Алексеев В.В. Физическое и математическое моделирование экосистем
Подождите немного. Документ загружается.


триггерные режимы в системе, где исход конкуренции зависит от
начальных условий.
Рассмотрим более подробно триггерные режимы в модели.
Для этого изобразим поведение траекторий на фазовой плоско-
сти (М, М'). Рассмотрим случай задания параметров Ai, Аг
в
секторе II плоскости биогенов. Из уравнений сохранения масс
(3.13)
следует,
что реально возможные значения биомасс ограни-
чены на фазовой плоскости областью под прямыми
о,М + а;М' = Д,
(3.43)
а
2
М + а
2
М' = Л
2
.
(3.44)
Условия (3.43),
(3.44)
определяют предельное увеличение биомасс
М, М', при которых концентрации биогенов в среде становятся
равными
нулю и дальнейшее развитие фитоценоза становится не-
возможным.
Учитывая дополнительно
(3.16)
и (3.18), получим, что область
возможных значений биомасс имеет вид четырехугольника, при-
мыкающего к осям координат. Вершины его на фазовой плоскости
расположены в точках (0, 0), (0,
A
2
/a'
z
),
(Ai/a
u
0), (|Я
2
|/б,
Hi/б),
как показано на рис. 3.8. Все возможные стационарные
точки находятся внутри четырехугольника, их положение опреде-
ляется из (3.30), (3.34), (3.38). Динамика, описываемая уравне-
ниями
(3.22), справедлива в треугольной области выше линии
M'=#i/6,
уравнения (3.23)—в области справа от линии М =
=
|#
2
|/6.
В прямоугольной области вблизи начала координат по-
ведение фитоценоза отражают уравнения (3.21).
Линии
перемены знаков производных
dJA/dt
и
dlA'/dt
для
уравнений
(3.22)
представляют собой две прямые, параллельные
,(3.44):
S
2
= 0
2
;
S
2
=--%.
(3.45)
Линии
смены знака производных для уравнений (3.23)—это
прямые,
параллельные (3.43):
s, = e,; s
2
= e;.
(3.46)
В уравнениях
(3.21)
производные меняют знаки на прямых
S, = e,; S
2
= 0
2
.
(3.47)
Скорость роста популяции положительна, если фазовая точка
расположена ниже прямой минимума биогенов для данного вида,
и
отрицательна в обратном случае. Несколько фазовых траекто-
рий,
полученных на ЭВМ, изображены на рис. 3.8. Интересно, что
траектории из начальной точки стремятся кратчайшим путем при-
близиться к ближайшей прямой минимума биогенов в среде и да-
лее движутся вдоль нее к стационарному значению. Специфиче-
ский
характер траекторий отражает явление аутостабилизации,
т. е. выхода на минимум концентрации лимитирующего компо-
-.101

нента
питания в среде задолго до установления равновесия в си-
стеме, независимо от начальных биомасс видов и начальной кон-
центрации
биогена в среде.
Нетрудно видеть (см. рис. 3.8), что каждая стационарная точка
на
фазовой плоскости может оказаться как устойчивой, так и не-
устойчивой в зависимости от взаимного расположения линий пере-
мены
знака
(3.45)
— (3.47). На рис. 3.8 а представлен случай, когда
все три стационара устойчивы — тройной триггер; рис. 3.8 6 — это
двойной
тригер; на рис. 3.8в все равновесные точки неустойчивы,
траектории выходят на предельный цикл.
Таким
образом, выбирая нужное сочетание физиологических
параметров на одном из вариантов рис. 3.8 и значения А\, Аг
У
соответствующие требуемому
исходу
конкуренции, можно автома-
тически получить доминирование нужного вида водоросли. Для
триггерных режимов начальные концентрации биомасс определя-
ются по изображению на фазовой плоскости.
Полное
исследование структуры конкурентных исходов
даже
в
рассмотренном простом примере фитоценоза из
двух
видов
с двумя лимитирующими компонентами питания показывает бога-
тые возможности получения разнообразных режимов культивиро-
вания
микроорганизмов и
дает
рекомендации для сознательного
управления ростом и конкурентными отношениями в биоценозах
одного трофического уровня.
