Caballero B. (ed.) Encyclopaedia of Food Science, Food Technology and Nutrition. Ten-Volume Set
Подождите немного. Документ загружается.

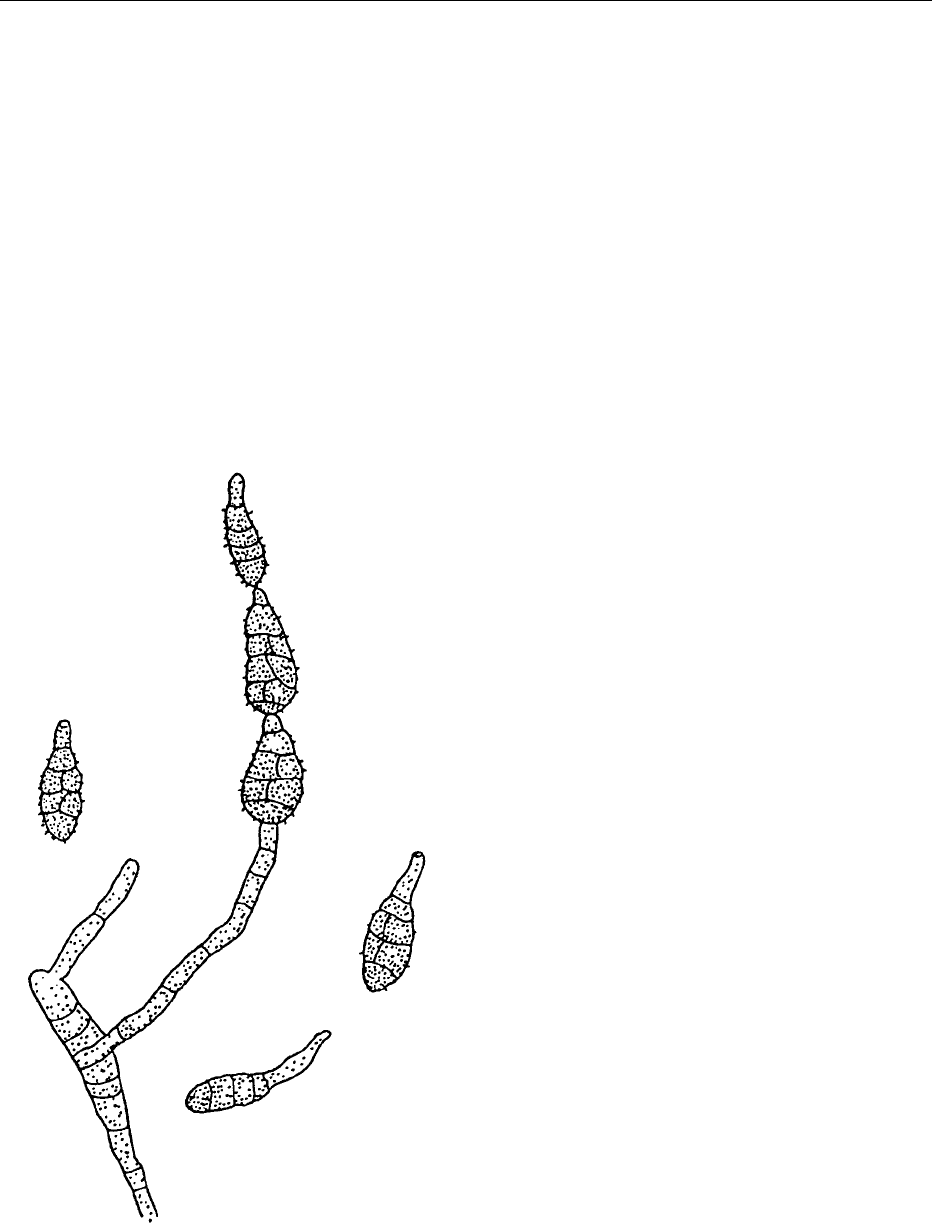
Alternaria
0026 Species of Alternaria occur worldwide on many foods
and plant materials. They have septate hyphae and
appear to be dark, gray-green in color, and almost
black on the reverse side of colonies growing on agar.
They produce septate conidia which are also dark-
colored (Figure 4). The septa are both transverse and
longitudinal, and spores are oblong in shape, some-
times with an elongated apical cell. The spores are
produced in chains. Alternaria can cause spoilage of a
variety of foods, including tomatoes and fresh vege-
tables, such as bell peppers. Alternaria species have
been found in the core area of apples, in nuts, includ-
ing peanuts, hazelnuts, and pecans, in grains such as
wheat and sorghum, and in cold-stored meat and
spices. Alternaria are capable of growing at low tem-
peratures on refrigerated foods. Common species in-
clude A. alternata, which is considered synonymous
with A. tenuis,andA. tenuissima. Alternaria species
are capable of producing several mycotoxins and thus
present both food spoilage and potential safety
hazards.
Trichothecium
0027Trichothecium is a common genus of mold which
occurs throughout the world. A common species is
T. roseum, which is pink in color, and has been found
growing on various fruits and vegetables, as well as
cereal grains, such as barley, wheat, and corn, and
cereal products such as flour. It has also been found
on beans, pecans, filberts, and meat. Trichothecium
species produce clusters of two-celled or single-
septate conidia that are elliptical to pear-shaped.
Some strains of Trichothecium have been shown to
produce trichothecenes.
Trichoderma
0028Two of the most common species of Trichoderma are
T. viride and T. harzianum. Trichoderma colonies
are bright green. Trichoderma contaminate many
food products, including stored grains such as barley,
wheat, and oats, nuts such as peanuts and pecans, and
various fruits and vegetables, such as tomatoes, sweet
potatoes, and citrus fruit.
Cladosporium
0029Cladosporium has dark mycelia which may be brown
to blackish-brown or gray-green in color. The reverse
side of a colony of Cladosporium on agar is very dark
greenish-black or blue-black. Cladosporium species
produce many one-celled conidia, but two- and
three-celled forms are also common. Two common
species are C. cladosporioides and C. herbarum. Cla-
dosporium species, which can grow at low tempera-
tures, tend to be slow-growing and form black spots
on foods. Cladosporium species have been isolated
from cereal grains, peanuts, fruits, and refrigerated
meat, particularly beef. Cladosporium is very common
and can grow under refrigerated storage conditions.
The main consequence of Cladosporium species in
foods is spoilage and discoloration.
