Березов Т.Т., Коровкин Б.Ф. Биологическая химия
Подождите немного. Документ загружается.

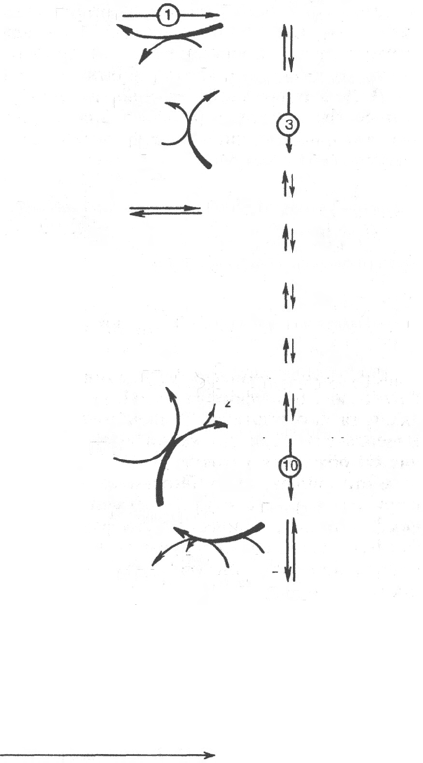
Рис. 10.7. Гликолиз и глюконеогенез.
Красными стрелками указаны «обходные»
пути глюконеогенеза при биосинтезе глю-
козы из пирувата и лактата; цифры в круж-
ках обозначают соответствующую стадию
гликолиза.
зо-6-фосфат. Последний может дефосфорилироваться (т.е. реакция идет
в обход гексокиназной реакции) под влиянием фермента глюкозо-6-фос-
фатазы:
На рис. 10.7 представлены «обходные» реакции глюконеогенеза при
биосинтезе глюкозы из пирувата и лактата.
Регуляция глюконеогенеза. Важным моментом в регуляции глюконеоге-
неза является реакция, катализируемая пируваткарбоксилазой. Роль по-
ложительного аллостерического модулятора этого фермента выполняет
ацетил-КоА. В отсутствие ацетил-КоА фермент почти полностью лишен
активности. Когда в клетке накапливается митохондриальный ацетил-КоА,
биосинтез глюкозы из пирувата усиливается. Известно, что ацетил-КоА
одновременно является отрицательным модулятором пируватдегидроге-
назного комплекса (см. далее). Следовательно, накопление ацетил-КоА
замедляет окислительное декарбоксилирование пирувата, что также спо-
собствует превращению последнего в глюкозу.
Другой важный момент в регуляции глюконеогенеза – реакция, ката-
лизируемая фруктозо-1,6-бисфосфатазой – ферментом, который ингибиру-
ется АМФ. Противоположное действие АМФ оказывает на фосфофрукто-
341
Глюкоза
Глюкозо-6-фосфат
Н
2
O
Н
3
РO
4
Н
3
РО
4
Н
2
O
Фруктозо-6-фосфат
Фруктозо-1,6-бисфосфат
Диоксиацетон-
фосфат
Глицеральдегид-3-фосфат
1,3-Бисфосфоглицериновая кислота
3-Фосфоглицериновая кислота
ГДФ
ГТФ
2-Фосфоглицериновая
кислота
CО
2
Фосфоенолпируват
Оксалоацетат
Пируват
Н
3
РО
4
АДФ
АТФ
СО
2
Лактат
Глюкозо-6-фосфат + Н
2
O
Глюкозо-6-фосфатаза
Глюкоза + Р
i
.

киназу, т. е. для этого фермента он является аллостерическим активатором.
При низкой концентрации АМФ и высоком уровне АТФ происходит
стимуляция глюконеогенеза. Напротив, когда величина отношения
АТФ/АМФ мала, в клетке наблюдается расщепление глюкозы.
