Urry D.W. (Ed.) What Sustains Life? : Consilient Mechanisms for Protein-Based Machines and Materials
Подождите немного. Документ загружается.

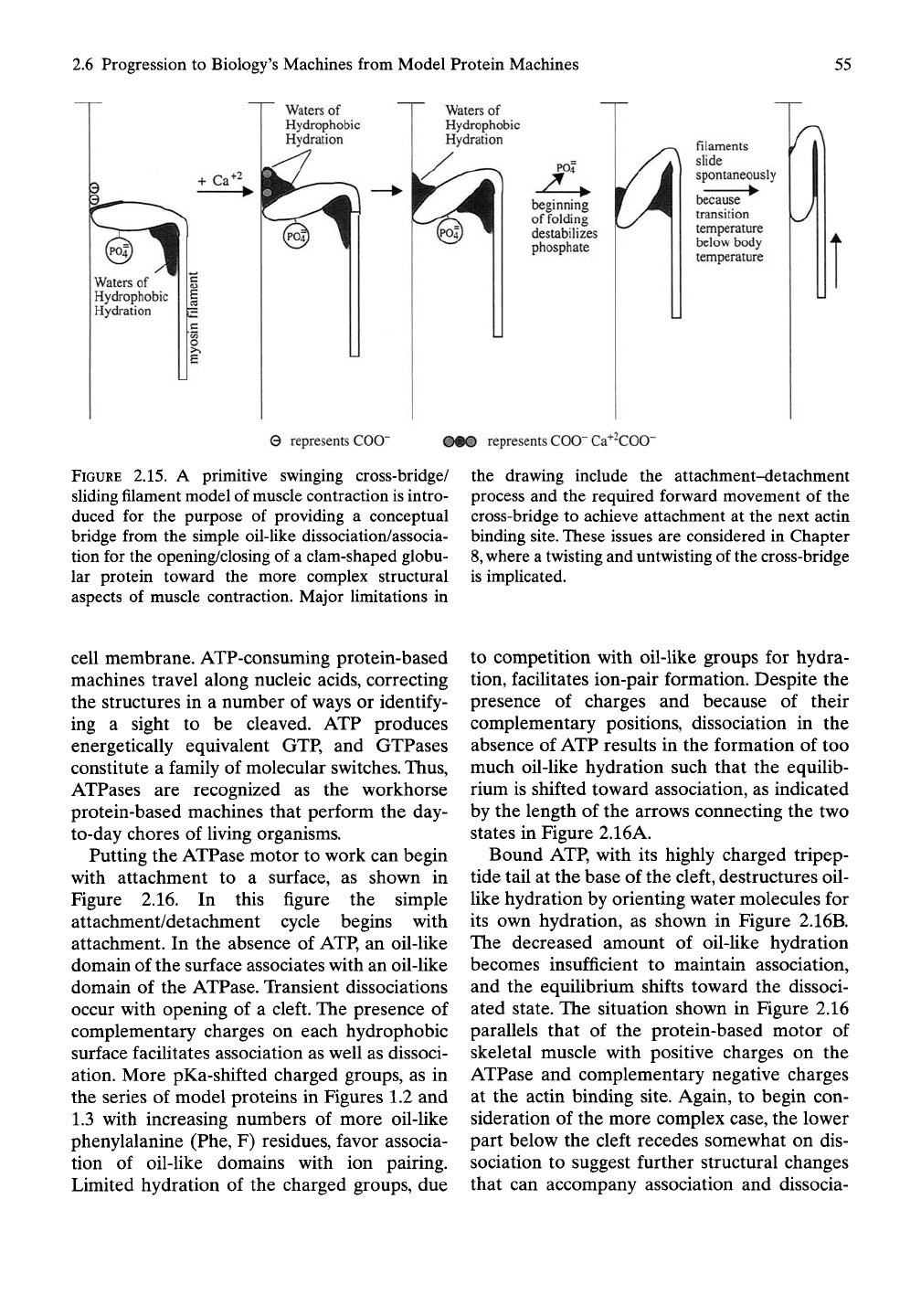
2.6 Progression to Biology's Machines from Model Protein Machines
55
Waters of
Hydrophobic
Hydration
Waters of
Hydrophobic
Hydration
PO4
beginning
of folding
destabilizes
phosphate
m
0 represents COO
FIGURE
2.15. A primitive swinging cross-bridge/
sUding filament model of muscle contraction is intro-
duced for the purpose of providing a conceptual
bridge from the simple oil-like dissociation/associa-
tion for the opening/closing of a clam-shaped globu-
lar protein toward the more complex structural
aspects of muscle contraction. Major limitations in
filaments
slide
spontaneously
because
transition
temperature
below body
temperature
»
t
represents
COO"
Ca"^^COO"
the drawing include the attachment-detachment
process and the required forward movement of the
cross-bridge to achieve attachment at the next actin
binding site. These issues are considered in Chapter
8,
where a twisting and untwisting of the cross-bridge
is implicated.
cell membrane. ATP-consuming protein-based
machines travel along nucleic acids, correcting
the structures in a number of ways or identify-
ing a sight to be cleaved. ATP produces
energetically equivalent GTP, and GTPases
constitute a family of molecular switches. Thus,
ATPases are recognized as the workhorse
protein-based machines that perform the day-
to-day chores of living organisms.
Putting the ATPase motor to work can begin
with attachment to a surface, as shown in
Figure 2.16. In this figure the simple
attachment/detachment cycle begins with
attachment. In the absence of ATP, an oil-like
domain of the surface associates with an oil-like
domain of the ATPase. Transient dissociations
occur with opening of a cleft. The presence of
complementary charges on each hydrophobic
surface facilitates association as well as dissoci-
ation. More pKa-shifted charged groups, as in
the series of model proteins in Figures 1.2 and
1.3 with increasing numbers of more oil-like
phenylalanine (Phe, F) residues, favor associa-
tion of oil-like domains with ion pairing.
Limited hydration of the charged groups, due
to competition with oil-like groups for hydra-
tion, facilitates ion-pair formation. Despite the
presence of charges and because of their
complementary positions, dissociation in the
absence of ATP results in the formation of too
much oil-like hydration such that the equilib-
rium is shifted toward association, as indicated
by the length of the arrows connecting the two
states in Figure 2.16A.