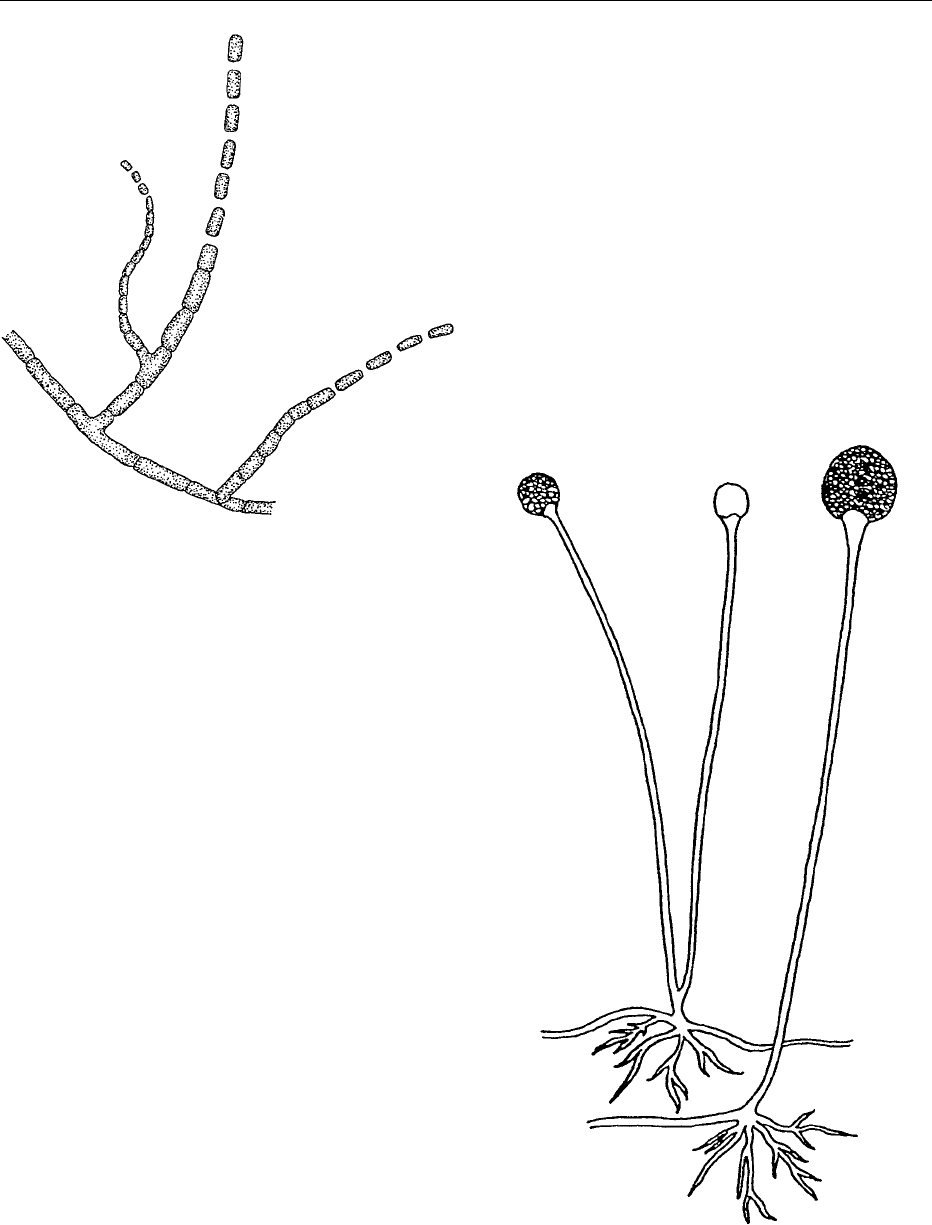
Geotrichum
0030Geotrichum has septate mycelia which readily frag-
ment into arthrospores (Figure 5), which are the
organism’s primary means of reproduction. The only
species that is significant in foods is G. candidum;
other names for this organism are Oidium lactis
and Oospora lactis; it produces ascospores, and the
fig0004 Figure 4 Multicelled conidia of Alternaria with transverse and
longitudinal septa. Reproduced from Spoilage: Fungi in Food –
An Overview. Encyclopaedia of Food Science, Food Technology and
Nutrition, Macrae R, Robinson RK and Sadler MJ (eds), 1993,
Academic Press.
SPOILAGE/Fungi in Food – An Overview 5517
telemorphic state is known as Endomyces geotri-
chum. G. candidum is referred to as a yeast-like
fungus because it exhibits a dimorphic form and
readily fragments into arthrospores, which appear to
be single-celled. In addition, colonies of this organism
can be white, soft, creamy, and yeast-like in appear-
ance. G. candidum is a problem on food-processing
equipment, particularly in vegetable-processing
plants, and is referred to as ‘machinery mold.’
Unclean equipment provides a favorable environment
for rapid growth of this organism. It attacks citrus
fruits, as well as other fruits. It is able to infect fruits
mainly through injuries to the skin, as well as growing
on overripe fruit. G. candidum has also been obtained
from cheese, meats, and frozen foods, particularly
vegetables.
Rhizopus
0031 Rhizopus differs from the molds previously described,
because it is nonseptate and produces sporangio-
spores rather than conidia. It is a very-fast-growing,
spreading type of mold which has white mycelia and
black sporangia. Rhizopus species form rhizoids at
the base of the sporangiophores, and columella in the
sporangium (Figure 6). Young sporangia are white
before turning black with age. Probably the most
common of the Rhizopus species is R. stolonifer, the
so-called bread mold. Besides bread, Rhizopus causes
spoilage of strawberries, other berries, fruits, and
vegetables. Rhizopus species have also been isolated
from cereal grains, nuts, and meat. R. oligosporus is
used in making tempeh and certain other mold-
fermented foods. Rhizopus is placed in the taxonomic
order Mucorales and is somewhat typical, and prob-
ably the most common genus of that order to contam-
inate foods. Other genera in the order Mucorales
which are nonseptate and produce sporangiospores
are Absidia, Mucor, Rhizomucor, Syncephalastrum,
and Thamnidium. All of the Mucorales genera
that contaminate food are found in the family
Mucoraceae. These genera are sometimes referred
to as mucoraceous fungi. They are a part of the
fig0005 Figure 5 Arthrospores typical of Geotrichum. Reproduced from
Spoilage: Fungi in Food – An Overview. Encyclopaedia of Food
Science, Food Technology and Nutrition, Macrae R, Robinson RK
and Sadler MJ (eds), 1993, Academic Press.
fig0006Figure 6 Sporangiospore typical of Rhizopus and other mucor-
aceous fungi. Reproduced from Spoilage: Fungi in Food – An
Overview. Encyclopaedia of Food Science, Food Technology and Nu-
trition, Macrae R, Robinson RK and Sadler MJ (eds), 1993, Aca-
demic Press.
