Voet D., Voet Ju.G. Biochemistry
Подождите немного. Документ загружается.


No prokaryotic or eukaryotic cytoplasmic tRNA is
known to participate in a nonwobble pairing combination.
There is, however, no known instance of such a tRNA with
an A in its third anticodon position, which suggests that the
consequent A U pair is not permitted.The structural basis
of wobble pairing is poorly understood, although it is clear
that it is influenced by base modifications.
A consideration of the various wobble pairings indi-
cates that at least 31 tRNAs are required to translate all 61
coding triplets of the genetic code (there are 32 tRNAs in
the minimal set because translational initiation requires a
separate tRNA; Section 32-3Ca). Most cells have 32
tRNAs, some of which have identical anticodons. In fact,
mammalian cells have 150 tRNAs. Nevertheless, all
isoaccepting tRNAs in a cell are recognized by a single
aminoacyl–tRNA synthetase.
c. Some Mitochondrial tRNAs Have More Permissive
Wobble Pairings than Other tRNAs
The codon recognition properties of mitochondrial
tRNAs must reflect the fact that mitochondrial genetic codes
are variants of the “standard” genetic code (Table 32-3). For
instance, the human mitochondrial genome, which consists
of only 16,569 bp, encodes 22 tRNAs (together with 2 ribo-
somal RNAs and 13 proteins). Fourteen of these tRNAs each
read one of the synonymous pairs of codons indicated in
Tables 32-2 and 32-3 (MNX, where X is either C or U or else
A or G) according to normal G U wobble rules:The tRNAs
have either a G or a modified U in their third anticodon po-
sition that, respectively, permits them to pair with codons
having X C or U or else X A or G. The remaining 8
tRNAs,which,contrary to wobble rules,each recognize one of
the groups of four synonymous codons (MNY, where Y A,
C, G, or U), all have anticodons with a U in their third posi-
tion. Either this U can somehow pair with any of the four
bases or these tRNAs read only the first two codon positions
and ignore the third.Thus, not surprisingly, many mitochon-
drial tRNAs have unusual structures in which, for example,
the GTCRA sequence (Fig. 32-9) is missing,or, in the most
bizarre case, a tRNA
Ser
lacks the entire D arm.
d. Frequently Used Codons Are Complementary to
the Most Abundant tRNA Species
The analysis of the base sequences of several highly ex-
pressed structural genes of S. cerevisiae has revealed a re-
markable bias in their codon usage. Only 25 of the 61 cod-
ing triplets are commonly used. The preferred codons are
those that are most nearly complementary, in the
Watson–Crick sense, to the anticodons in the most abundant
species in each set of isoaccepting tRNAs. Furthermore,
codons that bind anticodons with two consecutive G C
pairs or three A U pairs are avoided so that the preferred
codon–anticodon complexes all have approximately the
same binding free energies. A similar phenomenon occurs
in E. coli, although several of its 22 preferred codons differ
from those in yeast. The degree with which the preferred
codons occur in a given gene is strongly correlated, in both
organisms, with the gene’s level of expression (the meas-
ured rates of aminoacyl–tRNA selection in E. coli span a
25-fold range). This, it has been proposed, permits the
mRNAs of proteins that are required in high abundance to
be rapidly and smoothly translated.
e. Selenocysteine and Pyrrolysine Are Carried by
Specific tRNAs
Although it is widely stated, even in this text, that pro-
teins are synthesized from the 20 “standard” amino acids,
that is, those specified by the “standard” genetic code, some
organisms, as Theresa Stadtman discovered, use a twenty-
first amino acid, selenocysteine (Sec; alternatively SeCys),
in synthesizing a few of their proteins:
Selenium, a biologically essential trace element, is a com-
ponent of several enzymes in both prokaryotes and eu-
karyotes. These include thioredoxin reductase (Section 28-
3Ae) and the thyroid hormone deiodinases (which
participate in thyroid hormone synthesis; Section 19-1D) in
mammals and three forms of formate dehydrogenases in
E. coli, all of which contain Sec residues. The Sec residues
are ribosomally incorporated into these proteins by a unique
tRNA, tRNA
Sec
, bearing a UCA anticodon that is specified
by a particular (in the mRNA) UGA codon (normally the
opal Stop codon). The Sec–tRNA
Sec
is synthesized by the
aminoacylation of tRNA
Sec
with L-serine by the same
SerRS that charges tRNA
Ser
, followed by the enzymatic
selenylation of the resulting Ser residue.
How does the ribosomal system differentiate a Sec-spec-
ifying UGA codon from a normal opal Stop codon? As we
saw to be the case with Glu–tRNA
Gln
(Section 32-2Cf), EF-
Tu, the elongation factor that conducts most aminoacyl–
tRNAs to the ribosome in a GTP-dependent process, does
not bind Sec–tRNA
Sec
.Instead it is bound by a specific elon-
gation factor named SELB, a homolog of EF-Tu, that in its
complex with GTP is recruited to a ribosomally bound
mRNA stem–loop structure in the selenoprotein coding re-
gion on the 3¿ side of the UGA codon specifying Sec.
Certain methanogenic (methane-producing) archaea
express the enzyme methylamine methyltransferase, which
contains the amino acid residue pyrrolysine (Pyl), a Lys
with its ε-nitrogen in amide linkage to a pyrroline group:
Unlike post-translationally modified Lys residues, such as
5-hydroxylysine (Hyl; Section 8-2B) and ε-N-acetyllysine
N
OC
C
O
CH
NH
CH
2
CH
2
CH
2
CH
2
CH
3
NH
The pyrrolysine (Pyl) residue
The selenocysteine
(Sec) residue
CH CH
2
NH
Se
C
H
O
Section 32-2. Transfer RNA and Its Aminoacylation 1361
JWCL281_c32_1338-1428.qxd 9/7/10 2:22 PM Page 1361
(Section 4-3B), Pyl is directly incorporated into proteins
during translation. Pyl is specified by the codon UAG
(normally the amber Stop codon). Pyl is carried to the ri-
bosome by tRNA
Pyl
, which contains a CUA anticodon and
differs from typical tRNAs in having a D loop with five
rather than eight residues, an anticodon stem with six
rather than five base pairs, and a TC loop that lacks the
sequence TC. A specific aminoacyl–tRNA synthetase,
PylRS, that differs from known LysRSs, charges tRNA
Pyl
with pyrrolysine in an ATP-dependent reaction, the first
known example in nature of the direct aminoacylation of a
tRNA with a “nonstandard” amino acid. Unlike the case
for Ser–tRNA
Sec
, Pyl–tRNA
Pyl
is delivered to the ribo-
some by EF-Tu. This suggests that the mRNA contains a
signal that causes UAG to be read as a Pyl codon rather
than as a Stop codon.A conserved stem–loop structure lo-
cated on the 3¿ side of UAG codons specifying Pyl may
comprise this signal. Alternatively, a Pyl–tRNA
Pyl
may oc-
casionally read a UAG codon and is therefore a type of
nonsense suppressor (see below).
E. Nonsense Suppression
Nonsense mutations are usually lethal when they prema-
turely terminate the synthesis of an essential protein. An
organism with such a mutation may nevertheless be “res-
cued” by a second mutation on another part of the genome.
