
tectonic units: the Pacific active margin, the Atlantic
passive margin, the Andean Orogen, and the Patagon-
ian and South American platforms (Figure 1). The
continental margins and the Andean Orogen are the
younger portions of the continent; the platforms, on
the other hand, correspond to the mature and ancient
parts of South America.
Brazil is located entirely on the South American
platform, which is defined as the Precambrian core
of the continent, not affected by the Andean oroge-
nies (Figure 1). The exposures of the South American
platform are collectively referred to as the Brazilian
Shield, which in reality encompasses three distinct
morphotectonic domains: the Guyanas, the Central
Brazil (also called Guapore
´
) and the Atlantic shields
(Figure 1). Sedimentary basins, including large Palaeo-
zoic sags, Cretaceous passive and transform margins,
and Tertiary rifts make up the Phanerozoic cover of
the South American platform (Figure 1).
Figure 1 Tectonic map of South America, showing the main subdivisions. Modified from Almeida FFM, Hasui Y, Brito Neves BB, and
Fuck RA (1981) Brazilian structural provinces: an introduction. Earth Science Reviews 17: 1–29, with permission from Elsevier.
306 BRAZIL
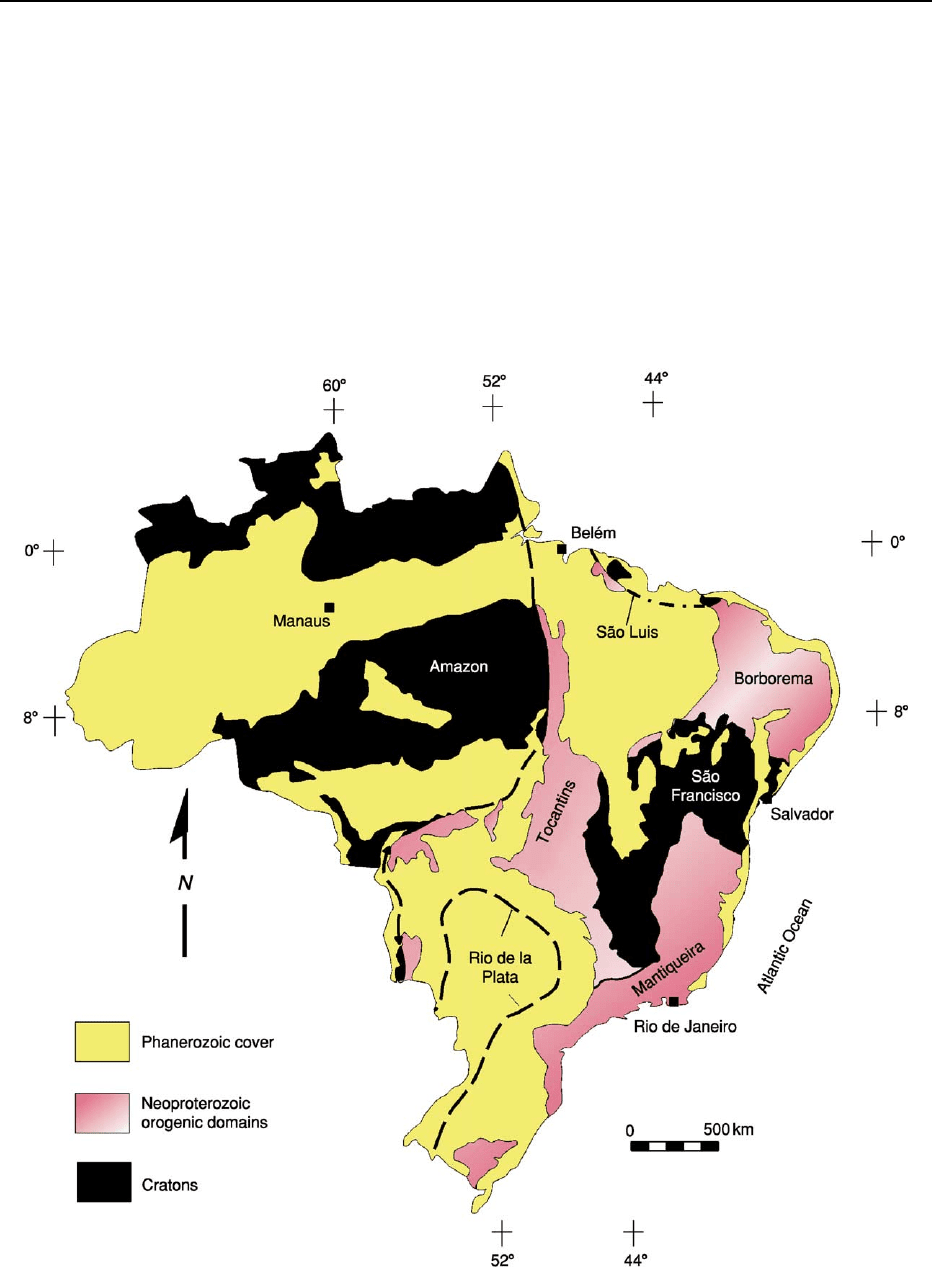
Two fundamentally distinct lithospheric compon-
ents, namely cratons and Neoproterozoic orogens,
form the Precambrian core of South America. The
cratons correspond to the relatively stable pieces of
the continent that escaped the Neoproterozoic oroge-
nies recorded in all the remaining shield areas of
Brazil. Four cratons have been delimited in the South
American platform: the Sa
˜
o Francisco, Amazon, Sa
˜
o
Luis, and Rio de la Plata cratons (Figure 2). The non-
cratonic segments of the platform are the Neopro-
terozoic Mantiqueira, Tocantins, and Borborema
orogenic domains (Figure 2). (In the Brazilian