Bound ATP, with its highly charged tripep-
tide tail at the base of the cleft, destructures oil-
like hydration by orienting water molecules for
its own hydration, as shown in Figure 2.16B.
The decreased amount of oil-like hydration
becomes insufficient to maintain association,
and the equilibrium shifts toward the dissoci-
ated state. The situation shown in Figure 2.16
parallels that of the protein-based motor of
skeletal muscle with positive charges on the
ATPase and complementary negative charges
at the actin binding site. Again, to begin con-
sideration of the more complex case, the lower
part below the cleft recedes somewhat on dis-
sociation to suggest further structural changes
that can accompany association and dissocia-
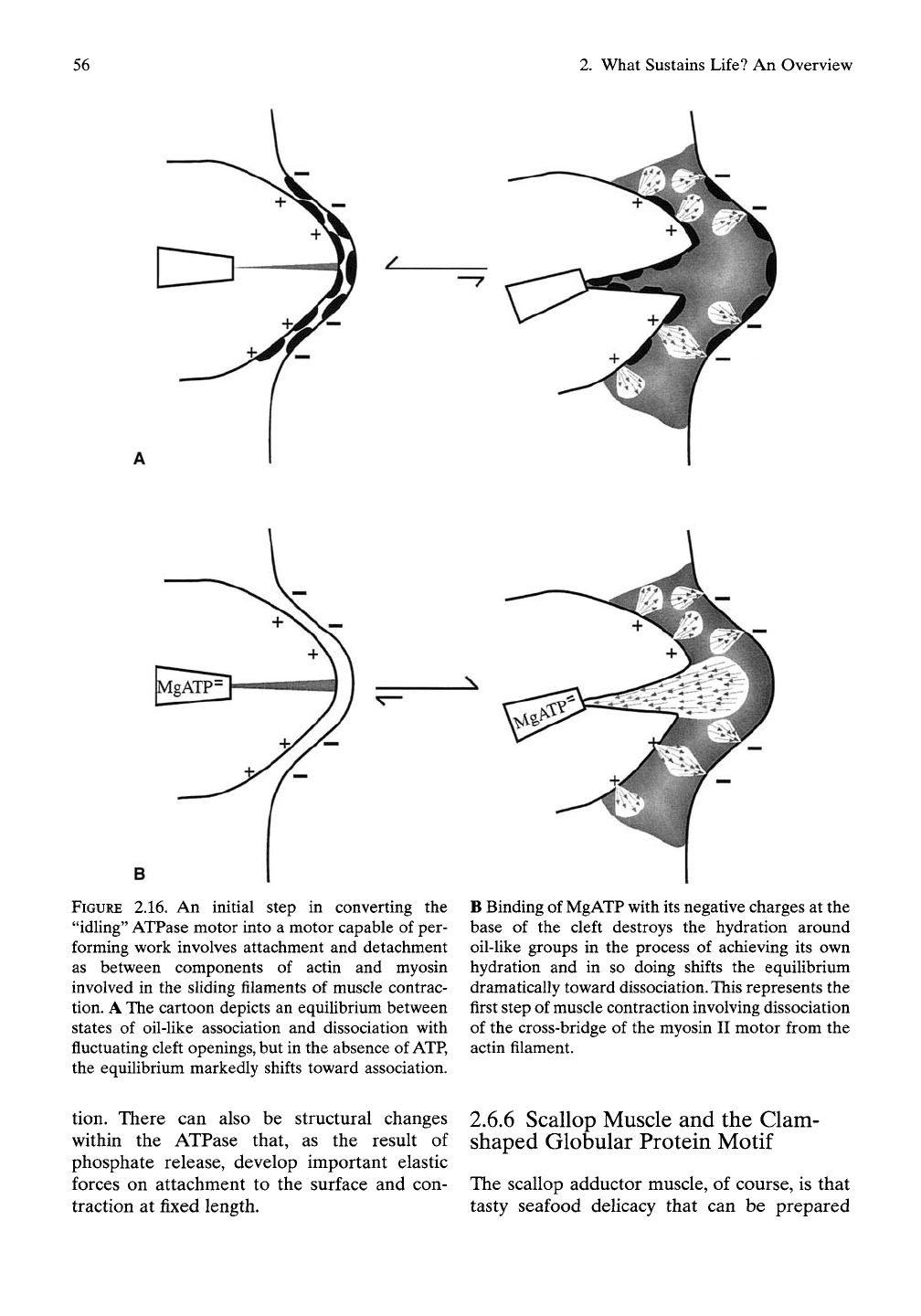
56
2.
What Sustains Life? An Overview
FIGURE 2.16. An initial step in converting the
"idUng"
ATPase motor into a motor capable of per-
forming work involves attachment and detachment
as between components of actin and myosin
involved in the sUding filaments of muscle contrac-
tion. A The cartoon depicts an equilibrium between
states of oil-like association and dissociation with
fluctuating cleft openings, but in the absence of
ATP,
the equilibrium markedly shifts toward association.
B Binding of MgATP with its negative charges at the
base of the cleft destroys the hydration around
oil-Uke groups in the process of achieving its own
hydration and in so doing shifts the equilibrium
dramatically toward dissociation. This represents the
first step of muscle contraction involving dissociation
of the cross-bridge of the myosin II motor from the
actin filament.
tion. There can also be structural changes
within the ATPase that, as the result of
phosphate release, develop important elastic
forces on attachment to the surface and con-
traction at fixed length.
2.6.6 Scallop Muscle and the Clam-
shaped Globular Protein Motif
The scallop adductor muscle, of course, is that
tasty seafood delicacy that can be prepared

2.6 Progression to Biology's Machines from Model Protein Machines
57
fried, baked, or even broiled. For the scallop it
functions to open and close the two halves of
this beautiful classic bivalve shell. Importantly,
the crystal structure data have been obtained of
the cross-bridge of scallop muscle from the
myosin filament to the site for actin binding,
and the cross-bridge has been obtained in two
different states, a near rigor state and a state
with an ATP
analogue.^^'^^
Somewhat ironically,
the structure of the cross-bridge provides an
actual analogue of the clam-shaped globular
protein in Figure 2.14A.
