Бейли Н. Математика в биологии и медицине
Подождите немного. Документ загружается.


ГЕНЕТИЧЕСКОЕ
СЦЕПЛЕНИЕ
И
ХРОМОСОМНЫЕ
КАРТЫ
241
легко видеть, что при отсутствии отклонений в жизнеспособности
соответствующая оценка частоты рекомбинаций у имеет вид
у = для фазы сцепления |
или
" !• (Ю.1)
у = для фазы отталкивания.
Оценка
дисперсии в обоих
случаях
имеет вид
o-
2
G/)=
V{i
~
y)
• (Ю.2)
Разумеется, можно начать с того, чтобы проверить, имеется
ли статистически значимое отклонение от простейшей гипотезы,
предполагающей
отсутствие
влияния рекомбинаций или различий
в
жизнеспособности. При у =
1
/
2
можно ожидать, что все четыре
класса фенотипов, рассматриваемые в табл. 10,
будут
встречаться
одинаково часто. Легко показать, что критерий %
2
с тремя степе-
нями
свободы можно разбить на три независимые компоненты:
%
2
А
— для проверки соотношения при расщеплении аллелей
пары Аа; %
в
— для проверки соотношения при расщеплении
аллелей пары В, b; %
2
L
— для проверки на сцепление:
л,2
л,
2 _ {а —
Ъ
— t , ..,
%
Ь
~ п
•
J
Каким
именно критерием для проверки значимости
следует
поль-
зоваться, зависит от того, какой информацией мы располагаем
вначале. Если известно, что лишь одно из соотношений откло-
няется
от ожидаемого, то можно использовать критерий %
2
L
для
проверки существования сцепления, но если известно, что откло-
няются оба соотношения, то нужно использовать описанный ниже
другой
критерий.
Допустим далее, что имеются три пары локусов и
соответствую-
щие пары генов А, а; В, Ъ и С, с. Сцепление
между
генами А я В
(с
частотой рекомбинаций г/j) можно обнаружить с помощью
возвратных скрещиваний, затрагивающих только эти два факто-
ра; сцепление
между
генами В и С (с частотой рекомбинаций у
2
)
можно обнаружить с помощью
другой
группы экспериментов,
в
которых затрагиваются только эти факторы. Что можно сказать

242
ГЛАВА
10
о
порядке расположения этих
трех
локусов? Ничего, если
у нас
нет данных
о
наличии сцепления
между
генами
А и С.
Таким
образом,
нам
нужна третья группа скрещиваний. Положим,
что
соответствующая частота рекомбинаций равна
у
3
.
Допустим теперь,
что
истинный порядок расположения локу-
сов
— АВС.
Легко показать,
что
между
генами
А и С
произойдет
рекомбинация,
если
она
произойдет
между
генами
А и В, но не
между
генами
В я С,
ж
наоборот. Если допустить,
что
рекомбина-
ция
в
соответствующих
двух
участках происходит независимо,
то
у
3
находится
по
формуле Троу:
Уз
= г/i
(1
—
г/г)
+ (1 — г/0 г/
2
= y
i
-j- г/
2
—
2у^
2
.
(10.4)
Очевидно,
что это
правило
для
частот множественных рекомбина-
ций
не
является аддитивным (чего следовало
бы
ожидать
на
основе
материала, рассмотренного
в
конце предыдущего раздела). Если
данных
не
очень много,
а
статистическая изменчивость велика,
то нелегко определить,
будет
ли
соотношение
(10.4)
более вероят-
но,
чем два
других,
соответствующих случаям
АСВ и ВАС.
Если
эти
три
локуса располагаются близко
друг
к
другу
и все три
частоты рекомбинаций малы,
то
справедливы следующие прибли-
женные равенства:
гУз
~ г/i +
г/г, порядок
АВС;
Л
гУ2~г/1
+ г/
3
,
порядок
ВАС; >
(10.5)
г/i
~
гуг
+
г/з, порядок
АСВ. J
Однако лучше определять порядок расположения локусов
на
потомстве, полученном
от
тригибридного возвратного скре-
щивания
типа
ABCIabc
X
abclabc.