В 1980 г. группой бельгийских исследователей (Г. Херс и др.) в ткани
печени был открыт фруктозо-2,6-бисфосфат, который является мощным
регулятором активности двух перечисленных ферментов:
Фруктозо-2,6-бисфосфат активирует фосфофруктокиназу и ингибирует
фруктозо-1,6-бисфосфатазу. Повышение в клетке уровня фруктозо-2,6-бис-
фосфата способствует усилению гликолиза и уменьшению скорости глю-
конеогенеза. При снижении концентрации фруктозо-2,6-бисфосфата отме-
чается обратная картина.
Установлено, что биосинтез фруктозо-2,6-бисфосфата происходит из
фруктозо-6-фосфата при участии АТФ, а распадается он на фруктозо-6-
фосфат и неорганический фосфат. Биосинтез и распад фруктозо-2,6-бис-
фосфата катализируется одним и тем же ферментом, т.е. данный фермент
бифункционален, он обладает и фосфокиназной, и фосфатазной актив-
ностью:
Показано также, что бифункциональный фермент в свою очередь ре-
гулируется путем цАМФ-зависимого фосфорилирования. Фосфорилирова-
ние приводит к увеличению фосфатазной активности и снижению фосфо-
киназной активности бифункционального фермента. Этот механизм объяс-
няет быстрое воздействие гормонов, в частности глюкагона, на уровень
фруктозо-2,6-бисфосфата в клетке (см. главу 16).
Активность бифункционального фермента регулируется также неко-
торыми метаболитами, среди которых наибольшее значение имеет гли-
церол-3-фосфат. Действие глицерол-3-фосфата на фермент по своей направ-
ленности аналогично эффекту, который наблюдается при его фосфори-
лировании с помощью цАМФ-зависимых протеинкиназ.
342
β-Фруктозо-2,6-бисфосфат
АТФ
АДФ
(фосфокиназное действие)
Фруктозо-6-фосфат
Бифункциональный
фермент
Фруктозо-2,6-бисфосфат
(фосфатазное действие)
P
i
Н
2
О
В настоящее время фруктозо-2,6-бисфосфат, помимо печени, обнаружен
и в других органах и тканях животных, а также у растений и микро-
организмов.
Показано, что глюконеогенез может регулироваться и непрямым путем,
т.е. через изменение активности фермента, непосредственно не участвую-
щего в синтезе глюкозы. Так, установлено, что фермент гликолиза пиру-
ваткиназа существует в 2 формах – L и М. Форма L (от англ. liver – печень)
преобладает в тканях, способных к глюконеогенезу. Эта форма ингиби-
руется избытком АТФ и некоторыми аминокислотами, в частности ала-
нином. М-форма (от англ. muscle – мышцы) такой регуляции не подвержена.
В условиях достаточного обеспечения клетки энергией происходит инги-
бирование L-формы пируваткиназы. Как следствие ингибирования замед-
ляется гликолиз и создаются условия, благоприятствующие глюконеоге-
незу.
Наконец, интересно отметить, что между гликолизом, интенсивно про-
текающим в мышечной ткани при ее активной деятельности, и глюко-
неогенезом, особенно характерным для печеночной ткани, существует
тесная взаимосвязь. При максимальной активности мышц в результате
усиления гликолиза образуется избыток молочной кислоты, диффунди-
рующей в кровь, в печени значительная ее часть превращается в глюкозу
(глюконеогенез). Такая глюкоза затем может быть использована как
энергетический субстрат, необходимый для деятельности мышечной ткани.
Взаимосвязь между процессами гликолиза в мышечной ткани и глю-
конеогенезом в печени может быть представлена в виде схемы:
АЭРОБНЫЙ МЕТАБОЛИЗМ ПИРУВАТА
Клетки, недостаточно снабжаемые кислородом, могут частично или пол-
ностью существовать за счет энергии гликолиза. Однако большинство
животных и растительных клеток в норме находится в аэробных условиях
и свое органическое «топливо» окисляет полностью до СО
2
и Н
2
О. В этих
условиях пируват, образовавшийся при расщеплении глюкозы, не восста-
343
Гликоген
Глюкозо-6-фосфат
Глюкоза
Лактат
Мышцы
Экзогенная
глюкоза из
пищеварительного
тракта
Глюкоза
Лактат
Кровяное русло
Гликоген
Глюкоза
Глюкозо-6-фосфат
Лактат
Печень
навливается до лактата, а постепенно окисляется до СО
2
и Н
2
О в аэробной
стадии катаболизма, при этом первоначально происходит окислительное
декарбоксилирование пирувата с образованием ацетил-КоА.