geological literature each component of the South
American platform and its cover (i.e. cratons, Brasi-
liano orogens, and Phanerozoic basins) is referred to
as a tectonic province.)
Together with Africa, the South American platform
once lay in the western portion of Gondwana, the
supercontinent assembled by the end of the Neo-
proterozoic and split apart in the Cretaceous, which
also encompassed Antarctica, India, and Australia
(Figure 3). The assembly of western Gondwana
resulted from a series of diachronic collisions, pre-
dominantly between 640 Ma and 520 Ma, the
Figure 2 Simplified tectonic map of Brazil, showing the subdivisions of the South American platform and its Phanerozoic cover.
Modified from Almeida FFM, Hasui Y, Brito Neves BB, and Fuck RA (1981) Brazilian structural provinces: an introduction. Earth Science
Reviews
17: 1–29, with permission from Elsevier.
BRAZIL 307
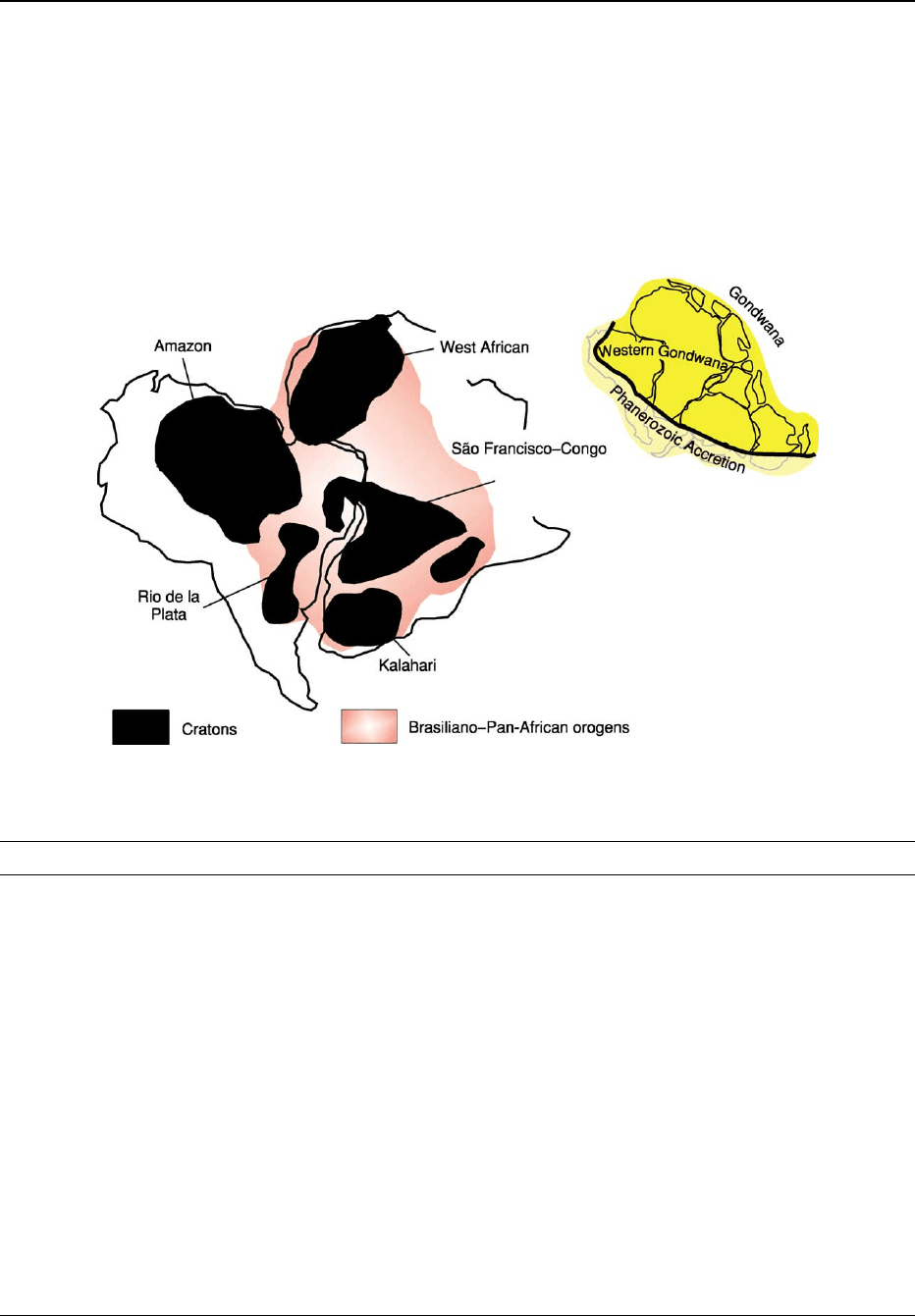
so-called Brasiliano or Pan-African orogenies. In this
context, the cratons of South America and Africa
are the preserved and more internal portions of the
plates that collided to build up western Gondwana;
the Neoproterozoic orogenic domains encompass
the margins of these plates and other lithospheric
pieces also involved in the tectonic collage that
made up western Gondwana. The dispersal of
western Gondwana and the opening of the South
Atlantic in the Lower Cretaceous broke apart Neo-
proterozoic orogens and cratons. Consequently, the
Neoproterozoic orogens and cratons of eastern Brazil
have African counterparts (Figure 3).
At first glance the geological panorama of Brazil
reflects only the Neoproterozoic assembly, Early
Palaeozoic to Jurassic permanence, and Cretaceous
Figure 3 Schematic map of western Gondwana, showing the cratons and Neoproterozoic Brasiliano–Pan-African belts.