3.2.3.
Конкуренция
в
проточной
системе
При
исследовании модели конкуренции в фитоценозе для зам-
кнутой системы рассматривались только установившиеся режимы,
процессы перехода к равновесному состоянию во внимание не
принимались.
Для проточных систем, в отличие от замкнутых,
сбалансированных по массе, характерны сложные и длительные
переходные процессы, занимающие в ряде случаев десятки дней
даже
в специальных лабораторных условиях [259]. Эта особен-
ность открытых систем приводит к существенным трудностям при
создании
искусственных экосистем и при прогнозировании изме-
нений
в естественных водоемах, связанных с антропогенным воз-
действием.
В настоящем параграфе мы установим соотношение
между
динамикой
проточной и замкнутой по веществу систем на при-
мере фитоценоза и проанализируем качественный характер пере-
ходных процессов в открытой экосистеме.
Запишем
уравнения динамики фитоценоза
(3.12)
для проточ-
ной
системы. Уравнения (3.12), описывающие динамику биомасс
М и М', остаются неизменными; для концентраций биогенных эле-
ментов Si и S
2
в среде имеем [25]
^S
I
,
S
2
)-a;M'O
2
(S
If
S
2
),
(3.48)
S
I
, S
2
)-a'
2
M'O
2
(S
v
S
2
).
102

Систему (3.12),
(3.48)
можно частично проинтегрировать, вы-
разив зависимости
двух
переменных, например Si и S2, от вре-
мени
в явном виде [4, 25].
Исключая
<Pi(Si, S
2
) и tf>
2
(Si, S
2
) из (3.12),
(3.48)
и делая за-
мену переменных
z,
= s, — s° +
О;М
+
<*;м\
систему
(3.48)
можно свести к простейшим уравнениям
dZ,
-^- + DZi = 0,
/=1,2,
(3.50)
откуда Zt{t)
=Z,(0)
exp (—Dt).
Возвращаясь к исходным переменным, получим для Si(i),
$2(0 выражения
S, (/) = S° - а,М — а;М' + Z, (0) exp (—Dt),
S
2
(t)
=
SO
- а
2
М - а^М' + Z
2
(0)exp
{-Dt),
(3<51)
где
Z,
(0) = S, (0) - SJ - a,M (0) + ajM' (0),
Z
2
(0) = S
2
(0) - S° + «
2
M (0) + a^M' (0).
(3
'
52)
Следует
отметить, что начальные концентрации биогенов
в
среде Si(0), S2(0)
могут
значительно отличаться от концентра-
ций
S°, S°
2
в поступающей с протоком питательной среде. Такие
ситуации возникают при смене питательных сред в процессе куль-
тивирования.
С
учетом
(3.51)
можно упростить выражения, стоящие под
знаком
sign,
а именно:
sign
(-J -1^)«
sign
(Я, - 6М' + [a,Z
2
(0) - a
2
Z, (0)] exp (-
(3.53)
sign
(ii _ 4) =
sign
{H
2
+ 6M - [a;z
2
(0) - a'
2
Z, (0)] exp
(-Dt)},
\ a
2
a
[
/
где Hi, Иг, б имеют смысл, аналогичный (3.15), где S° =A\,
S°
2
=Аг.
Легко видеть, что, за исключением экспоненциально
затухаю-
щего члена, выражения
(3.53)
совпадают с выражениями, стоя-
щими
под знаком
sign
в замкнутой системе (3.14). Таким образом,
проточная система имеет те же стационарные состояния, что и
замкнутая система с запасами биогенных элементов S
0
и S
0
.
Очевидно, однако, что за время перехода к равновесному состоя-
нию
знаки сигнатур
(3.53)
могут
неоднократно меняться, т. е.
10,3

будут
происходить смены лимитирующих факторов для каждой
популяции,
не характерные для замкнутой системы.