For many years after their discovery, the existence of such
intergenic suppressors was quite puzzling. It is now known,
however, that they usually arise from mutations in a tRNA
gene that causes the tRNA to recognize a nonsense codon.
Such a nonsense suppressor tRNA appends its amino acid
(which is the same as that carried by the corresponding
wild-type tRNA) to a growing polypeptide in response to
the recognized Stop codon, thereby preventing chain ter-
mination.For example,the E.coli amber suppressor known
as su3 is a tRNA
Ty r
whose anticodon has mutated from the
wild-type GUA (which reads the Tyr codons UAU and
UAC) to CUA (which recognizes the amber Stop codon
UAG).An su3
E. coli with an otherwise lethal amber mu-
tation in a gene coding for an essential protein would be vi-
able if the replacement of the wild-type amino acid residue
by Tyr does not inactivate the protein.
There are several well-characterized examples of amber
(UAG), ochre (UAA), and opal (UGA) suppressors in
E. coli (Table 32-6). Most of them, as expected, have mu-
tated anticodons. UGA-1 tRNA, however, differs from the
wild type only by a G S A mutation in its D stem, which
changes a G U pair to a stronger A U pair.This mutation
apparently alters the conformation of the tRNA’s CCA
anticodon so that it can form an unusual wobble pairing
with UGA as well as with its normal codon, UGG.
Nonsense suppressors also occur in yeast.
a. Suppressor tRNAs Are Mutants of Minor tRNAs
How do cells tolerate a mutation that both eliminates a
normal tRNA and prevents the termination of polypeptide
synthesis? They survive because the mutated tRNA is usu-
ally a minor member of a set of isoaccepting tRNAs and be-
cause nonsense suppressor tRNAs must compete for Stop
codons with the protein factors that mediate the termination
of polypeptide synthesis (Section 32-3F). Consequently, the
rate of suppressor-mediated synthesis of active proteins with
either UAG or UGA nonsense mutations rarely exceeds
50% of the wild-type rate, whereas mutants with UAA, the
most common termination codon,have suppression efficien-
cies of 5%. Many mRNAs, moreover, have two tandem
Stop codons so that even if their first Stop codon were sup-
pressed, termination could occur at the second. Neverthe-
less, many suppressor-rescued mutants grow relatively
slowly because they cannot make an otherwise prematurely
terminated protein as efficiently as do wild-type cells.
Other types of suppressor tRNAs are also known. Mis-
sense suppressors act similarly to nonsense suppressors but
substitute one amino acid in place of another. Frameshift
suppressors have eight nucleotides in their anticodon loops
rather than the normal seven.They read a four-base codon
beyond a base insertion thereby restoring the wild-type
reading frame.
3 RIBOSOMES AND POLYPEPTIDE
SYNTHESIS
Ribosomes were first seen in cellular homogenates by
dark-field microscopy in the late 1930s by Albert Claude
who referred to them as “microsomes.” It was not until the
mid-1950s, however, that George Palade observed them in
cells by electron microscopy, thereby disposing of the con-
tention that they were merely artifacts of cell disruption.
The name ribosome derives from the fact that these parti-
cles in E. coli consist of approximately two-thirds RNA and
one-third protein. (Microsomes are now defined as the ar-
tifactual vesicles formed by the endoplasmic reticulum on
cell disruption. They are easily isolated by differential cen-
trifugation and are rich in ribosomes.) The correlation be-
tween the amount of RNA in a cell and the rate at which it
synthesizes protein led to the suspicion that ribosomes are
the site of protein synthesis. This hypothesis was con-
firmed in 1955 by Paul Zamecnik, who demonstrated that
14
C-labeled amino acids are transiently associated with
1362 Chapter 32. Translation
Table 32-6 Some E. coli Nonsense Suppressors
Name Codon Suppressed Amino Acid Inserted
su1 UAG Ser
su2 UAG Gln
su3UAG Tyr
su4 UAA, UAG Tyr
su5 UAA, UAG Lys
su6 UAA Leu
su7 UAA Gln
UGA-1 UGA Trp
UGA-2 UGA Trp
Source: Körner,A.M., Feinstein, S.I., and Altman, S., in Altman, S. (Ed.),
Transfer RNA, p. 109, MIT Press (1978).
JWCL281_c32_1338-1428.qxd 8/19/10 10:05 PM Page 1362
ribosomes before they appear in free proteins. Further re-
search showed that ribosomal polypeptide synthesis has
three distinct phases: (1) chain initiation, (2) chain elonga-
tion, and (3) chain termination.
In this section we examine the structure of the ribosome
and then outline the ribosomal mechanism of polypeptide
synthesis. In doing so we shall compare the properties of ri-
bosomes from prokaryotes with those of eukaryotes.
A. Ribosome Structure
The E. coli ribosome, which has a particle mass of ⬃2.5
10
6
D and a sedimentation coefficient of 70S, is a spher-
oidal particle that is ⬃250 Å across in its largest dimension.
It may be dissociated, as James Watson discovered, into
two unequal subunits (Table 32-7).The small (30S) subunit
consists of a 16S rRNA molecule and 21 different polypep-
tides, whereas the large (50S) subunit contains a 5S and a
23S rRNA together with 31 different polypeptides. The up
to 20,000 ribosomes in an E. coli cell account for ⬃80% of
its RNA content and ⬃10% of its protein.
Structural studies of the ribosome through electron mi-
croscopy began soon after its discovery. Three-dimensional
(3D) structures of the ribosome and its subunits at low (⬃50
Å) resolution first became available in the 1970s through
image reconstruction techniques, pioneered by Klug, in
which electron micrographs of a single particle or ordered
sheets of particles taken from several directions are com-
bined to yield its 3D image. The small subunit is a roughly
mitten-shaped particle, whereas the large subunit is spher-
oidal with three protuberances on one side (Fig. 32-26).