Table 1 The main thermotectonic events recorded in the South American platform
Event Age Occurrence Manifestation Significance
Tertiary
reactivation
Eocene–Oligocene Eastern and
Northern Brazil
Rifting, alkaline magmatism
South Atlantic 130–78 Ma Continental margin and
interior
Flood basalts, followed by
rifting and a late phase of
alkaline magmatism
Gondwana breakup,
opening of the
South Atlantic
Brasiliano 640–520 Ma Non-cratonic areas Deformation, metamorphism,
magmatism, minor accretion
Western Gondwana
assembly
Macau
´
bas rifting 900–800 Ma Sa
˜
o Francisco Craton and
adjacent belts
Rifting, bimodal magmatism,
glaciation
Rodinia breakup
Rondonian/
San Igna
´
cio/
Sunsa
´
s
1500–1000 Ma South-western Amazon
Craton
Deformation, metamorphism,
magmatism
Rodinia assembly
Staterian rifting 1750 Ma Sa
˜
o Francisco Craton and
adjacent belts
Rifting, bimodal magmatism Atlantica breakup
Uatuma
˜
1800 Ma Amazon Craton Anorogenic magmatism Atlantica breakup,
plume
Transamazonian 2200–1950 Ma Amazon and Sa
˜
o Francisco
cratons, all Brasiliano
orogenic domains
Deformation, metamorphism,
magmatism, crustal
accretion
Assembly of
Atlantica
supercontinent
Jequie
´
, Rio Das
Velhas,
Aroense
2900–2780 Ma Sa
˜
o Francisco and Amazon
cratons, some Brasiliano
orogenic domains
Deformation, metamorphism,
magmatism, crustal
accretion
308 BRAZIL
dispersal of western Gondwana. A closer examination
of the cratons and Neoproterozoic orogenic belts
reveals, however, a long and diverse pre-Gondwana
history, as well as a whole series of post-Gondwana
features. Thus, in addition to the Brasiliano and South
Atlantic events, other events of the same significance
and extent are recorded in different ways in the South
American platform and its cover. The most important
of these events, together with their ages, areas of
occurrence, and geotectonic significance, are shown
in Table 1.
