Березов Т.Т., Коровкин Б.Ф. Биологическая химия
Подождите немного. Документ загружается.

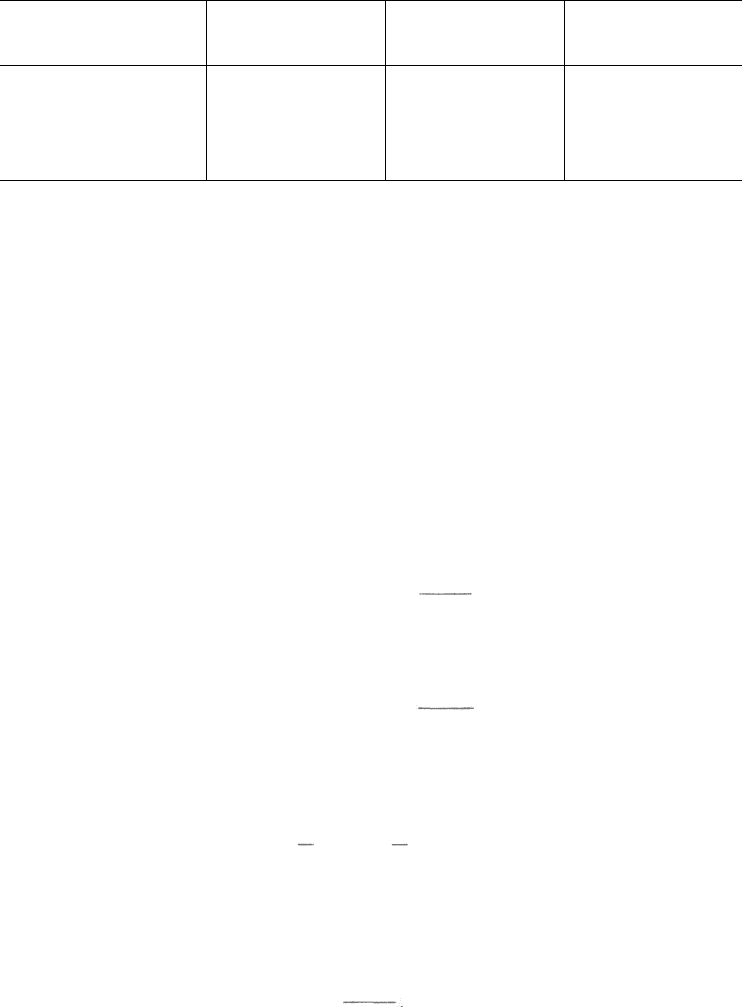
Таблица 3.2. Состав рибосомных РНК эукариот
Размер субчастицы,
ед. Сведберга (S)
60 (более 50 белков)
40 (более 30 белков)
Молекулярная
масса субчастицы
2,7•10
6
1,3•10
6
Размер РНК,
ед. Сведберга (S)
5
5,8
28
18
Молекулярная
масса РНК
35000
45000
1,5•10
6
750000
СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ
Для понимания ряда особенностей структуры ДНК важное значение имели
закономерности состава и количественного содержания азотистых осно-
ваний, установленные впервые Э. Чаргаффом. Оказалось, что азотистые
основания ДНК обычно варьируют у разных видов организмов, однако
почти не претерпевают изменений у одного и того же вида в процессе
развития или в зависимости от изменений окружающей среды либо харак-
тера питания. Показано также, что ДНК, выделенная из разных тканей
одного и того же вида, имеет одинаковый состав азотистых оснований.
Полученные количественные соотношения были названы правилами Чар-
гаффа. При анализе состава очищенной ДНК, выделенной из разных
источников, были сделаны следующие выводы:
1) молярная доля пуринов * равна молярной доле пиримидинов:
2) количество аденина и цитозина равно количеству гуанина и тимина:
3) количество аденина равно количеству тимина, а количество гуанина
равно количеству цитозина: А = Т и Г = Ц; соответственно
4) существенным для характеристики вида (таксономическое значение)
оказался так называемый коэффициент специфичности, отражающий от-
ношение
* Здесь и далее пуриновые и пиримидиновые основания обозначены начальными буквами
соответствующего названия.