Section 32-3. Ribosomes and Polypeptide Synthesis 1363
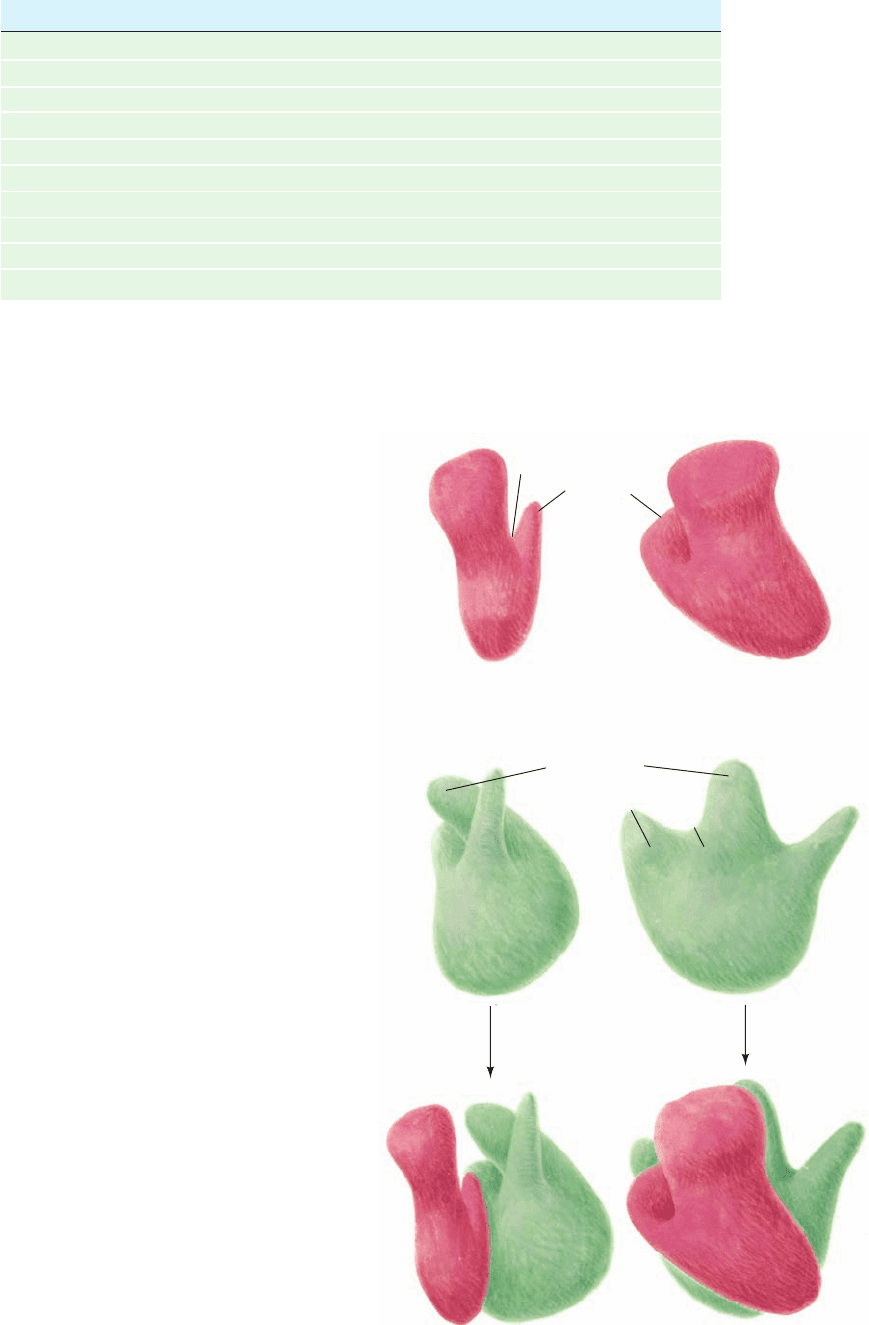
Figure 32-26 A low resolution model of the E. coli ribosome.
The small subunit (top; red) associates with the large subunit
(middle; green) to form the intact ribosome (bottom).Two
perpendicular views of each particle are provided.These models
are based on transmission electron micrographs of negatively
stained particles (in which the particle being imaged is embedded
in electron-absorbing heavy metal salts, thereby providing
contrast between the relatively electron-transparent particle and
the background).
Table 32-7 Components of E. coli Ribosomes
Ribosome Small Subunit Large Subunit
Sedimentation coefficient 70S 30S 50S
Mass (kD) 2520 930 1590
RNA
Major 16S, 1542 nucleotides 23S, 2904 nucleotides
Minor 5S, 120 nucleotides
RNA mass (kD) 1664 560 1104
Proportion of mass 66% 60% 70%
Proteins 21 polypeptides 31 polypeptides
Protein mass (kD) 857 370 487
Proportion of mass 34% 40% 30%
Head
Neck
Platform
Cleft
Body
Head
Neck
Body
Central
protuberance
Ridge
Valley
Stalk
Ribosome
++
JWCL281_c32_1338-1428.qxd 8/4/10 4:44 PM Page 1363
a. Ribosomal RNAs Have Complicated
Secondary Structures
The E. coli 16S rRNA, which was sequenced by Harry
Noller,consists of 1542 nucleotides.A computerized search
of this sequence for stable double helical segments yielded
many plausible but often mutually exclusive secondary
structures. However, the comparison of the sequences of
16S rRNAs from several prokaryotes, under the assump-
tion that their structures have been evolutionarily con-
served, led to the flowerlike secondary structure for 16S
rRNA seen in Fig. 32-27a. This four-domain structure is
54% base paired. Its double helical stems tend to be short
(8 bp) and many of them are imperfect. Intriguingly, elec-
tron micrographs of the 16S rRNA resemble those of the
complete 30S subunit, thereby suggesting that the 30S sub-
unit’s overall shape is largely determined by the 16S
rRNA. The large ribosomal subunit’s 5S and 23S rRNAs,
which consist of 120 and 2904 nucleotides, respectively,
have also been sequenced. As with the 16S rRNA, they
have extensive secondary structures (Fig. 32-27b).
b. Ribosomal Proteins Have Been
Partially Characterized
Ribosomal proteins are difficult to separate because
most of them are insoluble in ordinary buffers. By conven-
tion, ribosomal proteins from the small and large subunits
are designated with the prefixes S and L, respectively, fol-
lowed by a number indicating their position, from upper
left to lower right, on a two-dimensional gel electrophore-
togram (roughly in order of decreasing molecular mass;
Fig. 32-28). Only protein S20/L26 appears to be common to
both subunits. One of the large subunit proteins is partially
acetylated at its N-terminus so that it gives rise to two elec-
trophoretic spots (L7/L12). Four copies of this protein, a
dimer of dimers, are present in the large subunit. More-
over, these four copies of L7/L12 aggregate with L10 to
form a stable complex that was initially thought to be a
unique protein, “L8.” All the other ribosomal proteins oc-
cur in only one copy per subunit.