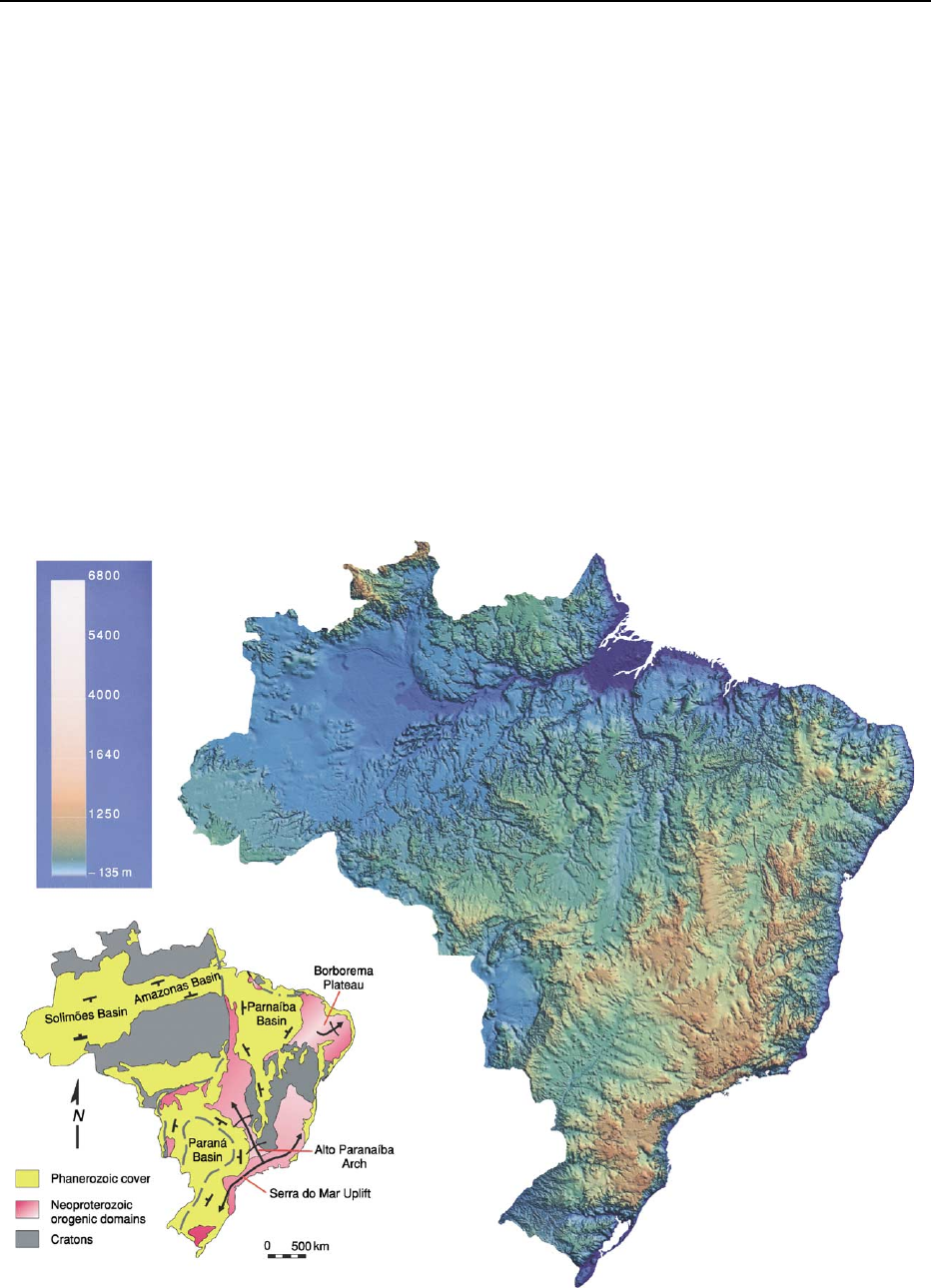
Regional Structures and Topography
of Brazil
The topography of Brazil to a large extent reflects the
constitution of the South American platform dis-
cussed in the previous section. The cratons underlie
the low areas, hosting the main river basins; the
highlands, on the other hand, have the Neoprotero-
zoic orogenic domains as their substrata (Figure 4).