Окислительное декарбоксилирование
пировиноградной кислоты
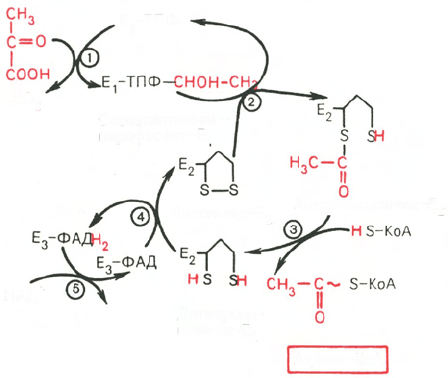
Окисление пирувата до ацетил-КоА происходит при участии ряда фермен-
тов и коферментов, объединенных структурно в мультиферментную систе-
му, получившую название «пируватдегидрогеназный комплекс».
На I стадии этого процесса пируват (рис. 10.8) теряет свою карбоксиль-
ную группу в результате взаимодействия с тиаминпирофосфатом (ТПФ)
в составе активного центра фермента пируватдегидрогеназы (E
1
). На II
стадии оксиэтильная группа комплекса E
1
–ТПФ–СНОН–СН
3
окисляется
с образованием ацетильной группы, которая одновременно переносится на
амид липоевой кислоты (кофермент), связанной с ферментом дигидроли-
поилацетилтрансферазой (Е
2
). Этот фермент катализирует III стадию – пе-
ренос ацетильной группы на коэнзим КоА (HS-KoA) с образованием
конечного продукта ацетил-КоА, который является высокоэнергетическим
(макроэргическим) соединением.
На IV стадии регенерируется окисленная форма липоамида из вос-
становленного комплекса дигидролипоамид–Е
2
. При участии фермента
дигидролипоилдегидрогеназы (Е
3
) осуществляется перенос атомов водо-
рода от восстановленных сульфгидрильных групп дигидролипоамида на
ФАД, который выполняет роль простетической группы данного фермента
и прочно с ним связан. На V стадии восстановленный ФАДН
2
дигидро-
липоилдегидрогеназы передает водород на кофермент НАД с образованием
НАДН + Н
+
.
Процесс окислительного декарбоксилирования пирувата происходит
в матриксе митохондрий. В нем принимают участие (в составе сложного
мультиферментного комплекса) 3 фермента (пируватдегидрогеназа, ди-
гидролипоилацетилтрансфераза, дигидролипоилдегидрогеназа) и 5 кофер-
ментов (ТПФ, амид липоевой кислоты, коэнзим А, ФАД и НАД), из
Рис. 10.8. Механизм действия
пируватдегидрогеназного комп-
лекса.
Е
1
- пируватдегидрогеназа; Е
2
- ди-
гидролипоилацетилтрансфсраза; Е
3
-
дигидролипоилдегидрогеназа; циф-
ры в кружках обозначают стадии
процесса.
344
Оксиэтилтиамин-
пирофосфат-Е
1
Липоамид-Е
2
Ацетиллипоамид-Е
2
Дигидроли-
поамид-Е
2
Ацетил-КоА
Е
1
-ТПФ
СO
2
НАД
+
НАДН+H
+
которых три относительно прочно связаны с ферментами (ТПФ-E
1
, ли-
поамид-Е
2
и ФАД-Е
3
), а два – легко диссоциируют (HS-KoA и НАД).