1364 Chapter 32. Translation
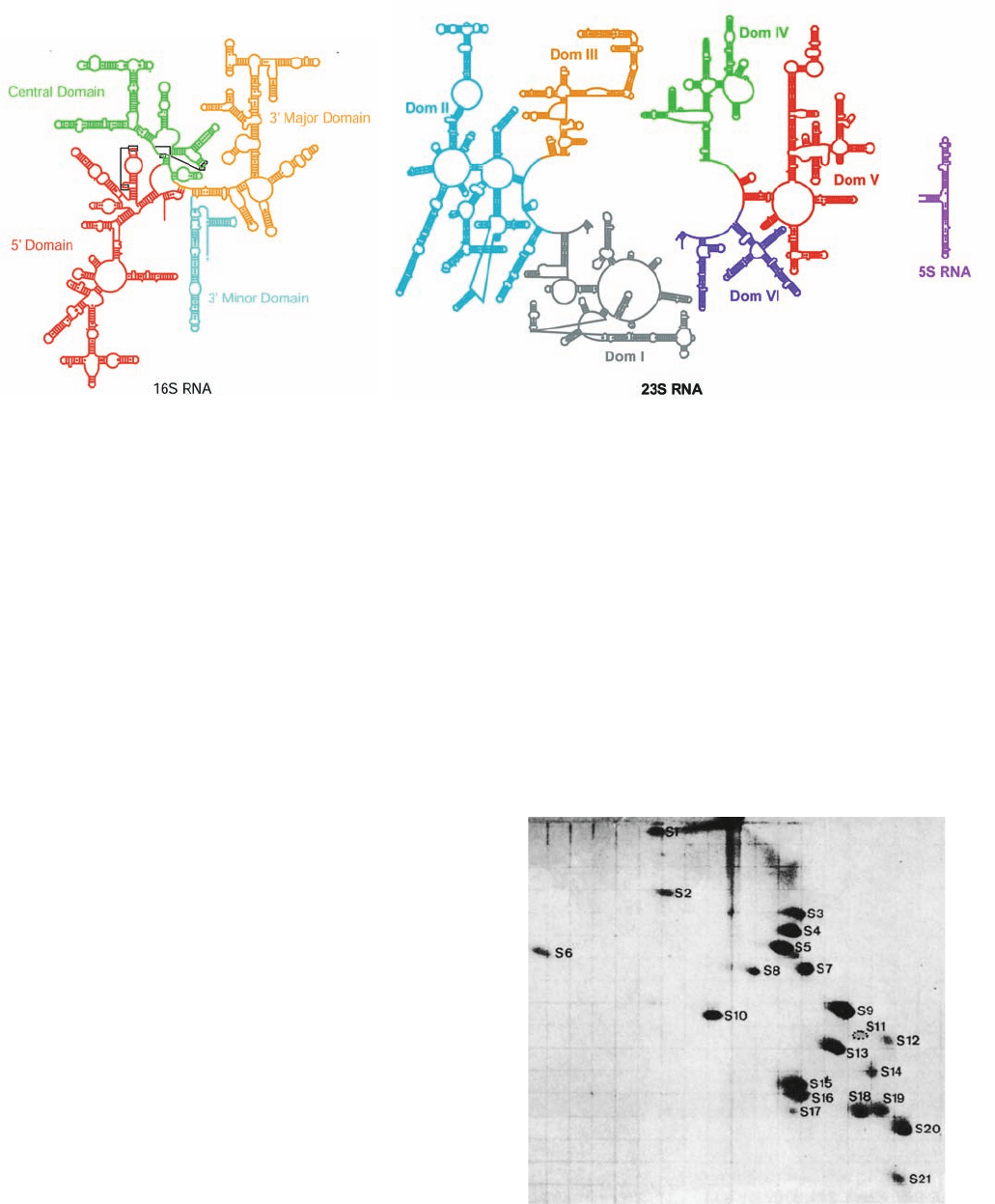
Figure 32-27 Secondary structures of the E. coli ribosomal
RNAs. (a) 16S RNA and (b) 23S and 5S RNAs. The rRNAs are
colored by domain with short lines spanning a stem representing
Watson–Crick base pairs, small dots representing G U base
pairs, and large dots representing other non-Watson–Crick base
Figure 32-28 Two-dimensional gel electrophoretogram of
E. coli small ribosomal subunit proteins. First dimension
(vertical): 8% acrylamide, pH 8.6; second dimension (horizontal):
18% acrylamide, pH 4.6. [From Kaltschmidt, E. and Wittmann,
H.G., Proc. Natl. Acad. Sci. 67, 1277 (1970).]
pairs. Note the flowerlike series of stems and loops forming each
domain. [Courtesy of Venki Ramakrishnan, MRC Laboratory of
Molecular Biology, Cambridge, U.K., and Peter Moore,Yale Uni-
versity. Adapted from diagrams in http://www.rna.ccbb.utexas.edu/.]
(a)
(b)
JWCL281_c32_1338-1428.qxd 8/19/10 10:05 PM Page 1364
The amino acid sequences of all 52 E. coli ribosomal pro-
teins were elucidated, mainly by Heinz-Günter Wittmann
and Brigitte Wittmann-Liebold. They range in size from 46
residues for L34 to 557 residues for S1.Most of these proteins,
which exhibit little sequence similarity with one another, are
rich in the basic amino acids Lys and Arg and contain few
aromatic residues as is expected for proteins that are closely
associated with polyanionic RNA molecules.
The X-ray and NMR structures of around half of the ri-
bosomal proteins or their fragments have been independ-
ently determined. These proteins have a wide variety of
structural motifs although most of their folds occur in other
proteins of known structure. Around one-third of these
ribosomal proteins contain the RNA-recognition motif
(RRM; Section 31-4Ab),which occurs in ⬎200 RNA-binding
proteins including rho factor (the transcriptional termination
protein, which contains four such motifs; Section 31-2Db),
poly(A) polymerase, poly(A)-binding protein (PABP), sev-
eral proteins involved in gene splicing (Section 31-4A), and
the translational initiation factor eIF4B (Section 32-3Cd).All
of these proteins presumably evolved from an ancient RNA-
binding protein.
c. Ribosomal Subunits Are Self-Assembling
Ribosomal subunits form, under proper conditions,
from mixtures of their numerous macromolecular compo-
nents. Ribosomal subunits are therefore self-assembling en-
tities. Masayasu Nomura determined the order in which
this occurs through partial reconstitution experiments. If
one macromolecular component is left out of an otherwise
self-assembling mixture of proteins and RNA, the other
components that fail to bind to the resulting partially as-
sembled subunit must somehow interact with the omitted
component.Through the analysis of a series of such partial
reconstitution experiments, Nomura constructed an as-
sembly map of the small (30S) subunit (Fig. 32-29). This
map indicates that the initial steps in small subunit assem-
bly are the independent binding to naked 16S rRNA of six
so-called primary (1°) binding proteins (S4, S7, S8, S15,
S17, and S20). The resulting assembly intermediates pro-
vide the molecular scaffolding for binding secondary (2°)
binding proteins, which after a significant conformational
change, form the attachment sites for tertiary (3°) binding
proteins.An analogous assembly map for the large subunit
was elucidated by Knud Nierhaus. The observation that
similar assembly intermediates occur in vivo and in vitro
suggests that in vivo and in vitro assembly processes are
much alike.
In the cell, the 16S RNA folds in an ordered manner
such that each domain is folded before the next domain is
transcribed.The assembly of the small ribosomal subunit is
then facilitated by a variety of assembly factors, proteins
that bind to immature complexes but not to mature sub-
units. Many assembly factors associate with segments of
the 16S RNA that change conformation during the latter
stages of assembly. Presumably, the assembly of the large
ribosomal subunit follows a similar course.
d. The Atomic Structure of the Prokaryotic
Ribosome Has Been Long in Coming
The elucidation of the ribosome’s atomic structure was
a tortuous affair extending over four decades in which
slow incremental improvements were occasionally punc-
tuated by significant technical gains. The process began in
the 1960s with shadowy transmission electron micro-
graphs that provided only rough 2D shapes. This was fol-
lowed in the 1970s by image reconstruction techniques
that generated 3D models although still at low resolution
(Fig. 32-26). Later in the 1970s, the sites of many of the ri-
bosome’s proteins were determined by James Lake and
Georg Stöffler through immune electron microscopy, a
technique in which antibodies raised against a particular
ribosomal protein are used to mark its position in electron
micrographs of the antibody complexed to a ribosomal
subunit. These results were improved and extended in the
1980s by neutron scattering experiments conducted by
Donald Engelman and Peter Moore on the 30S subunit,
which indicated the distances between the centers of mass of
its component proteins and hence their three-dimensional
distribution. These structural studies were supplemented by
a variety of chemical cross-linking and fluorescence transfer
studies that demonstrated the proximity of various riboso-
mal components.