Each of the large-scale topographical highs and
lows of the Brazilian territory is the expression of a
particular regional tectonic structure. Some of these
structures had already nucleated by the Palaeozoic.
The majority, however, were initiated in the Meso-
zoic and underwent significant reactivation during
the Cenozoic. The most prominent structural and
topographical lows correspond to the Parana
´
,Sa
˜
o
Francisco, Parnaı
´
ba, and Amazonas basins. The
Borborema Plateau, the Serra do Mar Uplift, and
the Alto Paranaı
´
ba Arch are the largest structural
and topographical highs (Figure 4).
Cratons
The cratons of the South American platform,
consisting of Archaean crust with substantial
Figure 4 Correlation between the large-scale tectonic structures and the topographical relief of Brazil. Relief map compiled by JBL
Franc¸ olim based on EROS GTOPO 30 dataset; reproduced with the permission of the author.
BRAZIL 309
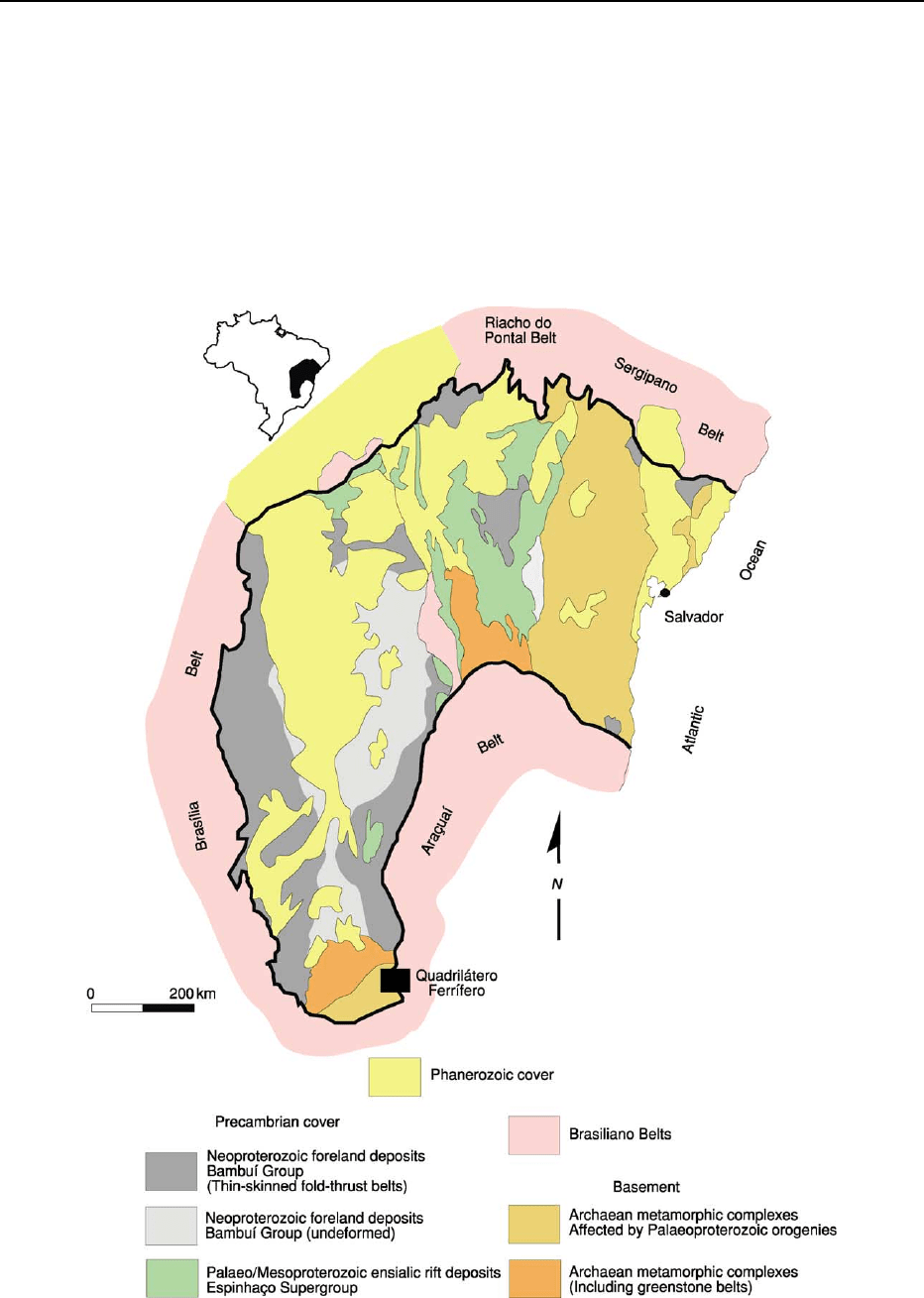
Palaeoproterozoic accretions, are the portions of the
Precambrian basement that were unaffected by the
Brasiliano orogenies. Their boundaries are defined
by the deformation styles of Neoproterozoic cover
strata, patterns of geophysical anomalies, and geo-
chronological data. Accordingly, the cratons of Brazil
are bounded on all sides by Brasiliano basement-
involved fold–thrust belts, but contain Brasiliano
thin-skinned foreland fold–thrust belts. In this regard
they differ from similar features delimited on other
continents. The cratons of Brazil do, however, exhibit
the typical attributes of their equivalents worldwide,
such as mantle roots, low heat-flow values, and high
lithospheric strength.