The crystal structure is of the SI cross-bridge
that contains the globular head, the a-helical
lever arm, and its wrappings, the essential
(ELC) and regulatory (RLC) hght chains,
which seem to contribute by strengthening the
single helical rod. For our purposes here, the
light chains will be removed in order to make
apparent the changes that occur when going
between contracted and ATP bound states. As
we will see, ATP binding opens the hydropho-
bic association between key components of
the cross-bridge, which bears analogy to the
binding of a single phosphate to open the
hydrophobically associated clam-shaped globu-
lar protein.
tion, that is, causes opening of the hydrophobi-
cally closed association.
2.6.6.1.2 Structural Reorientation on
Dephosphorylation
The long limb in Figure 2.17 is the a-helical
lever arm that in part connects the myosin fila-
ment to the globular part of the cross-bridge.
The stump or pedicle-like structure is the N-
terminal domain. In Figure 2.17A,C, the view is
in a direction along the myosin filament,
whereas in Figure 2.17B,D, the view is approx-
imately perpendicular to the myosin filament as
may be judged from the foot-like terminus that
is taken as being directed parallel to the myosin
filament.
Thus,
dephosphorylation results in a
rotation of the cross-bridge around the head of
the lever arm to tuck it under the stump of the
N-terminal domain. As expected from the con-
sihent mechanism, loss of the negatively
charged phosphate results in association of oil-
like domains. As considered in the text associ-
ated with Figures 8.47 through 8.57, this
hydrophobic association provides the power
stroke of muscle contraction.
2.6.6.1 Binding of ATP Analogue to
Cross-bridge ''Opens'' the Oil-like
Associated State as Occurs with the Clam-
shaped Globular Protein Motif
2.6.6.1.1 The Process of Muscle Contraction
Itself Can Be Viewed as a Variation on the
Clam-Shaped Globular Protein Theme
Figure 2.17 gives stereo views obtained from
the crystal structure of the cross-bridge of the
scallop myosin II motor in the contracted near-
rigor state in A (from the side of the ATP
binding site) and C (from the side opposite the
ATP binding site) and in the bound ATP ana-
logue state in B (from the side of the ATP
binding site) and in D (from the side opposite
the ATP binding site). Incredibly, as a direct
analogy to the hydrophobically closed clam-
shaped globular protein in Figure 2.14A, the
binding of the negatively charged ATP with its
triphosphate tail causes hydrophobic dissocia-
2.6.6.2 The Clefts Within Which the ATP
Resides Act as Conduits to Direct
the Forces Emanating from the
Thirsty Phosphates
The cleft at which the ATP resides runs in two
directions, with the y-phosphate positioned to
direct its thirst for water toward the actin
binding site and toward the open clam-like
region between upper lever arm and pedicle of
the N-terminal domain. Seen from the front, the
actin binding
side,
the ATP site resides in a cleft
under an overhang with its y-phosphate in the
depths at the aqueous edge of the cleft. The
cleft runs in two directions. In one direction in
Figure 2.17B the cleft runs outward and upward
at about 10 o'clock to the junction of the
myosin head with the actin filament, such that
ATP binding disrupts attachment to the actin
filament by the repulsion between oil-like and
charged species. In the other direction the cleft
runs to the oil-like association between the
head of the lever arm and the N-terminal
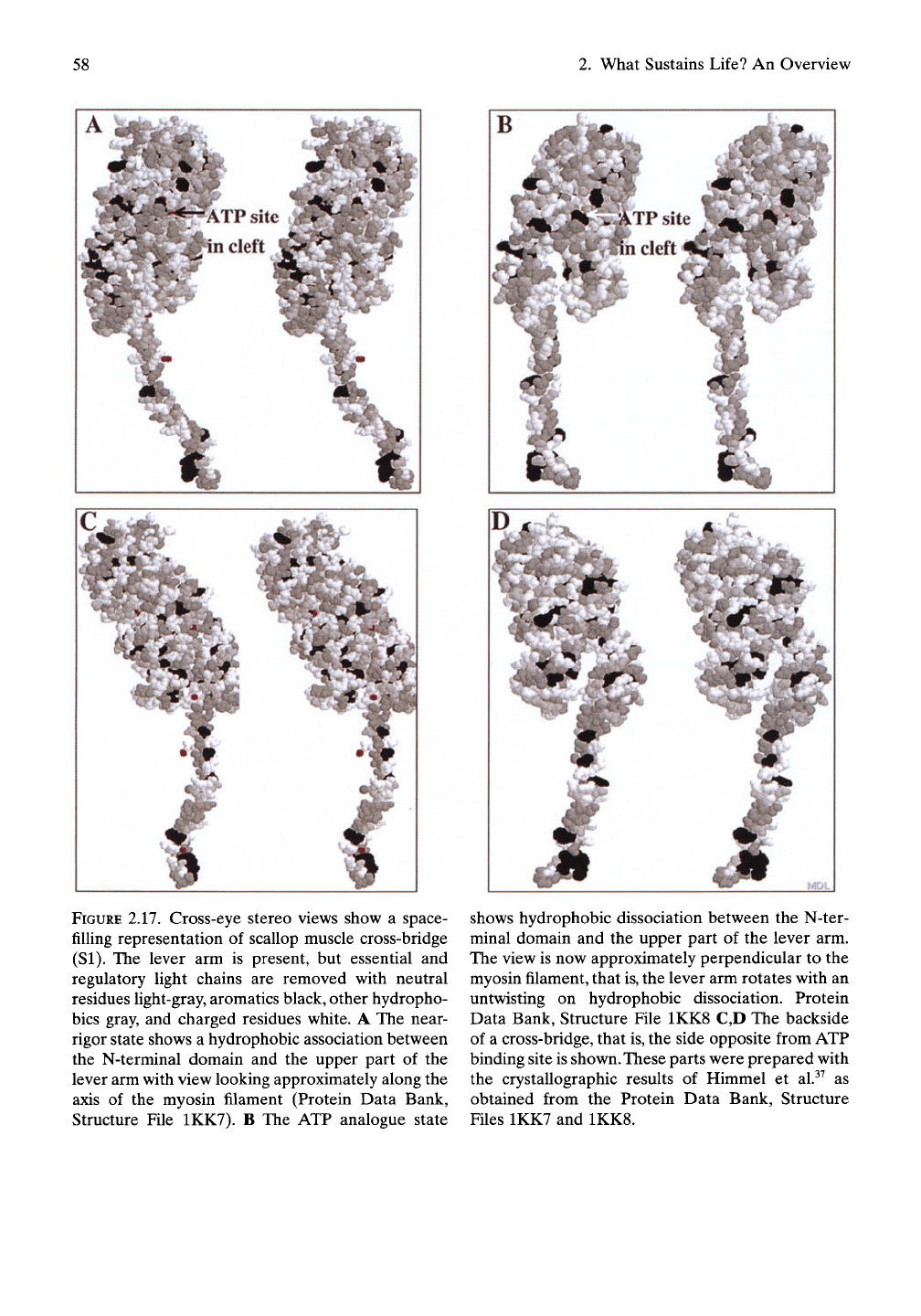
58
2.