Это
тригибридное скрещивание
в
фазе сцепления.
Не
представляет
труда
вычислить математиче-
ские
ожидания числа потомков
с
различными фенотипами.
Для
порядка
АВС эти
математические ожидания приведены
в
табл.
11.
Таблица
11
Тригибридное возвратное скрещивание со сцеплением
Классы фенотипов
Математическое
ожидание
для
ABC/abcXabc/abc
Наблюдаемое
чис-
ло
АВС,
abc
ге
(1—У])
(1 —г/2)
а
аВС
Abc
Щ\
(1—г/г)
Ъ
АВс,
abC
ге
(1—г/1)г/г
с
аВс,
АЬС
п
У\Уг
d
Всего
ге
п

ГЕНЕТИЧЕСКОЕ
СЦЕПЛЕНИЕ
И
ХРОМОСОМНЫЕ
КАРТЫ 243
Заметим, что мы объединили здесь комплементарные классы,
т. е. классы типа ABC и abc, для которых математические ожида-
ния
одинаковы. Аналогичным способом можно получить соот-
ветствующие результаты и для
трех
других
типов скрещивания.
Максимально правдоподобные оценки для у^ и у
2
, а также оценки
их дисперсий в этом
случае
имеют вид
(Ю.6)
Если наше предположение о том, что ABC — истинный поря-
док следования локусов, верно, то число потомков с фенотипом
(аВс, АЬС)
будет
наименьшим; оно имеет математическое ожида-
ние
пуху
2
и отражает рекомбинацию в обоих
участках
АВ и ВС
одновременно. Обычно при тригибридном возвратном скрещива-
нии
нетрудно установить таким способом наименее многочислен-
ный
класс фенотипов и определить вероятный порядок располо-
жения локусов. Так, если окажется, что наименьшее число потом-
ков
относится к классу (АВс, аЪС), то искомым порядком локусов
должен быть порядок АСВ.
Существуют, однако, и некоторые
другие
преимущества при-
менения
полигибридного возвратного скрещивания по сравнению
с попытками комбинировать результаты, полученные в нескольких
отдельных экспериментах с дигибридным скрещиванием. Эти пре-
имущества станут ясны, если
учесть
осложнения, связанные,
в частности, с дифференциальной жизнеспособностью, которую
мы сейчас и рассмотрим для случая простого дигибридного воз-
вратного скрещивания; более сложные эксперименты с тригибрид-
ным
скрещиванием мы обсудим в разд. 10.3.
Дифференциальная
жизнеспособность
Очень часто различные генотипы или фенотипы характери-
зуются различной жизнеспособностью, и во многих
случаях
важ-
но
учитывать этот фактор. Допустим, что относительная жизне-
способность фенотипа А по сравнению с фенотипом а равна и и что
соответствующая величина для фенотипа В по сравнению с
фено-
типом Ь равна v. Если величины и и v независимы
друг
от
друга
и
действуют
мультипликативно, то математические ожидания
числа потомков с различными фенотипами при дигибридном
возвратном скрещивании в фазе сцепления, представленные
в табл. .10, изменятся и примут значения, приведенные в табл. 12„

244
ГЛАВА
10
где делитель D определяется формулой
(10.7)
Эта таблица имеет три степени свободы, что позволяет оценить
три неизвестных; легко найти, что соответствующие максимально
правдоподобные оценки выражаются следующим образом:
где
У
=•
и =
(be)
1
ab
\ V2
ill
. _L
4-
—
4-
—
(10.8)
Если
имеются данные только по скрещиванию в фазе сцепления
(или
только в фазе отталкивания), то ни одной степени свободы
для проверки модели у нас не остается; но если мы располагаем
Таблица
12
Дигибридное возвратное скрещивание
в
фазе
сцепления
при
наличии
дифференциальной жизнеспособности
Классы
фенотипов
Математическое ожида-
ние
для
АВ/аЪхаЪ/аЬ
Наблюдаемое число
АВ
nuv
(i—y)
D
а
Ab
пиу
D
Ъ
аВ
nvy
D
с
ab
ге(1-г/)
D
d
Сум-
ма
n
n
данными
по
обеим фазам,
то
можно произвести проверку
с
помощью
критерия
%
2
.