Все эти ферменты, имеющие субъединичное строение, и коферменты
организованы в единый комплекс. Поэтому промежуточные продукты
способны быстро взаимодействовать друг с другом. Показано, что со-
ставляющие комплекс полипептидные цепи субъединиц дигидролипоил-
ацетилтрансферазы составляют как бы ядро комплекса, вокруг которого
расположены пируватдегидрогеназа и дигидролипоилдегидрогеназа. При-
нято считать, что нативный ферментный комплекс образуется путем са-
мосборки.
Суммарную реакцию, катализируемую пируватдегидрогеназным комп-
лексом, можно представить следующим образом:
Пируват + НАД
+
+ HS-KoA –> Ацетил-КоА + НАДН + Н
+
+ СO
2
.
Реакция сопровождается значительным уменьшением стандартной сво-
бодной энергии и практически необратима.
Образовавшийся в процессе окислительного декарбоксилирования аце-
тил-КоА подвергается дальнейшему окислению с образованием СО
2
и Н
2
О. Полное окисление ацетил-КоА происходит в цикле трикарбоновых
кислот (цикл Кребса). Этот процесс, так же как окислительное декарбо-
ксилирование пирувата, происходит в митохондриях клеток.
ЦИКЛ ТРИКАРБОНОВЫХ КИСЛОТ (ЦИКЛ КРЕБСА)
Цикл трикарбоновых кислот впервые был открыт английским биохимиком
Г. Кребсом *.
Он первым постулировал значение данного цикла для полного сгорания
пирувата, главным источником которого является гликолитическое пре-
вращение углеводов. В дальнейшем было показано, что цикл трикарбо-
новых кислот является тем центром, в котором сходятся практически все
метаболические пути. Таким образом, цикл Кребса – общий конечный путь
окисления ацетильных групп (в виде ацетил-КоА), в которые превращается
в процессе катаболизма большая часть органических молекул, играющих
роль «клеточного топлива»: углеводов, жирных кислот и аминокислот.
Образовавшийся в результате окислительного декарбоксилирования
пирувата в митохондриях ацетил-КоА вступает в цикл Кребса. Данный
цикл происходит в матриксе митохондрий и состоит из восьми последова-
тельных реакций (рис. 10.9). Начинается цикл с присоединения ацетил-КоА
к оксалоацетату и образования лимонной кислоты (цитрата). Затем лимон-
ная кислота (шестиуглеродное соединение) путем ряда дегидрирований
(отнятие водорода) и двух декарбоксилирований (отщепление СО
2
) теряет
два углеродных атома и снова в цикле Кребса превращается в оксалоацетат
(четырехуглеродное соединение), т.е. в результате полного оборота цикла
одна молекула ацетил-КоА сгорает до СО
2
и Н
2
О, а молекула окса-
лоацетата регенерируется. Рассмотрим все восемь последовательных реак-
ций (этапов) цикла Кребса.
* За это выдающееся открытие Г. Кребс получил Нобелевскую премию в 1953 г.
(совместно с Ф. Липманом). Цикл трикарбоновых кислот часто называют его именем - цикл
Кребса (цикл лимонной кислоты Кребса).