5518 SPOILAGE/Fungi in Food – An Overview

class Zygomycetes and form zygospores by sexual
processes and chlamydospores (Figure 7). They re-
quire high-moisture conditions for growth.
0032 For more detailed descriptions of these and other
genera of molds, refer to the references given in the
Further Reading Section at the end of this article.
Recognizing Molds
0033 Molds are recognized primarily as visible fuzzy, cot-
tony, or colored growth on the surfaces of substrates
such as foods or other organic materials. In grains,
seeds, and animal feeds, however, such growth may
not always be visible in this manner. Internal mold
contamination and growth in nuts, seeds, and grains
may not always be visible as filamentous growth.
However, mold growth in nuts, whole grains or
seeds often results in discoloration and altered ap-
pearance. In corn (including popcorn), for example,
invasion and growth of mold in the germ area
may produce darkened, greenish- or blueish-colored
germ areas in affected kernels. When this growth is
attributable to certain blue-colored Aspergillus or
Penicillium species, and a blueish line appears in
the germ area, the condition is known as ‘blue eye.’
In other grains, such as wheat, white to pinkish
discolored and shrivelled kernels may indicate mold
invasion. Likewise, with nuts, shrivelled or misshapen
kernels may be caused by mold, although that is not
the only cause of such problems.
Health Implications
0034Mold deterioration of foods and agricultural com-
modities is a serious problem worldwide. Besides
deterioration of food and feed products by causing
damage that reduces quality, grades, and prices,
resulting in economic loss, the presence of molds in
foods and commodities poses hazards to human and
animal health. A serious hazard associated with mold
growth in foods and feeds is the possible production
of mycotoxins – substances which are toxic and some
of which are carcinogenic. (See Aflatoxins; Myco-
toxins: Classifications.) Additional hazards arise
from the presence of the molds themselves. A wide
spectrum of human and animal diseases are attribut-
able to molds, ranging from mycotic abortion to afla-
toxin poisoning in animals, and from allergic
reactions to life-threatening systemic infections in
humans. In recent years, invasive mycoses have
become a more frequent and important cause of mor-
bidity and mortality in humans, particularly in indi-
viduals who are immunosuppressed as a result of
organ transplantation, underlying disease (acquired
immune deficiency syndrome or AIDS), chemother-
apy, radiation therapy, or age. The environment, in-
cluding foods, is the source of these molds. Certain
occupational niches, such as grain handling and stor-
age, pose additional hazards to agricultural workers
through exposure to high levels of mold spores and
endotoxins in grain dusts. These dusts may also be a
significant source of exposure to mycotoxins. Various
lung diseases in grain handlers, farmers, and food-
producing animals are associated with molds and
grain dust.
Yeasts
0035Yeasts are single-celled microscopic organisms that
are larger than bacteria, and are oval, elliptical, spher-
ical, or elongated into rod shapes. Most yeasts repro-
duce by a process called budding (Figure 8). Budding
is an asexual process in which a bulge (bud) develops
on the cell wall, and protoplasm, including replicated
nuclear material, fills the bulge. The bulge grows in
size and eventually a wall forms between the bulge
and the parent cell, forming a new cell. The new cell
then separates from the parent cell. Buds may form in
(a) (b)
fig0007 Figure 7 (a) Zygospore and (b) chlamydospores. Reproduced
from Spoilage: Fungi in Food – An Overview. Encyclopaedia of
Food Science, Food Technology and Nutrition, Macrae R, Robinson
RK and Sadler MJ (eds), 1993, Academic Press.
SPOILAGE/Fungi in Food – An Overview 5519
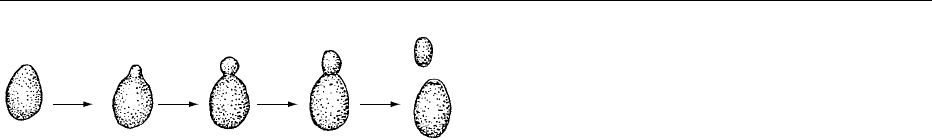
a polar position or multilaterally. Some yeasts repro-
duce by binary fission, similar to fission in bacteria,
but these are few in number. One yeast reproduces by
both fission and budding. So-called true yeasts also
reproduce by a sexual process involving conjugation
of two cells and the resultant formation of ascospores
within the cells. The cell containing ascospores is then
known as an ascus. A few yeasts produce ascospores
without conjugation, but the resulting ascospores
then undergo conjugation. True yeasts are referred
to as ascosporogenous yeasts and are classified in
the subdivision Ascomycotina, which used to be
called Ascomycetes. Asporogenous yeasts, also called
false yeasts, do not produce ascospores or other
sexual spores, and are placed in the subdivision Deu-
teromycotina, also called Fungi Imperfecti.
0036 Another type of spore formed by some yeasts is
called a chlamydospore. A chlamydospore forms
when a thick wall surrounds the yeast cell, and is
similar to chlamydospores formed by molds. These
appear to be resting or survival structures, and are
asexual. Yeasts that form pseudomycelia can also
produce by fragmentation spores known as arthro-
spores and blastospores.
Growth Requirements
0037 Individual yeasts vary in their growth requirements,
but some generalizations can be made. Yeasts are
aerobic and facultatively anaerobic. Some yeasts
are extremely aerobic and oxidative in their metabol-
ism. These oxidative yeasts grow on the surface of
liquids and are known as film yeasts. Yeasts that are
aerobic and facultatively anaerobic can grow both
ways. When growing aerobically, yeasts produce
mainly biomass, i.e., more yeast cells. When growing
anaerobically, yeasts have a fermentative metabolism
and produce carbon dioxide and ethanol. For the
most part, sugars are the best source of energy for
yeasts, but the film yeasts can oxidize organic acids
and alcohol for energy. Yeasts can utilize simple ni-
trogen compounds, such as ammonia, urea, and
amino acids. Some yeasts can also hydrolyze proteins
and polypeptides.
0038Yeasts require somewhat less available moisture
than bacteria, but more than molds. In terms of a
w
,
the lower limits for most yeasts are 0.88–0.94. Some
yeasts, known as osmophilic yeasts, can grow in the
presence of high concentrations of sugars or salt at a
w
as low as 0.62–0.65. Each yeast will have its own
characteristic optimal a
w
and a range of a
w
over
which it can grow. The minimum and optimum a
w
for a given yeast may change depending on outside
factors, such as nutrition, pH, temperature, oxygen,
and presence of inhibitors.
0039The optimum temperature for growth of yeasts is
in the mesophilic range of 25–30
C. Yeasts in general
can grow over a range of temperatures from 0
Cto
47
C. Yeasts grow well under acid conditions, at pH
4.0–4.5. They can grow at lower pH than most bac-
teria, but do not grow well under alkaline conditions.