Нетрудно провести проверку
на
сцепление, допустив,
что
оба моногибридных расщепления нарушены.
Так,
если
мы при-
нимаем нулевую гипотезу
у = у, то из
табл.
12
видно,
что при-
веденные
в ней
математические ожидания соответствуют
тем,
которые получаются
из
таблицы сопряженности признаков
2x2

ГЕНЕТИЧЕСКОЕ
СЦЕПЛЕНИЕ И ХРОМОСОМНЫЕ КАРТЫ 245
с независимыми рядами и столбцами. Эта таблица имеет вид
Ъ
(10.9)
d
В этом
случае
можно применять обычный критерий %
2
или точ-
ный
критерий значимости.
Однако вполне возможно, что величины и и v не
будут
неза-
висимы
друг
от
друга,
и поэтому
лучше
принять допущение
Таблица
13
Сбалансированное
возвратное
скрещивание
при
наличии
дифференциальной
жизнеспособности
Классы фенотипов
Математическое ожидание для
Ab/abxablab
Наблюдаемое число
Математическое ожидание для
Ab/aBxab/ab
Наблюдаемое число
АВ, ab
nia(i-y)
a(i — y) + y
а
п
2
ау
ау + {1 —
У)
с
АЬ, аВ
щу
a(i—y) + y
Ъ
«2(1 —г/)
щЛ-(\—у)
А
Сумма
ГЦ
щ
п
2
п
2
о том, что относительная жизнеспособность
двух
классов роди-
тельских особей АВ и ab по сравнению с жизнеспособностью
классов рекомбинантов АЬ я аВ равна, скажем, а. В этом
случае
важно иметь данные по скрещиванию как в фазе сцепления, так
и
в фазе отталкивания (сбалансированное скрещивание).
Допу-
стим, что получены величины, приведенные в табл. 13.
На
этот раз нужно получить оценки лишь для
двух
параметров,
и
имеются как раз две степени свободы — по одной для данных
по
каждой фазе. Максимально правдоподобные оценки имеют вид
у =•
(be)
1
<*'$)=•
а—
(10.10)
где
h а т Ъ • с
246
ГЛАВА
10
Формально
результаты
(10.8)
и
(10.10)
очень похожи, хотя они
имеют различную интерпретацию.