Для того чтобы понять закономерности смены факторов лими-
тирования
в переходном процессе, обратимся снова к формулам
(3.51). Поскольку общие запасы биогенов Ai и А
2
в экосистеме
можно выразить как
'"^"СТ"'!!''
(з
-
54)
уравнения (3.51) приобретают
вид
O)exn(-Dt),
(3.55)
следовательно, установление равновесия в проточной системе сво-
дится к изменению экосистемы таким образом, что содержание
каждого из биогенов становится равным его поступлению.
Изобразим
переходной процесс на плоскости биогенов (Л
4
, Ai).
Исключая
из
(3.55)
время, получим, что в течение перехода к рав-
новесию проточная система на плоскости (Ai, А
2
) движется вдоль.
отрезка прямой
A
=fw
(A
-
s?)+s
"
(3
-
56
>
в
направлении от начальной точки с координатами (Si(0); S2(0))
к
точке с координатами (S°, S°).
Как
видно из рис. 3.6, плоскость (А
и
А
2
) для системы кон-
курирующих водорослей представляет собой мозаичную совокуп-
ность областей существования различных устойчивых стационар-
ных состояний. Во время переходного процесса система, двигаясь
вдоль отрезка прямой (3.56), попадает последовательно в зоны
различного конкурентного доминирования, причем из рис. 3.6 мо-
жно определить, какие именно состояния
будет
проходить система.
В пределах каждой зоны доминирования знаки сигнатур не ме-
няются,
и система стремится перейти в соответствующее устойчи-
вое стационарное состояние. Поскольку скорость установления ло-
кального равновесия в проточной системе пропорциональна
ехр [—D + yS/(K+S)]t, а запасы биогенных элементов изменяются
как
ехр (—Dt), приближенно можно считать, что при прохожде-
нии
каждой зоны устойчивости, биомассы популяций и концентра-
ции
биогенов в среде выходят на соответствующие стационарные
значения.
Сами же численные значения стационаров медленно
«ползут»
вслед за изменением параметров Ai, A
2
до тех пор, пока
точка (A
x
(t); A
2
{t)) не войдет в
другую
зону устойчивости.
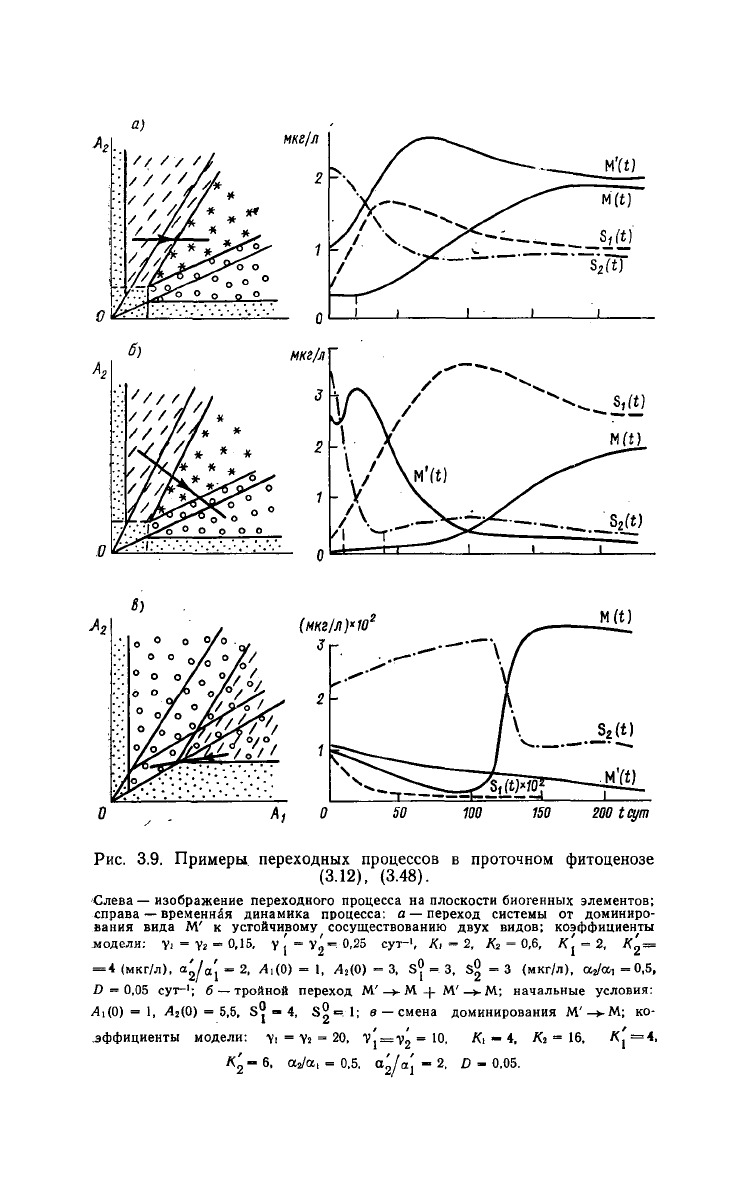
На
рис. 3.9 представлены переходные процессы в проточных
экосистемах, параметры которых соответствуют различным вари-
антам заполнения плоскости (Ai, A
2
) областями конкурентного
104
H'(t)
S
d
(t)
s'
t
W
Ж
1
/I
/*
•
о
1
*
* *
poo
M(t)
WO
200 ta/m
Рис. З.9.
Примеры
переходных
процессов
в
проточном
фитоценозе
(3.12), (3.48).