Yeasts are widely distributed in nature and commonly
occur on grapes and other fruits. They are also found
in dust and water, and may be found on the skin and
in the intestinal tract of humans and animals.
Food Spoilage and Effects of Processing
on Yeasts
0040Some yeasts, mostly in the genus Saccharomyces, are
very important industrially for leavening of bread
and production of wine, beer, alcohol, glycerol,
and invertase. Other yeasts, referred to as wild yeasts,
can cause spoilage problems in foods. Film yeasts can
oxidize enough lactic acid to allow spoilage in sauer-
kraut and pickle fermentations by nonacid-tolerant
organisms. Yeasts can grow in vacuum-packaged
wieners and other cured meats, forming visible col-
onies or slime. Other salt-tolerant yeasts can grow in
curing brines, salted meats and fish, soya sauce,
tamari sauce, and miso. Osmophilic yeasts can grow
well in foods with high concentration of solutes, es-
pecially sugars and salts, and cause spoilage of salad
dressings, honey, syrups, concentrated fruit juices,
and even dried fruits. Certain other yeasts, known
as apiculate or lemon-shaped yeasts, can contaminate
wine fermentations, causing off-flavors, low yields of
alcohol, and production of volatile acids. Yeasts can
also reach high numbers in frozen vegetables prior to
freezing or after thawing, especially if the vegetables
are mishandled.
0041Yeasts are not heat-resistant and are readily des-
troyed in most heat processes. Other food processes,
such as freezing, refrigeration, vacuum packaging,
and acidification, will not necessarily destroy yeasts.
Vacuum packaging will prevent growth of highly
aerobic film yeasts, but not of fermentative types.
Lower pH will not necessarily prevent the growth
of yeasts, but organic acids may prevent growth of
fig0008 Figure 8 Diagrammatic representation of budding in a typical
yeast cell. Reproduced from Spoilage: Fungi in Food – An Over-
view. Encyclopaedia of Food Science, Food Technology and Nutrition,
Macrae R, Robinson RK and Sadler MJ (eds), 1993, Academic
Press.
5520 SPOILAGE/Fungi in Food – An Overview

some yeasts. Sorbic acid and sorbates are particularly
effective in preventing yeast growth. Sulfur dioxide
can be used to kill wild-type yeasts in dried fruits and
in grapes for wine-making. Heating of dried fruits in
the package can also be used to kill yeasts and pas-
teurize the fruit.
Specific Genera of Yeasts
0042 There are a number of genera of yeasts that are im-
portant in foods. These include both beneficial and
harmful types. The following is a synopsis of 10
genera of yeasts that include the most important in-
dustrial yeast, and various spoilage yeasts, including
osmophilic yeasts, film yeasts, and asporogeneous
yeasts.
Saccharomyces
0043 Saccharomyces is the most important genus from an
industrial standpoint. The main species is S. cerevisiae
and different strains of it are used in many food indus-
tries for leavening bread and making ales, beer, wine,
alcohol, and other products. It has also been used to
produce single-celled protein, and food ingredients
such as autolyzed yeast. It is probably the single
most important yeast known. S. cerevisiae cells are
elliptical to ovoid in shape, with some spherical and
elongated cells as well. They reproduce by budding,
and form ascospores. S. cerevisiae can also cause
spoilage under certain conditions where its fermenta-
tive activity is not desired.
Zygosaccharomyces
0044 Zygosaccharomyces is considered to be a subgenus of
Saccharomyces, and some authors have reclassified
this genus as Saccharomyces. The yeasts in this
genus are osmophilic and can grow in high concen-
trations of sugar. They are responsible for spoilage of
molasses, syrups, honey, salad dressing, soya sauce,
and wine. They reproduce by budding, and form
ascospores.
Schizosaccharomyces
0045 Schizosaccharomyces species are found in sugar, mo-
lasses, honey, tropical fruit, and soil. They reproduce
by fission, forming arthrospores, or by sexual means,
forming ascospores. There are either four or eight
ascospores per ascus. The spores are oval, spherical,
or kidney-shaped.
Debaryomyces
0046 Yeasts in the genus Debaryomyces are film yeasts,
and they form pellicles of surface growth on meat
brines. They also grow on cheese and sausage
products. The cells of Debaryomyces are round to
oval-shaped and they form ascospores. They may
also form pseudomycelia.
Hansenula
0047Yeasts in the genus Hansenula are also film yeasts.
They are common in olive brines, fruit juice concen-
trates, citrus fruits, and grapes. Although they are
film yeasts that form pellicles, they are somewhat
fermentative. They produce spherical to oval elong-
ated cells. They reproduce by budding and by forming
ascospores. The ascospores have a characteristic
shape, similar to a bowler hat. They may form
pseudomycelia.
Pichia
0048Yeasts in the genus Pichia are also film yeasts that
form a pellicle on liquids, such as beer and wines.
Cells are oval to cylindrical-shaped and form asco-
spores which are either round or hat-shaped.
Torulopsis
0049The Torulopsis genus may also be called Torula and it
consists of asporogenous yeasts that are round to oval
in shape. They reproduce by budding. These yeasts
are common in nature, and contaminate many foods,
including refrigerated foods. Species of this genus can
ferment lactose and can spoil milk products, such as
sweetened condensed milk, as well as fruit juice
concentrates and acid foods.
Candida
0050Candida species are asporogenous yeasts that form
pseudomycelia or true mycelia. They are sometimes
categorized as Fungi Imperfecti in the family Mon-
iliaceae with other yeast-like genera, particularly
Trichothecium and Geotrichum. They reproduce by
budding and fragmentation of the mycelia into
blastospores. Members of this genus are common
in foods and occur in fresh meat, such as ground
beef, cured meats, and butter and margarine. They
can spoil foods high in acid and salt, and the lipo-
lytic species C. lipolytica can cause rancidity of
butter and margarine. Other species have been
grown for food and feed. At least one species, C.
albicans, is responsible for infections in humans and
animals.
Trichosporon
0051Trichosporon species are asporogenous yeasts that
reproduce by budding and form arthrospores. They
grow at lower temperatures and can be found in
many foods, including chilled beef, beer, and maple
sap.
SPOILAGE/Fungi in Food – An Overview 5521

Rhodotorula
0052 Yeasts in the genus Rhodotorula are sporogenous and
reproduce by budding. They sometimes also from
pseudomycelia. These organisms are widespread in
nature, often found in air and dust, and contaminate
many foods. They produce red, yellow, and pink pig-
ments and often discolor foods with red and yellow
spots on meats, particularly cured meats, and pink
areas in sauerkraut.