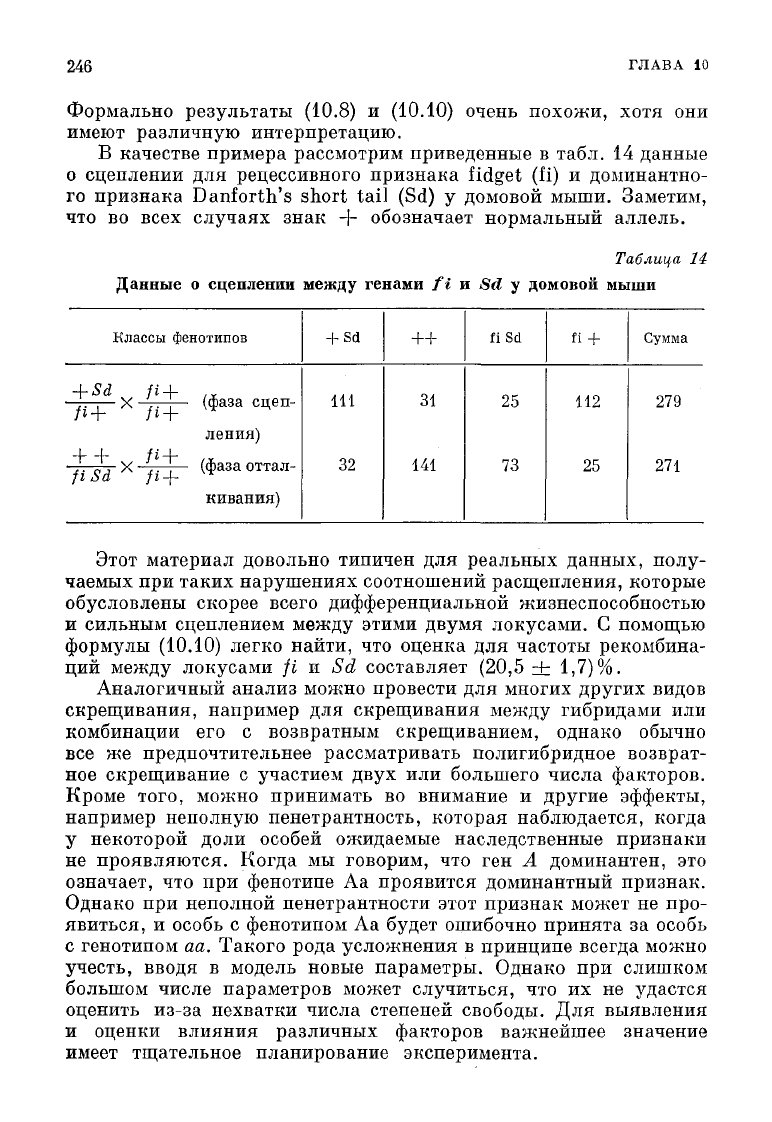
В качестве примера рассмотрим приведенные в табл. 14 данные
о
сцеплении для рецессивного признака
fidget
(fi) и доминантно-
го признака Danforth's short tail (Sd) у домовой мыши. Заметим,
что во
всех
случаях знак -f- обозначает нормальный аллель.
Таблица
14
Данные
о
сцеплении
между
генами
fi и Sd у
домовой
мыши
Классы фенотипов
+
Sd fi+ ,_,
ления)
кивания)
+
Sd
111
32
-н-
31
141
fi
Sd
25
73
fi +
112
25
Сумма
279
271
Этот материал довольно типичен для реальных данных, полу-
чаемых при таких нарушениях соотношений расщепления, которые
обусловлены скорее всего дифференциальной жизнеспособностью
и
сильным сцеплением
между
этими двумя локусами. С помощью
формулы
(10.10)
легко найти, что оценка для частоты рекомбина-
ций
между
локусами fi и Sd составляет (20,5 ±
1,7)%.
Аналогичный анализ можно провести для многих
других
видов
скрещивания,
например для скрещивания
между
гибридами или
комбинации
его с возвратным скрещиванием, однако обычно
все же предпочтительнее рассматривать полигибридное возврат-
ное
скрещивание с участием
двух
или большего числа факторов.
Кроме
того, можно принимать во внимание и
другие
эффекты,
например
неполную пенетрантность, которая наблюдается, когда
у некоторой доли особей ожидаемые наследственные признаки
не
проявляются. Когда мы говорим, что ген А доминантен, это
означает, что при фенотипе Аа проявится доминантный признак.
Однако при неполной пенетрантности этот признак может не про-
явиться,
и особь с фенотипом Аа
будет
ошибочно принята за особь
с генотипом аа. Такого рода усложнения в принципе всегда можно
учесть, вводя в модель новые параметры. Однако при слишком
большом числе параметров может случиться, что их не удастся
оценить
из-за нехватки числа степеней свободы. Для выявления
и
оценки влияния различных факторов важнейшее значение
имеет тщательное планирование эксперимента.