101
А + Г = Ц + Т или
А + Ц = Г + Т или
А + Г
Ц + Т
= 1;
А + Ц
Г + Т
= 1;
А
Т
= 1;
Г
= 1;
Г + Ц
А + Т
Ц
Это отношение часто выражают в молярных процентах (Г + Ц), или
процентах ГЦ-пар. Для животных и большинства растений этот коэф-
фициент ниже 1 (от 0,54 до 0,94), у микроорганизмов он колеблется
в значительных пределах (от 0,45 до 2,57).
Данные, полученные А. Н. Белозерским и его учениками, свидетельст-
вуют о существовании в природе АТ-типа ДНК (у хордовых и беспозво-
ночных животных, высших растений, ряда бактерий, дрожжевидных орга-
низмов) и ГЦ-типа ДНК (у недрожжевидных грибов, актиномицетов, ряда
бактерий и вирусов).
Известно, что структурными единицами нуклеиновых кислот являются
мономерные молекулы – мононуклеотиды. Следовательно, нуклеиновые
кислоты представляют собой полинуклеотиды. Это продукты полимериза-
ции мононуклеотидов, число и последовательность расположения которых
в цепях ДНК и РНК определяются в строгом соответствии с программой,
заложенной в молекуле матрицы (см. главу 14). Мононуклеотиды легко
образуются при гидролизе ДНК и РНК в присутствии нуклеаз, состоят из
трех специфических компонентов: азотистого основания, углевода и фос-
форной кислоты. В этой «триаде» мононуклеотида углевод занимает
среднее положение. Соединения азотистого (любого) основания и углевода
(рибозы или дезоксирибозы), получившие название нуклеозидов, легко
образуются из мононуклеотида при гидролитическом отщеплении фос-
форной кислоты в присутствии щелочи или при участии специфических
ферментов – нуклеотидаз.
Нуклеозиды содержат пуриновое или пиримидиновое основание,
соединенное с углеводом N-гликозидной связью. В составе нуклеиновых
кислот обнаруживаются только β-нуклеозиды. Примером могут служить
два мононуклеотида: аденозин-5'-монофосфорная кислота (АМФ) и ци-
тидин-5'-монофосфорная кислота (ЦМФ):
R у 2' углерода представлен Н- или ОН-группой в зависимости от типа
нуклеиновой кислоты – ДНК или РНК. В образовании N-гликозидной связи
в пуриновых нуклеотидах принимают участие N-9 пурина и С-1' пен-
тозы, а в пиримидиновых нуклеотидах – N-1 пиримидина и С-1' пентозы.
Чтобы отличить углеродные атомы рибозы или дезоксирибозы от уг-
леродных атомов, входящих в состав пуриновых и пиримидиновых
102
АМФ
ЦМФ
N-гликозидные
связи

оснований, первые принято обозначать символом «штрих»: например,
атомы у 3-го и 5-го углерода обозначают С-3' и С-5' или, чаще,
3' и 5'.
Следует отметить, что среди продуктов ферментативного гидролиза
ДНК и РНК обнаруживаются, помимо нуклеозид-5'-монофосфатов, также
нуклеозид-3'-монофосфаты. Положение фосфата определяется местом раз-
рыва фосфодиэфирной связи между соседними нуклеотидами, что ука-
зывает на характер связи нуклеотидов через остаток фосфорной кислоты,
соединяющий 3' и 5' углеродные атомы пентозы.
В табл. 3.3 приведены состав и названия (включая тривиальные), а также
сокращенные обозначения нуклеозидов и нуклеотидов (для РНК они назы-
ваются рибонуклеотидами, а для ДНК – дезоксирибонуклеотидами).