The molecular structure of the prokaryotic ribosome
began to come into focus in the mid-1990s through the de-
velopment of cryoelectron microscopy (cryo-EM). In this
technique, the sample is cooled to near liquid N
2
tempera-
tures (–196°C) so rapidly (in a few milliseconds) that the
water in the sample does not have time to crystallize but,
Section 32-3. Ribosomes and Polypeptide Synthesis 1365
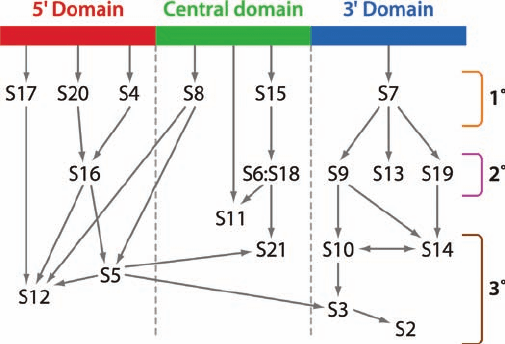
Figure 32-29 Assembly map of the E. coli small subunit. The
map is organized according to the domains of the 16S RNA (Fig.
32-27) with arrows indicating the facilitation of binding. For
example, the arrow from the 5¿ domain of the 16S rRNA to S20
indicates that S20 binds directly to the 16S rRNA in the absence
of other proteins and is therefore a primary (1°) binding protein;
the arrow from S20 to S16 indicates that S20 facilitates the
binding of S16, which is therefore a secondary (2°) binding
protein; and the arrows from S16 to S5 and S12 indicate that S5
and S12 are tertiary (3°) binding proteins. [Courtesy of James
Williamson, The Scripps Research Institute, La Jolla, California.]
JWCL281_c32_1338-1428.qxd 10/27/10 1:40 PM Page 1365
rather, assumes a vitreous (glasslike) state. Consequently,
the sample remains hydrated and hence retains its native
shape to a greater extent than in conventional electron mi-
croscopy (in which the sample is vacuum dried). Studies,
carried out in large part by Joachim Frank, revealed the po-
sitions where tRNAs and mRNA as well as various soluble
protein factors bind to the ribosome (Fig. 32-30). The high-
est resolution achieved by cryo-EM of ribosomes has grad-
ually improved over the years to ⬃8 Å.
Ribosomal subunits were first crystallized by Ada
Yonath in 1980 although they diffracted X-rays poorly.
Over the course of several years, however, the quality of
these crystals were incrementally improved until, in 1991,
Yonath reported crystals of the 50S subunit that diffracted
X-rays to 3-Å resolution. It was not until later in the 1990s,
however,that technology was up to the task of determining
the X-ray structures of these gargantuan molecular com-
plexes. In 2000, the annus mirabilis (miracle year) of ribo-
somology, Moore and Steitz reported the X-ray structure
of the 50S ribosomal subunit of the halophilic (salt-loving)
bacterium Haloarcula marismortui at atomic (2.4-Å) reso-
lution and Venki Ramakrishnan and Yonath independently
reported the X-ray structure of the 30S subunit of T. ther-
mophilus at ⬃3-Å resolution. In 2001, Noller reported the
5.5-Å resolution structure of the entire T. thermophilus ri-
bosome, which was gradually improved to 2.8 Å. In addi-
tion, the structures of the E. coli ribosome, by Jamie Cate,
and the large subunit from Deinococcus radiodurans, by
Yonath, have been determined. In the following para-
graphs we discuss the properties of these groundbreaking
structures. We consider their functional implications start-
ing in Section 32-3C.
e. Ribosomal Architecture
Several generalizations can be made about ribosomal
architecture based on the structures of the 30S and 50S
subunits:
1. Both the 16S and 23S rRNAs are assemblies of
coaxially stacked helical elements connected by loops,
most of which are irregular extensions of helices (Fig. 32-
31).These structures, which are in close accord with previ-
ous secondary structure predictions (Fig. 32-27), are stabi-
lized by interactions between helices such as minor
groove to minor groove packing, which also occur in the
structures of the group I intron and RNase P (Sections 31-
4Af and 31-4Ca; recall that A-form RNA has a very shal-
low minor groove); the insertion of a phosphate ridge into
a minor groove; and adenines that are distant in sequence
but often highly conserved that are inserted into minor
grooves. The overall shapes of these subunits are rela-
tively flat rather than globular with most regions having a
thickness of two or three helical diameters [in contrast,
other large (100 nt) RNAs of known structure, such as
the group I intron, RNase P, and the RNA from the signal
recognition particle (Section 12-4Bb), are only one helix
thick]. Although the determination of the structures of
the 30S and 50S ribosomal subunits increased the amount
of RNA structure that was then known at atomic resolu-
tion by ⬃10-fold, nearly all of the secondary structural
motifs seen in the ribosome also occur in smaller RNA
structures. This suggests that the repertoire of RNA sec-
ondary structural motifs is limited.