345
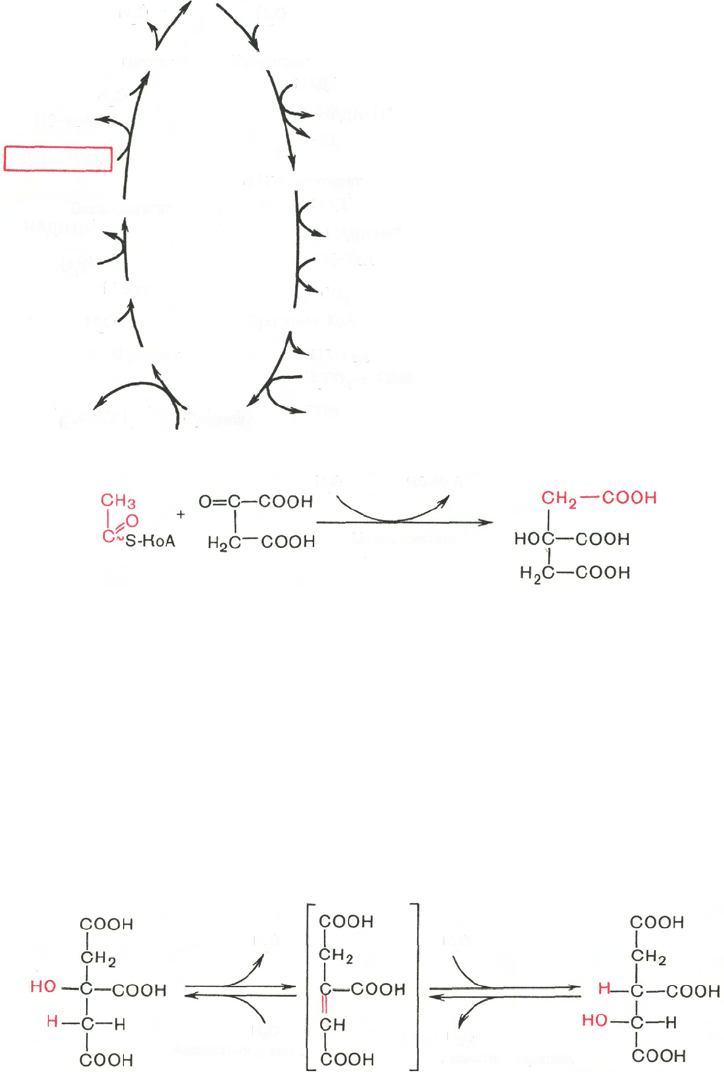
Рис. 10.9. Цикл трикарбоновых
кислот (цикл Кребса).
Первая реакция ката-
лизируется ферментом цит-
рат-синтазой, при этом аце-
тильная группа ацетил-КоА
конденсируется с оксалоаце-
татом, в результате чего об-
разуется лимонная кислота:
По-видимому, в данной реакции в качестве промежуточного продукта
образуется связанный с ферментом цитрил-КоА. Затем последний само-
произвольно и необратимо гидролизуется с образованием цитрата и
HS-KoA.
В результате второй реакции образовавшаяся лимонная кислота
подвергается дегидратированию с образованием цис-аконитовой кислоты,
которая, присоединяя молекулу воды, переходит в изолимонную кислоту
(изоцитрат). Катализирует эти обратимые реакции гидратации–дегидра-
тации фермент аконитатгидратаза (аконитаза). В результате происходит
взаимоперемещение Н и ОН в молекуле цитрата:
346
цис-Аконитат
Н
2
О
Н
2
О
Цитрат
Изоцитрат
НАД
+
НАДН + Н
+
СО
2
α-Кетоглутарат
НАД
+
НАДН + Н
+
HS-KoA
СО
2
Сукцинил-КоА
HS-KoA
Н
3
РО
4
+ ГДФ
ГТФ
Сукцинат
Е-ФАД
Е-ФАДН
2
Фумарат
Н
2
О
Малат
НАД
+
НАДН+ Н
+
Оксалоацетат
Ацетил-КоА
HS-KoA
Н
2
О
Н
2
О
HS-КoA
Цитрат-синтаза
Ацетил-КоА
Оксалоацетат
Цитрат
Цитрат
Н
2
О
Н
2
О
Аконитат-гидратаза
цис-Анонитат
Н
2
О
Н
2
О
Аконитат-гидратаза
Изоцитрат
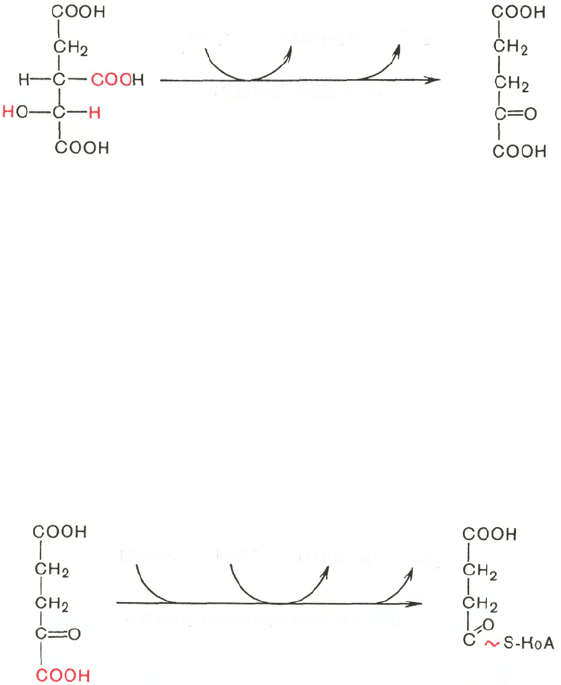
Третья реакция, по-видимому, лимитирует скорость цикла Кребса.