ГЕНЕТИЧЕСКОЕ
СЦЕПЛЕНИЕ
И ХРОМОСОМНЫЕ КАРТЫ 247
10.3. СБАЛАНСИРОВАННОЕ
ТРИГИБРИДНОЕ
ВОЗВРАТНОЕ
СКРЕЩИВАНИЕ
В предыдущем разделе мы убедились в значительных преиму-
ществах проведения анализа на сцепление
двух
факторов методом
сбалансированного возвратного скрещивания, поскольку здесь
требуются менее строгие допущения о характере влияния диф-
ференциальной
жизнеспособности. Если рассматривается порядок
расположения нескольких локусов, то в экспериментах с три-
гибридным возвратным скрещиванием
легче
определить истинный
порядок
их расположения, чем при изучении
трех
отдельных
дигибридных возвратных скрещиваний (хотя мы рассматривали
лишь
тот случай, когда влияние дифференциальной жизнеспособ-
ности
отсутствует). Существуют также и некоторые
другие
аргу-
менты в пользу тригибридных (или
даже
полигибридных) скрещи-
ваний.
Так, хотя частота рекомбинаций и является довольно
постоянной
величиной, у разных пород животных и в разных
условиях среды она может слегка варьировать. Поэтому
лучше исследовать как можно больше параметров в пределах
одного эксперимента. Кроме того, возникают большие сложности,
когда точки обмена не распределены вдоль хромосомы случайным
образом, т. е. имеет место интерференция. В этом
случае
соотно-
шение
(10.4)
уже не справедливо и
труднее
описать частоты различ-
ных типов рекомбинаций; поэтому здесь необходимо проводить
эксперименты
с использованием нескольких сцепленных факторов.
Многие
доводы в пользу такого рода экспериментов очень близки
к
доводам в пользу факторных экспериментов при агрономических
полевых испытаниях или
других,
по смыслу аналогичных иссле-
дованиях.
Полный
математический анализ
даже
тригибридного возврат-
ного скрещивания сопряжен с рядом трудностей (см.
другую
книгу автора [7], разд. 4.5), хотя интерпретация получаемых
результатов с точки зрения генетики вполне возможна. Рассмот-
рим
оценку параметров для одной модели по
методу
Р. Фишера.
Как
и ранее, допустим, что имеются три пары локусов А,а,
В,Ъ и С,с, и предположим, что истинным порядком локусов
является ABC. При образовании гамет рекомбинации
могут
вообще не происходить, происходить только на первом участке АВ,
только на втором участке ВС или на обоих участках одновременно.
Пусть вероятности каждого из этих четырех возможных случаев
равны
соответственно р, q, r и s, где
p-\-q-{-r-}-s=l.
Мы не
принимаем
здесь допущения о том, что рекомбинации на обоих
участках происходят независимо
друг
от
друга.
Ясно, что частоты
рекомбинаций
между
генами А и. В и
между
генами В ж С равны

248
ГЛАВА
10
соответственно
(10.11)
и
мы не предполагаем, что в общем
случае
s =
Если
мы хотим получить полностью сбалансированные дан-
ные,
нам необходимы данные по каждому из четырех тройных
гетерозигот:
ABC/abc, аВС/Abc,
АВс/abC
и
аВс/АЬС.