Таблица 3.3. Состав нуклеозидов и мононуклеотидов
Азотистые основания
Пуриновые
Пиримиди-
новые
Аденин
Гуанин
Урацил
Цитозин
Тимин
Нуклеозиды
(основание
+ углевод)
Аденозин
Гуанозин
Уридин
Цитидин
Тимидин
Мононуклеотиды
(нуклеозиды + Н
3
РО
4
)
Аденозинмонофосфат
(адениловая кислота)
Гуанозинмонофосфат
(гуаниловая кислота)
Уридинмонофосфат
(уридиловая кислота)
Цитидинмонофосфат
(цитидиловая кислота)
Тимидинмонофосфат
(тимидиловая кислота)
Сокращенное
обозначение
АМФ
ГМФ
УМФ
ЦМФ
ТМФ
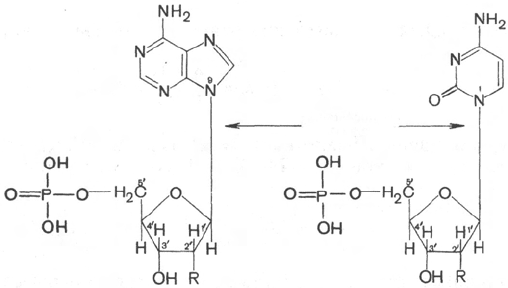
Мононуклеотиды и их производные, а также динуклеотиды присутст-
вуют в клетках в свободном виде и играют важную роль в обмене веществ.
В частности, нуклеотидную структуру имеют многие коферменты, включая
коферменты оксидоредуктаз. Мононуклеотиды, присоединяя еще один ос-
таток фосфата, образуют фосфоангидридную связь (наподобие связи,
имеющейся в пирофосфате) и превращаются в нуклеозиддифосфаты
(соответственно они обозначаются сокращенно АДФ, ГДФ, УДФ, ЦДФ
и ТДФ). Последние, присоединяя еще один остаток фосфата, образуют
нуклеозидтрифосфаты (соответственно обозначаются АТФ, ГТФ,
УТФ,
ЦТФ и
ТТФ).
Следует особо указать, что только свободные нуклеозидтрифосфаты
в клетках являются предшественниками ферментативного синтеза ДНК
и РНК (см. главу 13). Однако в клетках имеются свободные, также
природные нуклеозидтрифосфаты, не принимающие участия в синтезе
белка, но выполняющие жизненно важные функции. В частности, одной из
важнейших функций нуклеозидтрифосфатов и особенно АТФ является их
участие в биоэнергетике всех живых организмов (см. главу 13). Приводим
схему образования молекул аденозинди- и аденозинтрифосфатов (неко-
торые атомы водорода, как и углерода, в пуриновом ядре и в кольце
рибозы опущены):
103
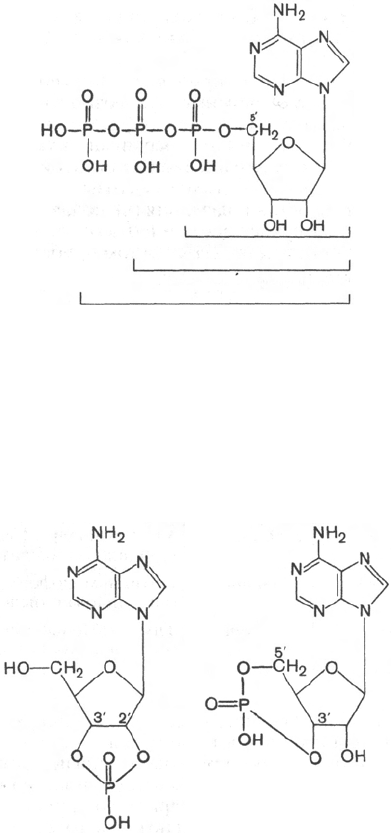
Необходимо указать на существование в организме еще двух типов
фосфорных эфиров нуклеотидов, когда фосфат связывает 2 атома кисло-
рода пентозного остатка в одном и том же нуклеотиде и когда фосфатный
мостик объединяет два разных мононуклеотида. Примером первого типа
являются циклические нуклеотиды 2',3'- и 3',5'-, т.е. два возможных класса
соединений, в которых кислородные атомы у С-2' и С-3' или у С-3' и С-5'
участвуют в образовании циклической структуры:
Первое из этих соединений, 2',3'-АМФ, образуется в качестве промежу-
точного продукта распада рибонуклеиновых кислот, в то время как цикли-
ческий 3',5'-АМФ (цАМФ) является естественно встречающимся рибо-
нуклеотидом (он образуется из АТФ в процессе реакции, катализируемой
ферментом аденилатциклазой). цАМФ наделен рядом уникальных функций
и высокой биологической активностью в регуляции процессов обмена,
выполняя роль медиатора внеклеточных сигналов в клетках животных.