Слева — изображение переходного процесса на плоскости биогенных элементов;
справа — временная динамика процесса: а — переход системы от доминиро-
вания
вида М' к устойчивому сосуществованию
двух
видов; коэффициенты
модели: Yi - V» - 0,15. у J = У^=0,25
сут-
1
,
Ki = 2, К2 = 0,6, к| = 2, к'
2
=
=4
(мкг/л), а^/а; - 2,
,4,(0)
= 1, /ЫО) - 3, S° = 3, S?, = 3 (мкг/л), aj/a, =0,5,
D
= 0,05
сут-';
б —тройной переход М'—>-М + М'—»-М; начальные условия:
J4I(0)
— 1, А
2
(0) = 5,5, S^ - 4, S9=. 1; в — смена доминирования М'
-*•
М; ко-
эффициенты
модели: Yi = Ys = 20, vj =
i>2
~
10
. /Ci — •*. Х2 = 16, Л| = 4.
«2 - 6, aj/a, = 0,5,
а'
2
[а\
- 2, D - 0,05.

доминирования. Начальные условия выбирались таким образом,
чтобы за время установления равновесия система проходила воз-
можно большее число областей доминирования. Для каждого из
вариантов рис. 3.9 дается параллельное изображение переходного
процесса на плоскости биогенов (A
it
Л2), а также результаты
интегрирования системы (3.12),
(3.48)
на ЭВМ.
Смену доминирующих видов, изображенную на рис. 3.9 а, б,
можно схематически представить соответственно следующим об-
разом:
На
рис. 3.9в изображен процесс смены доминирующего вида без:
промежуточной стадии сосуществования популяций:
Из
рис. 3.9 хорошо видно, что переходные процессы в проточ-
ных экосистемах
могут
быть весьма длительными, например, для
водорослей со средними скоростями роста (диатомовые, см.
рис.
3.9 6) установление равновесия занимает в ряде случаев до
100 дней. По-видимому, в естественных водоемах равновесные со-
стояния
в экосистемах не успевают установиться за летний период,
что подтверждает гипотезу Хатчинсона [149] о нестационарности
процессов в природных водоемах.
3.2.4.
Сравнение теории
с
результатами
долговременных
экспериментов
по
конкуренции
Экспериментальная проверка моделей фитопланктонных сооб-
ществ была выполнена в
1970-х
годах
в Мичиганском университете
Д. Тилманом [314, 315]. Тилманом были проведены долговремен-
ные эксперименты по определению исходов конкуренции
между
пресноводными водорослями
Asterionella
formosa
и
Cyclotella
meneghiniana
при лимитировании двумя минеральными компонен-
тами питания — фосфатами (РО|~) и силикатами (SiO
2
~~). Конку-
ренцию изучали на полунепрерывных
культурах
вдоль градиента
отношения кремний/фосфор, которое варьировали от 1000 до 1.