Пусть число
потомков,
полученных при этих четырех типах скрещивания,
равно соответственно п
и
п
2
, п
3
и n
k
. Математические ожидания
в
случае
тригибридного возвратного скрещивания в фазе сцепле-
ния
при отсутствии дифференциальной жизнеспособности приводи-
лись в табл. 11. Допустим, что средние значения относительной
жизнеспособности для четырех пар фенотипов, встречающихся
в
потомстве, равны соответственно и, v, w к t, где и 4- v 4- w 4-
+
t = 4. Тогда математические ожидания и наблюдаемые значе-
ния
для всей рассматриваемой совокупности сбалансированных
возвратных скрещиваний можно записать в виде, показанном
в
табл. 15, где приняты следующие обозначения:
D
i
= pu
-\-qv-\-
rw-\-st,
~\
Do = vv 4- аи 4- rt -f- sw, \
[ (10 12)
D
s
= pw 4- qt 4- ru 4- sv, \
D
t
= pt
-j-gw-fry
+ su. J
Таблица
15
Сбалансированное тригибридное возвратное скрещивание
при
наличии дифференциальной жизнеспособности
ABC Математическое ожидание
abc Наблюдаемое число
аВС Математическое ожидание
Abe Наблюдаемое число
АВс Математическое ожидание
аЬС Наблюдаемое число
аВс Математическое ожидание
АЬС Наблюдаемое число
Наблюдаемая сумма
Отсутст-
вие
ре-
комбина-
ций
a
i
n
2
pv/D
2
n
3
pw/D
3
«l
4
' dl
P
Рекомби-
нации
только
на
первом
участке
n
i
qvjD
i
а
2
n
2
qu/D
2
n
3
qt/D
3
n
i
qw/D
i
Q
Рекомби-
нации
только
на
втором
участке
«3
n
2
rt/D
2
n
3
rujD
3
R
Рекомби-
нации
на обоих
участках
n
i
st/D
i
Ч
n
2
sw/D
2
n
3
su/D
3
Ч
S
Сум-
ма
«1
"1
«2
«3
«3
П
4
N

ГЕНЕТИЧЕСКОЕ
СЦЕПЛЕНИЕ
И ХРОМОСОМНЫЕ КАРТЫ 249
Математические ожидания, приведенные в этой таблице, обнару-
живают очевидную комбинаторную симметрию, и можно надеять-
ся,
что с помощью какого-либо изящного математического приема
можно
будет
получить в явном виде максимально правдоподобные
оценки
и средние квадратические ошибки. Так, если для
удобства
принять
обозначения
U
= a
i
+
Ъ
2
+ с
3
+ d
it
~]
V
= а
2
-f bi + с
4
+ d
3
, I
2
, f
(10ЛЗ>
j
то функцию правдоподобия для рассматриваемой совокупности
наблюдаемых величин можно записать в следующем виде:
e
L
~
p
p
q
Q
r
R
s
s
u
u
v
v
w
w
t
T
D;
ni
D;
n
*D;
ns
D;
n
\
(10.14)
Это соотношение необходимо максимизировать
при
условиях
JD
Ц—
gr-|—
г
—f—
s = 1 и и^ v^ w-\-1 = 4.
Поэтому начнем
с
вычисления
таких основных характеристик,
как
дЬ Р п{и Й
2
У n
3
w n^t ..„ .,_.
~др~
=
~р
D] D~
2
Ih Ж (1U.10J
и
т. д.
К
сожалению, чисто математический анализ сопряжен здесь
с очень сложными алгебраическими выкладками, хотя можно
получить ряд интересных результатов для некоторых частных
случаев. Допустим, например, что необходимо начать с проверки
существования дифференциальной жизнеспособности. Принимаем
нулевую
гипотезу и = v = w = 1 и приступаем к вычислению
дхаксимально правдоподобных оценок для этих параметров, исполь-
зуя максимально правдоподобные оценки частот рекомбинаций
в
отсутствие дифференциальной жизнеспособности. Легко видеть,
что эти оценки имеют вид
p = P/N, q = Q/N, r = R/N, s = S!N.
(10.16)
Далее находим дифференциальные характеристики
^ = U- (щР + n
2
Q + n
3
R + n
k
S)IN,
4£ = V -
(щ(?