Аналогичной функцией наделены цГМФ, производные УДФ, ЦТФ и
нуклеотиды в составе кофакторов и коферментов (см. главу 4).
Примером, когда одна фосфатная группа связывает два разных рибо-
нуклеотида, является структура гуанозилтимидинфосфата:
104
Аденозин-5'-монофосфат (АМФ)
Аденозин-5'-дифосфат (АДФ)
Аденозин-5'-трифосфат (АТФ)
Циклический 2',3'-АМФ
Циклический 3',5'-АМФ

Это соединение хотя и не обнаружено в природе, но содержит тип связи,
характерный для природных нуклеиновых кислот. Видно, что С-3' рибозы
одного нуклеотида соединяется посредством фосфодиэфирной связи с
С-5' рибозы второго нуклеотида.
В медицинской практике, в частности в онкологии, нашли широкое
применение синтетические аналоги как азотистых оснований, так нуклео-
зидов и нуклеотидов. Эти аналоги, имеющие небольшие модификации
в структуре основания или углевода, встраиваясь в соответствующие
клеточные компоненты, оказывают заметный цитотоксический эффект.
К наиболее распространенным лекарственным препаратам – аналогам пу-
риновых и пиримидиновых оснований (и соответствующим нуклеотидам)
относятся 5-фторурацил, 6-тио- и 6-меркаптопурин, 8-азагуанин, 6-азаури-
дин и 6-азацитидин, а также 5-йодпроизводное дезоксиуридина.
Помимо сокращенных названий и обозначений нуклеозидов и нуклео-
тидов (см. табл. 3.3), приняты буквенные обозначения нуклеозидов (и
нуклеотидов): в частности, для аденозина (и АМФ) это А, для гуанозина (и
ГМФ) – Г, для цитидина (и ЦМФ) – Ц, для уридина (и УМФ) – У, для
тимидина (и ТМФ) – Т. Пользуясь этими символами, приведенный выше
дирибонуклеозидмонофосфат можно обозначить как Г–Т. Заметим, что как
по структуре, так и по свойствам Г–Т и Т–Г будут сильно отличаться друг
от друга (как и в случае дипептидов).
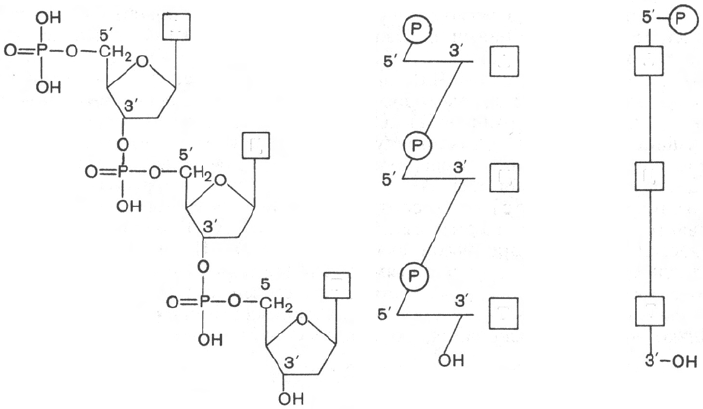
Первичная структура нуклеиновых кислот
Под первичной структурой нуклеиновых кислот понимают порядок, после-
довательность расположения мононуклеотидов в полинуклеотидной цепи
ДНК и РНК. Такая цепь стабилизируется 3',5'-фосфодиэфирными связями.
Поскольку молекулярная масса нуклеиновых кислот колеблется в широких
пределах
(от
2•10
4
до
10
10
–10
11
),
установить
первичную структуру всех
известных РНК и особенно ДНК весьма сложно. Тем не менее во всех
нуклеиновых кислотах (точнее, в одноцепочечной нуклеиновой кислоте)
имеется один и тот же тип связи – 3',5'-фосфодиэфирная связь между
соседними нуклеотидами. Эту общую основу структуры можно представить
следующим образом:
Установлено, что в образовании межнуклеотидной связи участвуют
гидроксильные группы в 3'- и 5'-положениях остатков углевода.