Sa
˜
o Francisco Craton
A map view of the Sa
˜
o Francisco Craton shows a
shape that resembles a horse’s head, with the north-
ern segment merging with the east coast of Brazil
(Figure 5). Except for the Atlantic coast, the Sa
˜
o
Figure 5 Simplified geological map of the Sa
˜
o Francisco Craton showing the distribution of the major lithostratigraphical units.
310 BRAZIL
Francisco Craton is bounded on all sides by the exter-
nal fold–thrust belts of the Mantiqueira, Borborema,
and Tocantins orogenic domains (Figures 2 and 5). As
indicated by reconstructions of western Gondwana,
the Sa
˜
o Francisco Craton and the Congo Craton of
Africa formed a single lithospheric unit from the
Palaeoproterozoic until the Cretaceous opening of
the South Atlantic.
The basement, made up of Archaean metamorphic
complexes (tonalite–trondhjemite–granodiorite associ-
ation and voluminous calc-alkaline plutons), Archaean
greenstone belts, and Palaeoproterozoic metasedi-
mentary successions, is exposed in the northern lobe
of the craton and in a smaller area close to the south-
ern boundary (Figure 5). In both exposures, Archaean
nuclei are bounded on the east and south-east by
Palaeoproterozoic Transamazonian fold–thrust belts,
which involve the Archaean basement and Palaeopro-
terozoic metasedimentary units. The second-largest
and best-studied mineral province of Brazil, the
Quadrila
´
tero Ferrı
´
fero (Iron Quadrangle), lies partly
in the southern portion of the craton and partly in
the adjacent Brasiliano Arac¸uaı
´
belt. In this pro-
vince, gold is found in an Archaean greenstone-belt
sequence, and high-grade iron-ore deposits occur in a
Palaeoproterozoic banded iron formation.
Alluvial to marine sediments with 1750 Ma acid-
volcanic intercalations at the base (representing the
fill of an ensialic rift system), glacial-influenced
Tonian (ca. 850 Ma) rift sediments, and Cryogenian
(720–600 Ma) foreland strata form the Precambrian
cover of the craton. The foreland-basin strata are
deformed in the areas adjacent to the craton bound-
aries, thereby defining two thin-skinned foreland
fold–thrust belts with opposite vergences. These
belts represent the orogenic fronts of the Neoproter-
ozoic Brası
´lia
and Arac¸uaı
´
belts, which fringe the
craton to the west and to the east, respectively.