+ п
2
Р + n
3
S + n,R)IN,
dv
[
(10.17)
^L
=
W- (n
t
R + n
2
S + n
3
P +
/i
4
<?)/JV,
4£-
= T - (n,S + n
2
R +
n
3
Q
+
n,P)IN,
250
ГЛАВА
10
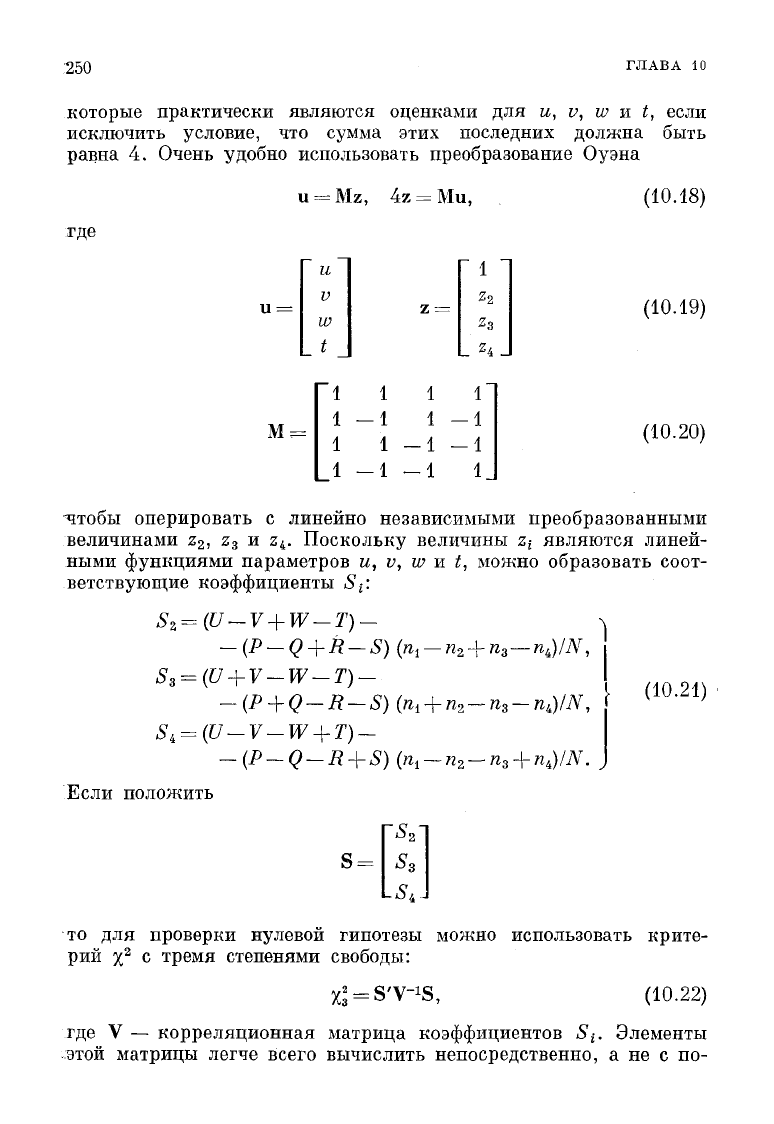
которые практически являются оценками для и, v, w и t, если
исключить условие, что сумма этих последних должна быть
равна 4. Очень удобно использовать преобразование Оуэна
=
Mz,
где
М =
и
V
w
t _
"1
1
1
1
z =
1
1
-1
1 -
1
-1 -
-1
-1
" 1
2
_Z
4
_
г
-1
-1
1
(10.18)
(10.19)
(10.20)
чтобы оперировать с линейно независимыми преобразованными
величинами z
2
, z
3
и z
4
. Поскольку величины z
t
являются линей-
ными
функциями параметров и, v, w и t, можно образовать соот-
ветствующие коэффициенты S
t
:
(10.21)
ЕСЛИ
ПОЛОЖИТЬ
S =
то для проверки нулевой гипотезы можно использовать крите-
рий
%
2
с тремя степенями свободы:
x
j =
S'V-
1
S,
(10.22)
где V — корреляционная матрица коэффициентов S
t
. Элементы
этой
матрицы
легче
всего вычислить непосредственно, а не с по-