What Sustains Life? An Overview
FIGURE 2.17. Cross-eye stereo views show a space-
fiUing representation of scallop muscle cross-bridge
(SI).
The lever arm is present, but essential and
regulatory light chains are removed with neutral
residues Hght-gray, aromatics black, other hydropho-
bics gray, and charged residues white. A The near-
rigor state shows a hydrophobic association between
the N-terminal domain and the upper part of the
lever arm with view looking approximately along the
axis of the myosin filament (Protein Data Bank,
Structure File 1KK7). B The ATP analogue state
shows hydrophobic dissociation between the N-ter-
minal domain and the upper part of the lever arm.
The view is now approximately perpendicular to the
myosin
filament,
that
is,
the lever arm rotates with an
untwisting on hydrophobic dissociation. Protein
Data Bank, Structure File 1KK8 C,D The backside
of a cross-bridge, that
is,
the side opposite from ATP
binding site is shown. These parts were prepared with
the crystallographic results of Himmel et al.^^ as
obtained from the Protein Data Bank, Structure
Files 1KK7 and 1KK8.

2.7 Consequences of Protein Machines Based on the Inverse Temperature Transitions
59
domain to cause the association to "open", that
is,
to dissociate. The cleft acts like a conduit
directing the thirst of the phosphate for hydra-
tion in both directions to disrupt hydrophobic
associations. The cleft is shown from several
dif-
ferent vantage points in Figures 8.58 and
8.59 and further discussed in their associated
text.
In Chapter
8,
more structural background and
molecular details of contraction exhibited by the
linear myosin II motor are considered after, in
Chapter 5, the physical basis for the apolar
(oil-like)-polar (vinegar-like) repulsive energy
that controls hydrophobic association is experi-
mentally and analytically developed. The crystal
structures of the cross-bridge of scallop muscle
provide remarkable examples of the consilient
mechanism functioning in this protein-based
machine!
2.7 Consequences of Protein
Machines Based on the Inverse
Temperature Transitions
We again draw comparison by quoting from
Schrodinger's What is Life! as published in
1944:
"What I wish to make clear in this last
chapter is, in short, that from all that we have
learnt about the structure of living matter, we
must be prepared to find it working in a manner
that cannot be reduced to the ordinary laws of
physics."^^ In the present volume, our founda-
tion
is,
of course, the somewhat counterintuitive
inverse temperature transition. It is by means
of the inverse temperature transition that the
energies essential to sustain Life can, in fact, be
accessed. This efficient consilient mechanism,
however, requires no new laws of physics. In
fact, the seeds of this mechanism are found in
a 1937 report of Butler,^^ in which he analyzed
the thermodynamic elements of the solubility
of oil-like groups in water.
The essential aspect of the capacity of the
inverse temperature transition to achieve
diverse energy conversions resides within large
chain molecules, which were just becoming
known when the first edition of Schrodinger's
book appeared. As we have sketched above, the
functional properties of the model protein-
based machines result from the interaction
between hydrophobic (oil-like) and hydrophilic
(vinegar-like) groups constrained to occur in
unique sequence along the chain molecule,
which combines with the capacity of vinegar-
like functional groups to shift between being
more vinegar-like and more oil-like.
The special effectiveness of proteins com-
pared with all other known chain molecules
arises from biology's capacity accurately to
produce diverse sequences. As for accuracy, the
error in placing the correct amino acid residue
at the specified sequence position depends on
the ability to delineate one R-group from
another at the stage of attachment to t-RNA
(see Chapter 4).^^ In general, the error is
remarkably small. It is also very important that
all of the residues be of the correct optical con-
figuration,"^^ as random mixing of mirror images
of amino acid residues would destroy structure
with the consequence of eliminating reliable
function (see the paragraphs on Pasteur and
mirror image molecules in section 3.1.6 of
Chapter 3).
These features of protein polymers result in
regular, nonrandom structures, which, as will be
shown later, are the reason for efficient energy
conversion. As for diversity, theoretically each
position in the sequence can be any one of 20
different amino acid residues. Again very sig-
nificantly, the protein sequence is specified
by the nucleic acid sequence and not by the
energetics within the sequence of a particular
protein. In this way, a sequence that would
normally be highly improbable becomes just as
probable as any other sequence. (Improbabili-
ties would otherwise arise by virtue of being a
single sequential arrangement of amino acids
among an inordinate number of possible
arrangements of the same collection of amino
acids and because of unfavorable interaction
energies between the R-groups and thus being
of high energy.) Of course, much energy has
been spent in producing the "improbable"
sequence, as discussed further in Chapter 4.
Production of unique and improbable
sequences might have been one place where
the far-from-equilibrium arguments of Pri-
gogine could have come into play. Order is
indeed achieved out of chaos, but it occurs at a

60
2.
What Sustains Life? An Overview
high price in energy—in the energy required to
copy the parent chain of DNA, in the energy
required to transcribe the DNA sequence into
RNA, and in the energy required to translate
the nucleic acid sequence into a protein
sequence.
To
the best of our present knowledge,
however, each of the many steps in the proce-
dure can be treated and studied in the test
tube as an equiUbrium process. Thus, once the
machinery of the cell is in place, the individual
steps within the cell appear to be reversible and
explicable in terms of the thermodynamics
of Boltzmann and Gibbs. However, as noted
below and treated in more detail in Chapter 4,
biology has employed a throw-away energy; the
pyrophosphate product of each step of the
assembly essential for reversibiUty is broken
down by the interesting enzyme pyrophos-
phatase to ensure unidirectionality to both
nucleic acid and protein synthesis.
Now, because it can cost just as much energy
to produce an inefficient and more limited
protein-based machine as it can to produce a
more efficient and/or a new machine that can
access a new energy source, obviously with
natural selection the arrow of time for biology
is toward greater complexity and diversity func-
tion (see Chapter 6).