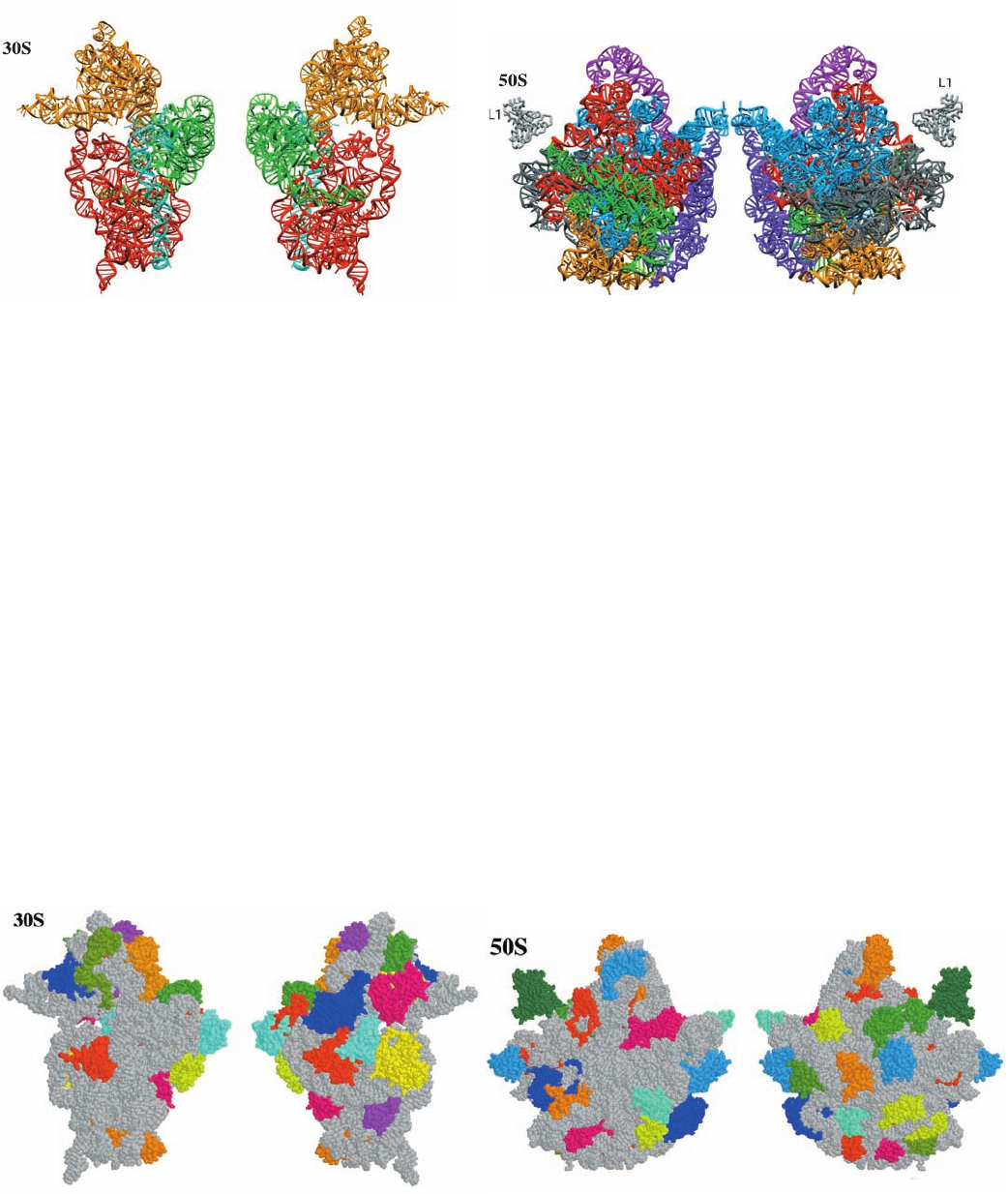
2. Each of the 16S RNA’s four domains, which extend
out from a central junction (Fig. 32-27a), forms a morpho-
logically distinct portion of the 30S subunit (Fig. 32-31a):
The 5¿ domain forms most of the body (Fig. 32-26), the
central domain forms the platform, the 3¿ major domain
forms the entire head, and the 3¿ minor domain, which
consists of just two helices, is located at the interface be-
tween the 30S and 50S subunits. In contrast, the 23S RNA’s
six domains (Fig. 32-27b) are intricately intertwined in the
50S subunit (Fig. 32-31b). Since the ribosomal proteins are
embedded in the RNA (see below), this suggests that the
domains of the 30S subunit can move relative to one
1366 Chapter 32. Translation
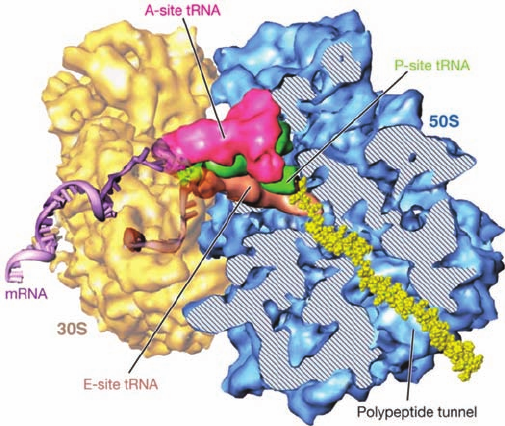
Figure 32-30 Cryoelectron microscopy–based image of the E.
coli ribosome. The 30S subunit (yellow) is on the left and the 50S
subunit (blue) is on the right.The tRNAs that occupy the A, P,
and E sites (Section 32-3B) are colored magenta, green, and
brown.A portion of the 50S subunit has been cut away to reveal
the polypeptide exit tunnel.A segment of mRNA (5¿ end brown
and 3¿ end lavender) and the nascent polypeptide chain (yellow)
have been modeled into the structure. [Courtesy of Joachim
Frank, State University of New York at Albany.]
JWCL281_c32_1338-1428.qxd 8/4/10 4:44 PM Page 1366
(a)
Interface view
Back view
Interface View Back View
another during protein synthesis, whereas the 50S subunit
appears to be rigid.
3. The distribution of the proteins in the two ribosomal
subunits is not uniform (Fig. 32-32). The vast majority of
the ribosomal proteins are located on the back and sides of
their subunits. In contrast, the face of each subunit that
forms the interface between the two subunits, particularly
those regions that bind the tRNAs and mRNA (see be-
low), is largely devoid of proteins.
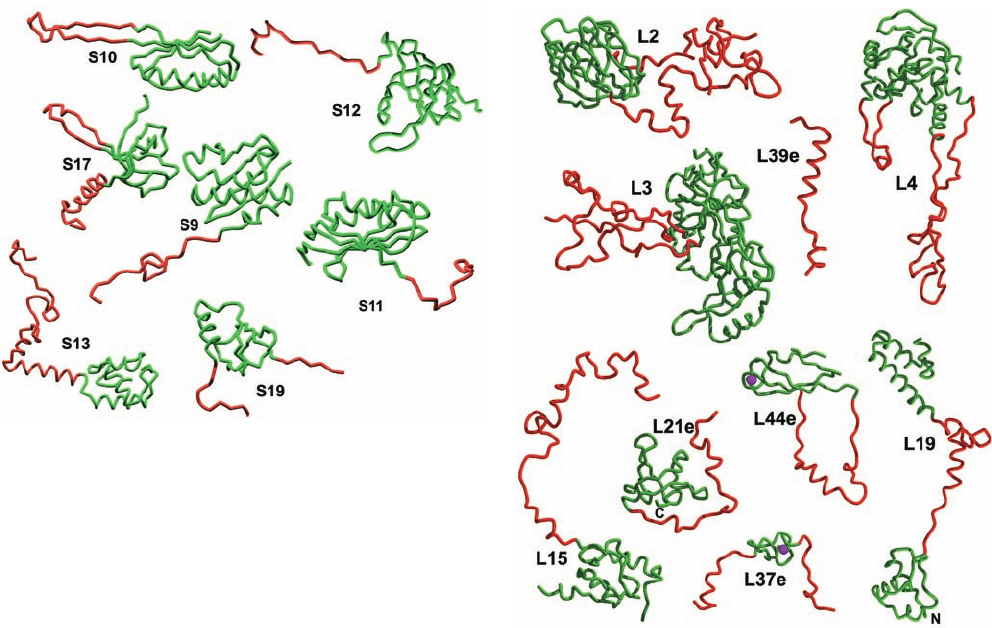
4. Most ribosomal proteins consist of a globular do-
main, which is, for the most part, located on a subunit sur-
Section 32-3. Ribosomes and Polypeptide Synthesis 1367
Figure 32-31 Tertiary structures of the ribosomal RNAs. (a)
The 16S rRNA of T. thermophilus. (b) The 23S rRNA of H.