Health Implications
0053 Yeasts are generally not associated with foodborne
illnesses, but certain yeasts can cause infections in
humans and animals. Probably the most common
infection caused by a yeast is candidiasis, caused by
C. albicans. This organism is a common inhabitant of
the mucous membranes of the mouth, vaginal canal,
and intestinal tract. Yeast infections can occur most
readily in persons receiving broad-spectrum antibi-
otic therapy and those who are immunocomprom-
ised. In the immunocompromised subjects, Candida
species and other yeasts can cause endocarditis, urin-
ary infections, kidney infections, esophageal infec-
tions, and septicemia or systemic candidiasis. As the
number of immunocompromised persons increases
from radiation and chemotherapy, AIDS, and simple
aging, it remains to be seen if foodborne yeasts may
become a problem for these individuals as opportun-
istic infectious agents.
See also: Aflatoxins; Antibiotics and Drugs: Uses in
Food Production; Cereals: Dietary Importance; Cheeses:
Chemistry and Microbiology of Maturation; Controlled-
atmosphere Storage: Applications for Bulk Storage of
Foodstuffs; Essential Oils: Properties and Uses;
Fungicides; Lactic Acid Bacteria; Mushrooms and
Truffles: Classification and Morphology; Mycotoxins:
Classifications; Occurrence and Determination;
Toxicology; Water Activity: Effect on Food Stability;
Yeasts
Further Reading
Bullerman LB (2000) Mold Growth in Bakery Products and
its Prevention. AIB Technical Bulletin, vol. XXII, issue 5.
Manhattan, KS: American Institute of Baking.
Doyle MP, Beuchat LR and Montville JT (2000) Food
Microbiology: Fundamentals and Frontiers, 2nd edn.
Washington, DC: ASM Press.
Klich MA and Pitt JI (1988) A Laboratory Guide to the
Common Aspergillus Species and Their Telemorphs.
North Ryde, Australia: Commonwealth Scientific and
Industrial Research Organization.
Nelson PE, Toussoun TA and Marasas WFO (1983) Fusar-
ium Species. An Illustrated Manual for Identification.
University Park, Pennsylvania: The Pennsylvania State
University Press.
Northolt MD and Bullerman LB (1982) Prevention of mold
growth and toxin production through control of envir-
onmental conditions. Journal of Food Protection 45:
519–526.
Pitt JI (1988) A Laboratory Guide to Common Penicillium
Species, 2nd edn. North Ryde, Australia: Common-
wealth Scientific and Industrial Research Organization.
Pitt JI and Hocking AD (1999) Fungi and Food Spoilage,
2nd edn. Gaithersburg, MD: Aspen Publishers.
Rhodes M (ed.) (1979) Food Mycology. Boston: GK Hall.
Samson RA, van Reenen-Hoekstra ES, Frisvad JC and Fil-
tenborg O (1996) Introduction to Food-borne Fungi, 5th
edn. Baarn, The Netherlands: Centraalbureau voor
Schimmelcultures.
Molds in Spoilage
F Ottaviani, State University of Bologna, Italy
M G Ottaviani, CONSAL – Advising in Food
Microbiology, Sermide, Italy
Copyright 2003, Elsevier Science Ltd. All Rights Reserved.
Basic Biology of Molds
0001Molds are eukaryotic organisms widely distributed in
nature, and are common contaminants of agricultural
commodities, food, beverages, and feed.
0002In contrast to the simple (prokaryotic) bacteria,
molds are complex organisms whose mycelium is
composed of individual filaments termed hyphae.
The hypha is essentially a tube, consisting of a rigid
wall and containing a moving mass of protoplasm; its
interior is frequently divided into compartments by
cross-walls which, being incomplete or perforated, do
not separate the content into individual cells. This
coenocytic condition gives molds an advantage over
the single-celled bacteria, permitting both a more
efficient distribution of nutrients and a better dilution
of external stresses over a large number of cells. Or-
ganelles inside the hypha are of the eukaryotic type,
i.e., more sophisticated than those of the bacterial
cell. Nuclei and energy-yielding mitochondria are
present.
0003The hyphal wall is predominantly composed of
polysaccharides which give the molds a more defined
barrier against environmental stress.
0004Chitin, a polymer of N-acetyl-d-glucosamine, is
the more frequently occurring polymer in the walls
of fungal spores and mycelia and it does not exist in
bacteria or most uncontaminated food and feed prod-
ucts. Glycoproteins such as galactomannan are either
5522 SPOILAGE/Molds in Spoilage

extracellular or bound to the fungal cell wall. Ergos-
terol is considered the principal sterol of fungi and
plays an important role in cell membranes.
0005 All these specific components of fungi are used to
estimate fungal activity by rapid chemical, biochem-
ical, and immunological methods even when fungal
biomass may have been killed, given that chitin and
fungal polysaccharidic antigens are heat-resistant and
they can survive after the death of fungal species.
0006 Because of their peculiar structure, molds behave in
a different way from bacteria, with regard both to
food preservation technologies and to laboratory
analytical techniques – they have better control of
ionic homeostasis (with regard to pH and water ac-
tivity (a
w
)), and insensitivity to antibiotics commonly
used to repress bacterial growth.
0007 With generation times ranging between 2 and 5 h,
fungal hyphae extend at their apices with their oldest
parts producing branches which fuse between adja-
cent hyphae. Branched vegetative hyphae form a
vegetative mycelium which appears as a light, ill-
defined spot on the food or culture medium surface
at the beginning of the spoilage or during the initial
stage of the colony development. Later, specialized
cells emerge from the flat hyphal mass bearing at
their ends the staminal components of the molds.
These constitute the reproductive mycelium of the
mold, whose cells are usually called conidiophores
and staminal components either conidia, if asexually
formed, or spores, if sexually generated. Sexual for-
mation of spores, which involves genetic rearrange-
ments, greatly contributes to the mold’s survival in
nature, but it is seldom observed in food and culture
media where asexual conidia are more likely to
appear. Sexual spores are frequently thick-walled
and able to withstand severe environmental condi-
tions, including heat processing, as in the case of the
Byssochlamys sexual spores which may spoil fruit
juices. In common with conidia, which possess a thin-
ner wall, spores differ from vegetative cells in having
a thicker tegument, a poorly differentiated cytoplasm,
and a low water content, resulting in the ability to
survive when environmental conditions become
lethal to living cells.
0008 Resting cells, which can withstand many adverse
conditions such as drying or oxygen deprivation, may
later restore their normal metabolic pathways, even-
tually giving rise to the first hyphal cell and so com-
mencing the mold life cycle when availability of
water, or air, or any other condition suitable to
growth, occurs.