Опыты проводились в стерильных условиях при постоянстве тем-
пературы и освещенности. Каждый опыт ставили для нескольких
скоростей разбавления
культур
(/=0,05; 0,15; 0,35; 0,5 сут"
1
).
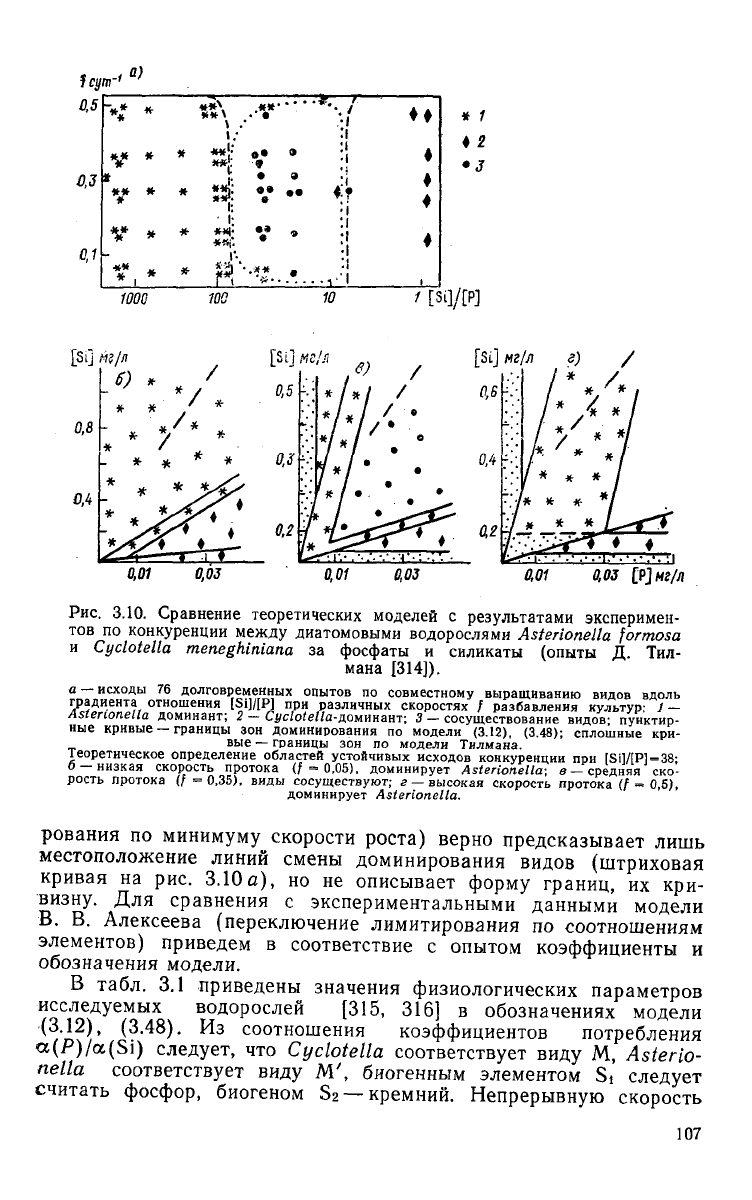
Исходы 76 экспериментов по конкуренции
между
A.
formosa
и
С.
meneghiniana
представлены на рис. 3.10, а. Граница
между
об-
ластью доминирования A.
formosa
и областью сосуществования
двух
видов имеет заметную кривизну как при высоких, так и при
очень низких скоростях разбавления культур. Как показал Тил-
ман [314, 315], модель конкуренции (3.8) (переключение лимити-
106
woo
10
1 [SQ/[P]
0,07
0,03
0,01
0,03
0,01
0,03
[р]нг/л
Рис.