105
Основание
Основание
Основание
Пентоза
Пентоза
Пентоза
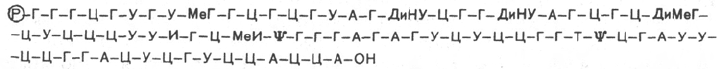
К настоящему времени удалось определить первичную структуру почти
всех тРНК, ряда молекул 5S рРНК, 16S рРНК E.coli, вирусных РНК,
в состав которых входят сотни и тысячи нуклеотидных остатков. Ниже
приводится примерная схема последовательности нуклеотидов в молекуле
РНК. Все клеточные РНК в основном состоят из одноцепочечной по-
линуклеотидной цепи:
5'-Г–У–Г–Ц–А–А–...–У–Ц–Г–Ц–Ц–А–3'
Полинуклеотидная цепь молекулы РНК имеет на одном конце почти
всегда свободный монофосфорный эфир, который принято обозначать как
5'-конец; на противоположном конце цепи такой фосфат отсутствует,
а содержится нуклеотид со свободными 2'- и 3'-гидроксильными группами.
Если подвергнуть щелочному гидролизу молекулу РНК, то в качестве
концевого нуклеотида будут обнаружены ЦМФ со свободным фосфатом
у 5'-конца и свободный аденозин в виде свободного нуклеозида у 3'-конца
полинуклеотидной цепи.
В выяснении первичной структуры РНК решающую роль сыграли
методы ступенчатого гидролиза, осуществленного в основном экзонуклеа-
зами и заключающегося в последовательном отщеплении по одному
мононуклеотиду с одного конца молекулы нуклеиновой кислоты. Ниже
представлена первичная структура первой РНК, имеющей 77 нуклеотидов,
для которой была расшифрована нуклеотидная последовательность в
1965 г. Р. Холли и сотр., а именно аланиновой тРНК:
В этой структуре Р – остаток фосфата, ψ – псевдоУМФ, МеГ – метилгуа-
нин, ДиНУ – дигидроурацил, ДиМеГ – диметилгуанин, МеИ – метилинозин.
Следует особо указать на две существенные особенности первичной
структуры всех тРНК. Первая из них заключается в том, что 5'-концом
всегда является гуаниловая (редко цитидиловая) кислота, несущая свобод-
ный остаток фосфата у С-5'. Вторая особенность – наличие на проти-
воположном конце молекулы остатков трех мононуклеотидов с одинаковой
последовательностью – ЦЦА, причем остаток адениловой кислоты содер-
жит свободную 3'-ОН-группу *.
Между этими структурами в строго определенной последовательности
располагаются все остальные нуклеотидные остатки, среди которых на
долю минорных нуклеотидов приходится до 10%. Полинуклеотидная цепь
разных типов тРНК содержит около 75 нуклеотидов.
Матричные (информационные) РНК относятся к наиболее гетероген-
ному классу нуклеиновых кислот, отличающихся по массе (см. табл. 3.1),
структуре, размерам, стабильности и функциям. Основной функцией мРНК
является перенос информации от ДНК (точнее, от гена) на белоксинте-
зирующую систему клетки. мРНК выполняет роль матрицы и, следо-
вательно, определяет первичную структуру синтезируемого белка (подроб-
нее см. главу 14). мРНК наделены рядом особенностей первичной струк-
туры; в частности, на 5'-конце все они содержат определенную после-
* Другие особенности структуры тРНК рассматриваются в главе 14.
106
довательность рибонуклеотидов, получившую название шапочки (кэп).
Первым нуклеотидом является 7-метилгуанозинтрифосфат, который при-
соединяется к 5'-гидроксилу соседнего мононуклеотида, представленного
2'-О-метилпуриновым нуклеотидом. На другом 3'-конце большинства (но
не всех) мРНК содержится полиадениловая последовательность (поли-А),
насчитывающая от 150 до 200 нуклеотидов.