0009 Nutrient requirements for mold growth are very
simple. A suitable organic carbon source is necessary
(this can be simple, such as glucose, or very complex,
like cellulose and hemicellulose composing the plant
cell wall). Many molds grow on poor substrates
containing only inorganic nutrients, e.g., inorganic
nitrogen or phosphorus. This explains why molds
may actively grow where bacteria cannot proliferate,
such as on the apparently hostile substrates of walls,
wood, fabrics, etc.
0010Many molds produce energy by reoxidation of
nicotinamide adenine dinucleotide/nicotinamide
adenine dinucleotide phosphate (NAD/NADP) co-
enzymes along an electron transport chain where an
inorganic compound serves as a terminal electron
acceptor (respiration); usually oxygen is the final ac-
ceptor, so molds need an aerobic environment for
their growth, but many common strains (e.g., asper-
gilli) use anaerobic respiration with nitrogen ions
as electron acceptors. Byssochlamys and a very few
other molds may break down sugars by fermentation,
in the complete absence of oxygen, initially producing
carbon dioxide, which can cause visible swelling of a
package. Growth of proteolytic and lipolytic strains
of molds is responsible for both the ripening and
distinctive flavor of the internally or surface mold-
ripened cheese and off-flavors and off-odors resulting
from fungal lipase activity, with an increase in the free
fatty acid (FFA) values.
0011Spoilage refers to any change that occurs in food
resulting in reduction in sensory quality, i.e., changes
in color, flavor, texture, or aroma brought about by
the growth of microorganisms in food and feed. De-
gradative enzymes, such as pectinases and cellulases,
are primarily responsible for degradation of the
main structural components of vegetables, fruits,
and legumes and related transformed commodities,
pectin, and cellulose.
0012Flavor-volatile metabolites produced by molds,
such as 3-methyl-butanol, 3-octanone, 3-octanol, and
many others, cause adverse changes in flavor and
aroma.
0013Besides the primary metabolic pathways supplying
the fungal cells with energy and cellular metabolites
necessary for growth, molds may show a range of
unusual metabolic processes (grouped as secondary
metabolism). These are not concerned with survival
and growth and take place when normal development
is restricted or the substrate composition is unbal-
anced (e.g., a very high carbon to nitrogen ratio, as
in the case of plant tissues). Some secondary metabol-
ites are of great commercial value (e.g., antibiotics
like penicillins) but others, such as mycotoxins, pose
a health risk to the consumer in crops and other plant
products. (See Mycotoxins: Occurrence and Deter-
mination.) Mold spoilage is so undesirable not solely
because of the resulting unesthetic changes, but also
because it may constitute a hazard to human and
animal health.
SPOILAGE/Molds in Spoilage 5523

Growth Requirements of Molds and
Effects of Processing
0014 The metabolism of molds, like that of other organ-
isms, is affected by environmental conditions which
may increase or decrease the rate of enzymatic reac-
tions. Physical and chemical conditions which play a
major role in controlling the mold’s metabolism and
growth are temperature, water availability, and ion
content.
0015 The nutritional, physical, and chemical attributes
of a given food substrate may select molds for pre-
dominant growth, these being the most highly
adapted organisms, e.g., to reduced water content in
high ionic strength (high acidity). Moreover, organ-
isms can survive to some degree in conditions that do
not allow growth. Spores and, to a lesser extent,
conidia may tolerate extreme conditions in food and
environment, retaining for extended times their abil-
ity to develop a mold when the substrate becomes
favorable again. Mold propagula held on a frozen
food may later undergo germination when mixed at
room temperature with other ingredients. Although
usually repressed by the high osmotic pressure (low
a
w
) in sugar-preserved or dried commodities, such as
in preserves in tanks for industrial use or in pasta in
its packaging, spores may give rise to a visible spoil-
age when water condenses at the mass surface.
0016 Most fungi are mesophilic, so they grow at moder-
ate temperatures, in the range 10–40
C. Lowering
the temperature decreases the rate of metabolism
and is often used to slow deterioration. A few fungi
grow at very low temperature. Cladosporium and
Thamnidium, sometimes colonizing animal hairs,
are important as spoilage agents of improperly
dressed meat that is held for long enough (usually
more than a month) at subzero temperatures above
10
C.
0017 Important intrinsic factors which influence the
microflora that develops in food and feed include
pH and a
w
.
0018 Fungi show a wide range of behavior with respect
to water availability, generally being more tolerant of
water stress than other organisms, including the ma-
jority of yeasts. The lower limit for growth of molds,
expressed as a
w
, vary from 0.93 for destructive rots of
fruits and vegetables like Rhizopus and Mucor;to
0.87 for toxigenic fusaria; to 0.83 for agents of
surface discoloration in fresh cheese and sausage
like Penicillum expansum or P. viridicatum. Xerophi-
lic fungi are characterized as being capable of growth
below an a
w
of 0.85 and are most commonly associ-
ated with intermediate-moisture food such as dried
meats, seafood, fruit concentrates, jellies, jams, pre-
serves, and syrups, spices, nuts, and cereals, where
their effective growth range can be as low as 0.65.
Conditions of reduced a
w
lower the final fungal bio-
mass produced because of reduction of the growth
rate and of increase of the time required for spore
germination up to 3 months or more. Properly dried
and stored commodities are inherently resistant to
spoilage due to their low a
w
; however, temperature
differentials, such as in bulk-stored grains, may lead
to moisture condensation and possible growth of
xerophilic molds. The majority of xerotolerant fungi
belong to genera Penicillium and Aspergillus, includ-
ing its perfect forms such as Eurotium (Aspergillus
glaucus), Emericella, and Wallemia sebi.(See Water
Activity: Effect on Food Stability.)
0019Media and diluents with reduced a
w
are necessary
for enumerating xerophilic fungi, otherwise vegeta-
tive cells and spores may be lysed by osmotic shock or
remain dormant when exposed to the relatively high
a
w
associated with the conventional diluents and
media used in microbiological analysis. Addition of
humectants, such as 50% glucose to the diluent, and
18% glycerol or 50% glucose to traditional media,
especially malt extract agar, lowers a
w
levels to about
0.94 and 0.92 respectively. Extended incubation
periods, such as 4–6 weeks at room temperature
instead of the traditional 5 days time, are required
for recovery.
0020Molds, like yeasts, tolerate a greater pH range than
bacteria do. Many fungi can grow over a wide pH
range of at least 3–8, while pH values below about 5
negatively affect the metabolism and growth of bac-
terial cells. Where the food substrate shows a high a
w
,
it is the acidic environment which plays a decisive role
in the competition between bacteria and filamentous
fungi. As fruits show pH values of 4.6 or lower, mold
growth is more likely. Spoilage thus arises from the
intense activity of pectic and proteolytic fungal
enzymes that have pH optima between 2 and 5. (See
pH – Principles and Measurement.)