2.7.1 Protein-based Machines as
Catalysts for Energy Conversion
When an energy input causes an inverse tem-
perature transition of hydrophobic folding, a
protein converts part of the energy into its own
folding and ordering; this constitutes one aspect
of a decrease in entropy. On completing a cycle
by returning to the unfolded and disordered
state,
the net entropy change, however, be-
comes zero for this cyclic folding and unfolding.
When in the process of cycling—mechanical,
chemical, electrical, or some other work—has
been catalyzed, then there results a useful
decrease in entropy resulting from the energy
input. A productive negative entropy change
does occur when, in the process of folding, the
protein picks up protons and deposits them at
a higher concentration, that is, pumps protons.
or the protein lifts a weight, that is, pumps iron,
for some useful purpose, or the protein phos-
phorylates ADP to form ATP. In each case
movement is away from disorder; a beneficial
increase in order, that is, a decrease in entropy,
has been achieved. Proteins that hydrophobi-
cally fold as the result of an energy input to
result in mechanical work, chemical work, or
some other energy output could reasonably be
classified as negative entropy machines, but this
is an incomplete characterization. An adequate
characterization would be to view protein-
based machines simply as catalysts of energy
conversion.
It is now possible to design and prepare
elastic-contractile model proteins capable of
interconverting the energies that sustain Life.
The set of six energies interconverted by living
organisms are represented symbolically in
Figure 2.18. Indicated by the bold intercon-
necting arrows are the pairwise energy conver-
sions,
directly demonstrated by designed model
proteins. Additional energy conversions have
been demonstrated less directly. These model
proteins are biomolecular machines of further
interest as advanced materials for medical and
nonmedical applications to enhance quality of
Life and to assist in sustaining an ever larger,
more complex and more productive society.
2.7.2 Efficiency and the Definition of
Chemical Energy
The efficiency, r|, of a process can be described
as the work achieved divided by the energy
input, that
is,
r\
=
work performed/energy input.
The work performed (w) can be the lifting of a
weight, which is the mass (m) of the object
times the force of gravity (g) times the height
(h) the object was raised, that is, w = mgh, as
indicated in Figure
2.5.
The energy input can be
due to the increase in concentration of protons,
which is proportional to the size of the step
along the horizontal axis in Figure 2.6B. In
Figure 2.6B, the size of step required to go com-
pletely from COO" to COOH for a less oil-like
protein with a broader sigmoid curve is greater
than for the more oil-like protein with its nar-
rower sigmoid
curve.
As the energy input comes
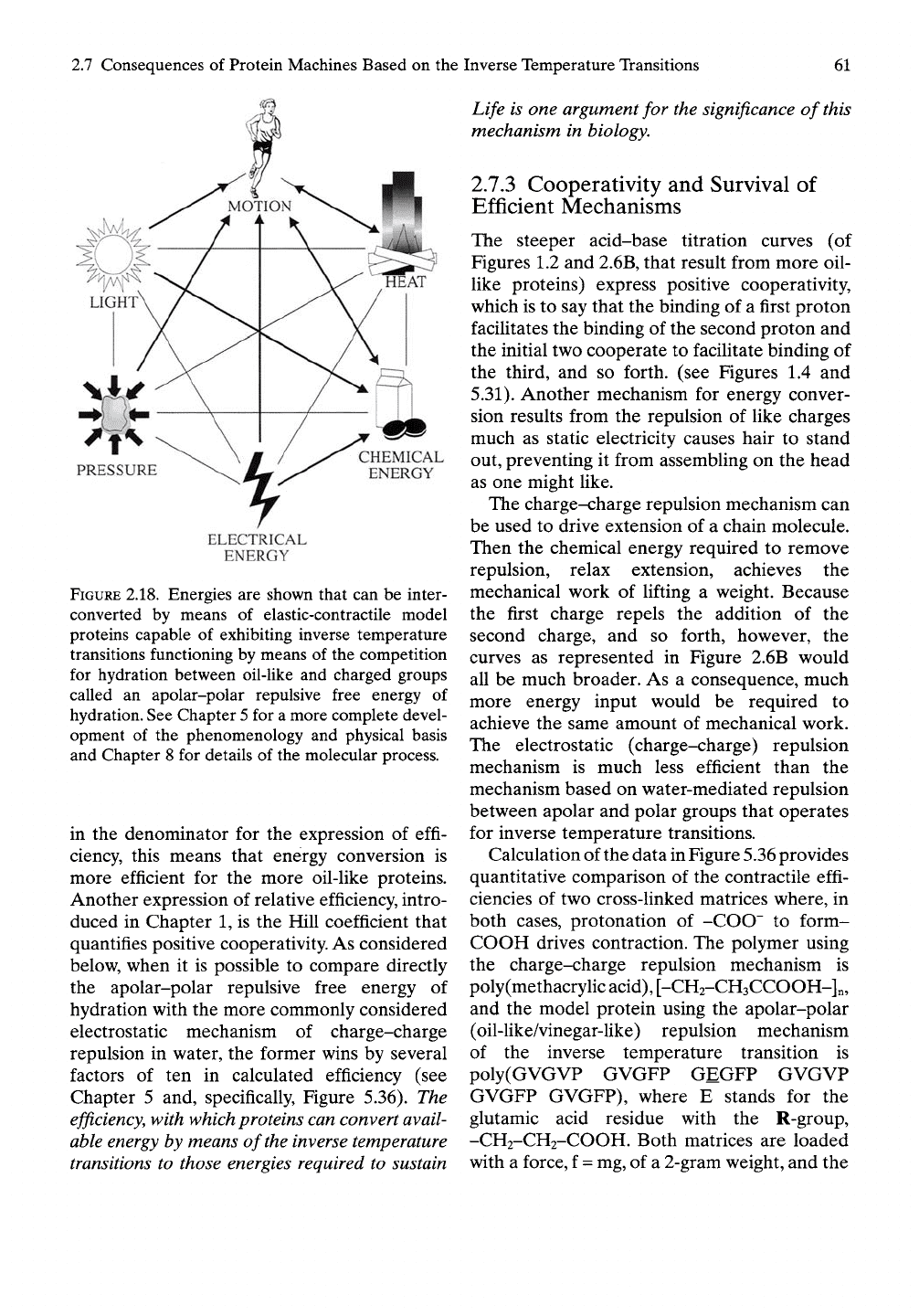
2.7 Consequences of Protein Machines Based on the Inverse Temperature Transitions
61
CHEMICAL
ENERGY
ELECTRICAL
ENERGY
FIGURE 2.18. Energies are shown that can be inter-
converted by means of elastic-contractile model
proteins capable of exhibiting inverse temperature
transitions functioning by means of the competition
for hydration between oil-like and charged groups
called an apolar-polar repulsive free energy of
hydration. See Chapter
5
for a more complete devel-
opment of the phenomenology and physical basis
and Chapter 8 for details of the molecular process.
in the denominator for the expression of effi-
ciency, this means that energy conversion is
more efficient for the more oil-like proteins.