Роль «кэпирования» и «полиаденилирования» мРНК в белковом синтезе
окончательно не выяснена. Предполагают, что кэп необходим для спе-
цифического узнавания в процессе трансляции, в то время как поли-А
отводится роль фактора стабилизации всей молекулы мРНК.
В последние годы расшифрована первичная структура не только низко-
молекулярных 5S рРНК разных бактерий и 5,8S рРНК клеток животных, но
и высокомолекулярных 16S и 18S рРНК, насчитывающих до 1200–1500
нуклеотидных звеньев. Более того, уже выяснены нуклеотидные после-
довательности 23S рРНК E.coli и 25S рРНК дрожжевой клетки, а также
первичные структуры высокомолекулярных (28S) рРНК клеток эукариот,
насчитывающих около 4700 нуклеотидов.
В настоящее время проводятся исследования первичных структур
различных молекул ДНК. Около 15 лет назад была полностью расшифро-
вана нуклеотидная последовательность митохондриальной ДНК человека
(16569 пар нуклеотидов). Известны полные нуклеотидные последователь-
ности ДНК ряда вирусов и плазмид. Совсем недавно завершено опреде-
ление нуклеотидных последовательностей геномов двух прокариотических
организмов (Haemophilus influenzae и Mycoplasma genitalum) и появились
сообщения о расшифровке генома первого эукариотического организма –
дрожжей. Близки к завершению аналогичные исследования генома E.coli
и генома нематоды Caenorhabditis elegans. Исследователи активно ра-
ботают над полной расшифровкой генома человека.
Результаты секвенирования (определение нуклеотидной последователь-
ности) разных молекул ДНК накапливаются в виде компьютерных банков
данных, которые уже доступны для пользователей международных
компьютерных сетей (например, «Internet»). Ниже представлены три ва-
рианта схемы нуклеотидной последовательности ДНК:
107
А
Ц
Т
А
Ц
Т
А
Ц
Т
В последнее время о первичной структуре ДНК (точнее, отдельных ее
фрагментов) судят по ряду косвенных данных, например, по степени
сплоченности нуклеотидных звеньев в молекуле ДНК (определение сводит-
ся в конечном счете к выяснению числа и структуры отдельных фракций
нуклеотидов, так называемых изоплитов), также по кинетике реассоциации
ДНК (метод позволяет выяснить наличие в молекуле повторяющихся
последовательностей нуклеотидов). О первичной структуре ДНК судят,
кроме того, по распределению минорных оснований (имеются данные
о существовании подобной закономерности) и обнаружению в ДНК и оп-
ределению последовательности палиндромов («обратно бегущие» после-
довательности, или перевертыши), которые обнаруживаются главным об-
разом в местах рестрикции (см. главу 13). Большие надежды в определении
первичной структуры ДНК исследователи возлагают на физические, хими-
ческие (синтез генов), генетические и другие методы, а также на методы
выделения некоторых генов (или их фрагментов) из природных источников
и синтеза генов на мРНК при участии фермента обратной транскриптазы.
Для установления первичной структуры ДНК недавно предложен экспресс-
метод, включающий применение двух ДНК-полимераз (из E.coli и из
бактериофага Т4). Однако во всех этих случаях определяется структура
небольшого участка ДНК, поэтому полная расшифровка первичной струк-
туры ДНК генома человека ждет своего решения.
Вторичная структура нуклеиновых кислот
В соответствии с моделью Дж. Уотсона и Ф. Крика, предложенной в 1953 г.
на основании ряда аналитических данных, а также рентгеноструктурного
анализа молекула ДНК состоит из двух цепей, образуя правовращающую
спираль, в которую обе полинуклеотидные цепи закручены вокруг одной
и той же оси. Удерживаются цепи благодаря водородным связям, обра-
зующимся между их азотистыми основаниями (рис. 3.1). Обе цепи поли-
нуклеотидов в биспиральной молекуле ДНК имеют строго определенное
пространственное расположение, при котором азотистые основания нахо-
дятся внутри, а фосфорильные и углеводные компоненты – снаружи *.