0021Because of the the acido-tolerant behavior of
molds, originally food mycologists relied on low-pH
media, such as acidified potato dextrose, Sabouraud,
or similar agars to detect fungi in clinical and food
specimens. However, in food samples acid pH may
adversely affect mold spore germination and give rise
to protein coagulation with the formation of artifacts,
and, therefore, it is more effective to use media of
neutral pH in combination with antibiotics. Anti-
biotics, such as chloramphenicol, gentamicin, and
others, are well-known biomolecules perturbing the
cell wall of bacteria which is different from that of
fungi.
0022Molds may alter the pH of the environment in
which they grow, by producing ammonium ions
that neutralize the substrate acidity and thus allowing
5524 SPOILAGE/Molds in Spoilage

(re)growth of normally repressed, dangerous
organisms. Such an example of metabiotic associ-
ation is given by the persistence of Listeria in cheese
ripened by selected molds. (See Listeria: Properties
and Occurrence.)
0023 Molds are aerobic organisms in that they require
oxygen and/or a food substrate with a high oxidation-
reduction potential (E
h
) for growth. Oxidized foods,
such as fruits with E
h
values betwee0n þ300 and
þ400 mV, support the growth of molds well, while
reduced ones, like yogurt with its E
h
value of about
100 mV, do not. In the latter case, molds may de-
velop a mycelium only at the surface in contact with
the air. Some fungi (e.g., strains of Paecylomyces
and Penicillium) are microaerophilic, growing in an
atmosphere containing only 0.1–0.5% oxygen. A few
species, such as Fusarium oxysporum, may show a
very delayed growth in the complete absence of
oxygen. It would appear, therefore, that the degree
of anaerobiosis attained under commercial processing
conditions is not always adequate to prevent com-
pletely the growth of molds.
0024 There is evidence that raising the level of carbon
dioxide in the atmosphere above 70–80% greatly
reduces the growth rate of most molds, with some
notable exceptions, such as Penicillium roquefortii
or Byssochlamyis nivea.
0025 Growth of most molds is restricted – sometimes
inhibited – by the use of preservatives at the legally
permitted concentrations (between 0.1 and 0.2%)
The acids and salts commonly used are propionic,
sorbic, and benzoic. Propionates are added to bakery
products and to certain processed cheeses. Benzoic
acid and its sodium salt are most widely used in
highly acidic foods such as fruit products and carbon-
ated beverages. The undissociated acid is the active
molecule and therefore the effectiveness increases
with decrease in pH while the organic acids exhibit
little fungistatic activity in neutral conditions. Inhib-
ition of growth by weak acid preservatives is due to a
number of actions, including membrane disruption,
inhibition of essential metabolic reactions, stress on
intracellular pH homeostasis, and accumulation of
toxic anions. Natamycin (pymaricin), a natural anti-
mycotic compound produced by Streptomyces nata-
lensis, is commercially available for the surface
treatment of cheese. In recent years, strains of preser-
vative-resistant molds (particularly aspergilli) have
been reported which tolerate high (0.5–1.5%) con-
centrations of organic acids due the induction of
an active transport system that pumps undissociated
acid molecules out of cells. The enzymatic de-
gradation of sorbic acid to pentadien by certain
fungal species is well documented. (See Preservation
of Food.)
0026Most molds possess little heat resistance, and are
easily destroyed when food with a
w
values above 0.93
are heated at pasteurization temperatures. Vegetative
cells and asexual propagules behave in the same
manner. Heat resistance of molds is enhanced in the
presence of high levels of solutes such as salt and
sugar. The decimal reduction time for Aspergillus
niger conidia heated at 100
C increases from about
1 ms in a moist substrate (a
w
¼0.98) to about 1 s in
intermediate-moisture substrates (a
w
between 0.85
and 0.90, such as cakes and most oven-processed
sweet products), and to 10 s when a
w
falls below
0.85 because of the addition of sugar (e.g., in fruit
preserves). In the latter products, the low pH, usually
between 3.2 and 3.5, partially counteracts the pro-
tective effect of sugar, thus decreasing the resistance
of mold to heat.
0027The responses of molds to the presence of sodium
chloride (salt) is somewhat different from that ob-
served with sucrose. Protection against heat inactiva-
tion of molds is increased even at lower a
w
values,
usually below 0.97, i.e., a salt concentration of about
3%. This should be kept in mind when setting up
pasteurization procedures for brines used in cheese
and sausage manufacture.
0028A few molds produce heat-resistant ascospores or
sclerotia. Ascomycetous species are widely distrib-
uted in soil and cause spoilage in heat-processed
food products containing fruits which have been ex-
tensively contaminated with soil. Ascospores pro-
duced by the two species of Byssochlamys, B. fulva
and B. nivea, and by Neosartorya fischeri are the
most resistant of all and they are typically associated
with spoilage of thermally processed fruit products,
mainly canned, where the can center temperature is
raised up to 80–90
C. The decimal reduction time of
Byssochlamys nivea spores at 80
C increases from
about 35 min (a
w
¼0.95) to about 4 h in neutral
foods with a
w
values between 0.85 and 0.86. Neosar-
torya fischeri var fischeri has a D
85
(decimal reduc-
tion time at 85
C) value of 6–10 min with a Z value
(Thermal Death Time curve’s slope) of 5–7
C while
the spinosa variety is more heat-resistant (D
85
value of 10–96 min with a Z value of 5–7
C). Symp-
toms observed with spoiled canned fruits include
breakdown of fruit texture, off-odors, gas, and
visible mold growth. (See Heat Treatment: Chem-
ical and Microbiological Changes; Pasteurization:
Principles.)
Classification of Molds
0029Only a small proportion of the thousands of genera
and species of molds identified in nature are of any
significance in foods.