Another expression of relative efficiency, intro-
duced in Chapter 1, is the Hill coefficient that
quantifies positive cooperativity. As considered
below, when it is possible to compare directly
the apolar-polar repulsive free energy of
hydration with the more commonly considered
electrostatic mechanism of charge-charge
repulsion in water, the former wins by several
factors of ten in calculated efficiency (see
Chapter 5 and, specifically. Figure 5.36). The
efficiency, with which proteins can convert avail-
able energy by means of the inverse temperature
transitions to those energies required to sustain
Life is one argument for the significance of this
mechanism in biology.
2.7.3 Cooperativity and Survival of
Efficient Mechanisms
The steeper acid-base titration curves (of
Figures 1.2 and 2.6B, that result from more oil-
like proteins) express positive cooperativity,
which is to say that the binding of a first proton
facilitates the binding of the second proton and
the initial two cooperate to facilitate binding of
the third, and so forth, (see Figures 1.4 and
5.31). Another mechanism for energy conver-
sion results from the repulsion of like charges
much as static electricity causes hair to stand
out, preventing it from assembling on the head
as one might like.
The charge-charge repulsion mechanism can
be used to drive extension of a chain molecule.
Then the chemical energy required to remove
repulsion, relax extension, achieves the
mechanical work of lifting a weight. Because
the first charge repels the addition of the
second charge, and so forth, however, the
curves as represented in Figure 2.6B would
all be much broader. As a consequence, much
more energy input would be required to
achieve the same amount of mechanical work.
The electrostatic (charge-charge) repulsion
mechanism is much less efficient than the
mechanism based on water-mediated repulsion
between apolar and polar groups that operates
for inverse temperature transitions.
Calculation of the data in Figure 5.36 provides
quantitative comparison of the contractile effi-
ciencies of two cross-linked matrices where, in
both cases, protonation of -COO" to form-
COOH drives contraction. The polymer using
the charge-charge repulsion mechanism is
poly(methacrylic acid), [-CHs-CHsCCOOH-]^,
and the model protein using the apolar-polar
(oil-like/vinegar-like) repulsion mechanism
of the inverse temperature transition is
poly(GVGVP GVGFP GEGFP GVGVP
GVGFP GVGFP), where E stands for the
glutamic acid residue with the R-group,
-CH2-CH2-COOH. Both matrices are loaded
with a force, f = mg, of a 2-gram weight, and the

62
2.
What Sustains Life? An Overview
contraction, that
is,
the decrease in length (AL)
as the pH is lowered, occurs on addition of
acid. Now efficiency can be written as r| = fAL/
(2.3RTApH)An, where R is the gas constant
(1.987
cal/mol-deg),
T is the temperature in
degrees K (centigrade degrees +
273),
and An is
the number of protons
used.
Based on the steep-
est slope for each curve in Figure 5.36, the ratio
of the efficiencies of the inverse temperature
transition mechanism,
r|ap,
to the charge-charge
repulsion mechanism, r|cc, is given by r|ap/r|cc =
[ApHAn]pMA/[ApHAn]E4F. On substitution of the
experimental numbers, r|ap/r|cc = [(2.24)(6.86 x
10-0]/[(0.59)(6.12
X
10-^)] «
40.
The mechanism
based on the inverse temperature transition of
the designed elastic-contractile model proteins
is more than 40 times more efficient. The elec-
trostatic mechanism in water is only a few
percent as effective as the mechanism that uses
the inverse temperature transition.
Due to the struggle to survive under circum-
stances of limited food
supply,
organisms evolve
to use the most efficient mechanism available to
their composition. The most efficient mecha-
nism available to the proteins that sustain Life
would seem to be the apolar-polar repulsive
free energy of hydration as observed for the
inverse temperature transitions for hydropho-
bic association. The efficiency of designed
elastic-contractile protein-based machines
and a number of additional properties make
designed protein-based materials of substantial
promise for the marketplace of the future.
2.8 "What Sustains Life?"
Becomes "What Sustains
Society?" (Biomolecular
Machines as Advanced Materials
for the Future)
Advances in physics, chemistry, and biology
spawned technologies that now support larger
populations at higher standards of living; they
have provided fossil fuel-powered, hydro-
powered, nuclear-powered, and chemical-
based energy sources (e.g., electricity, fuels,
explosives); they have given us electronics
and communications, agricultural enhancement
methods and products, medicines, new materi-
als such as plastics, and related medical de-
vices.
Now, fertile technologies, spawned from
advances in molecular biology and molecular
biophysics, promise opportunities to continue
to sustain society at higher population levels
with higher standards of living.
Here we utilize insights gained from the
capacity of living organisms to access and trans-
form energy and apply these protein-based
machines to problems of consequence to an
ever larger, more complex
society.
Three major
concerns of our society are an unsustainable
rise in medical care costs, the drug addiction
problem that drives an increasing crime rate
and supports terrorism, and pollution of the
environment.
The preceding design of model proteins that
emulate energy conversions of living organisms
constitutes useful knowledge for the design of
model proteins for medical and nonmedical
use.
In this brief overview, we mention three
appUcations that relate to the above-noted
problems in our
society.
Two
are medical appli-
cations, tissue reconstruction and drug delivery,
and the third is primarily a nonmedical appU-
cation of (bio)degradable plastics. These are
treated more extensively in Chapter 9.
The chosen three examples result from our
newly developed understanding of protein-
catalyzed energy conversion. This brings us to
the modern-day situation so well-stated by J.
Bronowski,"^^ "In effect, the modern problem is
no longer to design a structure from the mate-
rials,
but to design the materials for a structure."
For medical applications the biocompatibil-
ity of the model proteins must, of course, be
established; this has been done for three basic
compositions, but verification of nontoxicity
must be established for each specific composi-
tion designed for a particular human or veteri-
narian use.