Детальный анализ всевозможных вариантов образования водородных
связей между основаниями показал, что в биспиральной молекуле ДНК
основания уложены парами: пурин из одной цепи и пиримидин из другой
в соответствии с правилами Чаргаффа. Поскольку ориентация оснований на
плоскости не является, очевидно, произвольной, и основания в поли-
нуклеотидах представлены в лактамной форме, наиболее вероятными были
признаны пары аденин–тимин и гуанин–цитозин. Этот способ спаривания
получил в дальнейшем экспериментальное подтверждение. Избиратель-
ность взаимодействия пар А–Т и Г–Ц принято выражать термином
«комплементарность», а соответствующие азотистые основания на-
зывают комплементарными. Стабильность А–Т оснований обеспе-
чивается двумя водородными связями, а пар Г–Ц – тремя, что в свою
очередь определяется особенностями расположения функциональных групп
азотистых оснований. Длина водородных связей между основаниями со-
ставляет около 0,3 нм. Таким образом, комплементарными оказываются не
только отдельные основания, но и дезоксирибонуклеотидные цепи ДНК
* За это открытие Дж. Уотсон и Ф. Крик вместе с М. Уилкинсом получили Нобелевскую
премию в 1962 г.
108
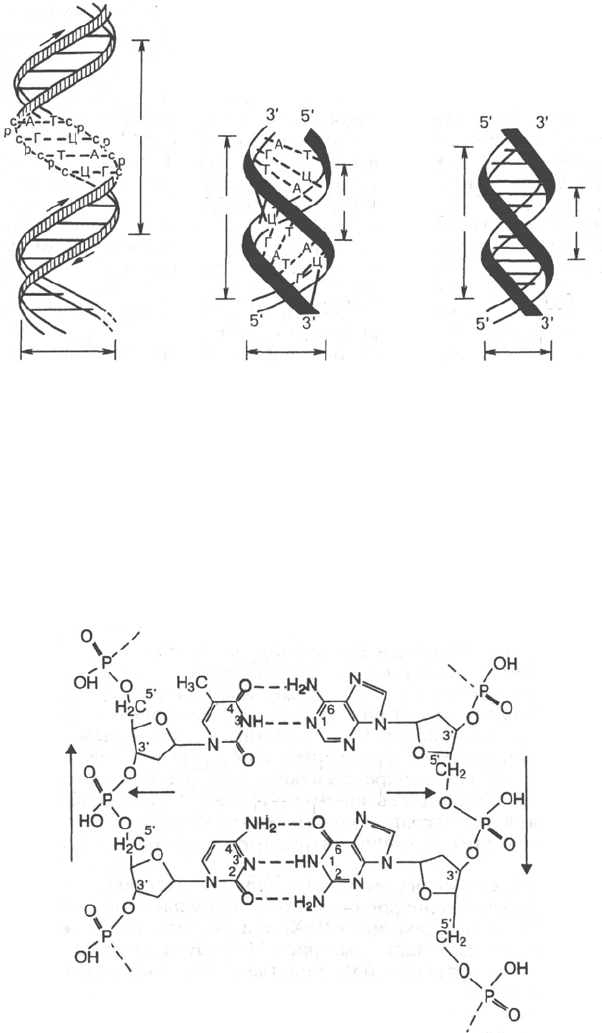
Рис. 3.1. Схематическое изображение двойной спирали ДНК.
а - по Уотсону и Крику: с - остаток дезоксирибозы, р - остаток фосфорной кислоты; б - А-фор-
ма ДНК; в - В-форма ДНК.
в целом, способствующие образованию весьма компактной структуры
и стабилизации всей молекулы *.
Обе цепи в молекуле ДНК имеют противоположную полярность. Это
означает, что межнуклеотидная связь в одной цепи имеет направление
5'–>3', а в другой – 3'–>5'. Подобная направленность цепей имеет важное
биологическое значение при репликации и транскрипции молекулы ДНК.
* Данные, полученные в последние годы, свидетельствуют, что в стабилизации биспи-
ральной структуры основную роль играют гидрофобные взаимодействия между компле-
ментарными основаниями, стыкующимися в центре двойной спирали. Водородные связи,
вероятнее всего, обеспечивают специфичность спаривания оснований.