Изолимонная кислота дегидрируется в присутствии НАД-зависимой изо-
цитратдегидрогеназы *.
В ходе изоцитратдегидрогеназной реакции изолимонная кислота одно-
временно декарбоксилируется. НАД-зависимая изоцитратдегидрогеназа
является аллостерическим ферментом, которому в качестве специфического
активатора необходим АДФ. Кроме того, фермент для проявления своей
активности нуждается в ионах Mg
2+
или Мn
2+
.
Во время четвертой реакции происходит окислительное декарбокси-
лирование α-кетоглутаровой кислоты с образованием высокоэнергетическо-
го соединения сукцинил-КоА. Механизм этой реакции сходен с таковым
реакции окислительного декарбоксилирования пирувата до ацетил-КоА,
α-кетоглутаратдегидрогеназный комплекс напоминает по своей структуре
пируватдегидрогеназный комплекс. Как в одном, так и в другом случае
в реакции принимают участие 5 коферментов: ТПФ, амид липоевой кис-
лоты, HS-KoA, ФАД и НАД
+
.
Пятая реакция катализируется ферментом сукцинил-КоА-синтета-
зой. В ходе этой реакции сукцинил-КоА при участии ГТФ и неорганического
фосфата превращается в янтарную кислоту (сукцинат). Одновременно
происходит образование высокоэргической фосфатной связи ГТФ ** за счет
высокоэргической тиоэфирной связи сукцинил-КоА:
* В митохондриях существует 2 типа изоцитратдегидрогеназ: НАД- и НАДФ-зависимый;
первый тип встречается только в митохондриях, второй - как в митохондриях, так и
в цитозоле.
** Образовавшийся ГТФ отдает затем свою концевую фосфатную группу на АДФ,
вследствие чего образуется АТФ. Образование высокоэргического нуклеозидтрифосфата в ходе
сукцинил-КоА-синтетазной реакции - еще один пример фосфорилирования на уровне субстрата
(субстратное фосфорилирование).
347
Изоцитратдегидрогеназа
Изоцитрат
α-Кетоглутарат
α-Кетоглутаратдегидрогеназный комплекс
α-Кетоглутарат
Сукцинил-КоА
НАД
+
НАДН + Н
+
СО
2
HS-KoA
НАД
+
НАДH + Н
+
СO
2
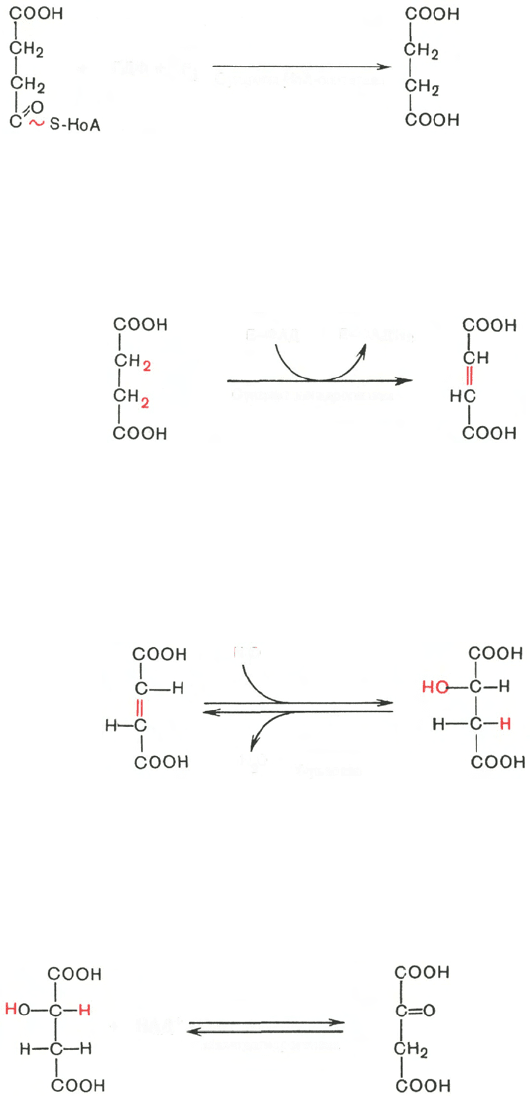
В результате шестой реакции сукцинат дегидрируется в фумаровую
кислоту. Окисление сукцината катализируется сукцинатдегидрогеназой,
в молекуле которой с белком прочно (ковалентно) связан кофермент ФАД.