The ultimate limitation to the utilization of
model proteins as advanced materials for the
future resides in low-cost, reliable production.
Proteins are unique polymers in that they can
be prepared by genetic engineering, allowing
for fidelity and diversity of sequence not pos-
sible for any other known chain molecules.
An overview would be incomplete without a
2.8 "What Sustains Life?" Becomes "What Sustains Society?"
63
few comments on this capacity as presently
demonstrated.
2.8.1 Genetic Engineering of
Model Proteins
On chemically synthesizing the first very long
chains of poly(GVGVP) early in 1984 with the
support from the Army Medical Research
Office, our peptide chemist, K. U. Prasad, sug-
gested that it would make a very nice chewing
gum, an idea dismissed with a laugh for one
stick of gum would have cost a few hundred
dollars. As the result of an Accelerated
Research Initiative for producing elastomers of
repeating peptide sequences by genetic engi-
neering, which was announced by the Office of
Naval Research in
1985,
the current anticipated
cost for an edible stick of gum could became as
little as a few pennies. This cost comparison
between chemical synthesis in the research lab-
oratory and microbial biosynthesis illustrates
the expansion of applications possible with
the recent success of recombinant DNA tech-
nology for producing (GVGVP)25/^ and other
protein-based polymers.
As discussed in subsequent chapters, the
error-free preparation resulting from microbial
biosynthesis is just as important for function as
lowering production cost is for expansion of
applications. The basic gene design and con-
struction occur with essentially no limitations
on sequence, and expression of the protein-
based polymer proceeds with few constraints
on sequence. The basic (monomer) gene is
polymerized enzymatically to provide genes for
the entire possible range of exact polymer
chain lengths. To date, model proteins con-
taining as many as 4,000 residues have been
produced with remarkably high quantities
prepared per volume of the fermentor. An
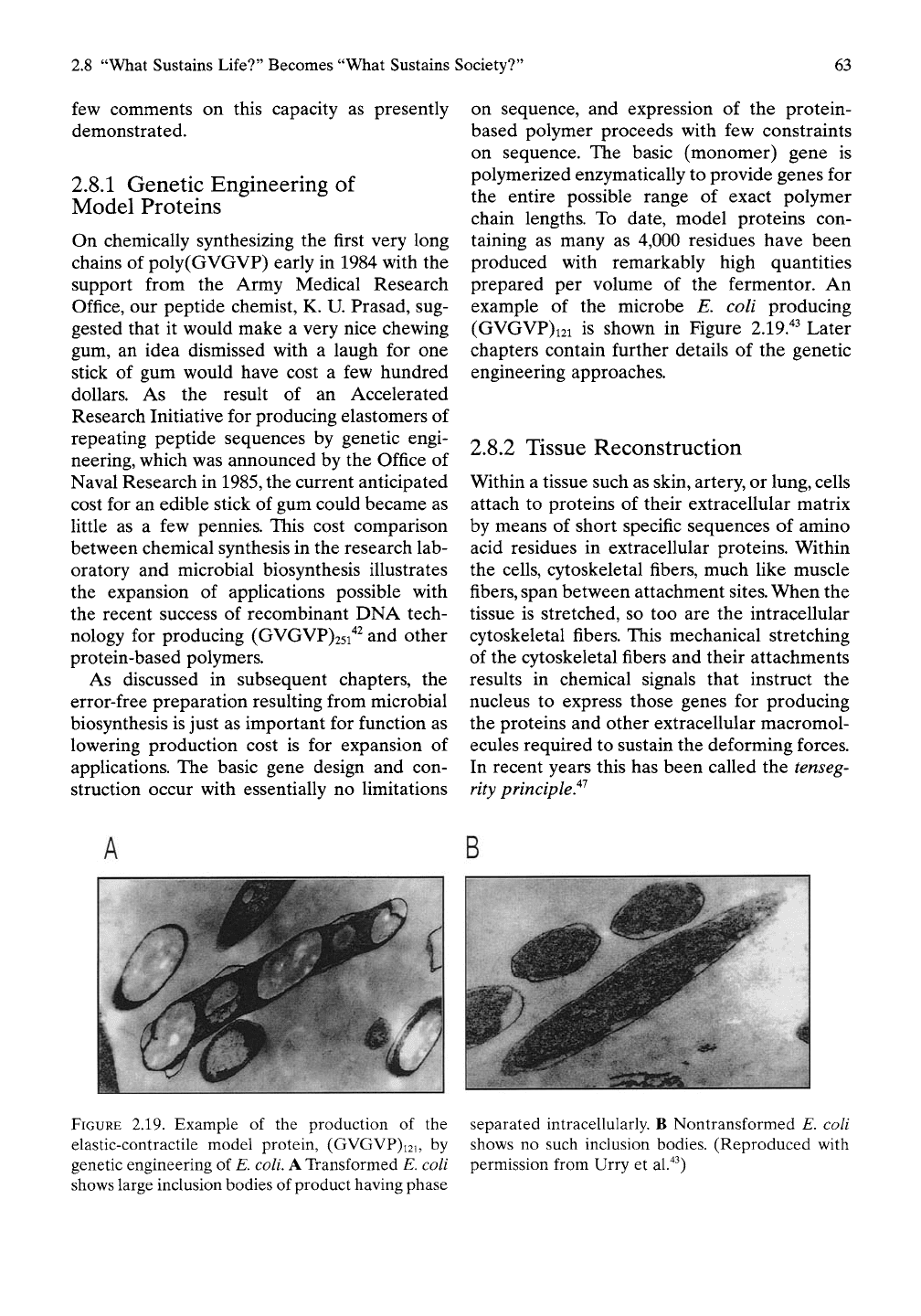
example of the microbe E. coli producing
(GVGVP)i2i is shown in Figure 2.19.^^ Later
chapters contain further details of the genetic
engineering approaches.
2.8.2 Tissue Reconstruction
Within a tissue such as skin, artery, or lung, cells
attach to proteins of their extracellular matrix
by means of short specific sequences of amino
acid residues in extracellular proteins. Within
the cells, cytoskeletal fibers, much like muscle
fibers, span between attachment
sites.
When the
tissue is stretched, so too are the intracellular
cytoskeletal fibers. This mechanical stretching
of the cytoskeletal fibers and their attachments
results in chemical signals that instruct the
nucleus to express those genes for producing
the proteins and other extracellular macromol-
ecules required to sustain the deforming forces.