SPOILAGE/Molds in Spoilage 5525
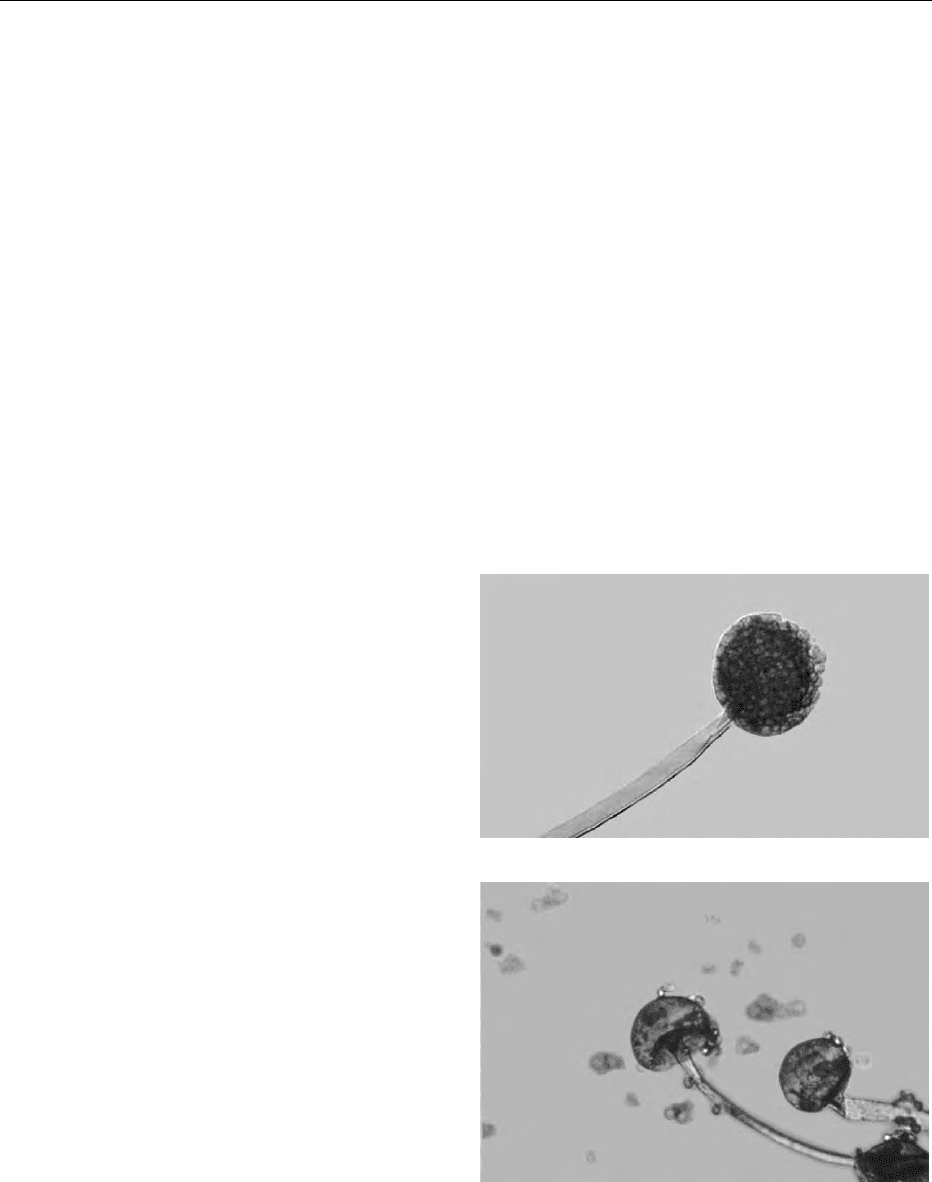
0030 Ranked taxonomically in the separate kingdom of
Fungi, molds are subdivided into further taxonomic
categories on the basis of ecological behavior, and
gross colony and microscopic morphology.
0031 The most important diagnostic factor relies on the
morphology of the asexual reproduction system
(the so-called anamorphic stage of the mold), best
observed under the microscope in slide-mounted
cultures. Spores formed through sexual reproduction
(the so-called telemorphic stage) are rarely used in
identifying foodborne molds, given the difficulty of
inducing gametogenesis and sexual fusion in vitro.
0032 Molecular tools based on the polymerase chain
reaction (PCR) may be applied in food mycology for
diagnostic purposes. The noncoding ITS (Internal
Transcribed Spacer) region of the rDNA unit, and
other genes coding for metabolic and structural func-
tions (e.g., the b-tubulin gene), have been investigated
to clarify the genetic structure of some species.
0033 Three subkingdoms include genera of significance
in food spoilage: Zygomycotina, Ascomycotina, and
Deuteromycotina. Most fungi, within the subking-
dom Zygomycotina, belong to the class Zygomycetes,
which includes, in the order Mucorales, three phyto-
pathogenic genera frequently involved in the spoilage
of moist wheat, flour, and cereal products: Absidia,
Mucor, and Rhizopus. Distinctive features of these
molds are (1) rapid growth when water availability
is above the limiting value of 0.92–0.93, including
filling the whole Petri dish with fine white to greyish
to blackish aerial mycelium, and (2) asexual spore
formation by cytoplasmic cleavage within a large
globose cell termed sporangium, from where sporan-
giospores are liberated by breakage or deliquescence
of the wall. Sporangia are located at the end of a
supporting hypha (sporangiophore), from where
they are separated by a septum forming a protrusion
called columella (Figure 1).
0034 A distinctive feature of the subkingdom Ascomy-
cotina is the development of sexual spores in a cell
(sporangium), more specifically termed the ascus
(bag). Asexual spores, called conidia, develop in
various ways directly from reproductive hyphae.
Ascospores of Byssochlamys species show a very
distinctive heat resistance.
0035 The subkingdom Deuteromycotina is used mainly
for convenience to accommodate all those fungi that
seldom, if ever, produce sexual stages. In all other
respects, however, these fungi are similar to the Asco-
mycotina, and where sexual stages are discovered
they are usually found to belong to the Ascomycotina.
In Deuteromycotina systematics the teleomorph
(perfect, sexual stage) name should preferentially
be used to indicate the species. Unfortunately, the
anamorphic name is firmly rooted in the literature
and in common parlance. The teleomorphic stage
cannot usually be seen in routine examination
of fungi unless special cumbersome techniques are
used, so the anamorph name is commonly retained:
Aspergillus, instead of its teleomorphs Eurotium and
Emericella; Penicillium, not the teleomorphs Eupeni-
cillium and Talaromyces, and so on.
0036The morphology of the fertile hypha is again the
most important factor in taxonomic characterization.
During development of the mold, asexual propaga-
tion occurs which starts with conidiogenesis: asexual
propagula, named conidia, are produced from the
mother cell (conidiogenous cell). The system com-
posed of a conidiogenous cell and its conidia is
known as a conidiophore. Conidiogenous cells are
sometimes supported by mono- or pluricellular stipes
whose shape may help in mold identification. Differ-
entiation of the conidiophore and conidial arrange-
ments serve as diagnostic features, and we may
distinguish the following:
(a)
(b)
fig0001Figure 1 (a) Large, round sporangium of Rhizopus oryzae.
(b) After disintegration of the sporangial wall, small clusters of
sporangiospores are released, leaving a naked umbrella-shaped
columella. Reproduced from Spoilage, Encyclopaedia of Food Sci-
ence, Food Technology and Nutrition, Macrae R, Robinson RK and
Sadler MJ (eds) 1993, Academic Press.
5526 SPOILAGE/Molds in Spoilage