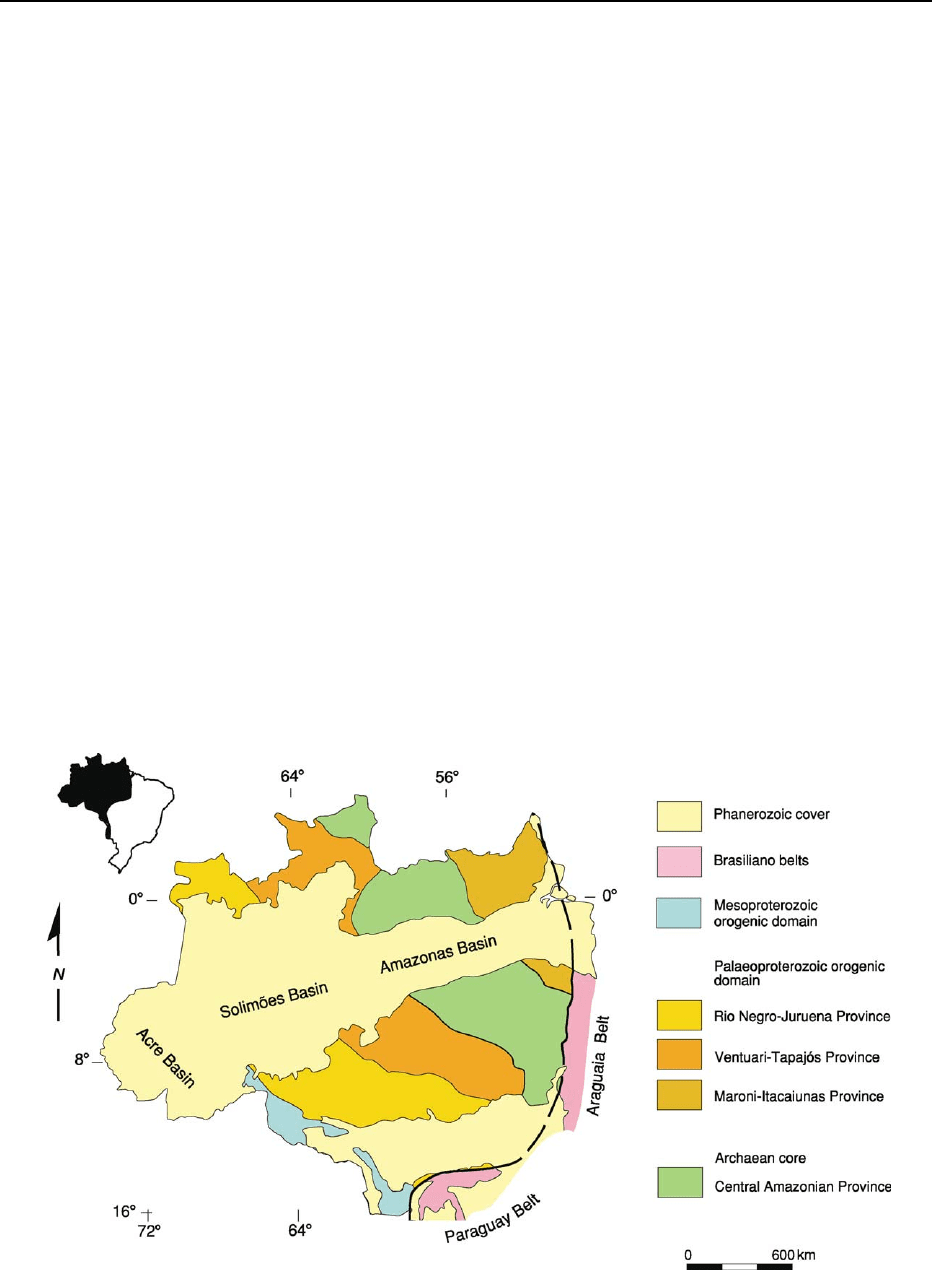
Amazon Craton
The Amazon Craton encompasses an area of approxi-
mately 430 000 km
2
in north-western South America,
extending far beyond the Brazilian borders into
Colombia, Venezuela, Guyana, Suriname, and French
Guiana. The eastern and south-eastern limits of the
craton are marked by the external portion of the Bra-
siliano Tocantins Orogen, represented by the Ara-
guaia and Paraguay belts, respectively. The western
and north-western boundaries are covered by the sub-
Andean basins, and the northern limit is marked
by the South American equatorial margin (Figures 2
and 6).
More than two-thirds of the Brazilian portion of
the craton is covered by Phanerozoic sediments and
the Amazon forest, so that the geological knowledge
Figure 6 Schematic map of the Amazon Craton, illustrating the main basement components and cover units. Modified from Tassinari
CG, Bettencourt S, Geraldes M, Macambira JB, and Lafon JM (2000) The Amazonian Craton. In: Cordani UG, Milani EJ, Thomaz FA, and
Campos DA (eds.)
Tectonic Evolution of South America, pp. 41–95. 31st International Geological Congress, Rio de Janeiro.
BRAZIL 311