В свою очередь сукцинатдегидрогеназа прочно связана с внутренней ми-
тохондриальной мембраной:
Седьмая реакция осуществляется под влиянием фермента фума-
ратгидратазы (фумаразы). Образовавшаяся при этом фумаровая кислота
гидратируется, продуктом реакции является яблочная кислота (малат).
Следует отметить, что фумаратгидратаза обладает стереоспецифичностью
(см. главу 4) – в ходе реакции образуется L-яблочная кислота:
Наконец, в ходе восьмой реакции цикла трикарбоновых кислот под
влиянием митохондриальной НАД-зависимой малатдегидрогеназы проис-
ходит окисление L-малата в оксалоацетат:
348
Сукцинил-КоА-синтетаза
Сукцинил-КоА
Сукцинат
Сукцинатдегидрогеназа
Сукцинат
Фумарат
Фумарат
Фумараза
L-малат
L- малат
Малатдегидрогеназа
Оксалоацетат
+ ГДФ + P
i
+ ГТФ + HS-KoA
Е-ФАД
Е-ФАДН
2
H
2
O
H
2
O
+ НАД
+
+ НАДН + Н
+
Как видно, за один оборот цикла, состоящего из восьми фермента-
тивных реакций, происходит полное окисление («сгорание») одной мо-
лекулы ацетил-КоА. Для непрерывной работы цикла необходимо постоян-
ное поступление в систему ацетил-КоА, а коферменты (НАД
+
и ФАД),
перешедшие в восстановленное состояние, должны снова и снова окис-
ляться. Это окисление осуществляется в системе переносчиков электронов
в дыхательной цепи (в цепи дыхательных ферментов), локализованной
в мембране митохондрий. Образовавшийся ФАДН
2
прочно связан с СДГ,
поэтому он передает атомы водорода через KoQ. Освобождающаяся
в результате окисления ацетил-КоА энергия в значительной мере сосре-
доточивается в макроэргических фосфатных связях АТФ. Из 4 пар атомов
водорода 3 пары переносят НАДН на систему транспорта электронов; при
этом в расчете на каждую пару в системе биологического окисления
образуется 3 молекулы АТФ (в процессе сопряженного окислительного
фосфорилирования), а всего, следовательно, 9 молекул АТФ (см. главу 9).
Одна пара атомов от сукцинатдегидрогеназы-ФАДН
2
попадает в систему
транспорта электронов через KoQ, в результате образуется только 2 моле-
кулы АТФ. В ходе цикла Кребса синтезируется также одна молекула ГТФ
(субстратное фосфорилирование), что равносильно одной молекуле АТФ.
Итак, при окислении одной молекулы ацетил-КоА в цикле Кребса и системе
окислительного фосфорилирования может образоваться 12 молекул АТФ.