In recent years this has been called the tenseg-
rity
principle f'^
B
FIGURE
2.19. Example of the production of the
elastic-contractile model protein, (GVGVP)i2i, by
genetic engineering of E. coli. A Transformed E. coli
shows large inclusion bodies of product having phase
separated intracellularly. B Nontransformed E. coli
shows no such inclusion bodies. (Reproduced with
permission from Urry et al."^^)
64
2.
What Sustains Life? An Overview
An equivalent conversion of mechanical
energy into chemical energy occurs with the
elastic model proteins. Stretching of properly
designed and cross-linked elastic model pro-
teins,
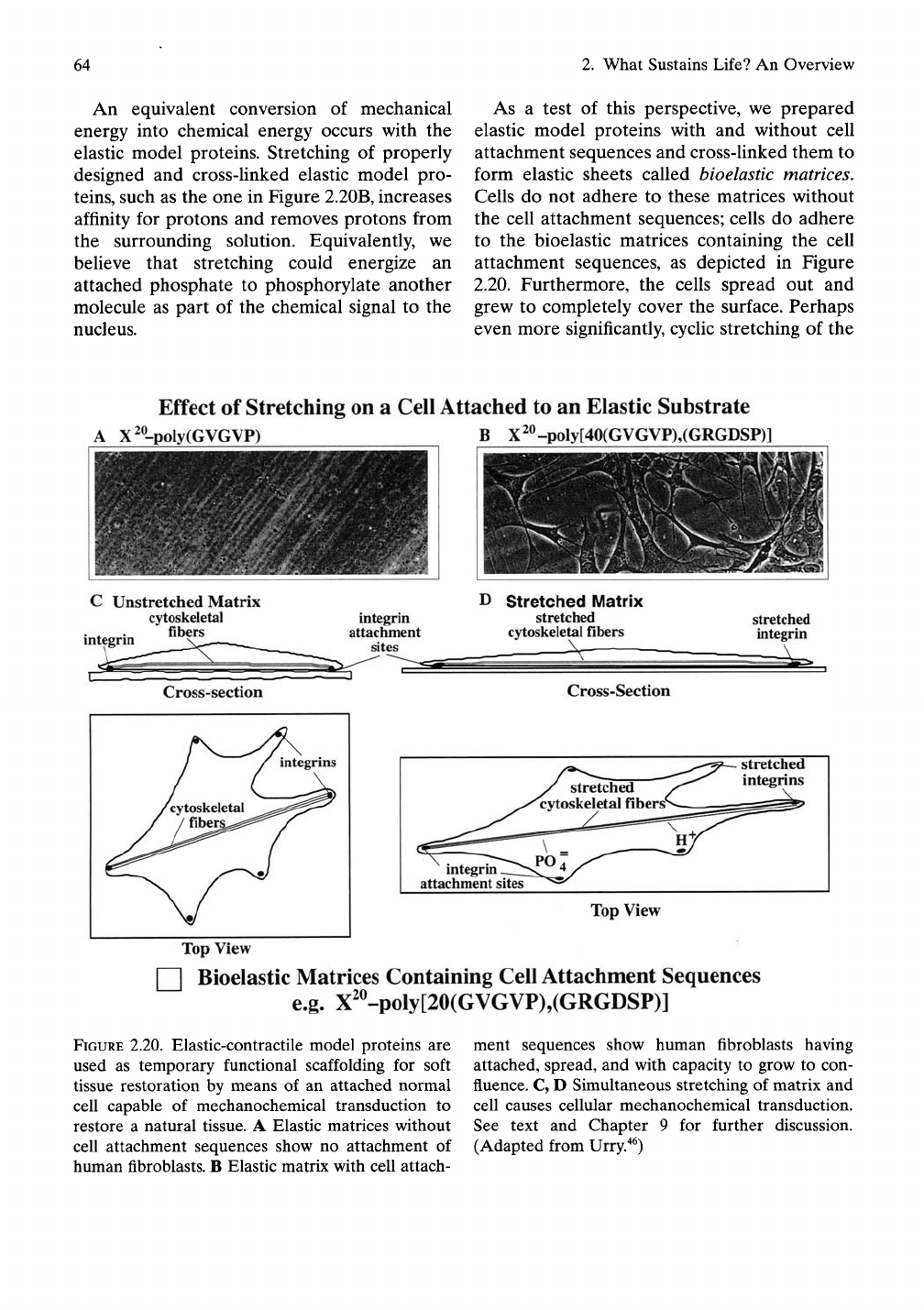
such as the one in Figure 2.20B, increases
affinity for protons and removes protons from
the surrounding solution. Equivalently, we
believe that stretching could energize an
attached phosphate to phosphorylate another
molecule as part of the chemical signal to the
nucleus.
As a test of this perspective, we prepared
elastic model proteins with and without cell
attachment sequences and cross-linked them to
form elastic sheets called bioelastic matrices.
Cells do not adhere to these matrices without
the cell attachment sequences; cells do adhere
to the bioelastic matrices containing the cell
attachment sequences, as depicted in Figure
2.20. Furthermore, the cells spread out and
grew to completely cover the surface. Perhaps
even more significantly, cyclic stretching of the
Effect of Stretching on a Cell Attached to an Elastic Substrate
B X^"-poly[40(GVGVP),(GRGDSP)]
C Unstretched Matrix
cytoskeletal
. . fibers
integrin
integrin
attachment
sites
D Stretched Matrix
stretched
cytoskeletal fibers
stretched
integrin
Cross-section Cross-Section
integrin
_^^N^
4
attachment sites
stretched
integrins
Top View
Top View
I I Bioelastic Matrices Containing Cell Attachment Sequences
e.g. X^^-poly[20(GVGVP),(GRGDSP)]
FIGURE 2.20. Elastic-contractile model proteins are
used as temporary functional scaffolding for soft
tissue restoration by means of an attached normal
cell capable of mechanochemical transduction to
restore a natural tissue. A Elastic matrices without
cell attachment sequences show no attachment of
human fibroblasts. B Elastic matrix with cell attach-
ment sequences show human fibroblasts having
attached, spread, and with capacity to grow to con-
fluence. C, D Simultaneous stretching of matrix and
cell causes cellular mechanochemical transduction.
See text and Chapter 9 for further discussion.
(Adapted from Urry."^^)