marismortui. The rRNAs are colored according to domain as in
Fig. 32-27. The interface view of a ribosomal subunit (left) is
toward its surface that associates with the other subunit in the
whole ribosome and the back view (right) is from the opposite
(solvent-exposed) side. Note that the secondary structure
Figure 32-32 Distribution of protein and RNA in the
ribosomal subunits. (a) The 30S subunit of T. thermophilus. (b)
The 50S subunit of H. marismortui. The subunits are drawn in
space-filling form with their RNAs gray and their proteins in
various colors. Note that the interface side of each subunit is
largely free of protein, particularly in its regions that interact
(b)
Interface view
Back view
with mRNA and tRNAs. [Part a based on an X-ray structure by
Venki Ramakrishnan, MRC Laboratory of Molecular Biology,
Cambridge, U.K. Part b based on an X-ray structure by Peter
Moore and Thomas Steitz, Yale University. PDBids 1J5E and
1JJ2.]
domains of the 16S rRNA fold as separate tertiary structure
domains, whereas in the 23S rRNA the secondary structure
domains are convoluted together. [Courtesy of Venki
Ramakrishnan, MRC Laboratory of Molecular Biology,
Cambridge, U.K., and Peter Moore, Yale University. PDBids
1J5E and 1JJ2.]
Interface View
Back View
face (Fig. 32-32), and a long segment that is largely devoid
of secondary structure and unusually rich in basic residues
that infiltrates between the RNA helices into the subunit
interior (Fig. 32-33). Indeed, a few ribosomal proteins lack
a globular domain altogether (e.g., L39e in Fig. 32-33b). Ri-
bosomal proteins make far fewer base-specific interactions
than do other known RNA-binding proteins. They tend to
interact with the RNA through salt bridges between their
positively charged side chains and the RNAs’ negatively
charged phosphate oxygen atoms, thereby neutralizing the
repulsive charge–charge interactions between nearby
RNA segments. This is consistent with the hypothesis that
(a)
(b)
JWCL281_c32_1338-1428.qxd 8/4/10 4:44 PM Page 1367
the primordial ribosome consisted entirely of RNA (the
RNA world) and that the proteins that were eventually ac-
quired stabilized its structure and fine-tuned its function.
The X-ray structure of the entire T. thermophilus ribo-
some in complex with three tRNAs and a 11-nt mRNA
segment was determined by Ramakrishnan (Fig. 32-34; ri-
bosomes, as we shall see in Section 32-3B, have three func-
tionally distinct tRNA-binding sites known as the A, P, and
E sites). The structures of the 30S and 50S subunits in this
enormous molecular machine closely resemble those of the
isolated subunits although there are several regions at the
subunit interface that exhibit significant conformational
shifts, which suggests that these changes occur as a conse-
quence of subunit association. In addition, several disor-
dered portions of the isolated H. marismortui 50S subunit
are ordered in the intact T. thermophilus ribosome, al-
though this may be a consequence of the latter’s greater
thermal stability.
The ribosome binds all three tRNAs in a similar manner
with their anticodon stem–loops bound to the 30S subunit
and their remaining portions, the D stem, elbow, and accep-
tor stem, bound to the 50S subunit. These interactions,
which mainly consist of RNA–RNA contacts, are made to
the tRNAs’ universally conserved segments, thereby per-
mitting the ribosome to bind different species of tRNAs in
the similar ways.
The small and large ribosomal subunits contact each
other at 12 positions via RNA–RNA, protein–protein, and
RNA–protein bridges. These intersubunit bridges have a
distinct distribution: The RNA–RNA bridges are centrally
located adjacent to the three bound tRNAs, whereas the
protein–protein and RNA–protein bridges are peripherally
located away from the ribosome’s functional sites. The
RNA–RNA contacts consist mainly of minor groove–minor
groove interactions although major groove, loop, and back-
bone contacts also occur. In the RNA–protein bridges, the
proteins contact nearly all types of RNA features including
major groove, minor groove, backbone, and loop elements.
We discuss the path of the mRNA and how it interacts
with the tRNAs in Section 32-3D. There we shall see that
the large subunit is mainly involved in mediating biochemi-
cal tasks such as catalyzing the reactions of polypeptide
elongation, whereas the small subunit is the major actor in
ribosomal recognition processes such as mRNA and tRNA
binding (although, as we have seen, the large subunit also
participates in tRNA binding).We shall also see that rRNA
has the major functional role in ribosomal processes (recall
that RNA has demonstrated catalytic properties; Sections
31-4Ae and 31-4Ca).
f. Eukaryotic Ribosomes Are Larger and More
Complex than Prokaryotic Ribosomes
Although eukaryotic and prokaryotic ribosomes resem-
ble one another in both structure and function, they differ
in nearly all details. Eukaryotic ribosomes have particle
masses in the range 3.9 to 4.5 10
6
D and have a nominal
1368 Chapter 32. Translation
Figure 32-33 Gallery of ribosomal protein structures. Proteins
from (a) the 30S subunit and (b) the 50S subunit.The proteins
are represented by their backbones with their globular portions
green and their highly extended segments red.The globular
portions are exposed on the surface of their associated subunit
(Fig. 32-32), whereas the extended segments are largely buried in
the RNA. The Zn
2
ions bound by L37e and L44e are represented
by magenta spheres. [Courtesy of Venki Ramakrishnan, MRC
Laboratory of Molecular Biology, Cambridge, U.K., and Peter
Moore, Yale University. PDBid 1J5E.]