Если подсчитать полный энергетический эффект гликолитического рас-
щепления глюкозы и последующего окисления двух образовавшихся моле-
кул пирувата до СО
2
и Н
2
О, то он окажется значительно большим.
Как отмечалось, одна молекула НАДН (3 молекулы АТФ) * образуется
при окислительном декарбоксилировании пирувата в ацетил-КоА. При
расщеплении одной молекулы глюкозы образуется 2 молекулы пирувата,
а при окислении их до 2 молекул ацетил-КоА и последующих 2 оборотов
цикла трикарбоновых кислот синтезируется 30 молекул АТФ (следователь-
но, окисление молекулы пирувата до СО
2
и Н
2
О дает 15 молекул АТФ).
К этому количеству надо добавить 2 молекулы АТФ, образующиеся при
аэробном гликолизе, и 6 молекул АТФ, синтезирующихся за счет окисления
2 молекул внемитохондриального НАДН, которые образуются при окис-
лении 2 молекул глицеральдегид-3-фосфата в дегидрогеназной реакции
гликолиза. Следовательно, при расщеплении в тканях одной молекулы
глюкозы по уравнению С
6
Н
12
О
6
+ 6О
2
—> 6СО
2
+ 6Н
2
О синтезируется 38
молекул АТФ. Несомненно, что в энергетическом отношении полное
расщепление глюкозы является более эффективным процессом, чем ана-
эробный гликолиз.
Необходимо отметить, что образовавшиеся в процессе превращения
глицеральдегид-3-фосфата 2 молекулы НАДН в дальнейшем при окислении
могут давать не 6 молекул АТФ, а только 4. Дело в том, что сами молекулы
внемитохондриального НАДН не способны проникать через мембрану
внутрь митохондрий. Однако отдаваемые ими электроны могут включаться
в митохондриальную цепь биологического окисления с помощью так
называемого глицеролфосфатного челночного механизма (рис. 10.10). Ци-
топлазматический НАДН сначала реагирует с цитоплазматическим ди-
гидроксиацетонфосфатом, образуя глицерол-3-фосфат. Реакция катализи-
* Напомним, что при прохождении по цепи дыхательных ферментов восстановительные
эквиваленты НАДН генерируют три высокоэнергетические фосфатные связи посредством
образования АТФ из АДФ в процессе окислительного фосфорилирования (см. главу 9).
349

Рис. 10.10. Глицеролфосфатный челноч-
ный механизм. Объяснение в тексте.
руется НАД-зависимой цитоплазматической глицерол-3-фосфат-дегидроге-
назой:
Дигидроксиацетонфосфат + НАДН + Н
+
<=> Глицерол-3-фосфат + НАД
+
.
Образовавшийся глицерол-3-фосфат легко проникает через митохонд-
риальную мембрану. Внутри митохондрии другая (митохондриальная)
глицерол-3-фосфат-дегидрогеназа (флавиновый фермент) снова окисляет
глицерол-3-фосфат до диоксиацетонфосфата:
Глицерол-3-фосфат + ФАД <=> Диоксиацетонфосфат + ФАДН
2
.
Восстановленный флавопротеин (фермент-ФАДН
2
) вводит на уровне
KoQ приобретенные им электроны в цепь биологического окисления
и сопряженного с ним окислительного фосфорилирования, а диоксиаце-
тонфосфат выходит из митохондрий в цитоплазму и может вновь взаимо-
действовать с цитоплазматическим НАДН + Н
+
. Таким образом, пара
электронов (из одной молекулы цитоплазматического НАДН + Н
+
), вво-
димая в дыхательную цепь с помощью глицеролфосфатного челночного
механизма, дает не 3, а 2 АТФ.
350
НАДН + Н
+
НАД
+
Цитоплазматическая
глицерол-3-фосфат-дегидрогеназа
Цитозоль
Диокси-
ацетонфосфат
Глицерол-3-
фосфат
Митохондриальная
глицерол-3-фосфат-дегидро-
Митохондрия
геназа
ФАДН
2
ФАД
KoQ
2е
-
АТФ
АТФ
О
2