3.10.
Сравнение
теоретических
моделей
с
результатами
эксперимен-
тов
по
конкуренции
между
диатомовыми
водорослями
Asterionella formosa
и
Cyclotella meneghiniana за
фосфаты
и
силикаты
(опыты
Д. Тил-
мана
[314]).
а — исходы
76
долговременных опытов
по
совместному выращиванию видов вдоль
градиента отношения [Si]/[P]
при
различных скоростях
/
разбавления культур-
/ -
Asterionella
доминант;
2
— С1/с/о/<Ша-доминант;
3
— сосуществование видов- пунктир-
ные
кривые — границы
зон
доминирования
по
модели (3.12), (3.48); сплошные
кри-
вые — границы
зон по
модели Тилмана.
Теоретическое определение областей устойчивых исходов конкуренции
при
[Si]/[P]=38;
О
—низкая
скорость протока (/-0,05), доминирует
Asterionella;
в
—средняя
ско-
рость протока (/=0,35), виды сосуществуют;
г
— высокая скорость протока
(/ =
0,5),
доминирует
Asterionella.
рования
по
минимуму скорости роста) верно предсказывает лишь
местоположение линий смены доминирования видов (штриховая
кривая
на рис. 3.10
а),
но не
описывает форму границ,
их кри-
визну.
Для
сравнения
с
экспериментальными данными модели
В.
В.
Алексеева
(переключение лимитирования
по
соотношениям
элементов) приведем
в
соответствие
с
опытом коэффициенты
и
обозначения модели.
В табл.
3.1
приведены значения физиологических параметров
исследуемых
водорослей
[315, 316] в
обозначениях модели
/о
/'
(?-
48
)-
Из
соотношения коэффициентов потребления
<x(P)/a(Si)
следует,
что
Cyclotella
соответствует
виду
М,
Asterio-
nella
соответствует
виду
М',
биогенным элементом
Si
следует
считать фосфор, биогеном
S
2
—кремний. Непрерывную скорость
107

Таблица
3.1
Физиологические
параметры
роста
фосфатом
и
Вид
Cyclotella
meneghiniana
Cyclotella
meneghiniana
Asterionella
formosa
Asterionella
formosa
Биоген
PO
4
SiO
2
PO
4
SiO
2
микроводорослей
при лимитировании
силикатом
[315]
К
мг/л
7,75-Ю-з
40,32-10-з
0,62-Ю-з
ПО-Ю-з
7
сут-
1
0,8
1,3
0,9
1,1
1/а
кл.(мг/л)-'
7-10
9
8,9-Ю
7
8,9-Ю
8
1,5-Ю
8
протока
D в
системе определяем через скорость периодического
разбавления
по
формуле
Концентрацию фосфора
в
поступающей питательной
среде
варьи-
ровали
в
ходе
экспериментов
от
0,0031
до
0,465
мг/л,
концентра-
цию кремния
— от 2,8 до
0,252
мг/л.
По
данным
из
табл.
3.1 для
нескольких значений скорости
разбавления
/
рассчитывались
6, и 6' (t=l,
2)—минимальные
концентрации
биогенов, необходимые
для
выживания каждого
из
видов водорослей
при
фосфатном
или
силикатном лимитирова-
нии
[см.
формулы (3.20), (3.26)]. После этого были изображены
распределения
зон
доминирования видов
на
плоскости биогенов
([Si],
[Р])
аналогично соответствующим вариантам
рис. 3.6 при
низкой
(f =
0,05 сут""
1
), средней
(f =
0,35
сут"
1
)
и
высокой
(/ =
=
0,5
сут"
1
)
скоростях разбавления
культур
(рис.
3.1 Об,в,г).
Рассмотрим распределение
зон
конкуренции
на рис.
ЗЛО
в.
При
заданной (средней) скорости разбавления имеется область
сосуществования видов
в
интервале отношений
78,6 >
[Si]
/
[Р]
>
>
5,5.