(a)
(b)
JWCL281_c32_1338-1428.qxd 8/4/10 4:45 PM Page 1368
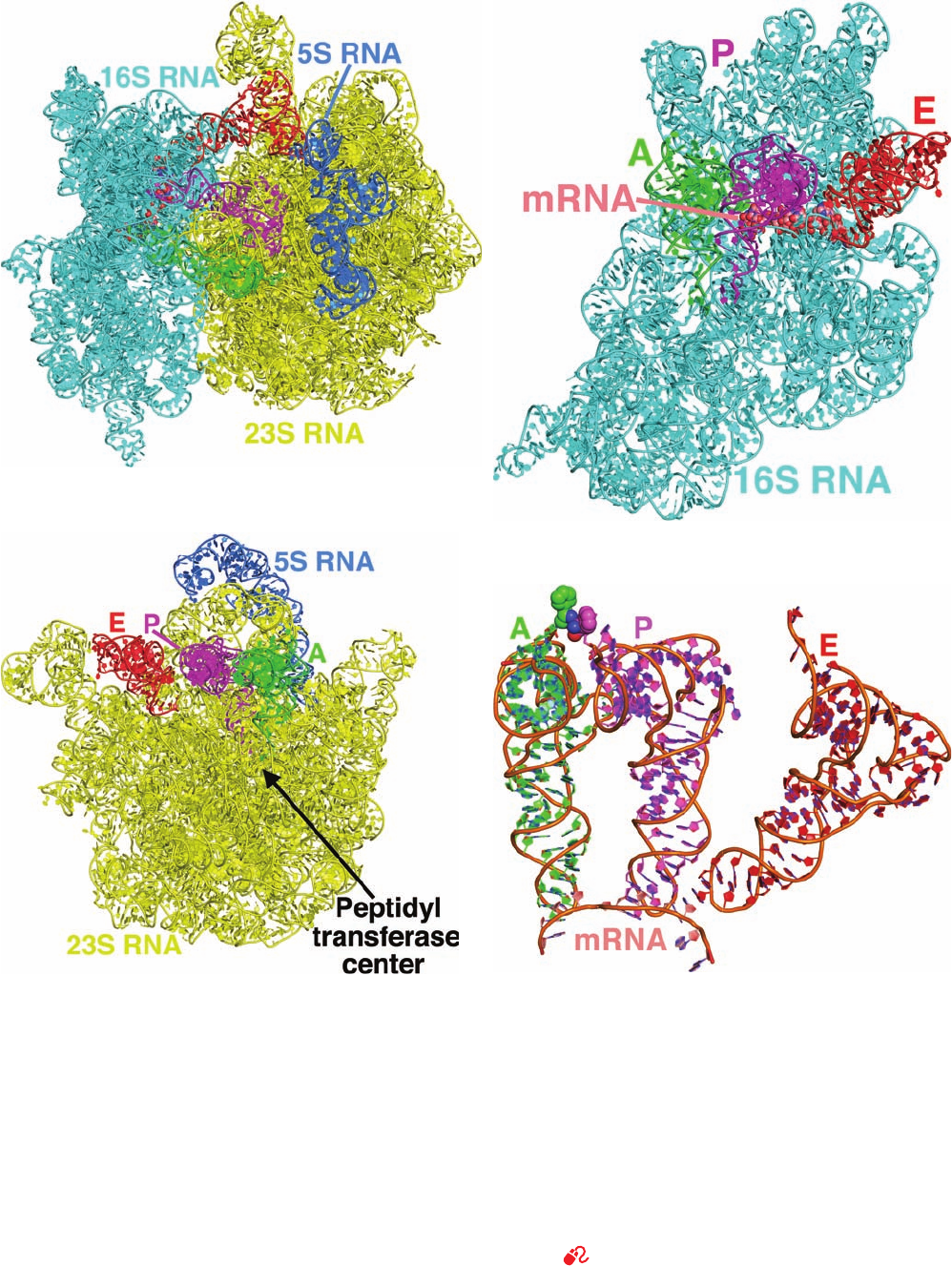
Section 32-3. Ribosomes and Polypeptide Synthesis 1369
(a) (b)
(c)
(d)
represented as in Part a.(c) The 23S RNA in interface view
(rotated 180° about the vertical direction relative to Part b) with
its bound tRNAs all represented as in Part a.(d) The interactions
of the tRNAs with the mRNA.This assembly is drawn in cartoon
form with A-site C green, P-site C magenta, E-site C red, mRNA
C pink, N blue, and O red and with successive P atoms connected
by orange rods.The Phe residues appended to the A- and P-site
tRNAs are represented in space-filling form. Note the close
approach of these Phe residues and that the tRNAs in the A and
P sites, but not that in the E site, form based paired codon–
anticodon interactions with the mRNA. [Based on an X-ray
structure by Venki Ramakrishnan, MRC Laboratory of
Molecular Biology, Cambridge, U.K. PDBids 2WDK and
2WDL.]
See Interactive Exercise 44.
Figure 32-34 X-ray structure of the T. thermophilus ribosome
in complex with tRNA and mRNA at 3.5 Å resolution. The E
site binds tRNA
Phe
and the A and P sites bind Phe–tRNA
Phe
(in
which the O atom forming an ester linkage from Phe to O3¿ of
the 3¿ terminal A has been replaced by an NH group to prevent
the hydrolysis of the Phe¬tRNA linkage). (a) The RNA
components of the ribosomal complex (its proteins are omitted
for clarity) drawn in cartoon form except for the 11-residue
mRNA, which is shown in space-filling form.The 16S RNA is
cyan, the 23S RNA is yellow, the 5S RNA is blue, the tRNAs in
the A, P, and E sites are green, magenta, and red, and the mRNA,
which is largely occluded by the 16S RNA, is colored according
to atom type with C pink, N blue, O red, and P orange. (b) The
16S RNA in interface view with its bound tRNAs and mRNA, all
JWCL281_c32_1338-1428.qxd 10/19/10 8:23 AM Page 1369

sedimentation coefficient of 80S. They dissociate into two
unequal subunits that have compositions that are distinctly
different from those of prokaryotes (Table 32-8; compare
with Table 32-7).The small (40S) subunit of the rat liver cy-
toplasmic ribosome, which together with the yeast ribo-
some is the most well-characterized eukaryotic ribosome,
consists of 33 unique polypeptides and an 18S rRNA. Its
large (60S) subunit contains 49 different polypeptides and
three rRNAs of 28S, 5.8S, and 5S. The additional complex-
ity of the eukaryotic ribosome relative to its prokaryotic
counterpart is presumably due to the eukaryotic ribo-
some’s additional functions: Its mechanism of translational
initiation is more complex (Section 32-3Cd); it must be
transported from the nucleus, where it is formed, to the cy-
toplasm, where translation occurs; and the machinery with
which it participates in the secretory pathway is more com-
plicated (Section 12-4B).
Sequence comparisons of the corresponding rRNAs
from various species indicates that evolution has conserved
their secondary structures rather than their base sequences
(Figs. 32-27a and 32-35). For example, a G C in a base
paired stem of E. coli 16S rRNA has been replaced by an A
U in the analogous stem of yeast 18S rRNA. The 5.8S
rRNA, which occurs in the large eukaryotic subunit in base
paired complex with the 28S rRNA, is homologous in se-
quence to the 5¿ end of prokaryotic 23S rRNA.Apparently
5.8S RNA arose through mutations that altered rRNA’s
post-transcriptional processing, producing a fourth rRNA.
1370 Chapter 32. Translation
Figure 32-35 Predicted secondary structures of evolutionarily
distant 16S-like rRNAs. (a) Archaebacteria (Halobacterium
volcanii), (b) eukaryotes (S. cerevisiae), and (c) mammalian
mitochondria (bovine). Compare them with Fig. 32-27a, the
secondary structure of 16S RNA from eubacteria (E. coli). Note
the close similarities of these assemblies; they differ mostly by
Table 32-8 Components of Rat Liver Cytoplasmic Ribosomes
Ribosome Small Subunit Large Subunit
Sedimentation coefficient 80S 40S 60S
Mass (kD) 4220 1400 2820
RNA
Major 18S, 1874 nucleotides 28S, 4718 nucleotides
Minor 5.8S, 160 nucleotides
5S, 120 nucleotides
RNA mass (kD) 2520 700 1820
Proportion of mass 60% 50% 65%
Proteins 33 polypeptides 49 polypeptides
Protein mass (kD) 1700 700 1000
Proportion of mass 40% 50% 35%
insertions and deletions of stem-and-loop structures.The
23S-like rRNAs from a variety of species likewise have similar
secondary structures. [After Gutell, R.R., Weiser, B., Woese, C.R.,
and Noller, H.F., Prog. Nucleic Acid Res. Mol. Biol. 32, 183
(1985).]
(a) (b) (c)
JWCL281_c32_1338-1428.qxd 8/4/10 4:45 PM Page 1370