109
Тимин
Аденин
Антипараллельные цепи
Цитозин
Гуанин
3,4 нм
1,8 нм
а
2,8 нм
1,4 нм
1,8 нм
б
3,4 нм
1,3 нм
1,7 нм
в
70°
90°
0,28
нм
0,30
нм
0,29
нм
0,30
нм
0,29
нм
На модели ДНК (см. рис. 3.1) видно, что расстояние между витками
(шаг спирали) равно 3,4 нм. На этом участке укладываются 10 нуклеотид-
ных остатков, размер одного нуклеотида составляет 0,34 нм; диаметр
биспиральной молекулы равен 1,8 нм.
Необходимо указать, что конфигурация двойной спирали ДНК сильно
меняется в зависимости от количественного содержания воды и ионной
силы окружающей среды. Методами рентгеноструктурного анализа дока-
зано существование по крайней мере 6 форм ДНК, названных А-, В-, С-, D-,
Е- и Z-формами. Конфигурация двух из них в простейшей форме представ-
лена на рис. 3.1, б и в. Можно увидеть, что у А-формы наблюдается
некоторое смещение пар оснований от оси молекулы к периферии, что
отражается на размерах (2,8 нм – длина одного витка, в котором вместо 10
содержится 11 мононуклеотидов; меняется расстояние между нуклеотидами
и др.). Если А- и В-формы представляют собой правозакрученную двойную
спираль, то Z-форма (зигзагообразная) ДНК имеет левозакрученную кон-
фигурацию, в которой фосфодиэфирный остов располагается зигзагообраз-
но вдоль оси молекулы. Параллельно фосфодиэфирному остову в структуре
А- и В-форм ДНК имеются большая и малая бороздки (желобки) –
сайты, где присоединяются белки, выполняющие, очевидно, регуляторные
функции при экспрессии генов. В настоящее время есть основание считать,
что между А- и В-формами ДНК осуществляются взаимные переходы при
изменении концентрации соли и степени гидратации. В-форма ДНК больше
всего подходит к модели Уотсона и Крика. В этих переходах, которые
могут быть вызваны растворителями или белками, очевидно, заключен
определенный биологический смысл. Предполагают, что в А-форме ДНК
выполняет роль матрицы в процессе транскрипции (синтез РНК на моле-
куле ДНК), а в В-форме – роль матрицы в процессе репликации (синтез
ДНК на молекуле ДНК).
В структуре ДНК, как и в структуре РНК, открыты нуклеотидные
последовательности, получившие название «палиндромы», или перевер-
нутые повторы. Они встречаются как внутри одной цепи, так и в двойной
спирали. Например, как слово ротатор, которое одинаково читается как
справа налево, так и обратно. Подобные обратные повторы могут служить
основой для образования структуры шпилек или других вариаций с изме-
ненным внутрицепочечным и межцепочечным спариванием и формирова-
нием на отдельных участках тройной спирали. Возможно, эти палинд-
ромные структуры имеют определенный биологический смысл в регуляции
экспрессии отдельных генов, выполняя роль сайтов для ДНК-связывающих
белков. Предстоит, однако, приложить немало усилий для установления как
точной структуры этих вариаций, так и для определения их функциональной
роли.
Менее охарактеризована вторичная структура матричных и рибосомных
РНК. Относительно вторичной структуры тРНК наиболее вероятной пред-
ставляется модель, предложенная Р. Холли, плоское изображение которой
напоминает клеверный лист (см. рис. 14.3). В настоящее время, когда
известна первичная структура большинства тРНК, последовательность всех
или почти всех природных тРНК как будто бы укладывается в эту схему
«клеверного листа» (см. главу 14). При сравнении этих структур выявляется
ряд закономерностей, несомненно, имеющих определенный биологический
смысл. Во всех тРНК есть участки, взаимодействующие с рибосомами,
места для связывания с аминокислотами и ферментами, а также специфи-
ческая последовательность трех нуклеотидов (триплет), называемая анти-
кодоном, которая оказывается комплементарной тринуклеотидной после-
110