Границы области сосуществования видов
в
данном
случае
совпадают
в
обоих вариантах моделей конкуренции
(6 <
< [Si]/ [P]
<; 80) и
соответствуют
экспериментальным данным.
Распределение
зон
конкурентного доминирования
при
высоких
скоростях разбавления
(f = 0,5
сут~')
показано
на рис.
3.10г.
Сосуществование
двух
видов становится невозможным,
при со-
отношении биогенов [Si]/[P]
=38
доминирует
вид A.
formosa,
т.
е.
граница доминирования вида искривляется
в
сторону мень-
ших значений этого отношения.
На
рис. 3.10 б
показаны области устойчивых исходов конку-
ренции
при
очень низкой скорости разбавления
культур
(f =
==0,05
сут"
1
).
В
этом режиме,
как
отмечает Тилман,
Asterionella
formosa,
лимитированная силикатом, может запасать
в 80 раз
больше фосфора,
чем
необходимо
при
данной скорости роста.
Химический состав водоросли меняется, значение отношения
[Si]/[P]
в
клетках
Asterionella
снижается
от
о^/^'^^б
до еди-
108

ницы.
Как видно из рис. 3.10 б, область сосуществования видов
отсутствует,
для отношения биогенов [Si]/[P] = 38 доминирует
A.
formosa.
Таким образом, граница доминирования этой водо-
росли при низких скоростях разбавления смещена в сторону мень-
ших отношений
[Si]/
[P],
что соответствует экспериментальным
данным.
Граница
между
доминированием
Cyclotella
и сосуществова-
нием
двух
видов в экспериментах Тилмана подробно не опреде-
лялась, теоретическое расположение границы, согласно модели
(3.12),
(3.48)
показано на рис. 3.10 а точечными линиями.
Из
проведенного сравнения
следует,
что модель В. В. Алексе-
ева (3.12),
(3.48)
достаточно хорошо согласуется с эксперимен-
тальными данными по конкуренции водорослей, правильно описы-
вает как местоположение, так и форму границ областей устойчи-
вых исходов конкуренции.

Глава
4
СИСТЕМЫ
С
ВЗАИМОДЕЙСТВИЯМИ
ТИПА
«ХИЩНИК—ЖЕРТВА»
4.1.
Элементарная
система
«хищник—жертва»
Первые модели трофического взаимодействия были построены
в
работах
В.
Вольтерра
[45].
Непосредственной предпосылкой
создания
модели явилась попытка теоретического объяснения
об-
наруженного зоологом Д'Анкона факта роста численности
хищ-
ных
рыб,
наблюдавшегося после временного прекращения рыбного
промысла
в
годы
первой мировой войны [163].
В
табл.
4.1
пред-
ставлены данные
о
процентном содержании хищных видов
рыб
в
уловах
нескольких морских портов
за
период
1905—1923
гг. Из
таблицы
следует,
что с 1915 по 1920 гг.,
когда вследствие войны
интенсивность
рыбной ловли снизилась, доля рыб-хищников
воз-
росла.
Таблица
4.1
Статистические
данные об уловах хищных видов рыб (% общего улова)
по
Адриатическому морю за
1905—1923
гг. [45]
Порт
Триест
Фиуме
Венеция
Год
1905
21,8
1910
5,7
1911
8,8
1912
9,5
1913
15,7
1914
14,6
11,9
1915
7,6
21,4
1916
16,2
22,1
Триест
Фиуме
Венеция
1917
15,4
21,2
1918
36,4
1919
19,9
27,3
30,9
1920
15,8
16
25,3
1921
13,3
15,9
25,9
1922
10,7
14,8
26,8
1923
10,2
10,7
26,6
Рассмотрим модель трофического взаимодействия
по
типу
«хищник—жертва», построенную
В.
Вольтерра. Пусть имеется
си-
стема, состоящая
из
двух
видов,
из
которых один поедает
другой.
Пусть рост популяции жертвы
при
отсутствии хищника ничто
не
ограничивает.
Тогда
скорость увеличения численности жертв
бу-
дет пропорциональна общей численности жертв. Естественно
ПО